Abnormal Expression of c-Myc Oncogene in NK Cells in Patients with Cancer

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
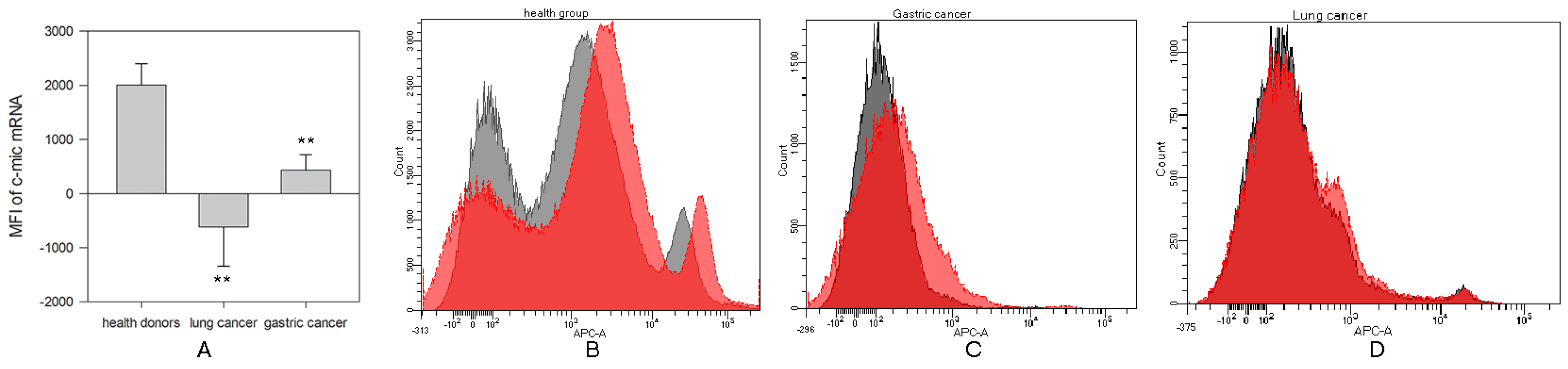
2.1. Reduced c-myc mRNA Expression in NK Cells in Cancer Patients
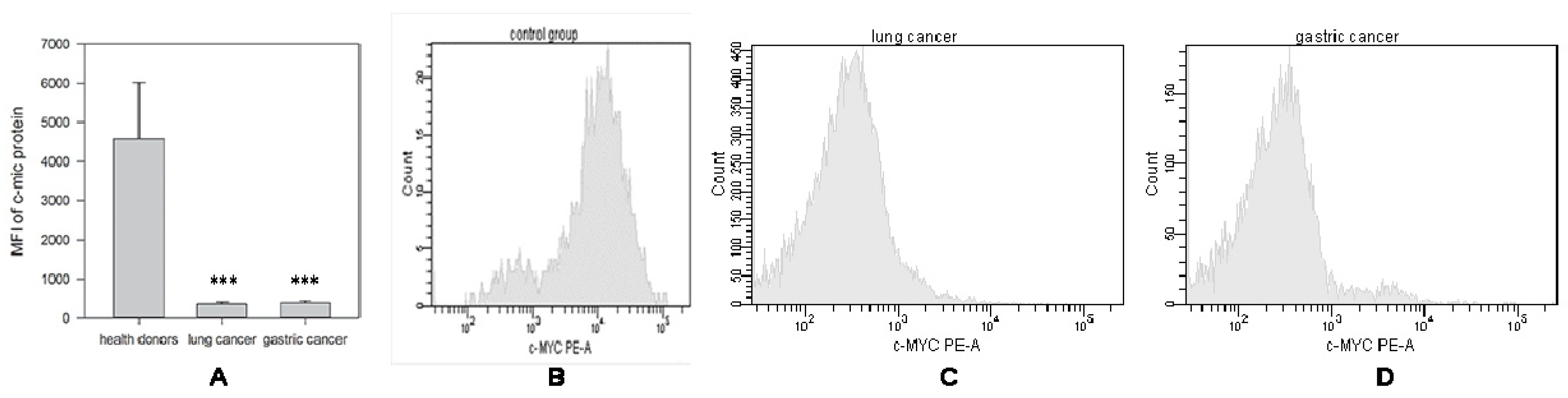
2.1.1. Decrease in c-Myc Protein Production in NK Cells from Cancer Patients
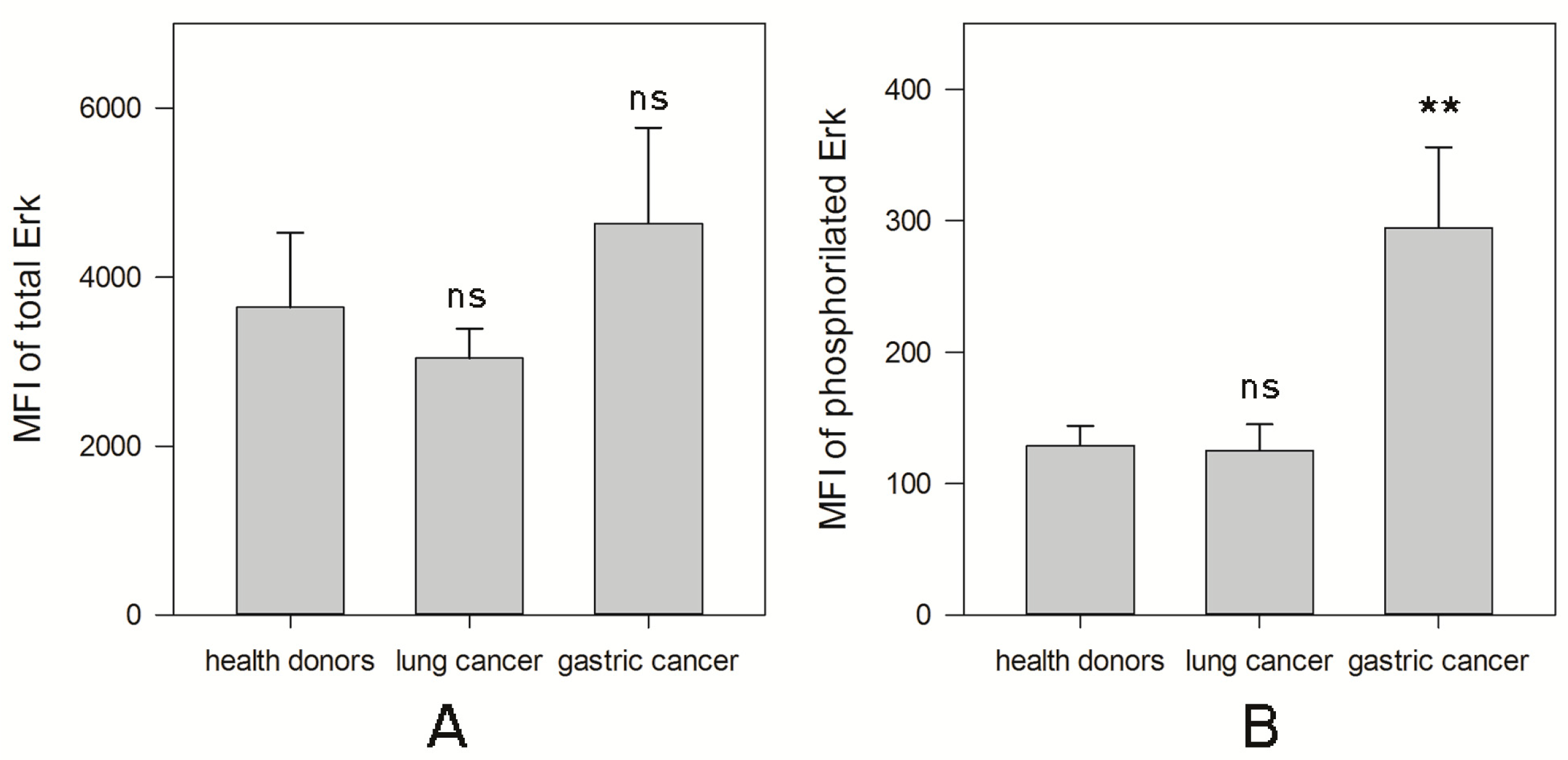
2.1.2. MAPK Activity in NK Cells from Cancer Patients
2.1.3. Defect of Cell Cycle in NK Cells from Cancer Patients
3. Discussion
4. Materials and Methods
4.1. Patients and Samples
4.2. Purification of NK Cells
4.3. SmartFlare™ RNA Detection Assay
4.4. Detection of c-Myc Protein Production
4.5. MAPK Activation Dual Detection Assay
4.6. Cell Cycle Assay
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, S.; Pineda, C.M.; Xin, T.; Boucher, J.; Suozzi, K.C.; Park, S.; Matte-Martone, C.; Gonzalez, D.G.; Rytlewski, J.; Beronja, S.; et al. Correction of aberrant growth preserves tissue homeostasis. Nature 2017, 548, 334–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavin, Y.; Kobayashi, S.; Leader, A.; Amir, E.D.; Elefant, N.; Bigenwald, C.; Remark, R.; Sweeney, R.; Becker, C.D.; Levine, J.H.; et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired Single-Cell Analyses. Cell 2017, 169, 750–765. [Google Scholar] [CrossRef] [PubMed]
- Degnim, A.C.; Hoskin, T.L.; Arshad, M.; Frost, M.H.; Winham, S.J.; Brahmbhatt, R.A.; Pena, A.; Carter, J.M.; Stallings-Mann, M.L.; Murphy, L.M.; et al. Alterations in the Immune Cell Composition in Premalignant Breast Tissue that Precede Breast Cancer Development. Clin. Cancer Res. 2017, 23, 3945–3952. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.E.; Hingorani, S.R.; Mick, R.; Combs, C.; Tuveson, D.A.; Vonderheide, R.H. Dynamics of the immune reaction to pancreatic cancer from inception to invasion. Cancer Res. 2007, 67, 9518–9527. [Google Scholar] [CrossRef] [PubMed]
- Zakiryanova, G.K.; Wheeler, S.; Shurin, M.R. Oncogenes in immune cells as potential therapeutic targets. Immunotargets Ther. 2018, 7, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Felsher, D.W. MYC Inactivation Elicits Oncogene Addiction through Both Tumor Cell-Intrinsic and Host-Dependent Mechanisms. Genes Cancer 2010, 1, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakiryanova, G.K.; Kustova, E.; Urazalieva, N.T.; Amirbekov, A.; Baimuchametov, E.T.; Nakisbekov, N.N.; Shurin, M.R. Alterations of oncogenes expression in, N.K. cells in patients with cancer. Immun. Inflamm. Dis. 2017, 5, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Levy, E.M.; Roberti, M.P.; Mordoh, J. Natural killer cells in human cancer: From biological functions to clinical applications. J. Biomed. Biotechnol. 2011, 676198. [Google Scholar] [CrossRef] [PubMed]
- Marcenaro, E.; Notarangelo, L.D.; Orange, J.S.; Vivier, E. Editorial: NK Cell Subsets in Health and Disease: New Developments. Front. Immunol. 2017, 8, 1363. [Google Scholar] [CrossRef] [PubMed]
- Messaoudene, M.; Frazao, A.; Gavlovsky, P.J.; Toubert, A.; Dulphy, N.; Caignard, A. Patient’s Natural Killer Cells in the Era of Targeted Therapies: Role for Tumor Killers. Front. Immunol. 2017, 8, 683. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, B.N.; Vivier, E. Natural killer cell tolerance: Control by self or self-control? Cold Spring Harb. Perspect. Biol. 2012, 4, 22383753. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Ugolini, S.; Blaise, D.; Chabannon, C.; Brossay, L. Targeting natural killer cells and natural killer T cells in cancer. Nat. Rev. Immunol. 2012, 12, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, W. Infusions of allogeneic natural killer cells as cancer therapy. Clin. Cancer Res. 2014, 20, 3390–3400. [Google Scholar] [CrossRef] [PubMed]
- Bachanova, V.; Miller, J.S. NK cells in therapy of cancer. Crit. Rev. Oncog. 2014, 19, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Oberschmidt, O.; Kloess, S.; Koehl, U. Redirected Primary Human Chimeric Antigen Receptor Natural Killer Cells As an “Off-the-Shelf Immunotherapy” for Improvement in Cancer Treatment. Front. Immunol. 2017, 8, 654. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.; Hodgins, J.J.; Marathe, M.; Nicolai, C.J.; Bourgeois-Daigneault, M.C.; Trevino, T.N.; Azimi, C.S.; Scheer, A.K.; Randolph, H.E.; Thompson, T.W.; et al. Contribution of NK cells to immunotherapy mediated by PD-1/PD-L1 blockade. J. Clin. Investig. 2018, 128, 4654–4668. [Google Scholar] [CrossRef] [PubMed]
- Meloche, S.; Pouyssegur, J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 2007, 26, 3227–3239. [Google Scholar] [CrossRef] [Green Version]
- Marampon, F.; Ciccarelli, C.; Zani, B.M. Down-regulation of c-Myc following MEK/ERK inhibition halts the expression of malignant phenotype in rhabdomyosarcoma and in non muscle-derived human tumors. Mol. Cancer 2006, 5, 31. [Google Scholar] [CrossRef]
- Zhu, J.; Blenis, J.; Yuan, J. Activation of PI3K/Akt and MAPK pathways regulates Myc-mediated transcription by phosphorylating and promoting the degradation of Mad1. Proc. Natl. Acad. Sci. USA 2008, 105, 6584–6589. [Google Scholar] [CrossRef] [Green Version]
- Obaya, A.J.; Mateyak, M.K.; Sedivy, J.M. Mysterious liaisons: The relationship between c-Myc and the cell cycle. Oncogene 1999, 18, 2934–2941. [Google Scholar] [CrossRef]
- Marrocco, I.; Romaniello, D.; Yarden, Y. Cancer Immunotherapy: The Dawn of Antibody Cocktails. Methods Mol. Biol. 2019, 11–51. [Google Scholar]
- Manca, P.; Raez, L.E.; Salzberg, M.; Sanchez, J.; Hunis, B.; Rolfo, C. The value of immunotherapy in head and neck cancer. Expert Opin. Biol. Ther. 2019, 19, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Ghysen, K.; Vansteenkiste, J. Immunotherapy in patients with early stage resectable nonsmall cell lung cancer. Curr. Opin. Oncol. 2019, 31, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Hamid, O.; Robert, C.; Daud, A.; Hodi, F.S.; Hwu, W.J.; Kefford, R.; Wolchok, J.D.; Hersey, P.; Joseph, R.W.; Weber, J.S.; et al. Safety and tumor responses with lambrolizumab (anti-PD-1) in melanoma. N. Engl. J. Med. 2013, 369, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Carvajal, R.D.; Sosman, J.A.; Atkins, M.B.; et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N. Engl. J. Med. 2012, 366, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.S.; Barroso-Sousa, R.; Tolaney, S.M.; Hodi, F.S.; Kaiser, U.B.; Min, L. Endocrine Toxicity of Cancer Immunotherapy Targeting Immune Checkpoints. Endocr. Rev. 2019, 40, 17–65. [Google Scholar] [CrossRef] [PubMed]
- Nishino, M.; Hatabu, H.; Hodi, F.S. Imaging of Cancer Immunotherapy: Current Approaches and Future Directions. Radiology 2019, 290, 9–22. [Google Scholar] [CrossRef]
- Sibbitt, W.L., Jr.; Bankhurst, A.D.; Jumonville, A.J.; Saiki, J.H.; Saiers, J.H.; Doberneck, R.C. Defects in natural killer cell activity and interferon response in human lung carcinoma and malignant melanoma. Cancer Res. 1984, 44, 852–856. [Google Scholar]
- Al Omar, S.Y.; Marshall, E.; Middleton, D.; Christmas, S.E. Increased killer immunoglobulin-like receptor expression and functional defects in natural killer cells in lung cancer. Immunology 2011, 133, 94–104. [Google Scholar] [CrossRef]
- Richards, J.O.; Chang, X.; Blaser, B.W.; Caligiuri, M.A.; Zheng, P.; Liu, Y. Tumor growth impedes natural-killer-cell maturation in the bone marrow. Blood 2006, 108, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Sung, P.S.; Jang, J.W. Natural Killer Cell Dysfunction in Hepatocellular Carcinoma: Pathogenesis and Clinical Implications. Int. J. Mol. Sci. 2018, 19, 3648. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Zou, J.; Feng, X.; Zhang, C.; Zheng, R.; Duanmu, W.; Saha-Mandal, A.; Ming, Z.; Wang, E. Inherited defects in natural killer cells shape tumor immune microenvironment, clinical outcome and immunotherapy response. BioRxiv 2018, 471912. [Google Scholar] [CrossRef]
- Tansey, W.P. Mammalian MYC Proteins and Cancer. New J. Sci. 2014, 27. [Google Scholar] [CrossRef]
- Casey, S.C.; Baylot, V.; Felsher, D.W. The MYC oncogene is a global regulator of the immune response. Blood 2018, 131, 2007–2015. [Google Scholar] [CrossRef] [PubMed]
- Muntasell, A.; Ochoa, M.C.; Cordeiro, L.; Berraondo, P.; Lopez-Diaz de Cerio, A.; Cabo, M.; Lopez-Botet, M.; Melero, I. Targeting NK-cell checkpoints for cancer immunotherapy. Curr. Opin. Immunol. 2017, 45, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef]
- Dahlberg, C.I.; Sarhan, D.; Chrobok, M.; Duru, A.D.; Alici, E. Natural Killer Cell-Based Therapies Targeting Cancer: Possible Strategies to Gain and Sustain Anti-Tumor Activity. Front. Immunol. 2015, 6, 605. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakiryanova, G.K.; Kustova, E.; Urazalieva, N.T.; Baimuchametov, E.T.; Nakisbekov, N.N.; Shurin, M.R. Abnormal Expression of c-Myc Oncogene in NK Cells in Patients with Cancer. Int. J. Mol. Sci. 2019, 20, 756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030756
Zakiryanova GK, Kustova E, Urazalieva NT, Baimuchametov ET, Nakisbekov NN, Shurin MR. Abnormal Expression of c-Myc Oncogene in NK Cells in Patients with Cancer. International Journal of Molecular Sciences. 2019; 20(3):756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030756
Chicago/Turabian StyleZakiryanova, Gulnur K., Elena Kustova, Nataliya T. Urazalieva, Emile T. Baimuchametov, Narymzhan N. Nakisbekov, and Michael R. Shurin. 2019. "Abnormal Expression of c-Myc Oncogene in NK Cells in Patients with Cancer" International Journal of Molecular Sciences 20, no. 3: 756. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030756