1. Introduction
Dry eye disease is currently classified into two major categories: evaporative dry eye (EDE) and aqueous tear-deficient dry eye (ADDE) [
1]. EDE is characterized by excessive water loss from the ocular surface despite normal lacrimal gland function, while the primary cause of ADDE is insufficient lacrimal secretion. The hyperosmolarity of the tear film observed in ADDE, a result of the reduction in lacrimal gland fluid secretion, inflames the underlying ocular surface. ADDE is further divided into two major subclasses based on etiological causes: Sjögren’s Syndrome Dry Eye (SSDE) and Non-Sjögren’s Syndrome Dry Eye (NSSDE).
Sjögren’s Syndrome (SS) is a prevalent autoimmune disorder [
2], affecting approximately 3–4% of adults [
1] and disproportionately affecting women at a ratio of approximately 9:1 [
2]. This disease involves the chronic inflammation of selected exocrine glands, producing xerostomia (dry mouth) and xerophthalmia (dry eye). The manifestations of dry eye are linked to the inflammation of the cornea, conjunctiva, and lacrimal gland [
1].
The cornea is one of the most densely innervated structures in the entire human body. Histological studies of the corneal epithelium reveal nerve bundles in the underlying stroma that penetrate the Bowman’s membrane and branch into smaller bundles subjacent to the epithelium. These nerves running parallel to the corneal surface are known as the subbasal plexus. From the subbasal nerves, smaller individual fibers penetrate into different epithelial cell layers, terminating close to the surface as free nerve endings [
3]. The majority of these nerves are sensory and the nerve endings release neurotransmitters, mainly Substance P (SubP) and calcitonin gene-related peptide (CGRP), in response to noxious stimuli mediating the inflammatory response.
Irritants interacting with the ocular surface and other stimuli, such as temperature, trigger the complex neural reflex arc that stimulates the production of tears. This arc consists of afferent sensory corneal nerves, the trigeminal ganglion, higher level ganglia and brain areas, and efferent parasympathetic and sympathetic nerves that innervate the conjunctiva, lacrimal gland, and meibomian gland. Activation of the efferent nerves causes fluid production from these tissues that combine to form tears. Thus, loss or changes in the sensitivity of the afferent corneal nerves would decrease their activity, lower tear volume, and alter the composition of the tears. This change in tear production can cause dry eye [
2,
4]. Using confocal microscopy, Villani et al. reported that SS patients show a decrease in both the number and density of corneal nerve fibers in the subbasal plexus [
5].
Thrombospondin-1 (TSP-1) is a matricellular protein that activates latent transforming growth factor (TGF)-β, a potent anti-inflammatory cytokine. TSP-1 interacts with cell surface receptors, growth factors, and extracellular matrix proteins to regulate extracellular and intracellular signaling complexes [
6]. TSP-1 is expressed in the corneal epithelium, stroma, and endothelium. To demonstrate the role of TSP-1 in ocular surface function, we used the TSP-1-deficient (TSP-1
−/−) mouse model. In both sexes this mouse appears to be normal at birth, but with age progressively develops dry eye and the aberrant histopathology associated with Sjögren’s Syndrome, making it a novel mouse model of dry eye [
7]. In the lacrimal glands of TSP-1
−/− mice the peripheral nerves are injured, preventing the release of neurotransmitters and decreasing protein secretion, thus leading to dry eye [
8]. In the malfunctioning lacrimal glands, an increase in apoptosis and deterioration is observed with the production of CD4+ T-rich inflammatory infiltrates, similar to those seen with Sjögren’s Syndrome. On the ocular surface, the lack of TSP-1 leads to the disruption of the corneal epithelial layer, corneal edema, and a reduction in conjunctival goblet cell density [
7].
Around the age of 12 weeks, the presence of anti-Sjögren’s Syndrome antigen A and B antibodies were confirmed in the serum of TSP-1
−/− mice. A loss of function of the lacrimal glands and therefore the tear production was suggested to explain the changes observed in the cornea [
7,
9]. A decrease in corneal nerve function could contribute to the decreased tear film and the development of dry eye; however, the structure and function of corneal nerves, specifically the sensory nerves and nerve endings, in the TSP-1
−/− mouse has yet to be investigated.
The present study aims to determine if the structure of nerves, specifically neurotransmitter-containing nerves, and infiltrating cells was altered in the corneas of TSP-1−/− compared to the control wild type mice. We performed histological, immunohistochemical, and in vivo confocal analyses to assess any changes in nerve structure and we observed a decrease in corneal nerve density in TSP-1−/− mice compared to their WT counterparts in the control group. Cytokine analysis also revealed an increase in the expression of dry eye-associated inflammatory mediators in the cornea.
3. Discussion
Dry eye from lacrimal gland and ocular surface dysfunction is widely associated with inflammation. Especially in autoimmune dry eye, an upregulation of proinflammatory cytokines, chemokines, and metalloproteinases has been demonstrated [
11]. The increase in these inflammatory factors leads to a rapid expansion of autoreactive T cells which then migrate to the conjunctiva, lacrimal gland, and cornea, causing inflammation and dysfunction [
12]. In TSP-1
−/− mice there is an age-dependent influx of pro-inflammatory lymphocytes that can alter lacrimal gland and conjunctival structure and function, leading to decreased secretion and ocular surface disease [
7,
8,
13]. In the present study, we demonstrated that similar changes in nerve structure and pro-inflammatory cytokine influx occur in the cornea, as in the lacrimal gland.
Tear secretion onto the ocular surface is regulated by a complex neural reflex that consists of the afferent sensory nerves of the cornea, trigeminal ganglion, trigeminal nucleus, superior salivatory nucleus, and the efferent parasympathetic and sympathetic nerves that innervate the lacrimal gland, conjunctival goblet, squamous epithelial cells, and corneal epithelial cells. Previously, we studied the efferent pathway in the absence of TSP-1 in mice, providing a model of autoimmune dry eye in which ocular surface homeostasis is disrupted [
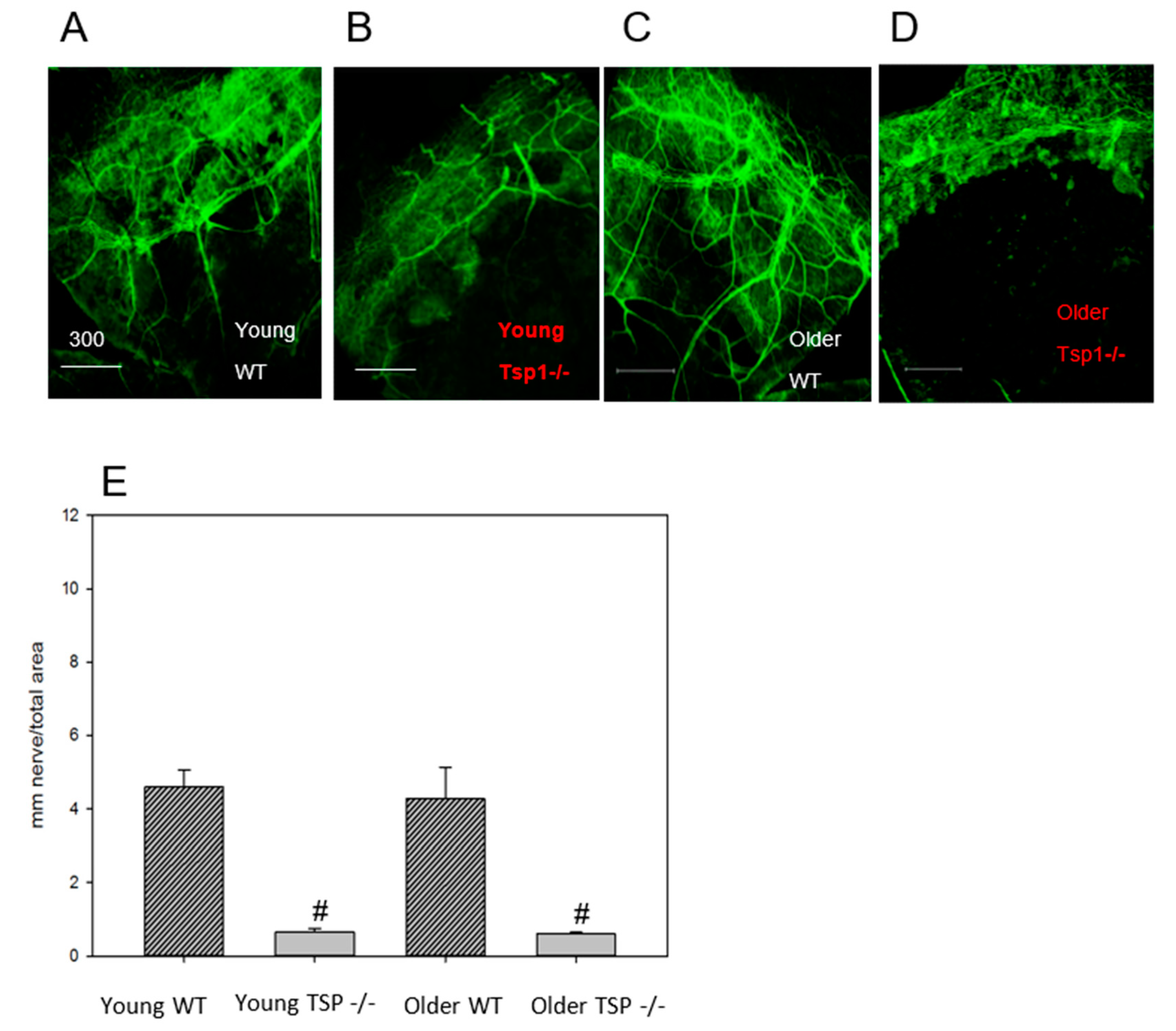
7]. Using this model, we found that TSP-1 is needed for the normal structure of the efferent parasympathetic and probably sympathetic nerves in the lacrimal gland as well as for function, as measured by depolarization-induced protein secretion. This dysfunction of lacrimal gland secretion contributes to the development of dry eye. In the present study, we focused on the afferent component of the neural reflex. Similar to the efferent nerves, we found that afferent nerve structure in the cornea is dependent upon TSP-1. We used two different methods, one in vivo confocal microscopy and one in vitro immunohistochemistry technique, that detect all the different types of nerves in the cornea. Both of these methods demonstrated fewer and discontinued nerve fibers in older TSP-1
−/− mice compared to WT control mice. Although we did not test function, because the structural changes in efferent lacrimal gland and afferent corneal nerves were similar, we propose that a reduction in the number and function of corneal nerves contributes to a suboptimal neural tear reflex in TSP-1
−/− mice. Thus, corneal afferent and lacrimal gland efferent nerves seem to be similarly affected in TSP-1
−/− mice.
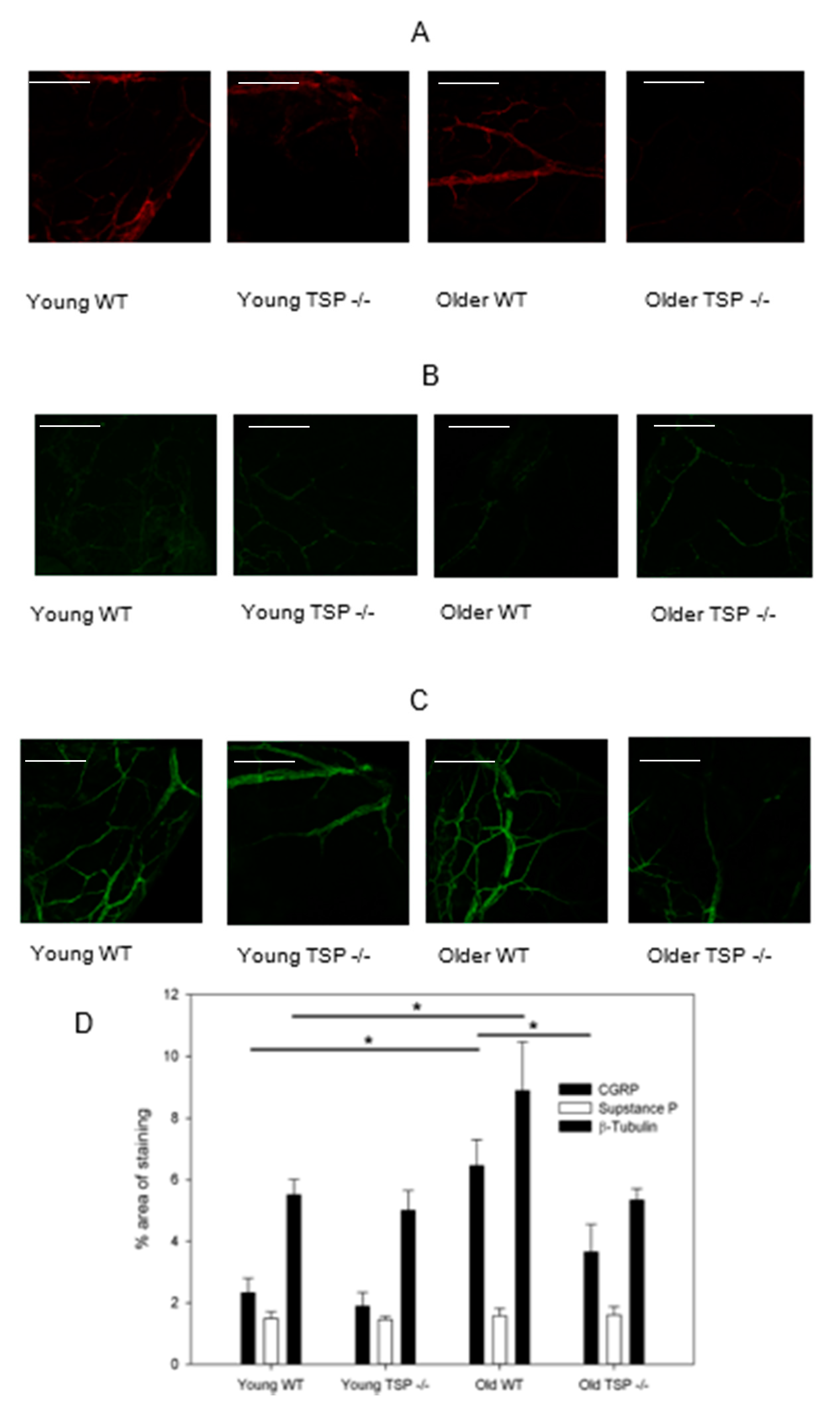
Approximately half of primary afferent neurons in the cornea are reported to be peptidergic nociceptors containing SubP and/or CGRP [
14,
15,
16], where the concentration and distribution of CGRP are reported higher than those of SubP [
17,
18,
19]. Our data on these two neuropeptides showed a substantially decreased amount of CGRP in the corneas of older TSP-1
−/− as compared to WT mice of the same age. In contrast, no changes in SubP were found. Both SubP and CGRP are known to be involved in corneal wound healing, and delayed wound healing has been observed in TSP-1
−/− mice [
20,
21,
22]. Importantly for the present study, CGRP is believed to play a key role in normal immunological functions by maintaining an anti-inflammatory state [
23]. CGRP can function as an anti-inflammatory mediator that dampens excessive immune responses in septic shock, autoimmune diabetes, inflammatory bowel disease, and UV radiation-induced immunosuppression [
24]. This neurotransmitter appears to work by preventing the development of T-cell driven autoimmune responses, as occurs in Sjögren’s Syndrome. Like in superior cervical ganglia neurons, CGRP works to increase TGF-β to produce regulatory T cells that dampen inflammation [
25]. TSP-1 activates TGF-β to prevent inflammation in the ocular surface tissues. As CGRP has the same function as TGF-β, the decrease in corneal CGRP-containing nerves with disease progression in TSP-1
−/− mice is consistent with an increased immune response and could contribute to this increased response. It is not known, however, how the loss of TSP-1 decreases CGRP-containing nerves.
In contrast to CGRP, which is anti-inflammatory, SubP is pro-inflammatory. The blockage of SubP or its receptor in knockout mice or by pharmacological blockade has an anti-inflammatory effect in many chronic inflammatory diseases including arthritis, type 1 diabetes, inflammatory bowel disease, HSV-1-corneal infection, and corneal neovascularization [
26]. SubP release stimulates Th1 and Th17 autoreactive T cells to produce IL17 and increases chemokines that recruit immune cells to the site of inflammation. In the present study, the level of SubP-containing neurons is unchanged with disease progression in TSP-1
−/− mice, allowing this neurotransmitter to continue its role in increasing inflammation. Our results suggest that CGRP- and SubP-containing nerves are differentially regulated, consistent with their distinct roles in the regulation of immune response.
It is possible, as in our study, to measure CGRP- and SubP-containing nerves separately in animal models. When studying corneal wound healing in mouse cornea, Cortina et al. found that after wounding, CGRP-containing nerves returned with PEDF + DHA treatment, but SubP-containing nerves did not [
27]. This is further evidence that CGRP- and SubP-containing nerves are differentially regulated, consistent with their different functions.
Our results from the present study, combined with the different immunological roles of SubP and CGRP, suggest that it is critical to determine which type of neurotransmitter is altered when a decrease in corneal nerves is detected in various corneal pathologies or surgical treatments, including dry eye, corneal transplantation, and LASIK surgery. Although it is not possible to differentially evaluate SubP- and CGRP-containing nerves in humans, tear CGRP and SubP levels can be measured in tears. When CGRP and SubP were measured in tears after LASIK, CGRP levels increased whereas SubP levels were unchanged [
28]. These findings could explain why the ocular surface remains relatively quiet after this type of surgery. This result is consistent with our current study in that CGRP- and SubP-containing nerves are differentially regulated.
In the present study we found that the structure of corneal afferent nerves was disrupted in the absence of TSP-1, suggesting that TSP-1 is needed as an extracellular matrix protein either for appropriate peripheral nerve development or for nerve repair. There is little published work on the effect of TSP-1 on peripheral nerves, except our previous study on lacrimal gland efferent parasympathetic nerves in which TSP-1 was also needed for nerve growth or repair [
8]. Several publications have found, however, that TSP-1 is needed for synapse formation in the central nervous system, with TSP-1 being secreted by astrocytes [
29,
30,
31]. The secreted TSP-1 interacts with the neuronal calcium channel subunit
α26-1 that may act as the on switch for synapse formation. The synapses for afferent corneal nerves project to the trigeminal ganglion. As trigeminal ganglion neurons do not have synapses, TSP-1 must work by a different mechanism to affect corneal efferent nerves. TSP-1 does have multiple binding partners in addition to
α26-1, including ApoER2, CD36, CD47, heparin, latent TGF-β, LRP1/CRT, neuroligin, notch, and VLDR [
29], that could function in the regulation of corneal afferent nerve growth. More research is needed to determine the mechanism by which TSP-1 affects corneal sensory nerves, including the cellular source of TSP-1, the neural binding partner, and the cellular location (trigeminal ganglion, nerve axon, nerve endings, or supporting cells) of the interaction.
In addition to TSP-1 directly affecting corneal afferent nerve growth or repair by a TSP-1 protein interaction, TSP-1 could alter these nerves indirectly by its effect on inflammation. In the conjunctiva, in the absence of TSP-1, there is an increase in inflammatory cell infiltration and in pro-inflammatory cytokine production from the lack of activation of TGF-βII (the main TGF-β isoform in the ocular surface) [
13,
32]. In the cornea, the expression of three different pro-inflammatory cytokines—TNF-α, MCP-1, and MIP-2—was examined and only MIP-2 was elevated in young TSP-1 compared to WT cornea; however, all three cytokines were increased in older mice. Furthermore, CD11b+ cells were not altered in older TSP-1
−/− mouse corneas. These results argue against an indirect effect of TSP-1 on nerves via the activation of MCP and TNF-α-dependent pro-inflammatory pathways in young mice. TSP-1 could play an effect in addition to the direct effect of TSP-1 binding for MIP-2-dependent action in young mice and for all three cytokines measured in older mice. More research, however, on different time points, cytokines, and immune cells is needed to determine if the activation of inflammation plays a role in the damage to corneal nerve growth and repair.
In conclusion, we found that TSP-1 is needed for the development or repair of corneal afferent nerves, especially in the population of nerves containing CGRP as a neurotransmitter. The changes in these nerves occur in older TSP-1−/− mice as the corneal expression of pro-inflammatory cytokines increases, but there is no indication of inflammatory cell activation. TSP-1 could control corneal CGRP-containing nerve development and repair by a direct effect as a matricellular protein interacting with its binding partners on the nerves and/or indirectly by preventing the development of an inflammatory environment.
4. Materials and Methods
4.1. Animals
Female C57BL/6 and TSP-1−/− mice (C57BL/6 background) aged 4–7 weeks (young, pre disease onset) and 9–12 weeks (older, post disease onset) were used in this study. C57BL/6 mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA). TSP-1−/− mice used were originally developed by Dr. Jack Lawler (Harvard Medical School, Boston, MA, USA) and were bred in-house in a pathogen-free facility at Schepens Eye Research Institute, Boston, MA or were obtained from Dr. Sharmila Masli (Boston University, Boston, MA, USA). All experiments conformed to the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 8023, revised 1978) and were approved by the Schepens Eye Research Institute Animal Care and Use Committee, project idenfitication code S472-1219, approval date 28 March 2017.
4.2. Hemotoxylin and Eosin Staining
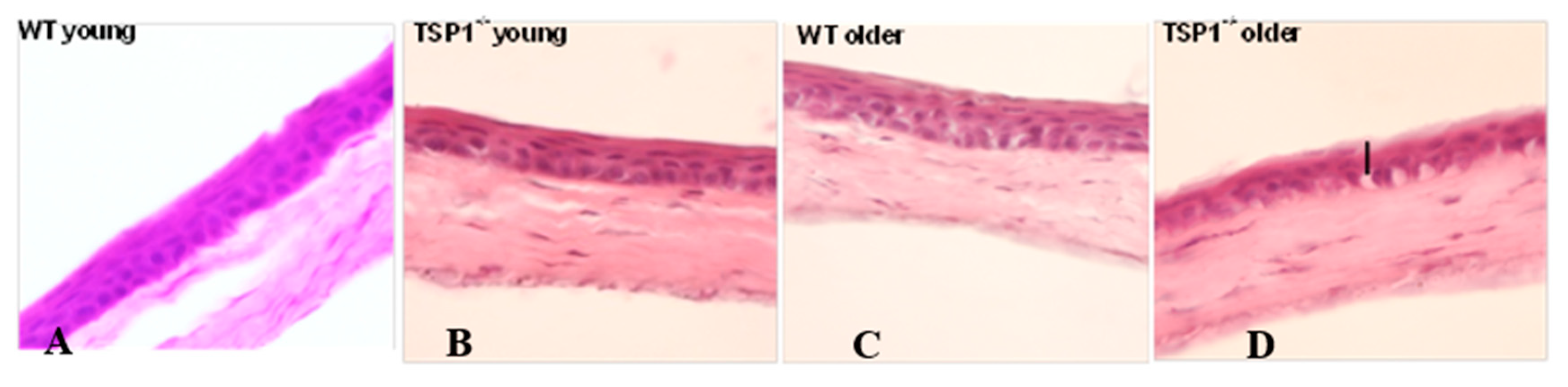
Whole eyes were excised and fixed in 4% paraformaldehyde at room temperature for 40 min. Fixed eyes were washed in phosphate buffered saline (PBS) and cryoprotected with sucrose. The tissue was embedded in optimal cutting temperature (OCT) media (Tissue-Tek, Torrance, CA, USA), and snap-frozen as previously described. Six to eight micron corneal sections were cut and stained with hematoxylin and eosin reagent. The slides were viewed by light microscopy (Eclipse E80i; Nikon, Tokyo, Japan).
4.3. Quantitative Real-Time PCR
Three corneas were harvested from each group: 4-week-old (young, before disease onset), 12-week-old (older, after disease onset) TSP-1−/− mice, (C57BL/6 background) and C57BL/6 mice. Total RNA was isolated from the corneas using a commercially available kit (KAPA SYBR® FAST qPCR Kit (KK4602)) and reverse transcribed using a Super Script® III Kit (Invitrogen Life Technologies, Carlsbad, CA, USA). Real-time PCR was performed to determine the relative quantitative expression of dry eye-associated inflammatory molecules (TNF-α, MCP-1, and MIP-2) by using Universal PCR Mastermix (Invitrogen, Carlsbad, CA, USA) and predesigned primers on an Ependorf Mastercycler (Ependorf North America, New York, NY, USA). Data were analyzed by a comparative threshold cycle method and normalized by GAPDH as an internal control.
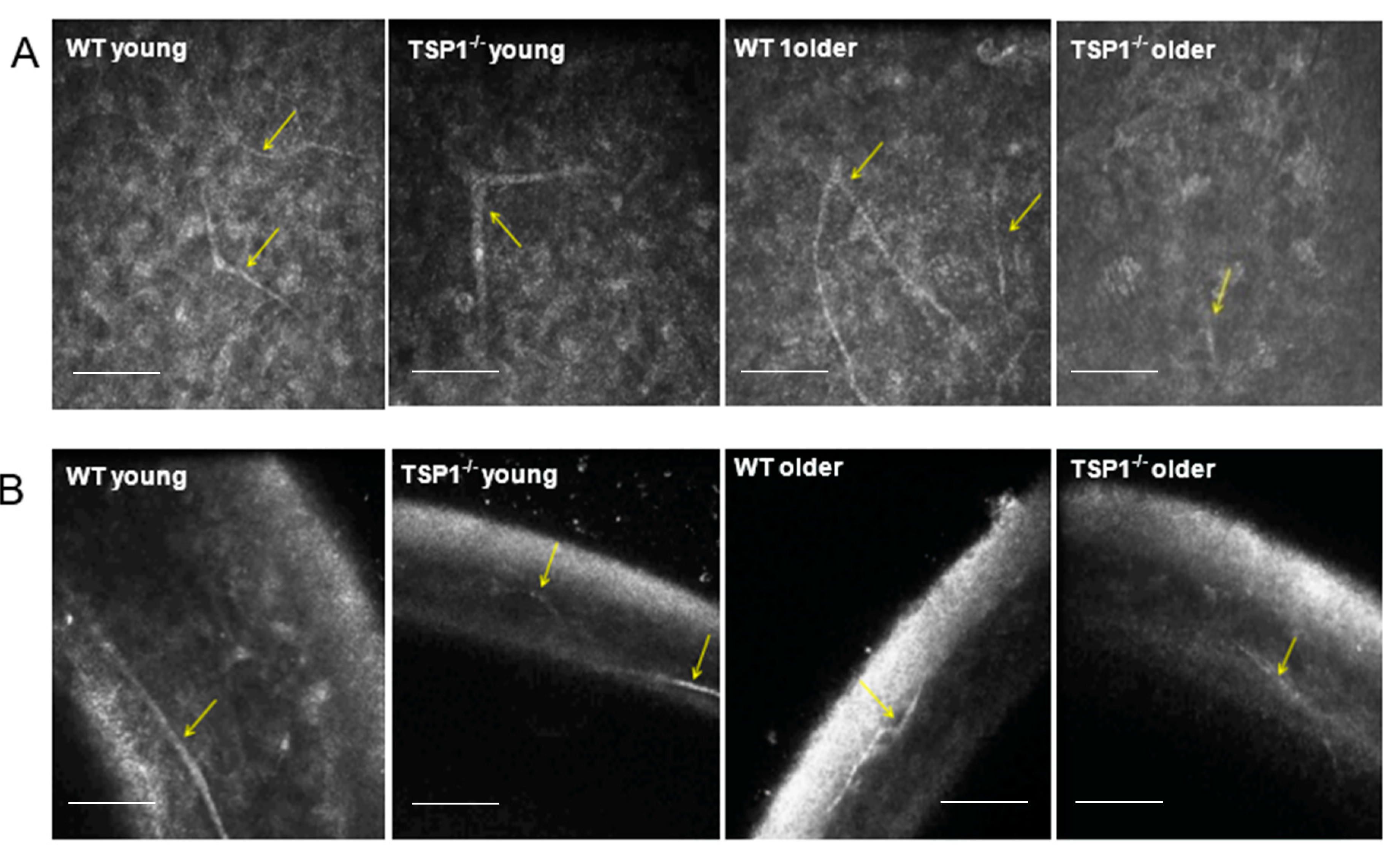
4.4. In Vivo Confocal Microscopy
For in vivo confocal microscopy, mice were anesthetized via an intraperitoneal injection of ketamine (120 mg/kg; Phoenix Scientific, St. Joseph, MO, USA) and xylazine (20 mg/kg; Phoenix Scientific). Eyes were treated with a drop of Proparacaine 0.5% Ophthalmic Solution, a topical anesthetic. A drop of Genteal (Novartis, St. Louis, MO, USA) lubrican puralube eye gel was placed on the tip of the objective lens to maintain immersion contact between the objective lens and the eye. The contralateral eye was moisturized with the same gel and subsequently imaged. During the procedure, anesthetized mice were placed on a heated platform to maintain physiological temperature. Images were acquired using an HRT II/RCM (Heidelberg Engineering GmbH, Heidelberg, Germany) in vivo confocal microscope with a diode laser with a wavelength of 670 nm and a 60× objective immersion lens. An area of 400 × 400 μm with a transverse optical resolution of approximately 1 mm/pixel was imaged. For all eyes, 30 images of each layer of the cornea (total 150 images), including the superficial and basal epithelium, anterior and posterior stroma, and endothelium, were recorded.
4.5. In Vitro Immunohistochemical Analysis
To analyze corneal nerve structure and number, at least three corneas were collected from each group: young (aged between 4 and 7 weeks) WT and young TSP-1−/−, older (aged between 9 and 12 weeks) WT and older TSP-1−/− mice. Corneas were dissected and fixed in 4% paraformaldehyde for 40 min at room temperature. Four radial relaxing incisions were made before the corneas were washed with PBS with 0.1% bovine serum albumin (BSA), and then blocked for 2 h with 1% BSA in PBS and 0.1% Triton X-100. The corneas were then incubated with either anti-beta-tubulin Class III (Millipore/Chemicon rabbit anti-beta III Tubulin or T2200 Sigma, St. Louis, MO, USA) at a dilution of 1:200, anti-CGRP (Goat polyclonal Cat# 1720-9007 BioRad, Hercules, CA, USA) or anti-SubP (rat monoclonal Cat# MAB356 EMD Millipore, Burlington, MA, USA) at a dilution of 1:200 primary antibody for 72 h at 4 °C.
After four washes in PBS containing 0.1% BSA, corneas were either incubated with the appropriate secondary antibody: DyLight 488 (Donkey Anti-Rabbit IgG, Cat# 711-485-152, Jackson Immuno Research, West Grove, PA, USA) at a dilution of 1:100 or DyLight 594 (Goat Anti-Rabbit IgG, Jackson Immuno Research, Cat# 111-515-003) at a dilution of 1:200 overnight at 4 °C. The corneas stained with anti-SubP antibody were incubated with secondary antibody Cy2 (Goat Anti-Rat, Cat# ab6952, Abcam, Cambridge, UK) at a dilution of 1:200, and those stained with anti-CGRP antibody were incubated with secondary antibody Cy3 (Anti-Goat) at a dilution of 1:400. The corneas were then rinsed in PBS and mounted in a medium containing DAPI (VectaShield; Vector Laboratories, Burlingame, CA, USA).
Corneas were viewed by confocal imaging (TCS-SP2, Leica Microsystems, Bannockburn, IL, USA). Four fields of view at the level of the subbasal nerve plexus near the periphery of the cornea were photographed (2007 IOVS YuCQ Transgenic corneal neurofluorescence in mice). To view CGRP and SubP, images were acquired at 200× magnification. The density of corneal nerves was measured using NIH ImageJ with the NeuronJ plugin using established methods [
10]. Two different methods were used, one for analyzing studies using anti-beta III Tubulin only and another when analyzing CGRP and SubP in the same study. The first method involved the quantification of neural density by tracing nerves according to established methods based on the tubular nature of the staining with anti-Class III β-tubulin [
10]; the results are reported as average mm nerve/mm
2 ± SD. The second method involved the calculation of the average percent of area staining for the specific neurotransmitter using ImageJ (Fiji,
https://imagej.nih.gov/ij/), due to the discrete nature of neurotransmitter-containing vesicles stained by anti-CGRP or anti-SubP antibodies.
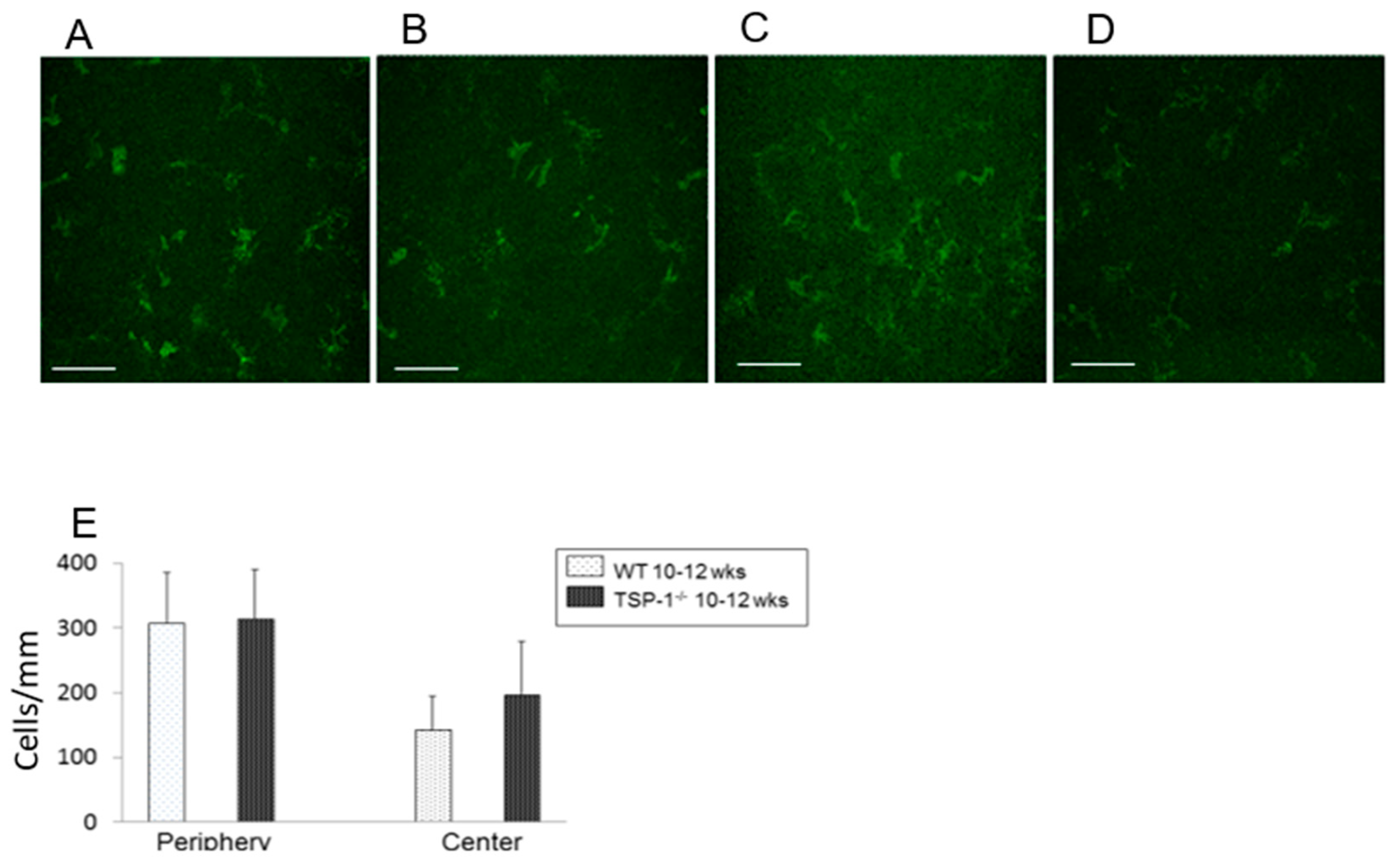
4.6. Anti-CD11b Antibody Analysis
To analyze the monocyte number of corneas, one eye from three mice per group were excised and fixed in acetone. Corneas were blocked in 2% bovine serum albumin for 45 min and then incubated with fluorescein isothiocyanate (FITC)-conjugated rat anti–mouse CD11b (marks monocyte derived cells, BD Pharmingen, San Diego, CA, USA) at ratio of 1:100 overnight at 4° [
33]. Corneas were washed and mounted using a medium containing DAPI to indicate cell nuclei. The slides were viewed by a fluorescence microscope (Eclipse E80i; Nikon, Tokyo, Japan) at a magnification of 40×. CD11b+ cells were counted in eight areas in the periphery (0.5-µm area from the limbus) and two areas in the center (central 2-µm area) of each cornea in a masked fashion [
33]. The mean number of cells was obtained by averaging the total number of cells in all the areas studied, and the result was expressed as the number of positive cells per square millimeter.
4.7. Statistical Analyses
CD 11b+ cells/mm2 were measured and the results were analyzed by unpaired t-test. A p-value < 0.05 was deemed statistically significant. Results for in vitro immunohistochemical analysis of corneas were analyzed by one-way analysis of variance (ANOVA), as appropriate, followed by the Scheffe post-hoc test. A p-value < 0.005 was deemed statistically significant. Results of qRT-PCR were analyzed by one-way analysis of variance (ANOVA), as appropriate, followed by the Scheffe post-hoc test. A p-value < 0.05 was deemed statistically significant.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





