A Novel Peroxidase Mimics and Ameliorates Alzheimer’s Disease-Related Pathology and Cognitive Decline in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
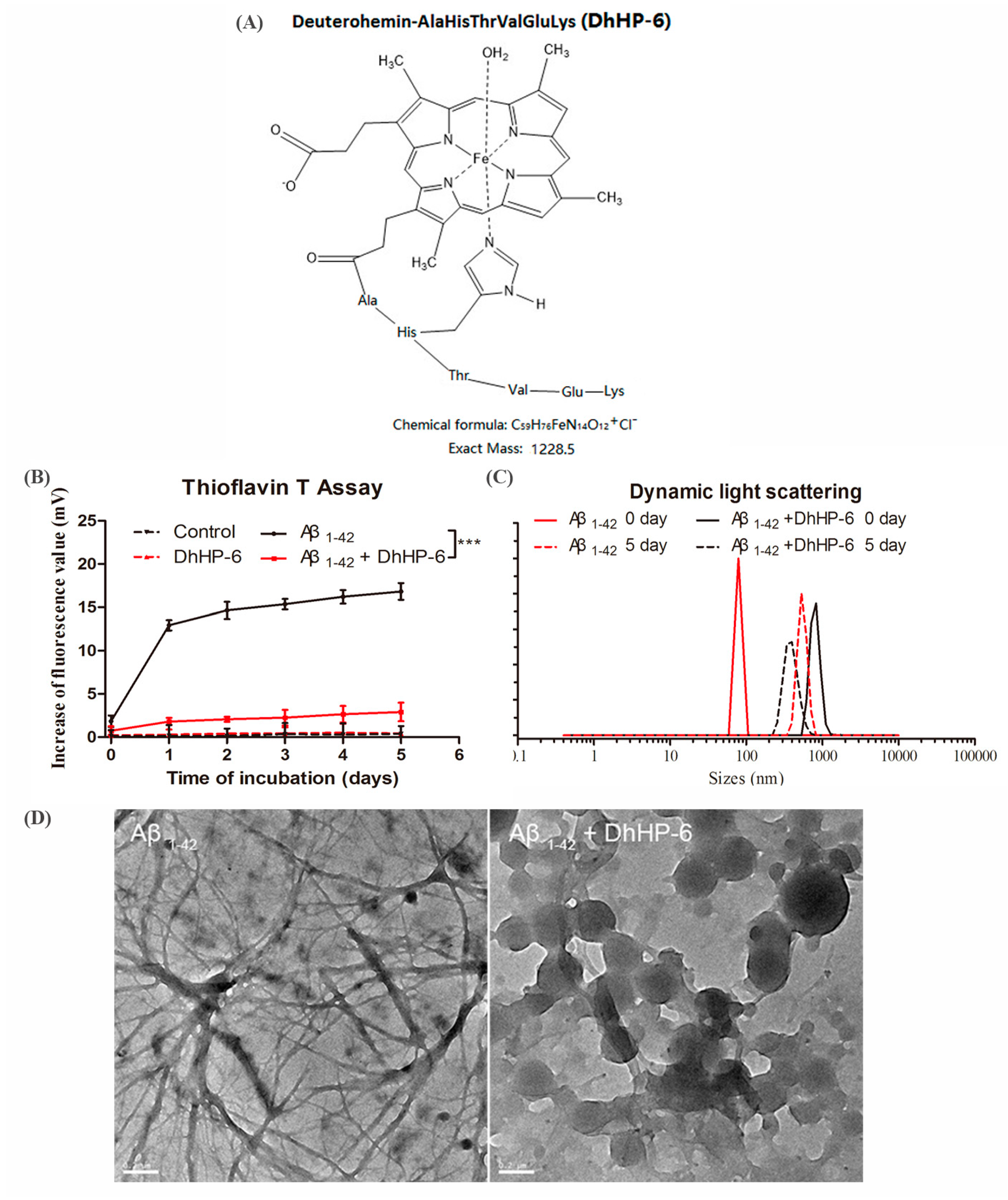
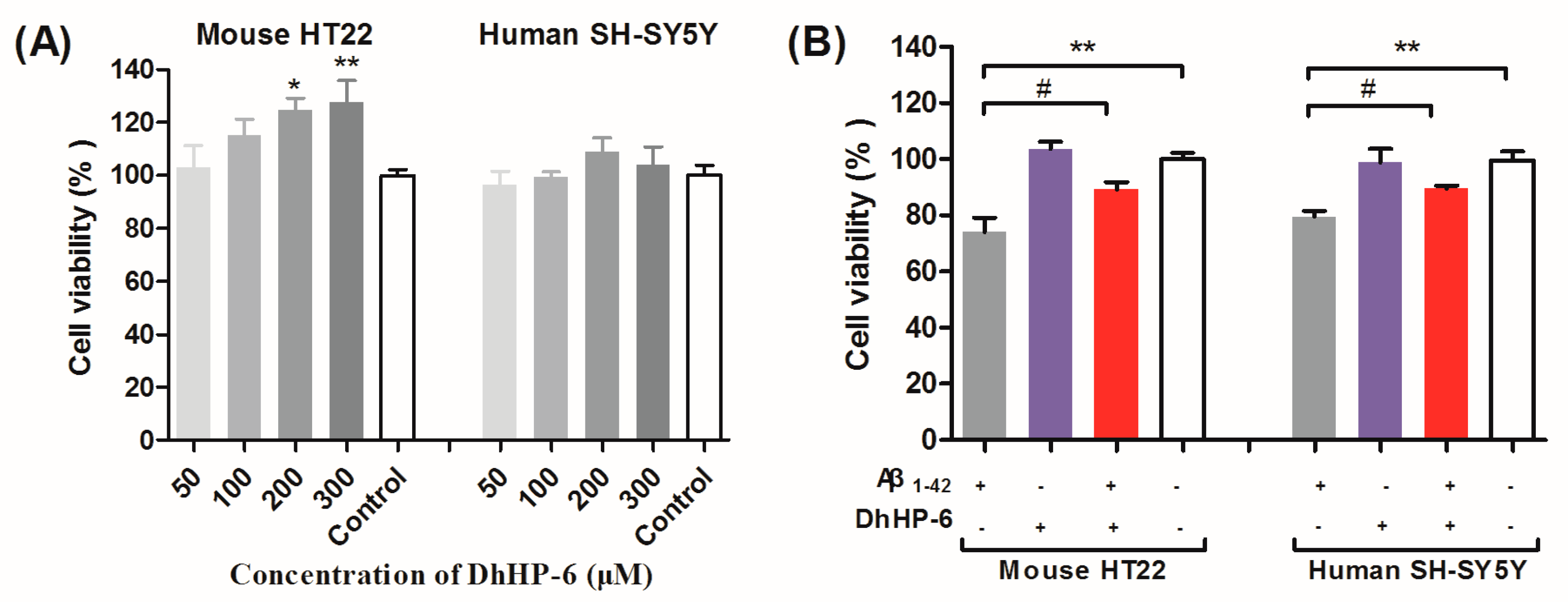
2.1. DhHP-6 Reduces the Aggregation of Aβ1-42 and Protects Neurons from Aβ Oligomers Toxicity In Vitro
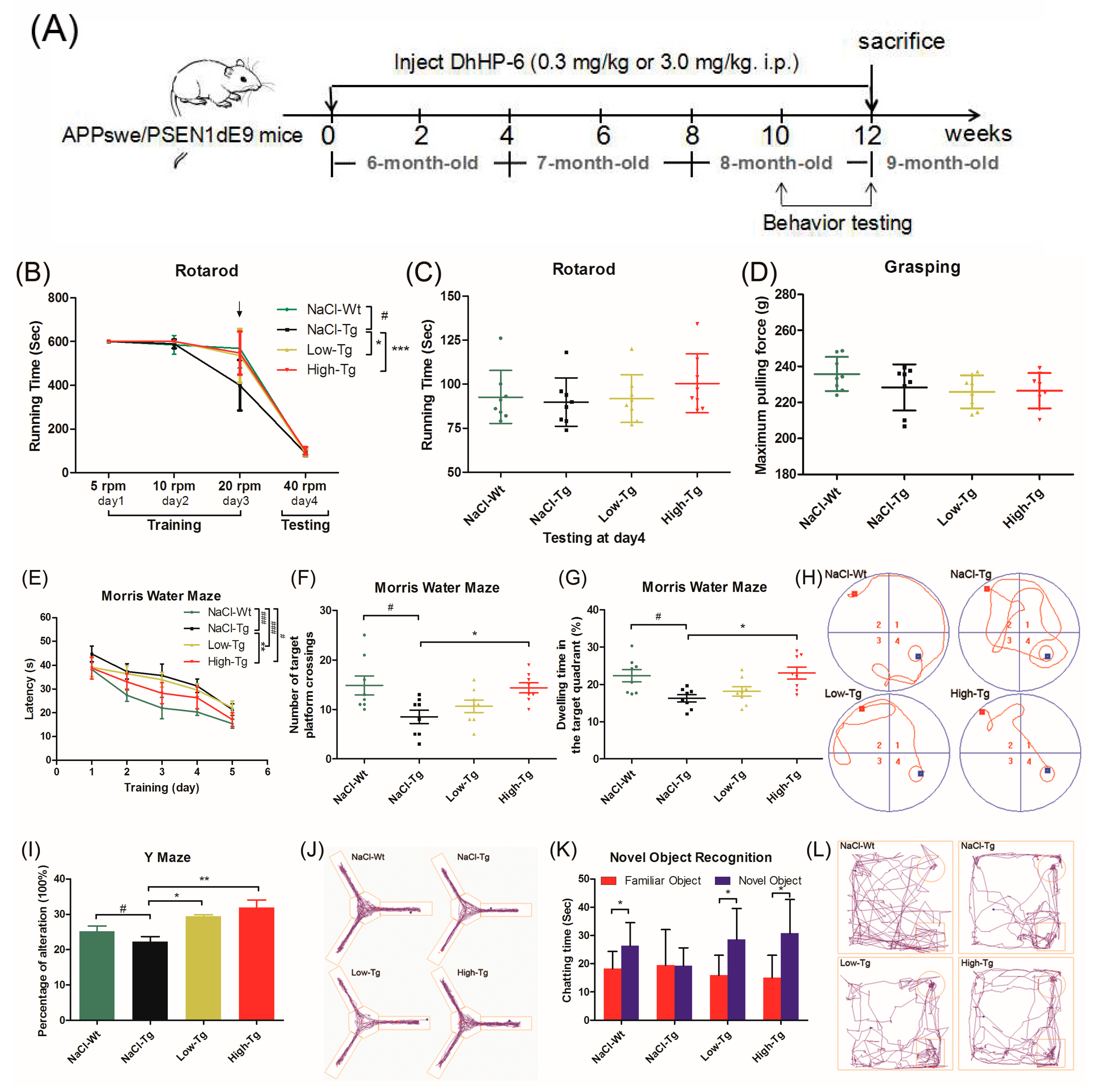
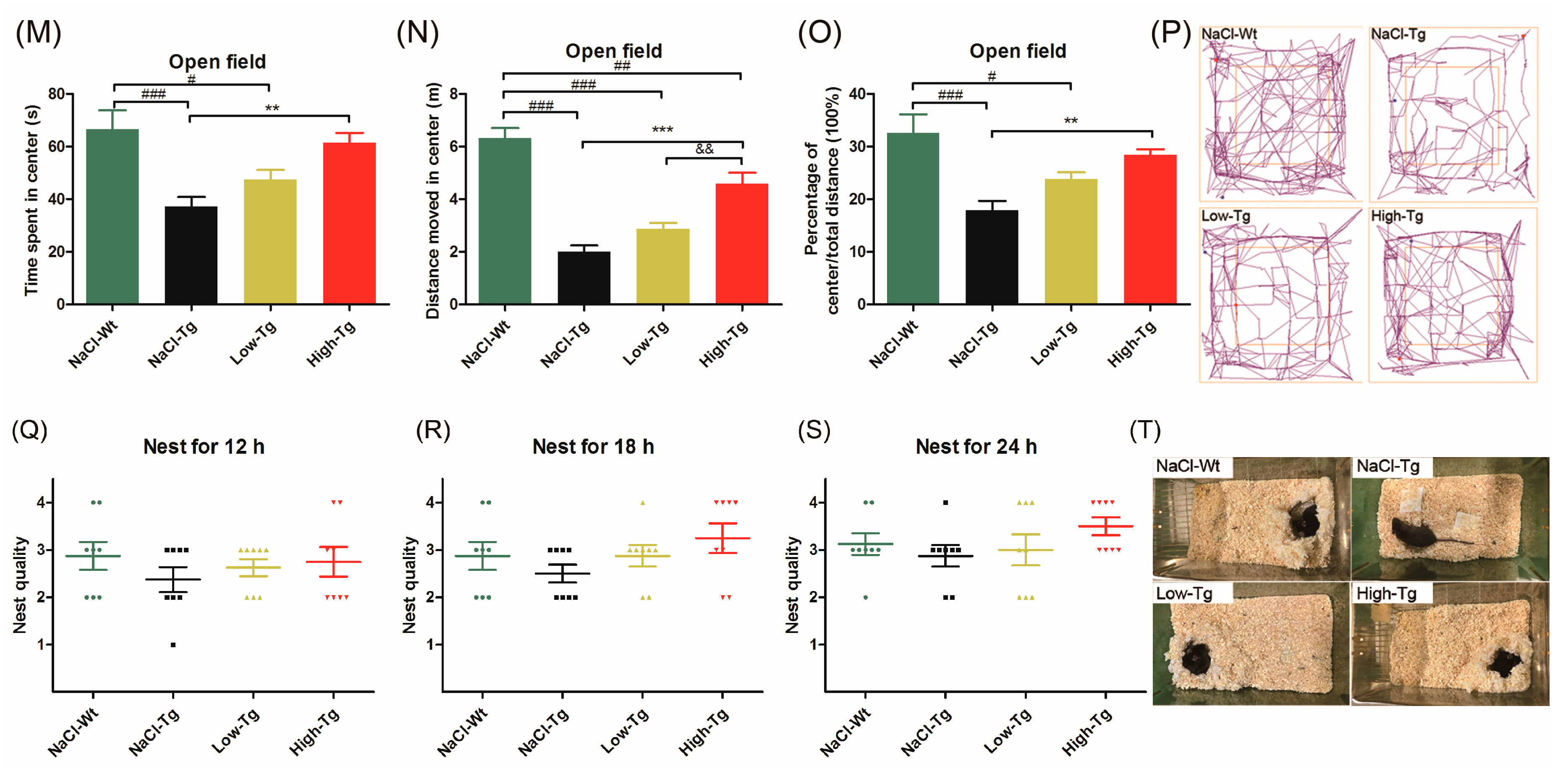
2.2. DhHP-6 Ameliorates Cognitive Decline and Improves Hippocampus Damage in APPswe/PSEN1dE9 Mice
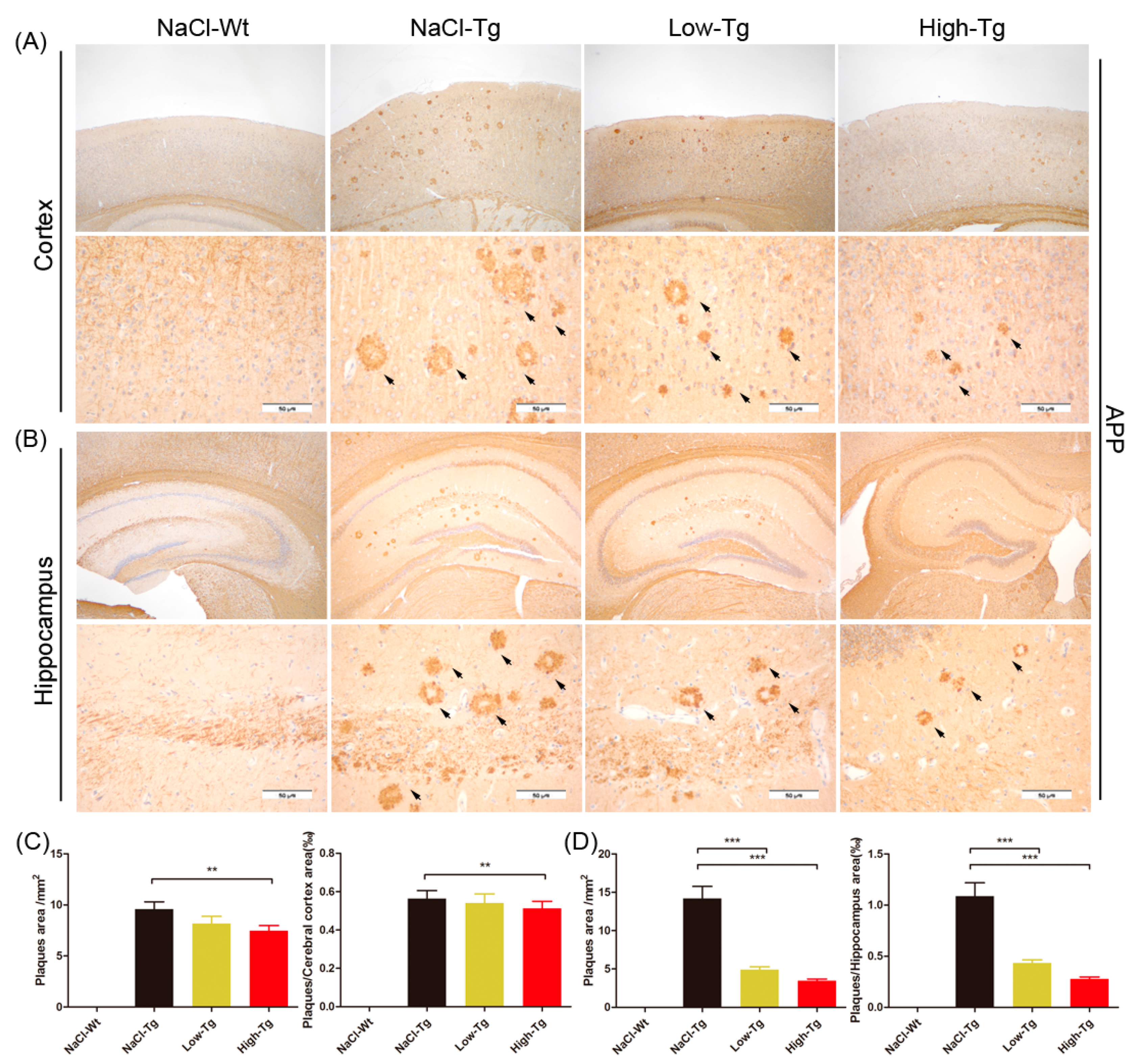
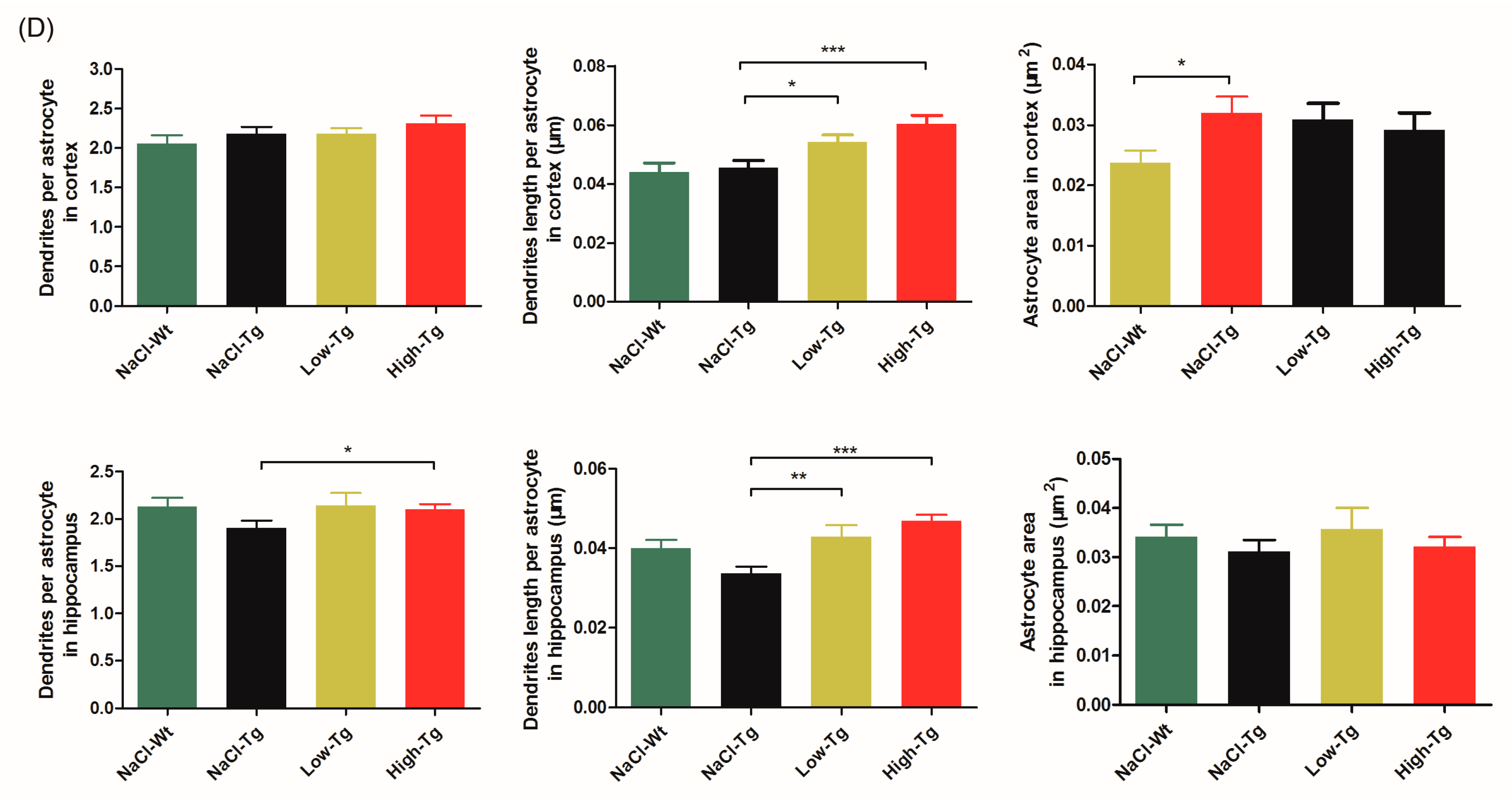
2.3. DhHP-6 Significantly Reduces Amyloid Deposition and Reverses Neuronal Atrophy in the Cortex and Hippocampus of APPswe/PSEN1dE9 Mice
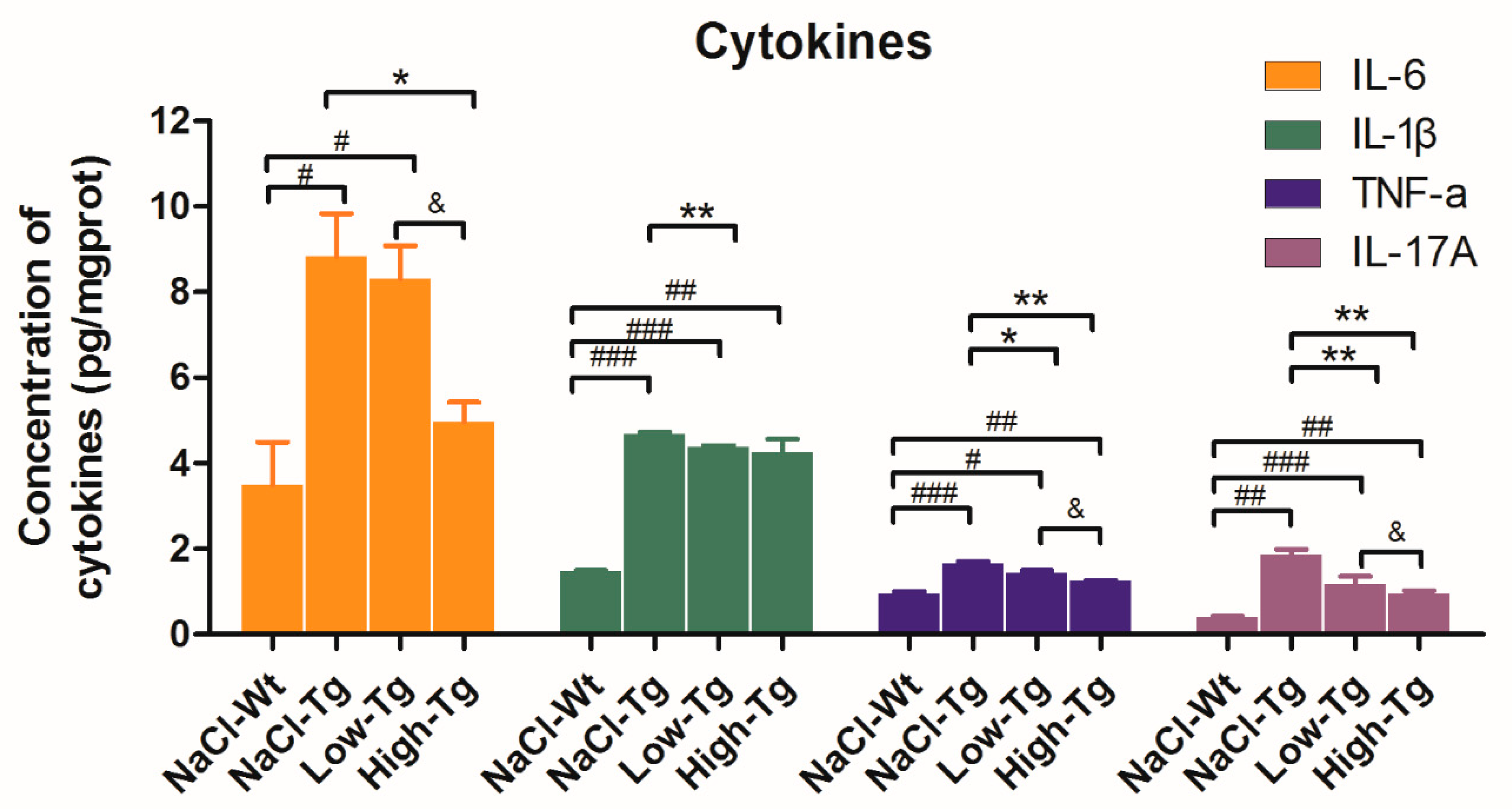
2.4. Pro-Inflammatory Cytokine Levels in DhHP-6 Treated AD Mice
3. Discussion
4. Materials and Methods
4.1. Materials and Regents
4.2. Design and Synthesis of Peroxidase Mimetic
4.3. Measurement of Aβ Fibrils by Thioflavin T Assay
4.4. Dynamic Light Scattering (DLS)
4.5. Transmission Electron Microscopy (TEM)
4.6. Cell Viability Assay
4.7. Transgenic Mice and Treatment
4.7.1. Rotary Rod
4.7.2. Grasping
4.7.3. Morris Water Maze
4.7.4. Y-Maze
4.7.5. Novel Object Recognition (NOR)
4.7.6. Open Field
4.7.7. Nest-Building Test
4.8. Immunohistochemistry
4.9. Inflammatory Cytokines
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical Approval and Informed Consent
Abbreviations
| AD | Alzheimer’s disease |
| Aβ | amyloid β |
| DhHP-6 | Deuterohemin-AlaHisThrValGluLys |
| MP-11 | microperoxidase-11 |
| C. elegans | Caenorhabditis elegans |
| APP | amyloid precursor protein |
| ROS | reactive oxygen species |
| BBB | blood-brain barrier |
| ELISA | enzyme-linked immunosorbent assay |
| Th-T | Thioflavin T |
| HFIP | hexafluoroisopropanol |
| AβO | Aβ1-42 oligomers |
| NOR | Novel Object Recognition |
| DLS | Dynamic light scattering |
| ANOVA | one-way analysis of variance |
References
- Blennow, K.; Zetterberg, H. Biomarkers for Alzheimer Disease–Current Status and Prospects for the Future. J. Intern. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fares, A.; Borrmann, D. Neurochemical Aspects of Alzheimer’s Disease and Movement Disturbances: A Theory of Beta-Amyloid and Tau-Protein. Am. J. Alzheimers Dis. Other Demen. 2018. [Google Scholar] [CrossRef] [PubMed]
- Francioso, A.; Punzi, P.; Boffi, A.; Lori, C.; Martire, S.; Giordano, C.; D’Erme, M.; Mosca, L. Beta-Sheet Interfering Molecules Acting against Beta-Amyloid Aggregation and Fibrillogenesis. Bioorg. Med. Chem. 2015, 23, 1671–1683. [Google Scholar] [CrossRef] [PubMed]
- Karran, E.; Mercken, M.; de Strooper, B. The Amyloid Cascade Hypothesis for Alzheimer’s Disease: An Appraisal for the Development of Therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Musiek, E.S.; Holtzman, D.M. Three Dimensions of the Amyloid Hypothesis: Time, Space and ‘Wingmen’. Nat. Neurosci. 2015, 18, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.P.; Clark, I.A.; Vissel, B. Inconsistencies and Controversies Surrounding the Amyloid Hypothesis of Alzheimer’s Disease. Acta Neuropathol. Commun. 2014, 2, 135. [Google Scholar] [CrossRef] [PubMed]
- Omar, S.H.; Scott, C.J.; Hamlin, A.S.; Obied, H.K. The Protective Role of Plant Biophenols in Mechanisms of Alzheimer’s Disease. J. Nutr. Biochem. 2017, 47, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Free Radicals, Reactive Oxygen Species, Oxidative Stress and Its Classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.D.; Sun, G.L.; Zhou, T.T.; Xu, X.; Zhu, Z.Y.; Rukachaisirikul, V.; Hu, L.H.; Shen, X. Small Molecule Lx2343 Ameliorates Cognitive Deficits in Ad Model Mice by Targeting Both Amyloid Beta Production and Clearance. Acta Pharmacol. Sin. 2016, 37, 1281–1297. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.D.; Hooten, N.N.; Tadokoro, T.; Lohani, A.; Barnes, J.; Evans, M.K. Alzheimer’s Disease-Associated Polymorphisms in Human Ogg1 Alter Catalytic Activity and Sensitize Cells to DNA Damage. Free Radic. Biol. Med. 2013, 63, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Albrekkan, F.M.; Kellyworden, M. Mitochondrial Dysfunction and Alzheimer’s Disease. Open J. Endocr. Metab. Dis. 2013, 3, 14–19. [Google Scholar] [CrossRef]
- Klein, W.L.; Stine, W., Jr.; Teplow, D.B. Small Assemblies of Unmodified Amyloid Beta-Protein Are the Proximate Neurotoxin in Alzheimer’s Disease. Neurobiol. Aging 2004, 25, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Mendes, D.; Oliveira, M.M.; Moreira, P.I.; Coutinho, J.; Nunes, F.M.; Pereira, D.M.; Valentao, P.; Andrade, P.B.; Videira, R.A. Beneficial Effects of White Wine Polyphenols-Enriched Diet on Alzheimer’s Disease-Like Pathology. J. Nutr. Biochem. 2018, 55, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Mucke, L. Alzheimer Mechanisms and Therapeutic Strategies. Cell 2012, 148, 1204–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auti, S.T.; Kulkarni, Y.A. A Systematic Review on the Role of Natural Products in Modulating the Pathways in Alzheimer’s Disease. Int. J. Vitam. Nutr. Res. 2018, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sawda, C.; Moussa, C.; Turner, R.S. Resveratrol for Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 2017, 1403, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.T.; Nguyen, T.H. Natural Product for the Treatment of Alzheimer’s Disease. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, W.; Zhang, Q.; Zhang, S.; Yuan, Z. Study on the Efficiency and Interaction Mechanism of a Decapeptide Inhibitor of Beta-Amyloid Aggregation. Biomacromolecules 2014, 15, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, G.; Grimaldi, M.; D’Ursi, A.M.; D’Errico, G. The Iabeta5p Beta-Breaker Peptide Regulates the Abeta(25–35) Interaction with Lipid Bilayers through a Cholesterol-Mediated Mechanism. Biochem. Biophys. Res. Commun. 2012, 417, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Sinopoli, A.; Giuffrida, A.; Tomasello, M.F.; Giuffrida, M.L.; Leone, M.; Attanasio, F.; Caraci, F.; de Bona, P.; Naletova, I.; Saviano, M.; et al. Ac-Lpffd-Th: A Trehalose-Conjugated Peptidomimetic as a Strong Suppressor of Amyloid-Beta Oligomer Formation and Cytotoxicity. Chembiochem 2016, 17, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Xiong, N.; Zhao, Y.; Dong, X.; Zheng, J.; Sun, Y. Design of a Molecular Hybrid of Dual Peptide Inhibitors Coupled on Aunps for Enhanced Inhibition of Amyloid B-Protein Aggregation and Cytotoxicity. Small 2017, 13, 1601666. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Park, S.; Allington, G.; Prelli, F.; Sun, Y.; Marta-Ariza, M.; Scholtzova, H.; Biswas, G.; Brown, B.; Verghese, P.B.; et al. Targeting Apolipoprotein E/Amyloid Beta Binding by Peptoid Cpo_Abeta17-21 P Ameliorates Alzheimer’s Disease Related Pathology and Cognitive Decline. Sci. Rep. 2017, 7, 8009. [Google Scholar] [CrossRef] [PubMed]
- Bachurin, S.O.; Bovina, E.V.; Ustyugov, A.A. Drugs in Clinical Trials for Alzheimer’s Disease: The Major Trends. Med. Res. Rev. 2017, 37, 1186–1225. [Google Scholar] [CrossRef] [PubMed]
- Jouroukhin, Y.; Ostritsky, R.; Assaf, Y.; Pelled, G.; Giladi, E.; Gozes, I. Nap (Davunetide) Modifies Disease Progression in a Mouse Model of Severe Neurodegeneration: Protection against Impairments in Axonal Transport. Neurobiol. Dis. 2013, 56, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Boxer, A.L.; Lang, A.E.; Grossman, M.; Knopman, D.S.; Miller, B.L.; Schneider, L.S.; Doody, R.S.; Lees, A.; Golbe, L.I.; Williams, D.R. Davunetide in Patients with Progressive Supranuclear Palsy: A Randomised, Double-Blind, Placebo-Controlled Phase 2/3 Trial. Lancet Neurol. 2014, 13, 676–685. [Google Scholar] [CrossRef]
- Anand, R.; Seiberling, M.; Kamtchoua, T.; Pokorny, R. Tolerability, Safety and Pharmacokinetics of the Fgll Peptide, a Novel Mimetic of Neural Cell Adhesion Molecule, Following Intranasal Administration in Healthy Volunteers. Clin. Pharmacokinet. 2007, 46, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A.J.; Wade, L.A.; Coy, D.H.; Schally, A.V.; Olson, R.D. Peptides and the Blood-Brain Barrier. Peptides 2015, 72, 16–19. [Google Scholar]
- Momenteau, M.; Rougee, M.; Loock, B. Five-Coordinate Iron-Porphyrin as a Model for the Active Site of Hemoproteins. Characterization and Coordination Properties. Eur. J. Biochem. 1976, 71, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, J.; Nakano, K. Totally Synthetic Microperoxidase-11. R. Soc. Open Sci. 2018, 5, 172311. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Guo, L.; Roeske, R.; Luo, G.M.; Wei, L.I. The Method Improvement of Synthyesis and Purification of Deuterohemin. Acta Sci. Nat. Univ. Jilinensis 2001, 91–92. [Google Scholar]
- Wang, L.P.; Liu, Y.L.; Yang, H.; Wei, L.I. Synthesis and Anti-Cataract Activity of a Novel Peroxidase Mimetics. Chem. Res. Chin. Univ. 2004, 25, 2171–2173. [Google Scholar]
- Guan, S.; Li, P.; Luo, J.; Li, Y.; Huang, L.; Wang, G.; Zhu, L.; Fan, H.; Li, W.; Wang, L. A Deuterohemin Peptide Extends Lifespan and Increases Stress Resistance in Caenorhabditis Elegans. Free Radic. Res. 2010, 44, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, P.; Wang, G.; Guan, S.; Sun, X.; Wang, L. Dhhp-6 Extends Lifespan of Caenorhabditis Elegans by Enhancing Nuclear Translocation and Transcriptional Activity of Daf-16. Free Radic. Res. 2013, 47, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Rajasekhar, K.; Mehta, K.; Govindaraju, T. Hybrid Multifunctional Modulators Inhibit Multifaceted Abeta Toxicity and Prevent Mitochondrial Damage. ACS Chem. Neurosci. 2018, 9, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Scholtzova, H.; Do, E. Innate Immunity Stimulation Via Toll-Like Receptor 9 Ameliorates Vascular Amyloid Pathology in Tg-Swdi Mice with Associated Cognitive Benefits. J. Neurosci. 2017, 37, 936–959. [Google Scholar] [CrossRef] [PubMed]
- Garcez, M.L.; Mina, F.; Bellettini-Santos, T.; da Luz, A.P.; Schiavo, G.L.; Macieski, J.M.C.; Medeiros, E.B.; Marques, A.O.; Magnus, N.Q.; Budni, J. The Involvement of Nlrp3 on the Effects of Minocycline in an Ad-Like Pathology Induced by Beta-Amyloid Oligomers Administered to Mice. Mol Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, J.; Urdzikova, L.M.; Svobodova, B.; Amin, A.G.; Karova, K.; Dubisova, J.; Zaviskova, K.; Kubinova, S.; Schmidt, M.; Jhanwar-Uniyal, M. Does Combined Therapy of Curcumin and Epigallocatechin Gallate Have a Synergistic Neuroprotective Effect against Spinal Cord Injury? Neural Regen. Res. 2018, 13, 119–127. [Google Scholar] [PubMed]
- Fu, L.; Li, Y.; Hu, Y.; Zheng, Y.; Yu, B.; Zhang, H.; Wu, J.; Wu, H.; Yu, X.; Kong, W. Norovirus P Particle-Based Active Abeta Immunotherapy Elicits Sufficient Immunogenicity and Improves Cognitive Capacity in a Mouse Model of Alzheimer’s Disease. Sci. Rep. 2017, 7, 41041. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P. Burrowing and Nest Building Behavior as Indicators of Well-Being in Mice. J. Neurosci. Methods 2014, 234, 139–146. [Google Scholar] [PubMed]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s Disease: The Amyloid Cascade Hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, D.S.; Murai, K.K. Synergistic Actions of Microglia and Astrocytes in the Progression of Alzheimer’s Disease. J. Alzheimers Dis. 2015, 45, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate Mapping Analysis Reveals That Adult Microglia Derive from Primitive Macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, A.; de Chaves, E.P. Abeta Internalization by Neurons and Glia. Int. J. Alzheimers Dis. 2011, 2011, 127984. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active Sensor and Versatile Effector Cells in the Normal and Pathologic Brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- McKoy, A.F.; Chen, J.; Schupbach, T.; Hecht, M.H. A Novel Inhibitor of Amyloid Beta (Abeta) Peptide Aggregation: From High Throughput Screening to Efficacy in an Animal Model of Alzheimer Disease. J. Biol. Chem. 2012, 287, 38992–39000. [Google Scholar] [CrossRef] [PubMed]
- Glabe, C.G. Structural Classification of Toxic Amyloid Oligomers. J. Biol. Chem. 2008, 283, 29639–29643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, C.; Zhao, L.; Zhu, B.; Li, Q.; Yew, D.T.; Yao, Z.; Xu, J. Protective Effects of Ginkgo Biloba Extract (Egb761) and Its Constituents Quercetin and Ginkgolide B against Beta-Amyloid Peptide-Induced Toxicity in Sh-Sy5y Cells. Chem. Biol. Interact. 2009, 181, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Necula, M.; Kayed, R.; Milton, S.; Glabe, C.G. Small Molecule Inhibitors of Aggregation Indicate That Amyloid Beta Oligomerization and Fibrillization Pathways Are Independent and Distinct. J. Biol. Chem. 2007, 282, 10311–10324. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Paul, S. Action of Caffeine as an Amyloid Inhibitor in the Aggregation of Abeta16-22 Peptides. J. Phys. Chem. B 2016, 120, 9019–9033. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ma, H.; Wang, X.; Zhao, Z.; Zhang, Y.; Zhao, B.; Guo, Y.; Xu, L. A Tetrapeptide from Maize Protects a Transgenic Caenorhabditis Elegans Aβ1-42 Model from Aβ-Induced Toxicity. RSC Adv. 2016, 6, 56851–56858. [Google Scholar] [CrossRef]
- Barsyte, D.; Lovejoy, D.A.; Lithgow, G.J. Longevity and Heavy Metal Resistance in Daf-2 and Age-1 Long-Lived Mutants of Caenorhabditis Elegans. FASEB J. 2001, 15, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of Aging and Age-Related Disease by Daf-16 and Heat-Shock Factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Balbach, J.J.; Ishii, Y.; Antzutkin, O.N.; Leapman, R.D.; Rizzo, N.W.; Dyda, F.; Reed, J.; Tycko, R. Amyloid Fibril Formation by a Beta 16-22, a Seven-Residue Fragment of the Alzheimer’s Beta-Amyloid Peptide, and Structural Characterization by Solid State Nmr. Biochemistry 2000, 39, 13748–13759. [Google Scholar] [CrossRef] [PubMed]
- Jankowsky, J.L.; Slunt, H.H.; Ratovitski, T.; Jenkins, N.A.; Copeland, N.G.; Borchelt, D.R. Co-Expression of Multiple Transgenes in Mouse Cns: A Comparison of Strategies. Biomol. Eng. 2001, 17, 157–165. [Google Scholar] [CrossRef]
- Huang, H.; Nie, S.; Cao, M.; Marshall, C.; Gao, J.; Xiao, N.; Hu, G.; Xiao, M. Characterization of Ad-Like Phenotype in Aged Appswe/Ps1de9 Mice. Age (Dordr) 2016, 38, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Ono, F.; Nakamura, S.; Matsumoto, S.E.; Jin, H.; Hattori, N.; Tabira, T. An Oral Abeta Vaccine Using a Recombinant Adeno-Associated Virus Vector in Aged Monkeys: Reduction in Plaque Amyloid and Increase in Abeta Oligomers. J. Alzheimers Dis. 2016, 54, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Arndt, J.W.; Qian, F.; Smith, B.A.; Quan, C.; Kilambi, K.P.; Bush, M.W.; Walz, T.; Pepinsky, R.B.; Bussiere, T.; Hamann, S.; et al. Structural and Kinetic Basis for the Selectivity of Aducanumab for Aggregated Forms of Amyloid-Beta. Sci. Rep. 2018, 8, 6412. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, T.; Castano, E.M.; Golabek, A.; Vogel, T.; Frangione, B. Acceleration of Alzheimer’s Fibril Formation by Apolipoprotein E in Vitro. Am. J. Pathol. 1994, 145, 1030–1035. [Google Scholar] [PubMed]
- Sathya, S.; Shanmuganathan, B.; Manirathinam, G.; Ruckmani, K.; Devi, K.P. A-Bisabolol Loaded Solid Lipid Nanoparticles Attenuates Aβ Aggregation and Protects Neuro2a Cells from Aβ Induced Neurotoxicity. J. Mol. Liq. 2018, 264, 431–441. [Google Scholar] [CrossRef]
- Parsons, C.G.; Ruitenberg, M.; Freitag, C.E.; Sroka-Saidi, K.; Russ, H.; Rammes, G. MRZ-99030–A Novel Modulator of Abeta Aggregation: I–Mechanism of Action (MoA) Underlying the Potential Neuroprotective Treatment of Alzheimer’s Disease, Glaucoma and Age-Related Macular Degeneration (AMD). Neuropharmacology 2015, 92, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Stine, W.B.; Jungbauer, L.; Yu, C.; LaDu, M.J. Preparing Synthetic Abeta in Different Aggregation States. Methods Mol. Biol. 2011, 670, 13–32. [Google Scholar] [PubMed]
- Zhang, J.H.; Yu, L.J.; Yang, H.; Hui, Z.; Jiang, S.; Chen, L.; Zhao, Y.; Wang, S.L.; Liu, Y.; Xu, Y. Huatuo Zaizao Pill Ameliorates Cognitive Impairment of App/Ps1 Transgenic Mice by Improving Synaptic Plasticity and Reducing Abeta Deposition. BMC Complement Altern Med. 2018, 18, 167. [Google Scholar] [CrossRef] [PubMed]
- Machova, U.L.; Karova, K.; Ruzicka, J.; Kloudova, A.; Shannon, C.; Dubisova, J.; Murali, R.; Kubinova, S.; Sykova, E.; Jhanwar-Uniyal, M. The Anti-Inflammatory Compound Curcumin Enhances Locomotor and Sensory Recovery after Spinal Cord Injury in Rats by Immunomodulation. Int. J. Mol. Sci. 2016, 17, 49. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The Novel Object Recognition Memory: Neurobiology, Test Procedure, and Its Modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Short, J.L.; Newman, S.A.; Choy, K.H.C.; Tiwari, D.; Yap, C.; Senyschyn, D.; Banks, W.A.; Nicolazzo, J.A. Cognitive Benefits of Lithium Chloride in APP/PS1 Mice Are Associated with Enhanced Brain Clearance of Beta-Amyloid. Brain Behav. Immun. 2018, 70, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Christakis, D.A.; Ramirez, J.S.; Ramirez, J.M. Overstimulation of Newborn Mice Leads to Behavioral Differences and Deficits in Cognitive Performance. Sci. Rep. 2012, 2, 546. [Google Scholar] [CrossRef]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Wang, K.; Yuan, Y.; Li, H.; Zhang, R.; Guan, S.; Wang, L. A Novel Peroxidase Mimics and Ameliorates Alzheimer’s Disease-Related Pathology and Cognitive Decline in Mice. Int. J. Mol. Sci. 2018, 19, 3304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113304
Xu J, Wang K, Yuan Y, Li H, Zhang R, Guan S, Wang L. A Novel Peroxidase Mimics and Ameliorates Alzheimer’s Disease-Related Pathology and Cognitive Decline in Mice. International Journal of Molecular Sciences. 2018; 19(11):3304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113304
Chicago/Turabian StyleXu, Jia, Kai Wang, Ye Yuan, Hui Li, Ruining Zhang, Shuwen Guan, and Liping Wang. 2018. "A Novel Peroxidase Mimics and Ameliorates Alzheimer’s Disease-Related Pathology and Cognitive Decline in Mice" International Journal of Molecular Sciences 19, no. 11: 3304. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113304