Comparative Analyses of Cytochrome P450s and Those Associated with Secondary Metabolism in Bacillus Species

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
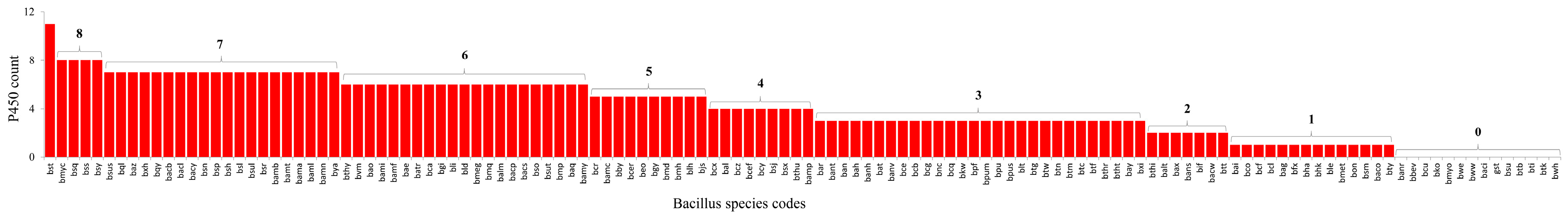
2.1. Bacillus Species Have the Lowest Number of P450s
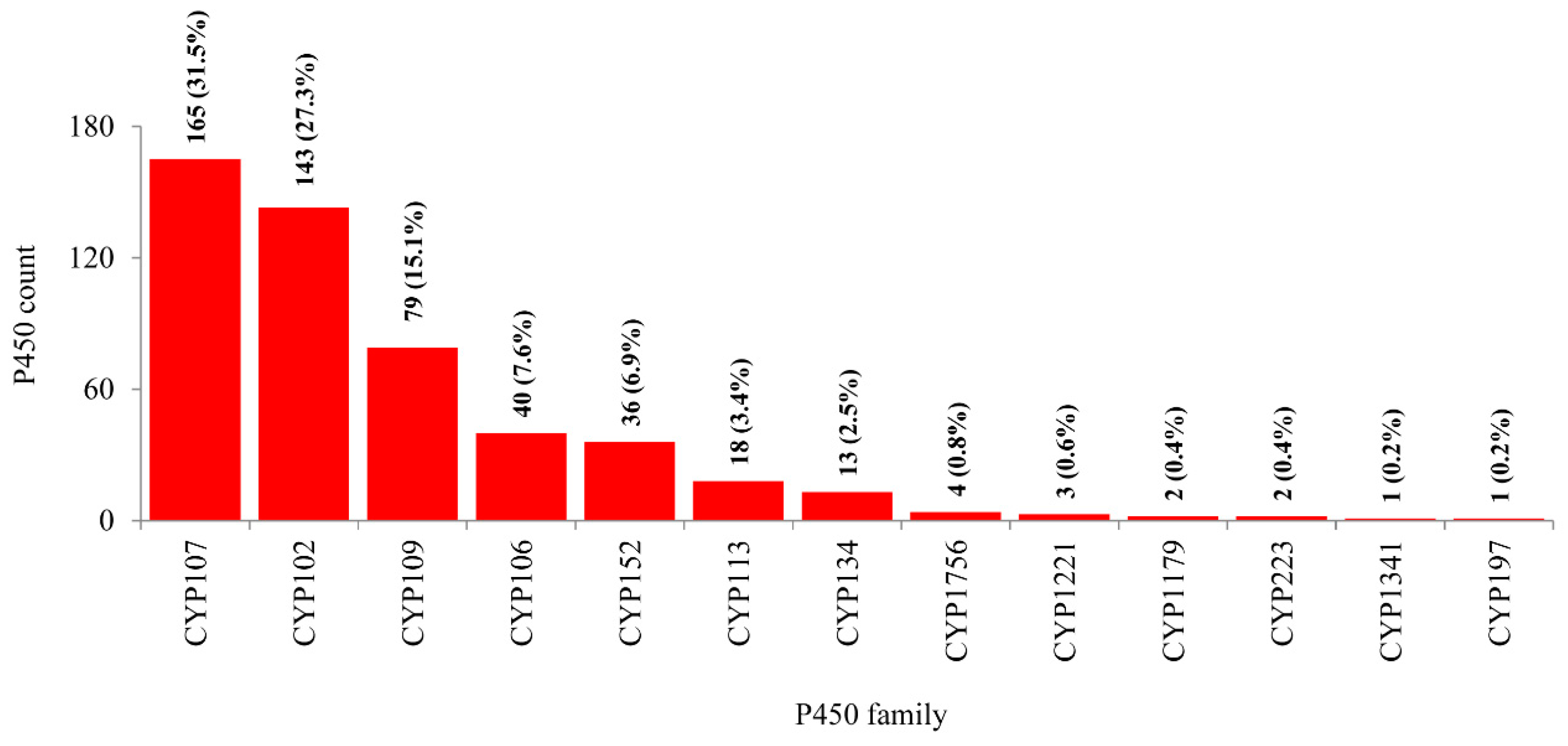
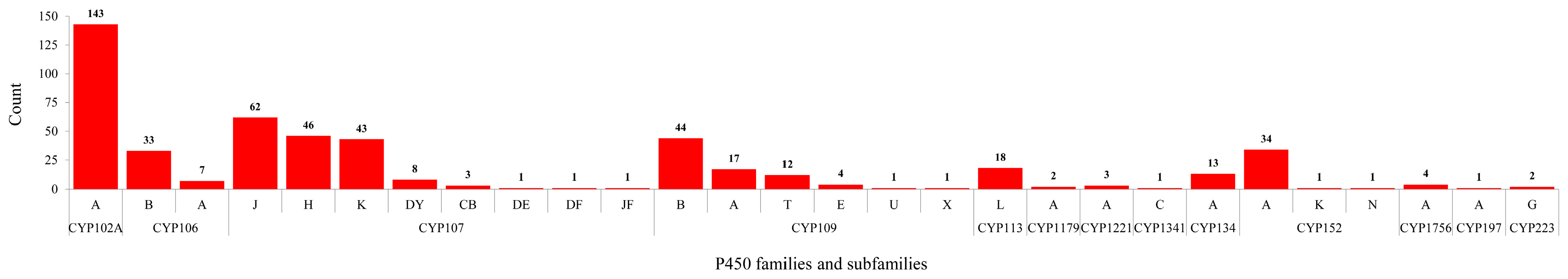
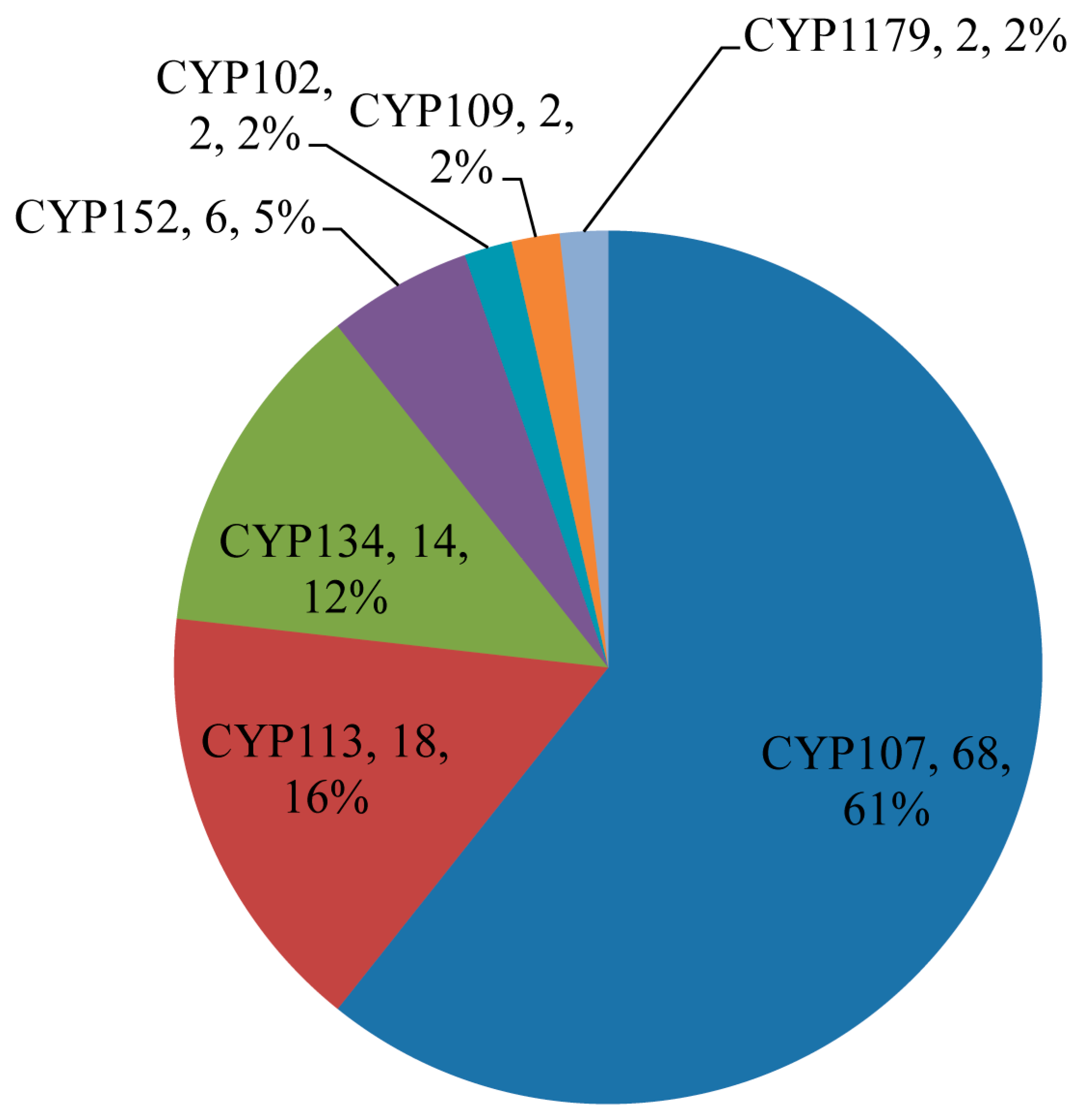
2.2. Bacillus Species Have the Lowest Number of P450 Families and Subfamilies’
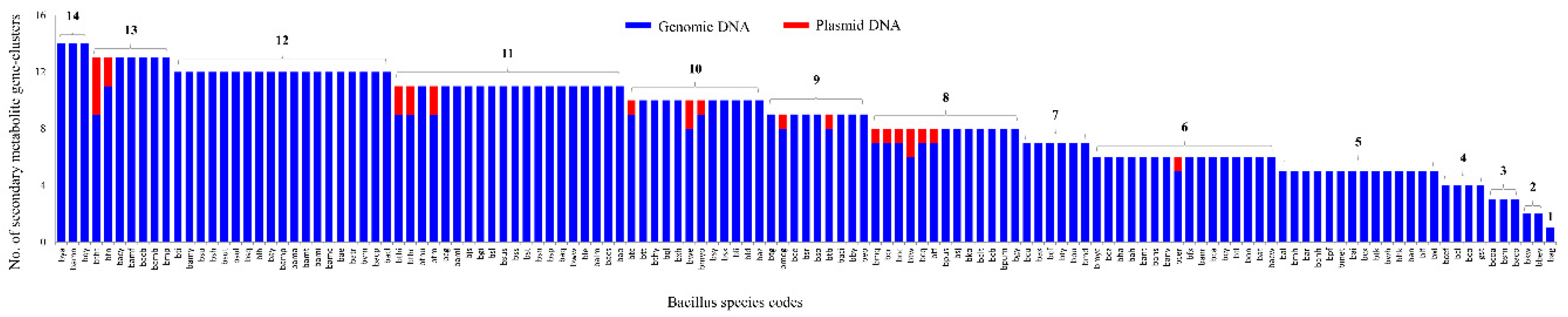
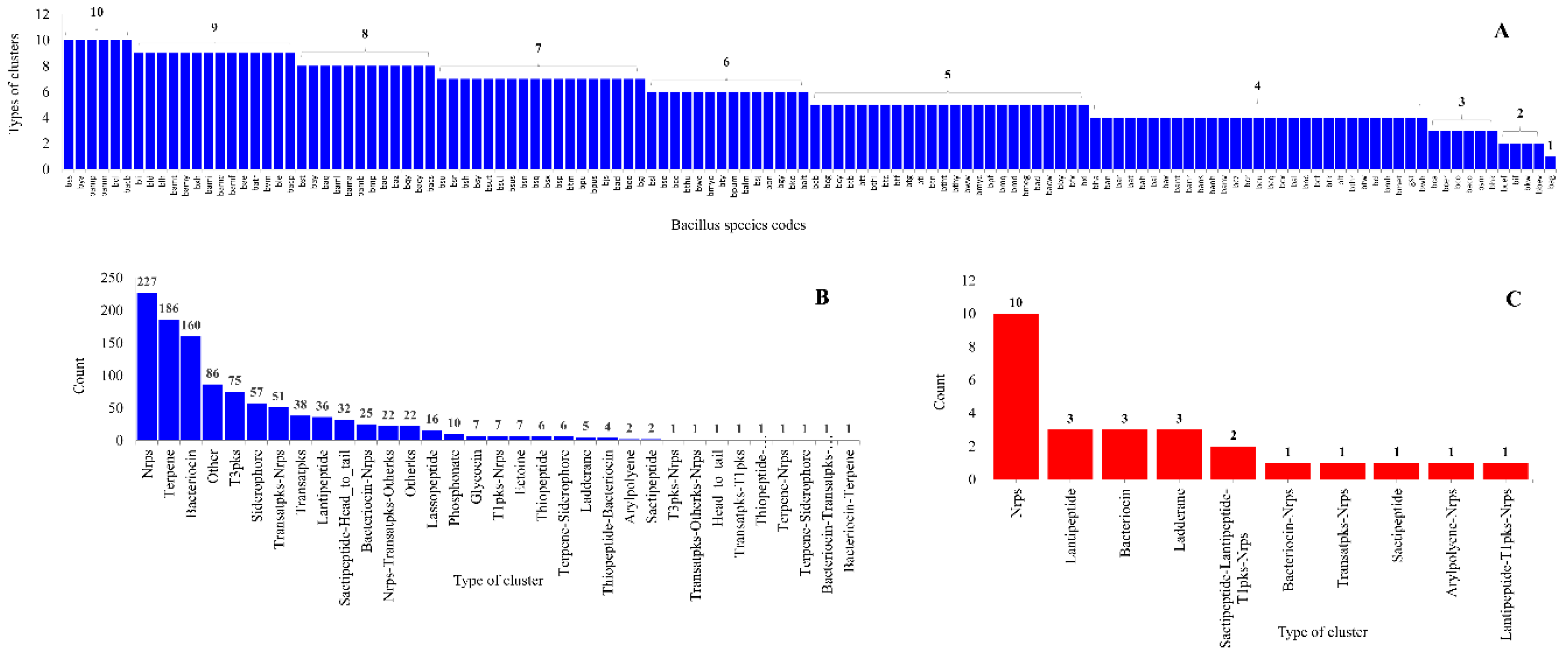
2.3. Bacillus Species Have the Lowest Number of Secondary Metabolite BGCs
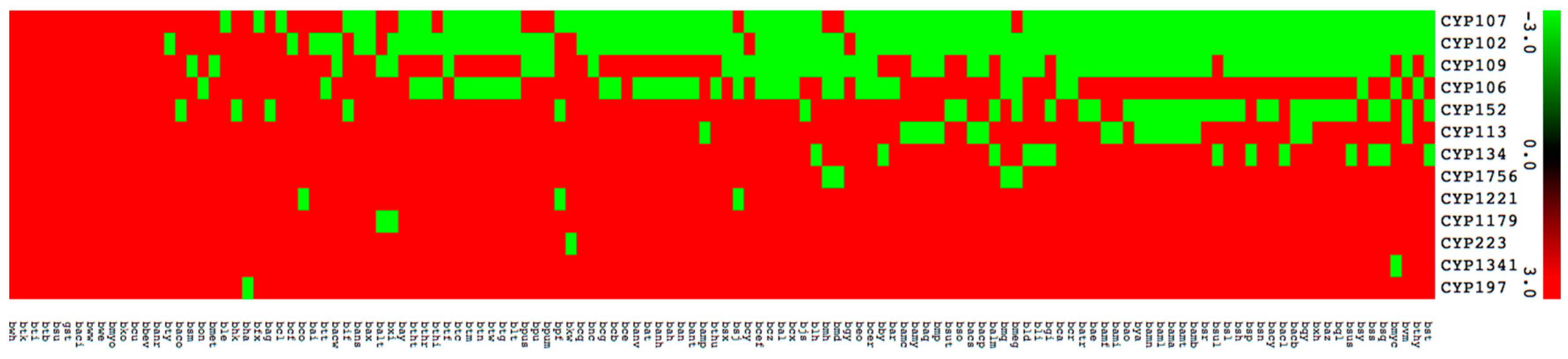
2.4. Large Number of P450s Found to Be Part of Secondary Metabolites BGCs in Bacillus Species
2.5. Bacillus P450s Indeed Involved in the Synthesis of Secondary Metabolites
3. Materials and Methods
3.1. Species and Database
3.2. Genome Data Mining and Annotation of P450s
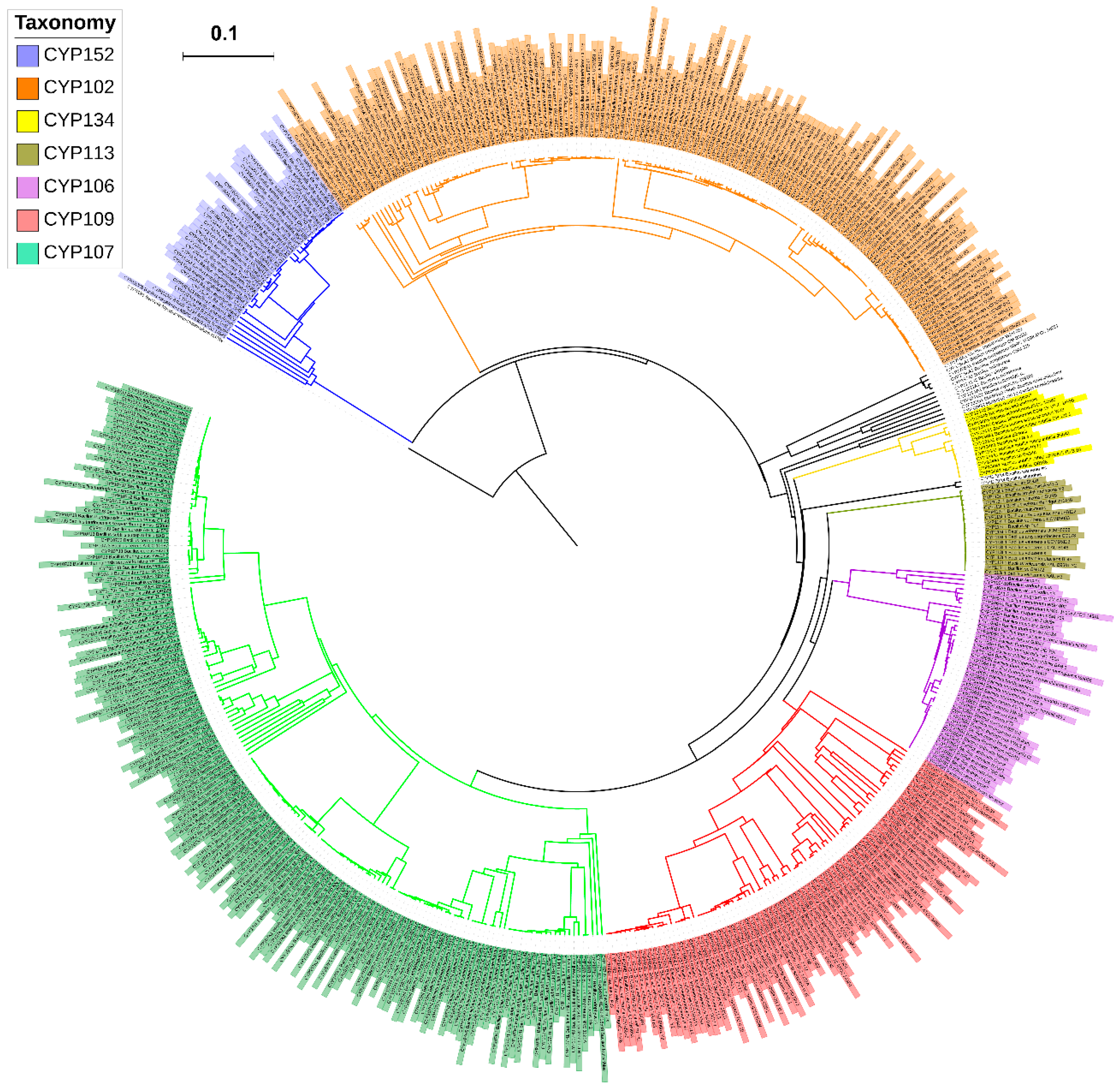
3.3. Phylogenetic Analysis of P450s
3.4. P450 Diversity Percentage Analysis
3.5. Generation of P450 Profile Heat-Maps
3.6. Secondary Metabolite BGCs Analysis
3.7. Comparative Analysis of P450s
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sono, M.; Roach, M.P.; Coulter, E.D.; Dawson, J.H. Heme-containing oxygenases. Chem. Rev. 1996, 96, 2841–2888. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Isin, E.M.; Guengerich, F.P. Complex reactions catalyzed by cytochrome P450 enzymes. Biochim. Biophys. Acta (BBA) Gen. Subj. 2007, 1770, 314–329. [Google Scholar] [CrossRef] [PubMed]
- Fasan, R. Tuning P450 enzymes as oxidation catalysts. ACS Catal. 2012, 2, 647–666. [Google Scholar] [CrossRef]
- Syed, K.; Porollo, A.; Lam, Y.W.; Grimmett, P.E.; Yadav, J.S. CYP63A2, a catalytically versatile fungal P450 monooxygenase capable of oxidizing higher-molecular-weight polycyclic aromatic hydrocarbons, alkylphenols, and alkanes. Appl. Environ. Microbiol. 2013, 79, 2692–2702. [Google Scholar] [CrossRef] [PubMed]
- Le-Huu, P.; Heidt, T.; Claasen, B.; Laschat, S.; Urlacher, V.B. Chemo-, regio-, and stereoselective oxidation of the monocyclic diterpenoid β-cembrenediol by P450 BM3. ACS Catal. 2015, 5, 1772–1780. [Google Scholar] [CrossRef]
- Syed, K.; Yadav, J.S. P450 monooxygenases (P450ome) of the model white rot fungus Phanerochaete chrysosporium. Crit. Rev. Microbiol. 2012, 38, 339–363. [Google Scholar] [CrossRef] [PubMed]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [PubMed]
- McLean, K.J.; Leys, D.; Munro, A.W. Microbial cytochrome P450s. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Montellano, P.R.O., Ed.; Springer: Basel, Switzerland, 2015; pp. 261–407. ISBN 978-3-319-12108-6. [Google Scholar]
- Guengerich, F.P. Human cytochrome P450 enzymes. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Montellano, P.R.O., Ed.; Springer: Basel, Switzerland, 2015; pp. 523–785. ISBN 978-3-319-12108-6. [Google Scholar]
- Girhard, M.; Bakkes, P.J.; Mahmoud, O.; Urlacher, V.B. P450 Biotechnology. In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Montellano, P.R.O., Ed.; Springer: Basel, Switzerland, 2015; pp. 451–520. ISBN 978-3-319-12108-6. [Google Scholar]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Lei, L.; Warrilow, A.G.; Lepesheva, G.I.; Mullins, J.G.; Waterman, M.R.; Kelly, S.L. The first virally encoded cytochrome p450. J. Virol. 2009, 83, 8266–8269. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.B.; Eiben, S. Cytochrome P450 monooxygenases: Perspectives for synthetic application. Trends Biochem. Sci. 2006, 24, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.B.; Girhard, M. Cytochrome P450 monooxygenases: An update on perspectives for synthetic application. Trends Biochem. Sci. 2012, 30, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Brown, L.C.W.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, D.R.; Perkins, J.B. The genus Bacillus. In Practical Handbook of Microbiology, 2nd ed.; Goldman, E., Green, L.H., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2009; pp. 309–337. ISBN 978-0-8493-9365-5. [Google Scholar] [CrossRef]
- Graumann, P. Bacillus: Cellular and Molecular Biology, 2nd ed.; Caister Academic Press: Haverhill, UK, 2012; ISBN 978-1-904455-97-4. [Google Scholar]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [PubMed]
- Grubbs, K.J.; Bleich, R.M.; Santa Maria, K.C.; Allen, S.E.; Farag, S.; Team, A.; Shank, E.A.; Bowers, A.A. Large-scale bioinformatics analysis of Bacillus genomes uncovers conserved roles of natural products in Bacterial physiology. MSystems 2017, 2, e00040-17. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Shibata, D.; Kino, K. Phylogenetic analysis of Bacillus P450 monooxygenases and evaluation of their activity towards steroids. Steroid 2009, 74, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Ruettinger, R.T.; Wen, L.P.; Fulco, A.J. Coding nucleotide, 5′ regulatory, and deduced amino acid sequences of P-450BM-3, a single peptide cytochrome P-450: NADPH-P-450 reductase from Bacillus megaterium. J. Biol. Chem. 1989, 264, 10987–10995. [Google Scholar] [PubMed]
- Munro, A.W.; Leys, D.G.; McLean, K.J.; Marshall, K.R.; Ost, T.W.; Daff, S.; Miles, C.S.; Chapman, S.K.; Lysek, D.A.; Moser, C.C.; et al. P450 BM3: The very model of a modern flavocytochrome. Trends Biochem. Sci. 2002, 27, 250–257. [Google Scholar] [CrossRef]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2015, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. AntiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed]
- Syed, K.; Shale, K.; Pagadala, N.S.; Tuszynski, J. Systematic identification and evolutionary analysis of catalytically versatile cytochrome P450 monooxygenase families enriched in model basidiomycete fungi. PLoS ONE 2014, 9, e86683. [Google Scholar] [CrossRef] [PubMed]
- Kgosiemang, I.K.R.; Mashele, S.S.; Syed, K. Comparative genomics and evolutionary analysis of cytochrome P450 monooxygenases in fungal subphylum Saccharomycotina. J. Pure Appl. Microbiol. 2014, 8, 291–302. [Google Scholar]
- Qhanya, L.B.; Matowane, G.; Chen, W.; Sun, Y.; Letsimo, E.M.; Parvez, M.; Yu, J.H.; Mashele, S.S.; Syed, K. Genome-wide annotation and comparative analysis of cytochrome P450 monooxygenases in Basidiomycete biotrophic plant pathogens. PLoS ONE 2015, 10, e0142100. [Google Scholar] [CrossRef] [PubMed]
- Ngwenya, M.L.; Chen, W.; Basson, A.K.; Shandu, J.S.; Yu, J.H.; Nelson, D.R.; Syed, K. Blooming of unusual cytochrome P450s by tandem duplication in the pathogenic fungus Conidiobolus coronatus. Int. J. Mol. Sci. 2018, 19, 1711. [Google Scholar] [CrossRef] [PubMed]
- Sello, M.M.; Jafta, N.; Nelson, D.R.; Chen, W.; Yu, J.H.; Parvez, M.; Kgosiemang, I.K.R.; Monyaki, R.; Raselemane, S.C.; Qhanya, L.B.; et al. Diversity and evolution of cytochrome P450 monooxygenases in Oomycetes. Sci. Rep. 2015, 5, 11572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.C.; Loveridge, E.J. CYP 105—Diverse structures, functions and roles in an intriguing family of enzymes in Streptomyces. J. Appl. Microbiol. 2014, 117, 1549–1563. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Poulos, T.L. The structure of the cytochrome p450BM-3 haem domain complexed with the fatty acid substrate, palmitoleic acid. Nat. Struct. Biol. 1997, 4, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Noble, M.A.; Miles, C.S.; Reid, G.A.; Chapman, S.K.; Munro, A.W. Catalytic properties of key active site mutants of flavocytochrome P-450 BM3. Biochem. Soc. Trans. 1999, 27, A44. [Google Scholar] [CrossRef]
- Lee, D.S.; Yamada, A.; Matsunaga, I.; Ichihara, K.; Adachi, S.I.; Park, S.Y.; Shiro, Y. Crystallization and preliminary X-ray diffraction analysis of fatty-acid hydroxylase cytochrome P450BSβ from Bacillus subtilis. Acta Crystallogr. D Struct. Biol. 2002, 58, 687–689. [Google Scholar] [CrossRef]
- Lee, D.S.; Yamada, A.; Sugimoto, H.; Matsunaga, I.; Ogura, H.; Ichihara, K.; Adachi, S.I.; Park, S.Y.; Shiro, Y. Substrate recognition and molecular mechanism of fatty acid hydroxylation by cytochrome P450 from Bacillus subtilis: Crystallographic, spectroscopic and mutational studies. J. Biol. Chem. 2003, 278, 9761–9767. [Google Scholar] [CrossRef] [PubMed]
- Cryle, M.J.; Bell, S.G.; Schlichting, I. Structural and biochemical characterization of the cytochrome P450 CypX (CYP134A1) from Bacillus subtilis: A cyclo-l-leucyl-l-leucyl dipeptide oxidase. Biochemistry 2010, 49, 7282–7296. [Google Scholar] [CrossRef] [PubMed]
- Cryle, M.J.; Schlichting, I. Structural insights from a P450 carrier protein complex reveal how specificity is achieved in the P450BioI ACP complex. Proc. Natl. Acad. Sci. USA 2008, 105, 15696–15701. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Z.; Li, X.F.; Yang, W.; Dong, X.; Yu, J.; Zhu, S.L.; Li, M.; Xie, L.; Tong, W.Y. Identification and functional analysis of cytochrome P450 complement in Streptomyces virginiae IBL14. BMC Genom. 2013, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhang, T.; Hall, E.A.; Hutchinson, S.; Cryle, M.J.; Wong, L.L.; Zhou, W.; Bell, S.G. The crystal structure of the versatile cytochrome P450 enzyme CYP109B1 from Bacillus subtilis. Mol. Biosyst. 2015, 11, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Savino, C.; Montemiglio, L.C.; Sciara, G.; Miele, A.E.; Kendrew, S.G.; Jemth, P.; Gianni, S.; Vallone, B. Investigating the structural plasticity of a cytochrome P450: Three dimensional structures of P450 EryK and binding to its physiological substrate. J. Biol. Chem. 2009. [Google Scholar] [CrossRef] [PubMed]
- Montemiglio, L.C.; Gianni, S.; Vallone, B.; Savino, C. Azole drugs trap cytochrome P450 EryK in alternative conformational states. Biochemistry 2010, 49, 9199–9206. [Google Scholar] [CrossRef] [PubMed]
- Merson-Davies, L.A.; Cundiiffe, E. Analysis of five tyiosin biosynthetic genes from the tyllBA region of the Streptomyces fradiae genome. Mol. Microbiol. 1994, 13, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Fouces, R.; Mellado, E.; Díez, B.; Barredo, J.L. The tylosin biosynthetic cluster from Streptomyces fradiae: Genetic organization of the left region. Microbiology 1999, 145, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Cluster Number | Type of BGCs | P450 Name |
|---|---|---|---|
| Bacillus subtilis subsp. subtilis 168 | 4 | Nrps-Transatpks-Otherks | CYP134A1 |
| 10 | Other | CYP134A1 | |
| Bacillus subtilis subsp. subtilis RO-NN-1 | 3 | Nrps-Transatpks-Otherks | CYP107K1 |
| Bacillus subtilis subsp. subtilis BSP1 | 8 | Transatpks-Otherks-Nrps | CYP107K1 |
| Bacillus subtilis subsp. subtilis 6051-HGW | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 10 | Other | CYP107K1 | |
| Bacillus subtilis subsp. subtilis BAB-1 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| Bacillus subtilis subsp. subtilis AG1839 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 10 | Other | CYP107K1 | |
| Bacillus subtilis subsp. subtilis JH642 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 10 | Other | CYP134A1 | |
| Bacillus subtilis subsp. subtilis OH 131.1 | 1 | Lantipeptide | CYP152A1 |
| 4 | Nrps-Transatpks-Otherks | CYP107K1 | |
| 9 | Other | CYP134A1 | |
| Bacillus subtilis subsp. spizizenii W23 | 1 | Phosphonate | CYP152A1 |
| 4 | Nrps-Transatpks-Otherks | CYP107K1 | |
| 10 | Other | CYP134A1 | |
| Bacillus subtilis subsp. spizizenii TU-B-10 | 3 | Nrps-Transatpks-Otherks | CYP107K1 |
| 9 | Other | CYP134A1 | |
| Bacillus subtilis BSn5 | 4 | Other | CYP102A48 |
| 8 | Lantipeptide | CYP152A1 | |
| 11 | Nrps-Transatpks-Otherks | CYP107K1 | |
| Bacillus subtilis QB928 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 10 | Other | CYP134A1 | |
| Bacillus subtilis XF-1 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| Bacillus subtilis PY79 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 9 | Other | CYP134A1 | |
| Bacillus licheniformis ATCC 14580 | 7 | Other | CYP134A5 |
| Bacillus licheniformis DSM 13 = ATCC 14580 | 7 | Other | CYP134A5 |
| Bacillus paralicheniformis | 10 | Other | CYP134A5 |
| Bacillus velezensis FZB42 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| 9 | Transatpks | CYP113L1 | |
| Bacillus velezensis CAU B946 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| 9 | Transatpks | CYP113L1 | |
| Bacillus velezensis YAU B9601-Y2 | 5 | Transatpks | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107K3 | |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus velezensis AS43.3 | 6 | Transatpks | CYP107K3 |
| 7 | Transatpks-Nrps | CYP113L1 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus velezensis UCMB5036 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| 9 | Bacteriocin-Nrps | CYP113L1 | |
| Bacillus velezensis UCMB5033 | 6 | Transatpks-Nrps | CYP107K3 |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus velezensis UCMB5113 | 7 | Transatpks-Nrps | CYP107K3 |
| 8 | Transatpks-Nrps | CYP107H4 | |
| 11 | Transatpks | CYP113L1 | |
| Bacillus velezensis NAU-B3 | 3 | Transatpks | CYP113L1 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| 7 | Transatpks-Nrps | CYP107K3 | |
| Bacillus velezensis TrigoCor1448 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| Bacillus velezensis SQR9 | 6 | Transatpks-Nrps | CYP107K3 |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus velezensis | 6 | Transatpks-Nrps | CYP107K3 |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus amyloliquefaciens DSM 7 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H2 | |
| Bacillus amyloliquefaciens TA208 | 7 | Transatpks-Nrps | CYP107H2 |
| 8 | Transatpks-Nrps | CYP107K3 | |
| Bacillus amyloliquefaciens LL3 | 5 | Transatpks-Nrps | CYP107K3 |
| 6 | Transatpks-Nrps | CYP107H2 | |
| Bacillus amyloliquefaciens XH7 | 7 | Transatpks-Nrps | CYP107H2 |
| 8 | Transatpks-Nrps | CYP107K3 | |
| Bacillus amyloliquefaciens Y2 | 6 | Transatpks-Nrps | CYP107K3 |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus amyloliquefaciens IT-45 | 3 | Transatpks | CYP113L1 |
| 6 | Transatpks-Nrps | CYP107H4 | |
| 7 | Transatpks-Nrps | CYP107K3 | |
| Bacillus amyloliquefaciens CC178 | 6 | Transatpks-Nrps | CYP107H4 |
| 9 | Transatpks | CYP113L1 | |
| Bacillus amyloliquefaciens LFB112 | 7 | Transatpks-Nrps | CYP107K3 |
| 8 | Transatpks-Nrps | CYP107H4 | |
| 11 | Transatpks | CYP113L1 | |
| Bacillus atrophaeus 1942 | 3 | Nrps-Transatpks-Otherks | CYP107K2 |
| 10 | Nrps | CYP152A9 | |
| Bacillus atrophaeus NRS 1221A | 3 | Nrps-Transatpks-Otherks | CYP107K2 |
| 10 | Nrps | CYP152A9 | |
| Bacillus vallismortis | 6 | Transatpks-Nrps | CYP107K3 |
| 7 | Transatpks-Nrps | CYP107H4 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus pumilus SH-B9 | 8 | Nrps | CYP109B6 |
| Bacillus sp. JS | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| Bacillus sp. Pc3 | 1 | Bacteriocin-Transatpks-Nrps | CYP107H4 |
| 2 | Transatpks-Nrps | CYP107K3 | |
| 10 | Transatpks | CYP113L1 | |
| Bacillus sp. BH072 | 8 | Transatpks-Nrps | CYP107K3 |
| 9 | Transatpks-Nrps | CYP107H4 | |
| 12 | Transatpks | CYP107H4 | |
| Bacillus sp. YP1 | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| Bacillus sp. BS34A | 4 | Nrps-Transatpks-Otherks | CYP107K1 |
| 10 | Other | CYP134A1 | |
| Bacillus sp. LM 4-2 | 3 | Nrps-Transatpks-Otherks | CYP107K1 |
| 7 | Other | CYP102A48 | |
| 9 | Other | CYP134A1 | |
| Bacillus gibsonii | 1 | Nrps-Transatpks-Otherks | CYP107K1 |
| 6 | Other | CYP134A1 | |
| 9 | Lantipeptide | CYP152A1 | |
| Bacillus xiamenensis | 2 | Nrps | CYP1179A4 |
| Bacillus altitudinis | 2 | Nrps | CYP1179A4 |
| 8 | Nrps | CYP109B5 | |
| Bacillus sp. SDLI1 | 3 | Transatpks-Nrps | CYP107H4 |
| 4 | Transatpks-Nrps | CYP107K3 | |
| 11 | Transatpks | CYP113L1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mthethwa, B.C.; Chen, W.; Ngwenya, M.L.; Kappo, A.P.; Syed, P.R.; Karpoormath, R.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comparative Analyses of Cytochrome P450s and Those Associated with Secondary Metabolism in Bacillus Species. Int. J. Mol. Sci. 2018, 19, 3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113623
Mthethwa BC, Chen W, Ngwenya ML, Kappo AP, Syed PR, Karpoormath R, Yu J-H, Nelson DR, Syed K. Comparative Analyses of Cytochrome P450s and Those Associated with Secondary Metabolism in Bacillus Species. International Journal of Molecular Sciences. 2018; 19(11):3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113623
Chicago/Turabian StyleMthethwa, Bongumusa Comfort, Wanping Chen, Mathula Lancelot Ngwenya, Abidemi Paul Kappo, Puleng Rosinah Syed, Rajshekhar Karpoormath, Jae-Hyuk Yu, David R. Nelson, and Khajamohiddin Syed. 2018. "Comparative Analyses of Cytochrome P450s and Those Associated with Secondary Metabolism in Bacillus Species" International Journal of Molecular Sciences 19, no. 11: 3623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113623