Novel Antioxidant Properties of Doxycycline

 , , and
, , and 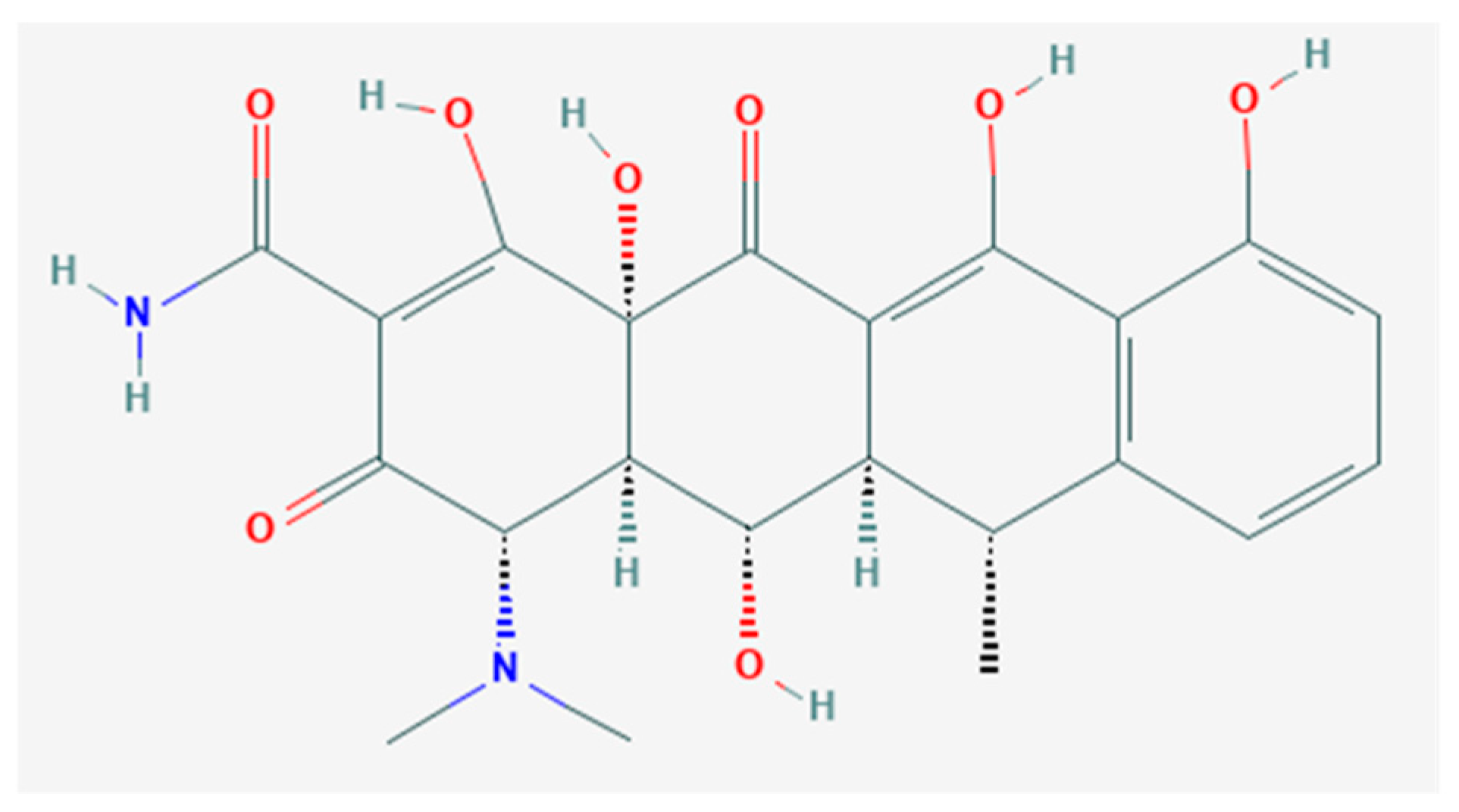{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
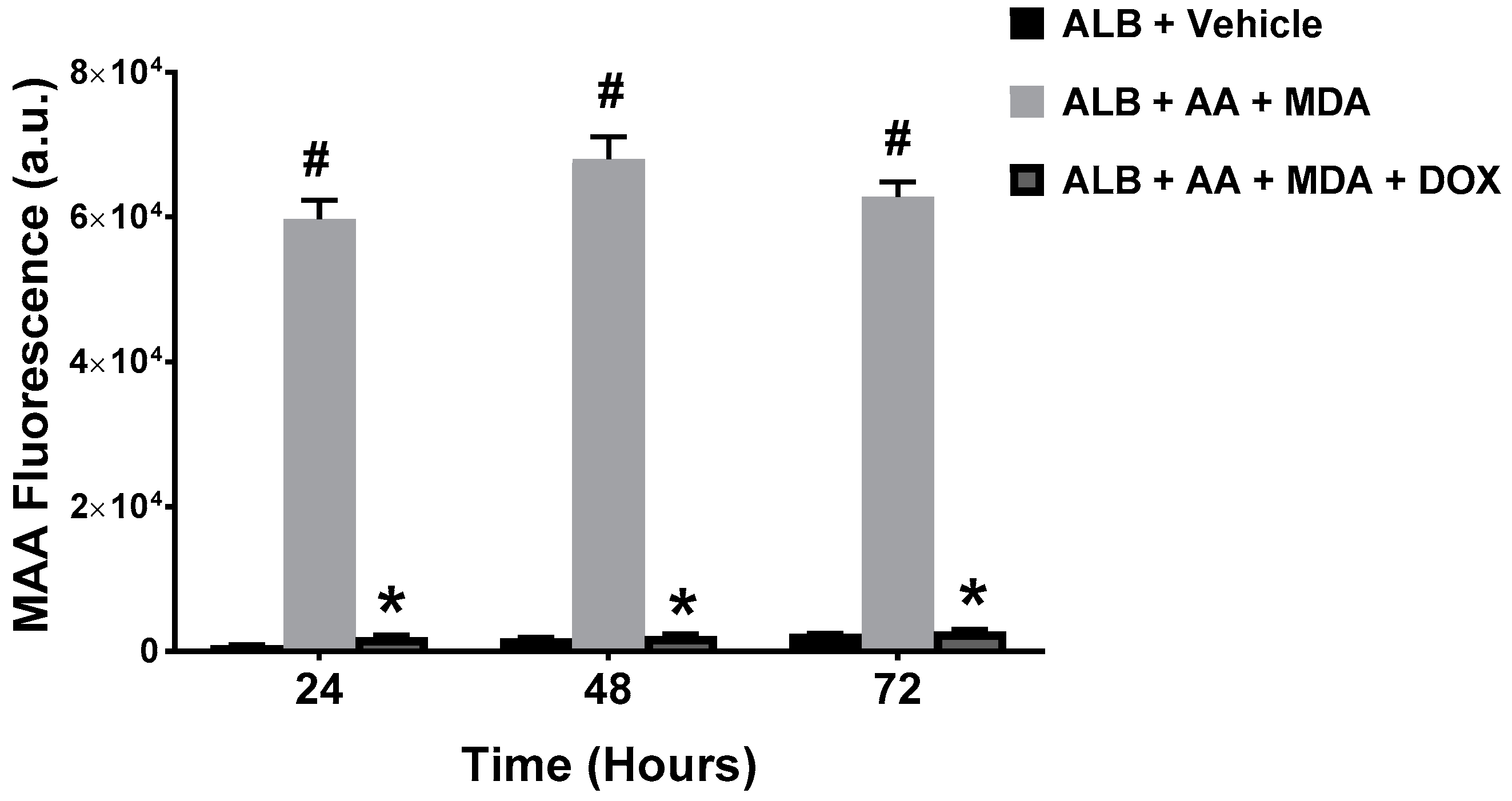
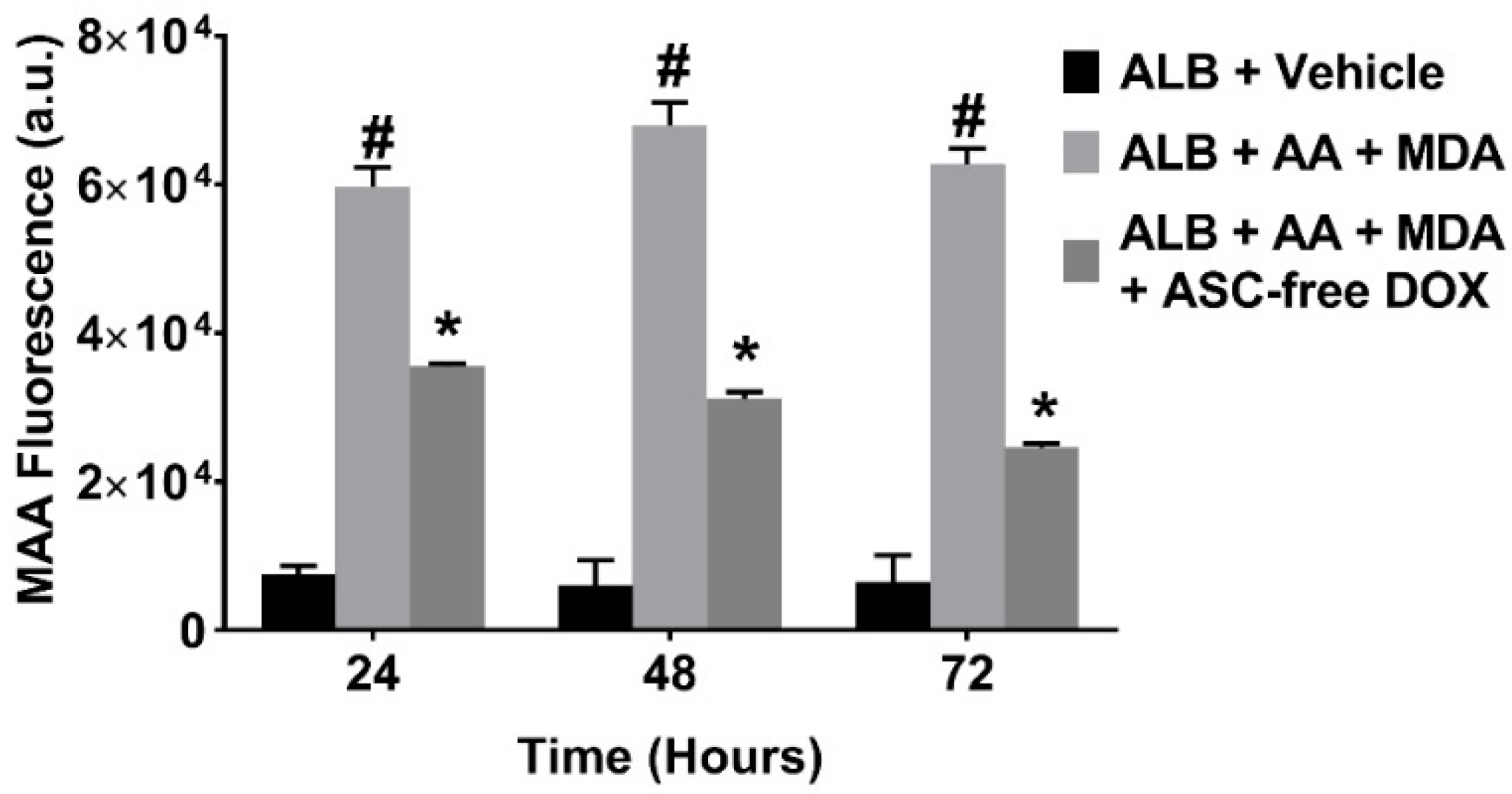
2.1. Doxycycline Attenuates MAA-Protein Adduct Formation
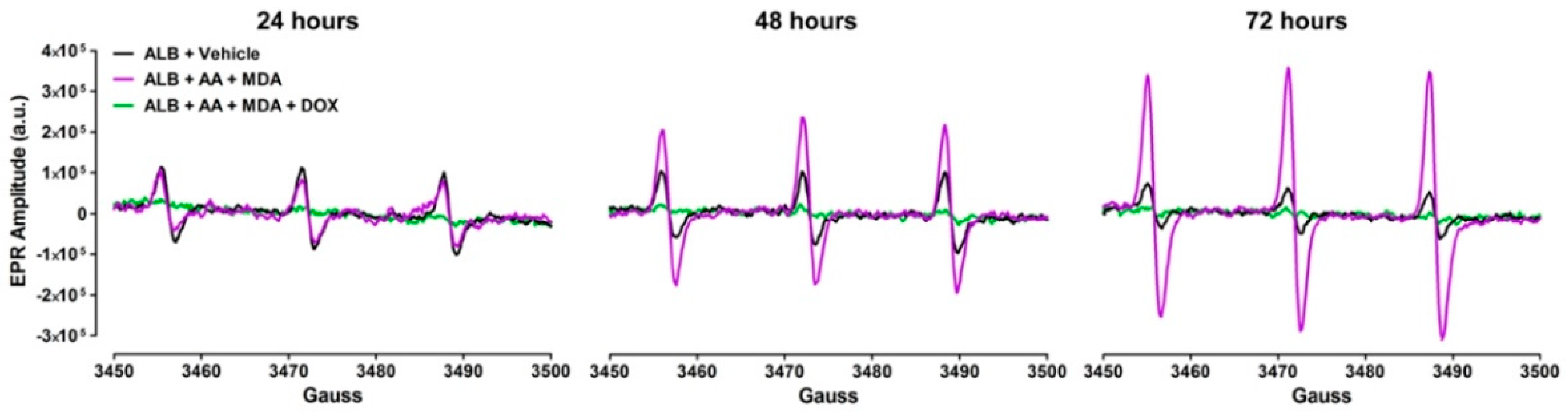
2.2. Doxycycline Decreases ROS Produced during the Generation of MAA-Adducts
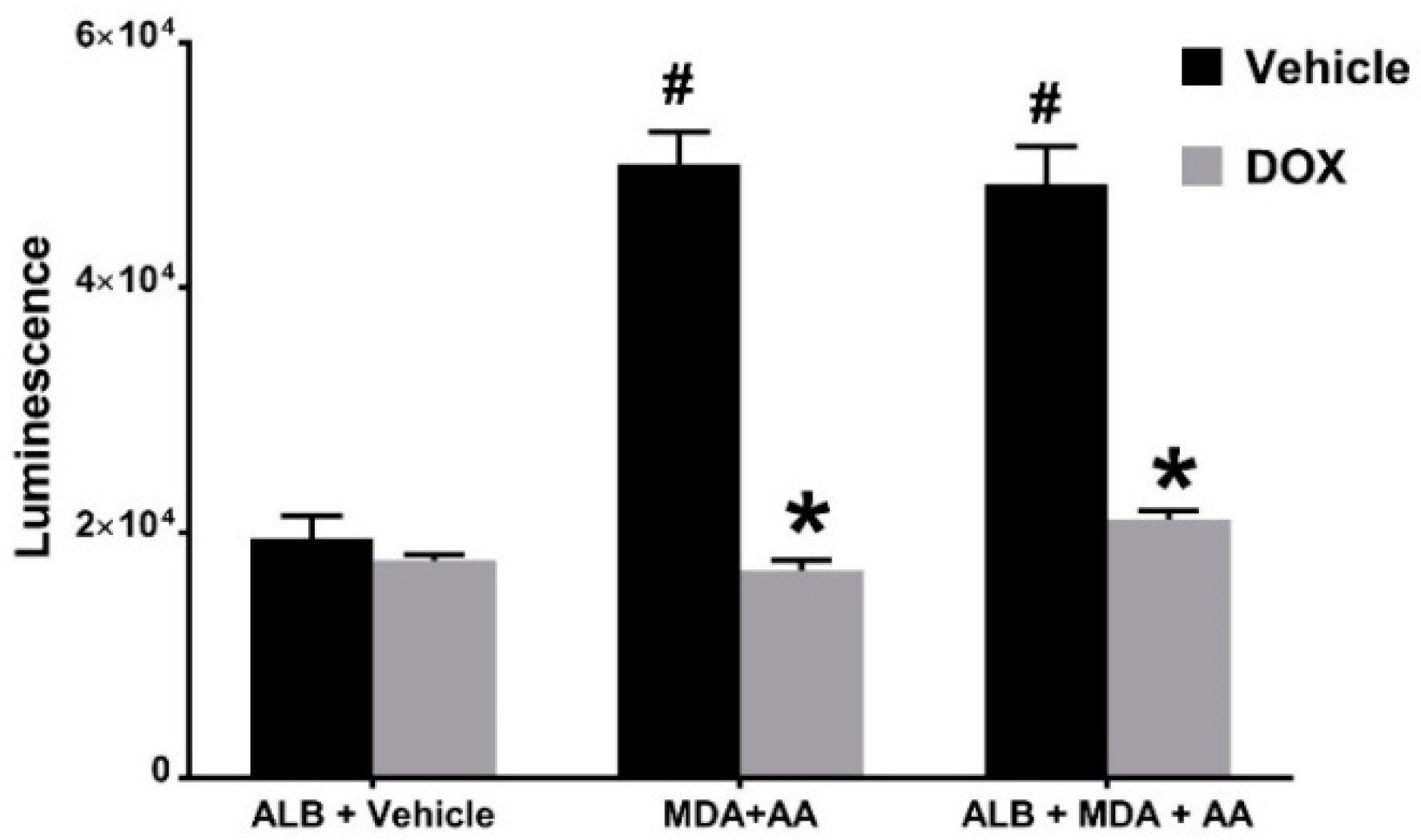
2.3. Doxycycline Inhibits MAA-ALB-Induced Activation of Nrf2.
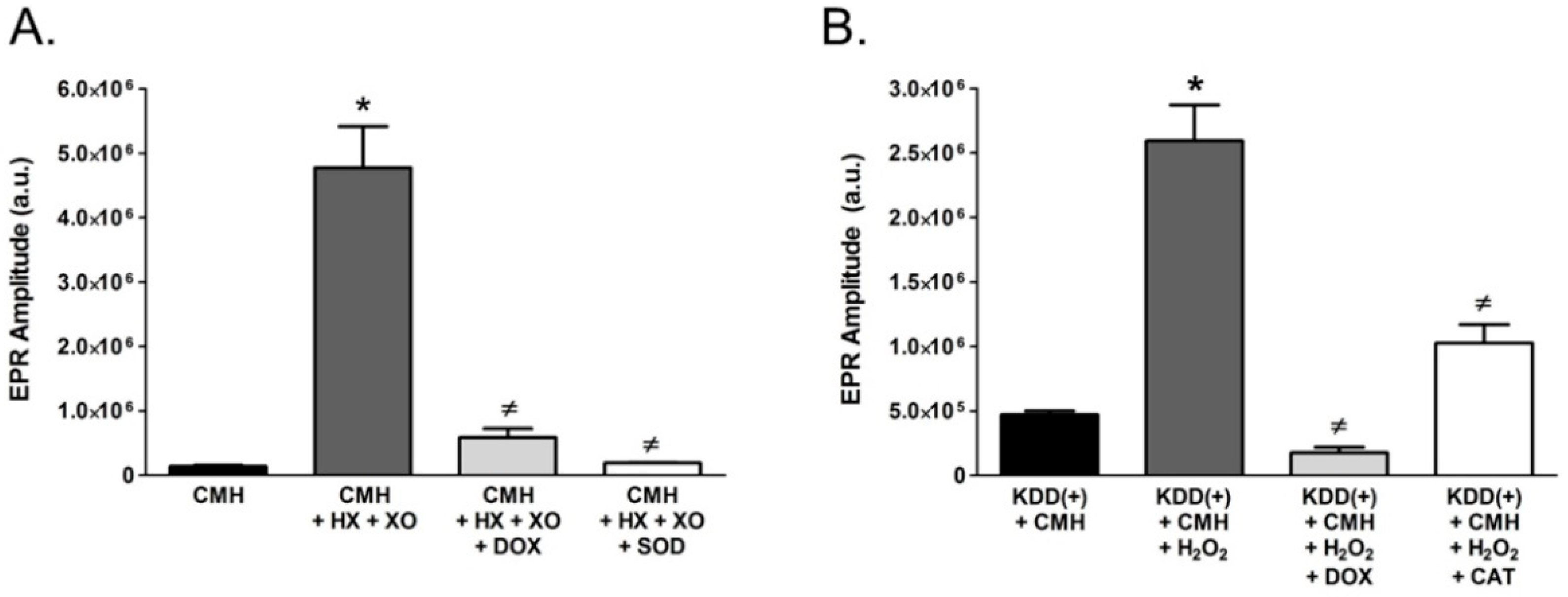
2.4. Pharmaceutical Doxycycline Directly Scavenges Superoxide and Hydrogen Peroxide
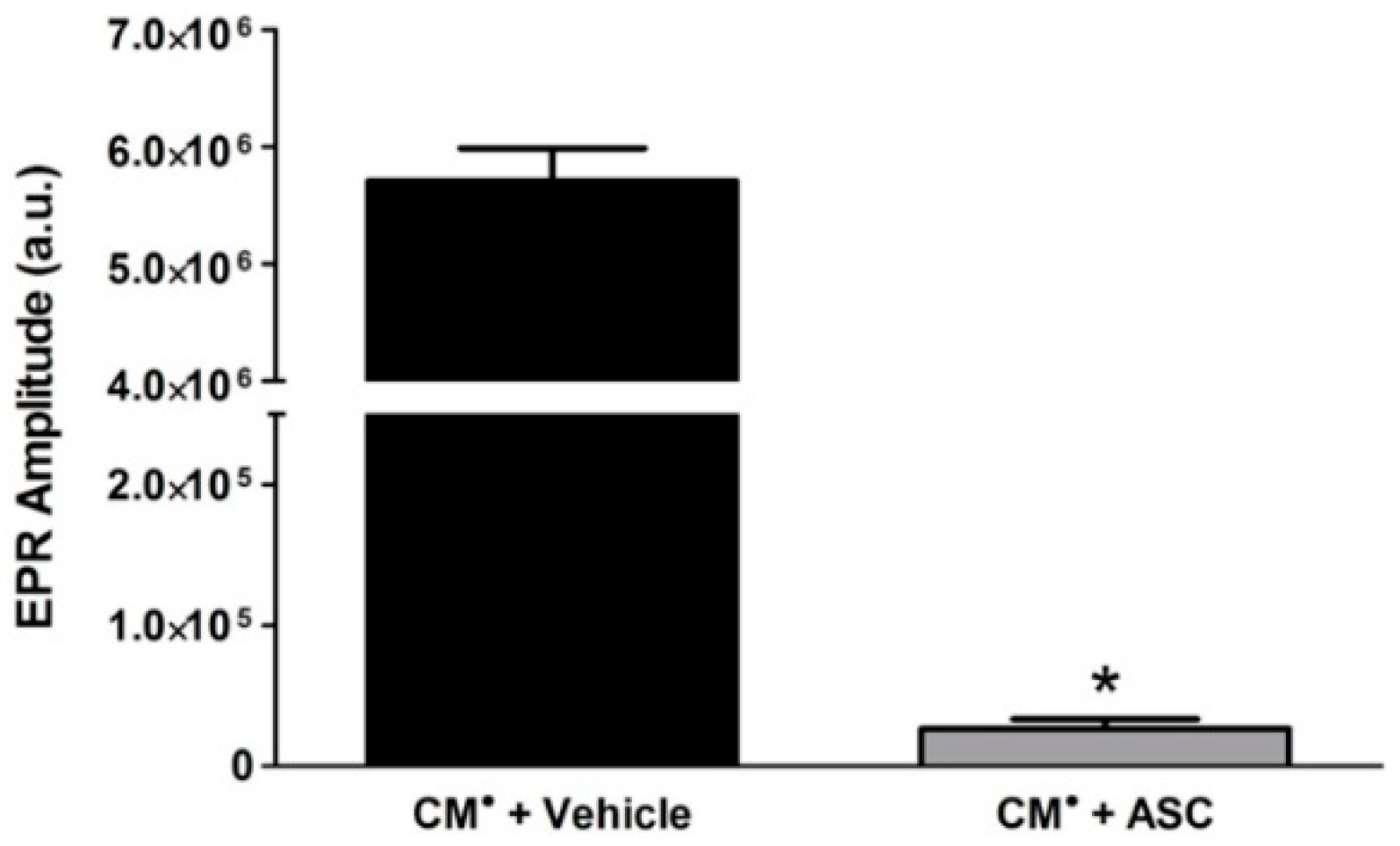
2.5. Ascorbic Acid (ASC) in Pharmaceutical DOX Reduces the CM Radical Spin Probe to Non-Radical CMH
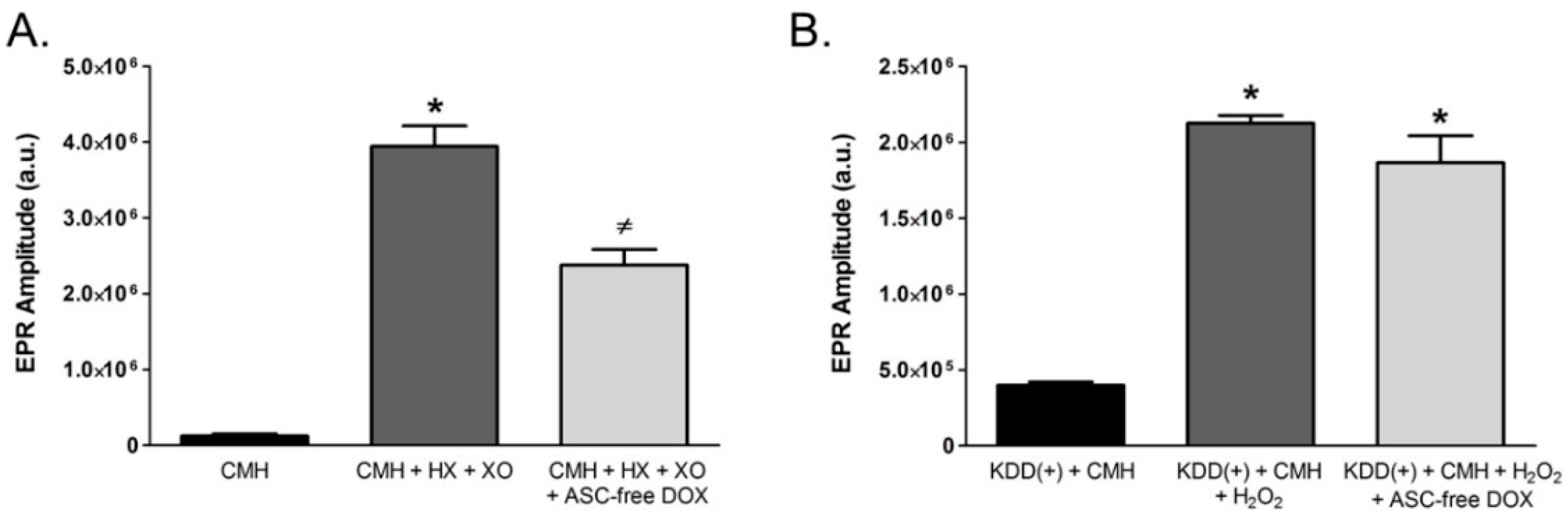
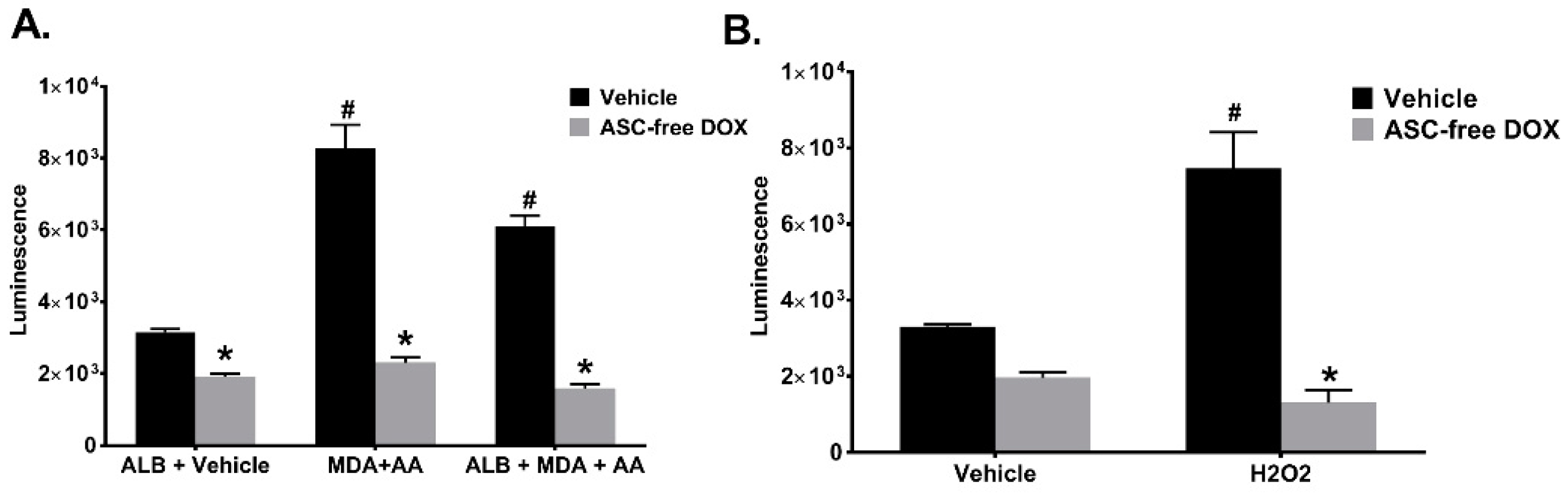
2.6. Doxycycline in the Absence of Ascorbic Acid Scavenges Superoxide and Inhibits Intracellular Redox Signaling
3. Discussion
4. Materials and Methods
4.1. Malondialdehyde-Acetaldehyde (MAA)-Protein Adduct Formation
4.2. Electron Paramagnetic Resonance (EPR) Spectroscopy
4.3. Cellular Redox Signaling
4.4. Statistical Analysis
5. Clinical Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | acetaldehyde |
| ALB | human serum albumin |
| ARE | antioxidant response element |
| ASC | ascorbic acid |
| CAT | catalase |
| CM• | stable nitroxide radical |
| CMH | 1-hydroxy-3-methoxycarbonyl-2,2,5,5-tetramethylpyrrolidine |
| DETC | diethyldithiocarbamate |
| DTPA | diethylenetriaminepentaacetic acid |
| DOX | doxycycline |
| EPR | electron paramagnetic resonance |
| H2O2 | hydrogen peroxide |
| HX | hypoxanthine |
| ROS | reactive oxygen species |
| MDA | malondialdehyde |
| MAA | malondialdehyde-acetaldehyde |
| MTX | methotrexate |
| Nrf2 | nuclear factor (erythroid derived 2)-like 2 |
| O2•− | superoxide |
| SOD | superoxide dismutase |
| XO | xanthine oxidase |
References
- Griffin, M.O.; Fricovsky, E.; Ceballos, G.; Villarreal, F. Tetracyclines: A pleitropic family of compounds with promising therapeutic properties. Review of the literature. Am. J. Physiol. Cell Physiol. 2010, 299, C539–C548. [Google Scholar] [CrossRef] [PubMed]
- Metz, L.M.; Li, D.K.B.; Traboulsee, A.L.; Duquette, P.; Eliasziw, M.; Cerchiaro, G.; Greenfield, J.; Riddehough, A.; Yeung, M.; Kremenchutzky, M.; et al. Trial of Minocycline in a Clinically Isolated Syndrome of Multiple Sclerosis. N. Engl. J. Med. 2017, 376, 2122–2133. [Google Scholar] [CrossRef]
- O’Dell, J.R.; Elliott, J.R.; Mallek, J.A.; Mikuls, T.R.; Weaver, C.A.; Glickstein, S.; Blakely, K.M.; Hausch, R.; Leff, R.D. Treatment of early seropositive rheumatoid arthritis: Doxycycline plus methotrexate versus methotrexate alone. Arthritis Rheum. 2006, 54, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stechmiller, J.; Cowan, L.; Schultz, G. The role of doxycycline as a matrix metalloproteinase inhibitor for the treatment of chronic wounds. Biol. Res. Nurs. 2010, 11, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.W.; Baxter, B.T. MMP inhibition in abdominal aortic aneurysms. Rationale for a prospective randomized clinical trial. Ann. N. Y. Acad. Sci. 1999, 878, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Baxter, B.T.; Pearce, W.H.; Waltke, E.A.; Littooy, F.N.; Hallett, J.W.; Kent, K.C., Jr.; Upchurch, G.R., Jr.; Chaikof, E.L.; Mills, J.L.; Fleckten, B.; et al. Prolonged administration of doxycycline in patients with small asymptomatic abdominal aortic aneurysms: Report of a prospective (Phase II) multicenter study. J. Vasc. Surg. 2002, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiong, W.; Baca-Regen, L.; Nagase, H.; Baxter, B.T. Mechanism of inhibition of matrix metalloproteinase-2 expression by doxycycline in human aortic smooth muscle cells. J. Vasc. Surg. 2003, 38, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Ravandi, A.; Boekholdt, S.M.; Mallat, Z.; Talmud, P.J.; Kastelein, J.J.; Wareham, N.J.; Miller, E.R.; Benessiano, J.; Tedgui, A.; Witztum, J.L.; et al. Relationship of IgG and IgM autoantibodies and immune complexes to oxidized LDL with markers of oxidation and inflammation and cardiovascular events: Results from the EPIC-Norfolk Study. J. Lipid Res. 2011, 52, 1829–1836. [Google Scholar] [CrossRef]
- Tsimikas, S.; Brilakis, E.S.; Lennon, R.J.; Miller, E.R.; Witztum, J.L.; McConnell, J.P.; Kornman, K.S.; Berger, P.B. Relationship of IgG and IgM autoantibodies to oxidized low density lipoprotein with coronary artery disease and cardiovascular events. J. Lipid Res. 2007, 48, 425–433. [Google Scholar] [CrossRef]
- Kikugawa, K.; Ido, Y. Studies on peroxidized lipids. V. Formation and characterization of 1,4-dihydropyridine-3,5-dicarbaldehydes as model of fluorescent components in lipofuscin. Lipids 1984, 19, 600–608. [Google Scholar] [CrossRef]
- Tuma, D.J.; Newman, M.R.; Donohue, T.M.; Sorrell, M.F., Jr. Covalent binding of acetaldehyde to proteins: Participation of lysine residues. Alcohol. Clin. Exp. Res. 1987, 11, 579–584. [Google Scholar] [CrossRef]
- Binder, C.J.; Papac-Milicevic, N.; Witztum, J.L. Innate sensing of oxidation-specific epitopes in health and disease. Nat. Rev. Immunol. 2016, 16, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.J.; Binder, C.J. Malondialdehyde epitopes as mediators of sterile inflammation. Biochim. Biophys. Acta 2017, 1862, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Tuma, D.J.; Thiele, G.M.; Xu, D.; Klassen, L.W.; Sorrell, M.F. Acetaldehyde and malondialdehyde react together to generate distinct protein adducts in the liver during long-term ethanol administration. Hepatology 1996, 23, 872–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.R.; Duryee, M.J.; Shurmur, S.W.; Um, J.Y.; Bussey, W.D.; Hunter, C.D.; et al. Unique antibody responses to malondialdehyde-acetaldehyde (MAA)-protein adducts predict coronary artery disease. PLoS ONE 2014, 9, e107440. [Google Scholar] [CrossRef] [PubMed]
- Duryee, M.J.; Klassen, L.W.; Schaffert, C.S.; Tuma, D.J.; Hunter, C.D.; Garvin, R.P.; Anderson, D.R.; Thiele, G.M. Malondialdehyde-acetaldehyde adduct is the dominant epitope after MDA modification of proteins in atherosclerosis. Free Radical Biol. Med. 2010, 49, 1480–1486. [Google Scholar] [CrossRef]
- Willis, M.S.; Thiele, G.M.; Tuma, D.J.; Klassen, L.W. T cell proliferative responses to malondialdehyde-acetaldehyde haptenated protein are scavenger receptor mediated. Int. Immunopharmacol. 2003, 3, 1381–1399. [Google Scholar] [CrossRef]
- Emeto, T.I.; Moxon, J.V.; Au, M.; Golledge, J. Oxidative stress and abdominal aortic aneurysm: Potential treatment targets. Clin. Sci. 2016, 130, 301–315. [Google Scholar] [CrossRef]
- Rosenbaugh, E.G.; Roat, J.W.; Gao, L.; Yang, R.F.; Manickam, D.S.; Yin, J.X.; Schultz, H.D.; Bronich, T.K.; Batrakova, E.V.; Kabanov, A.V.; et al. The attenuation of central angiotensin II-dependent pressor response and intra-neuronal signaling by intracarotid injection of nanoformulated copper/zinc superoxide dismutase. Biomaterials 2010, 31, 5218–5226. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M.C.; Clemens, D.L.; Duryee, M.J.; Sarmiento, C.; Chiou, A.; Hunter, C.D.; Tian, J.; Klassen, L.W.; O’Dell, J.R.; Thiele, G.M.; et al. Direct antioxidant properties of methotrexate: Inhibition of malondialdehyde-acetaldehyde-protein adduct formation and superoxide scavenging. Redox Biol. 2017, 13, 588–593. [Google Scholar] [CrossRef]
- Antoniak, D.T.; Duryee, M.J.; Mikuls, T.R.; Thiele, G.M.; Anderson, D.R. Aldehyde-modified proteins as mediators of early inflammation in atherosclerotic disease. Free Radical Biol. Med. 2015, 89, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.I.; Dikalova, A.E.; Bikineyeva, A.T.; Schmidt, H.H.; Harrison, D.G.; Griendling, K.K. Distinct roles of Nox1 and Nox4 in basal and angiotensin II-stimulated superoxide and hydrogen peroxide production. Free Radical Biol. Med. 2008, 45, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Monacelli, F.; Acquarone, E.; Giannotti, C.; Borghi, R.; Nencioni, A.; Vitamin, C. Aging and Alzheimer’s Disease. Nutrients 2017, 9. [Google Scholar]
- Kunsch, C.; Medford, R.M. Oxidative stress as a regulator of gene expression in the vasculature. Circ. Res. 1999, 85, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Spychalowicz, A.; Wilk, G.; Sliwa, T.; Ludew, D.; Guzik, T.J. Novel therapeutic approaches in limiting oxidative stress and inflammation. Curr. Pharm. Biotechnol. 2012, 13, 2456–2466. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.J.; Hendrikx, T.; Weismann, D.; Jackel, S.; Walenbergh, S.M.; Rendeiro, A.F.; Weißer, J.; Puhm, F.; Hladik, A.; Göderle, L.; et al. Malondialdehyde epitopes are sterile mediators of hepatic inflammation in hypercholesterolemic mice. Hepatology 2017, 65, 1181–1195. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Freeman, T.L.; Haver, A.; Duryee, M.J.; Tuma, D.J.; Klassen, L.W.; Hamel, F.G.; White, R.L.; Rennard, S.I.; Thiele, G.M. Aldehydes in cigarette smoke react with the lipid peroxidation product malonaldehyde to form fluorescent protein adducts on lysines. Chem. Res. Toxicol. 2005, 18, 817–824. [Google Scholar] [CrossRef]
- Hill, G.E.; Miller, J.A.; Baxter, B.T.; Klassen, L.W.; Duryee, M.J.; Tuma, D.J.; Thiele, G.M. Association of malondialdehyde-acetaldehyde (MAA) adducted proteins with atherosclerotic-induced vascular inflammatory injury. Atherosclerosis 1998, 141, 107–116. [Google Scholar] [CrossRef]
- McCaskill, M.L.; Kharbanda, K.K.; Tuma, D.J.; Reynolds, J.D.; DeVasure, J.M.; Sisson, J.H.; Wyatt, T.A. Hybrid malondialdehyde and acetaldehyde protein adducts form in the lungs of mice exposed to alcohol and cigarette smoke. Alcohol. Clin. Exp. Res. 2011, 35, 1106–1113. [Google Scholar] [CrossRef]
- Rolla, R.; Vay, D.; Mottaran, E.; Parodi, M.; Traverso, N.; Arico, S.; Sartori, M.; Bellomo, G.; Klassen, L.W.; Thiele, G.M.; et al. Detection of circulating antibodies against malondialdehyde-acetaldehyde adducts in patients with alcohol-induced liver disease. Hepatology 2000, 31, 878–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiele, G.M.; Duryee, M.J.; Anderson, D.R.; Klassen, L.W.; Mohring, S.M.; Young, K.A.; Benissan-Messan, D.; Sayles, H.; Dusad, A.; Hunter, C.D.; et al. Malondialdehyde-acetaldehyde adducts and anti-malondialdehyde-acetaldehyde antibodies in rheumatoid arthritis. Arthritis Rheumatol. 2015, 67, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.S.; Xiong, W.; Dale, M.; Yu, F.; Duryee, M.J.; Anderson, D.R.; Thiele, G.M.; Baxter, B.T. Antibodies against malondialdehyde-acetaldehyde adducts can help identify patients with abdominal aortic aneurysm. J. Vasc. Surg. 2016, 63, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Thiele, G.M.; Beckenhauer, J.L.; Klassen, L.W.; Sorrell, M.F.; Tuma, D.J. Detection of circulating antibodies to malondialdehyde-acetaldehyde adducts in ethanol-feed rats. Gastroenterology 1998, 115, 686–692. [Google Scholar] [CrossRef]
- Willis, M.S.; Klassen, L.W.; Tuma, D.J.; Sorrell, M.F.; Thiele, G.M. Adduction of soluble proteins with malondialdehyde-acetaldehyde (MAA) induces antibody production and enhances T.-cell proliferation. Alcohol. Clin. Exp. Res. 2002, 26, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Kraus, R.L.; Pasieczny, R.; Lariosa-Willingham, K.; Turner, M.S.; Jiang, A.; Trauger, J.W. Antioxidant properties of minocycline: Neuroprotection in an oxidative stress assay and direct radical-scavenging activity. J. Neurochem. 2005, 94, 819–827. [Google Scholar] [CrossRef]
- Budni, J.; Garcez, M.L.; de Medeiros, J.; Cassaro, E.; Bellettini-Santos, T.; Mina, F.; Quevedo, J. The Anti-Inflammatory Role of Minocycline in Alzheimer’s Disease. Curr. Alzheimer Res. 2016, 13, 1319–1329. [Google Scholar] [CrossRef]
- Soory, M. A role for non-antimicrobial actions of tetracyclines in combating oxidative stress in periodontal and metabolic diseases: A literature review. Open Dent. J. 2008, 2, 5–12. [Google Scholar] [CrossRef]
- Yagan, A.; Kesim, S.; Liman, N. Effect of low-dose doxycycline on serum oxidative status, gingival antioxidant levels, and alveolar bone loss in experimental periodontitis in rats. J. Periodontol. 2014, 85, 478–489. [Google Scholar] [CrossRef]
- Ozcinar, E.; Okatan, E.N.; Tuncay, E.; Eryilmaz, S.; Turan, B. Improvement of functional recovery of donor heart following cold static storage with doxycycline cardioplegia. Cardiovasc Toxicol. 2014, 14, 64–73. [Google Scholar] [CrossRef]
- Antonio, R.C.; Ceron, C.S.; Rizzi, E.; Coelho, E.B.; Tanus-Santos, J.E.; Gerlach, R.F. Antioxidant effect of doxycycline decreases MMP activity and blood pressure in SHR. Mol. Cell. Biochem. 2014, 386, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vascul. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Dikalov, S.; Griendling, K.K.; Harrison, D.G. Measurement of reactive oxygen species in cardiovascular studies. Hypertension 2007, 49, 717–727. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clemens, D.L.; Duryee, M.J.; Sarmiento, C.; Chiou, A.; McGowan, J.D.; Hunter, C.D.; Schlichte, S.L.; Tian, J.; Klassen, L.W.; O’Dell, J.R.; et al. Novel Antioxidant Properties of Doxycycline. Int. J. Mol. Sci. 2018, 19, 4078. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124078
Clemens DL, Duryee MJ, Sarmiento C, Chiou A, McGowan JD, Hunter CD, Schlichte SL, Tian J, Klassen LW, O’Dell JR, et al. Novel Antioxidant Properties of Doxycycline. International Journal of Molecular Sciences. 2018; 19(12):4078. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124078
Chicago/Turabian StyleClemens, Dahn L., Michael J. Duryee, Cleofes Sarmiento, Andrew Chiou, Jacob D. McGowan, Carlos D. Hunter, Sarah L. Schlichte, Jun Tian, Lynell W. Klassen, James R. O’Dell, and et al. 2018. "Novel Antioxidant Properties of Doxycycline" International Journal of Molecular Sciences 19, no. 12: 4078. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124078