MIST1, an Inductive Signal for Salivary Amylase in Mesenchymal Stem Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
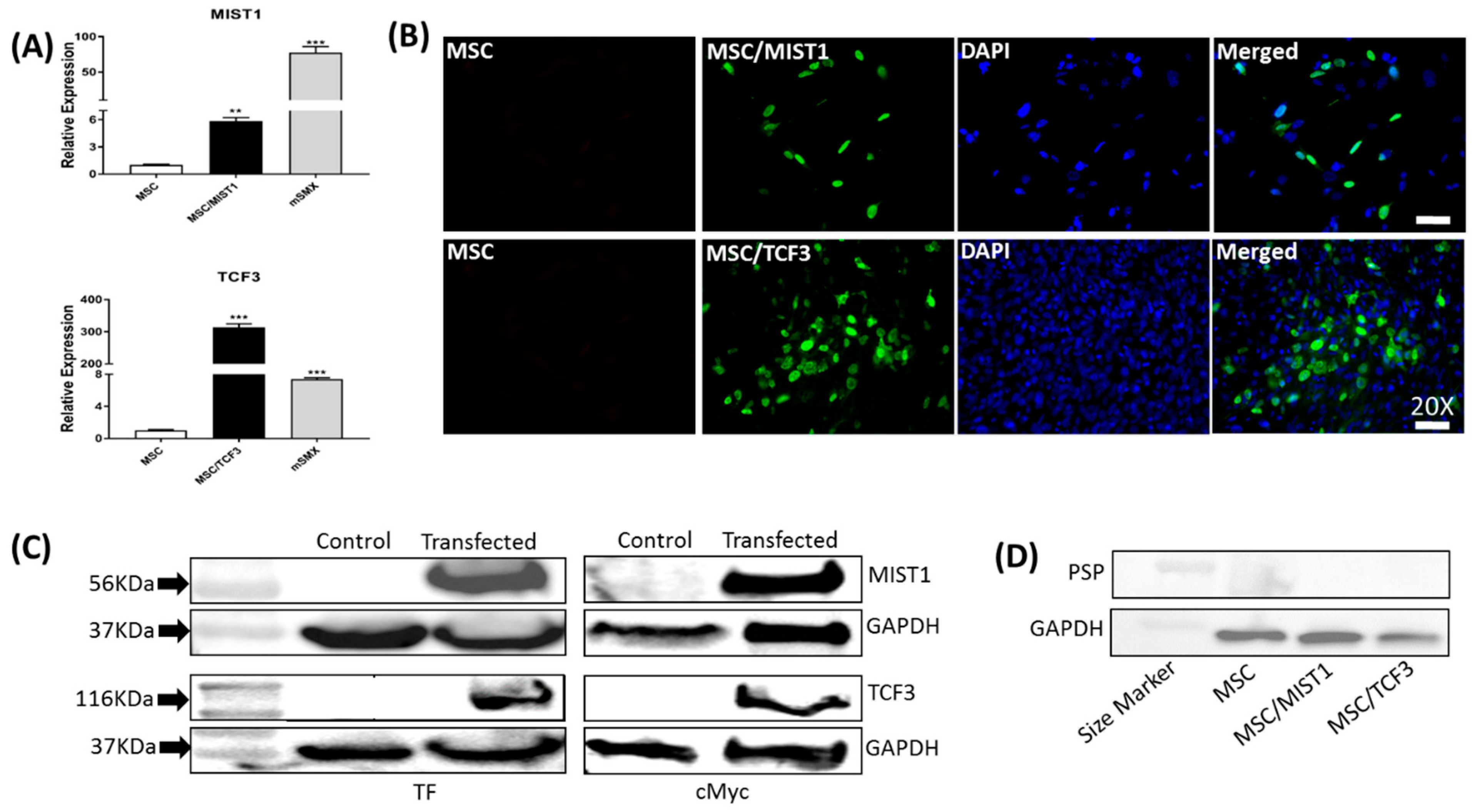
2.1. Recombinant MIST1 or TCF3 Expression Was Confirmed in mMSCs
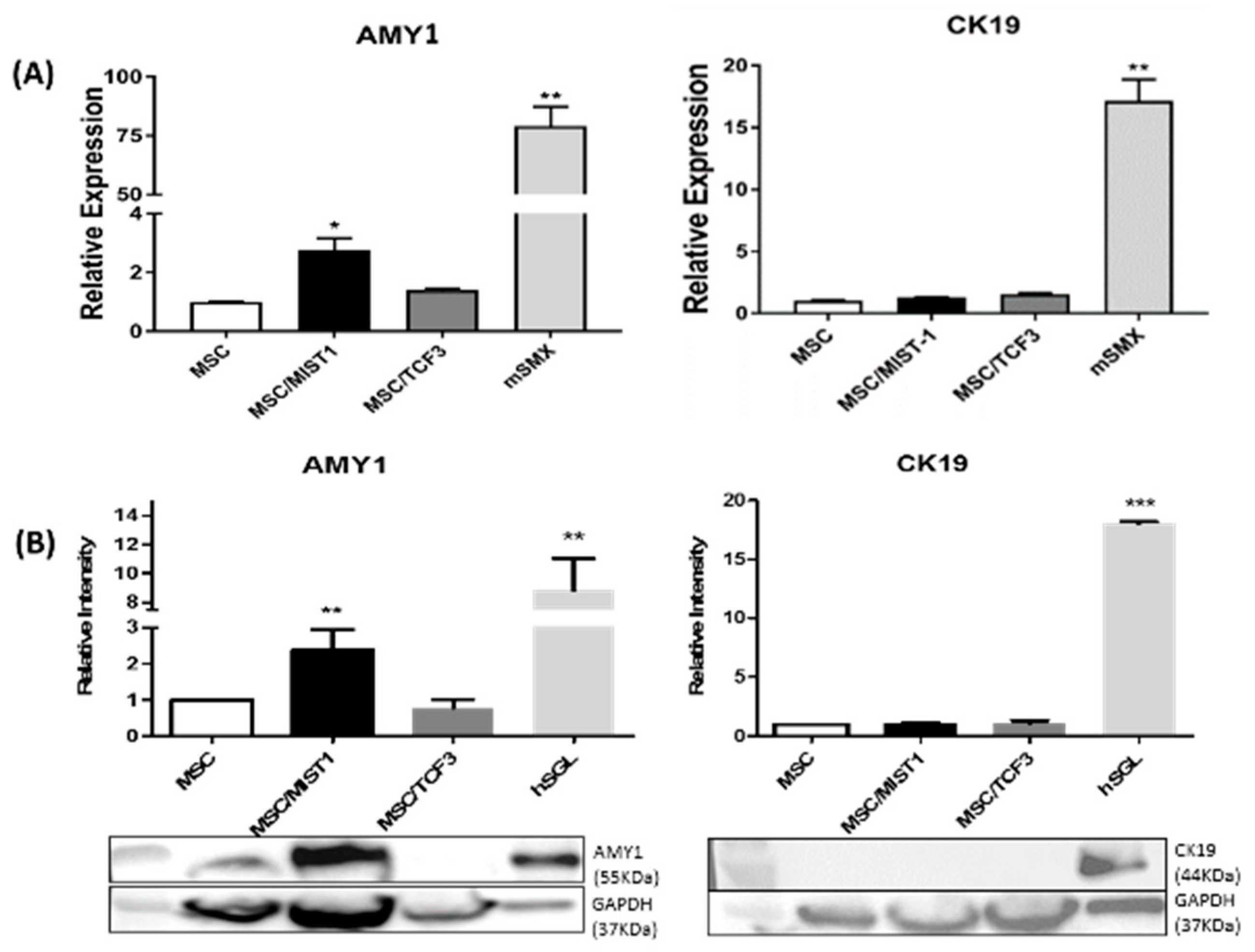
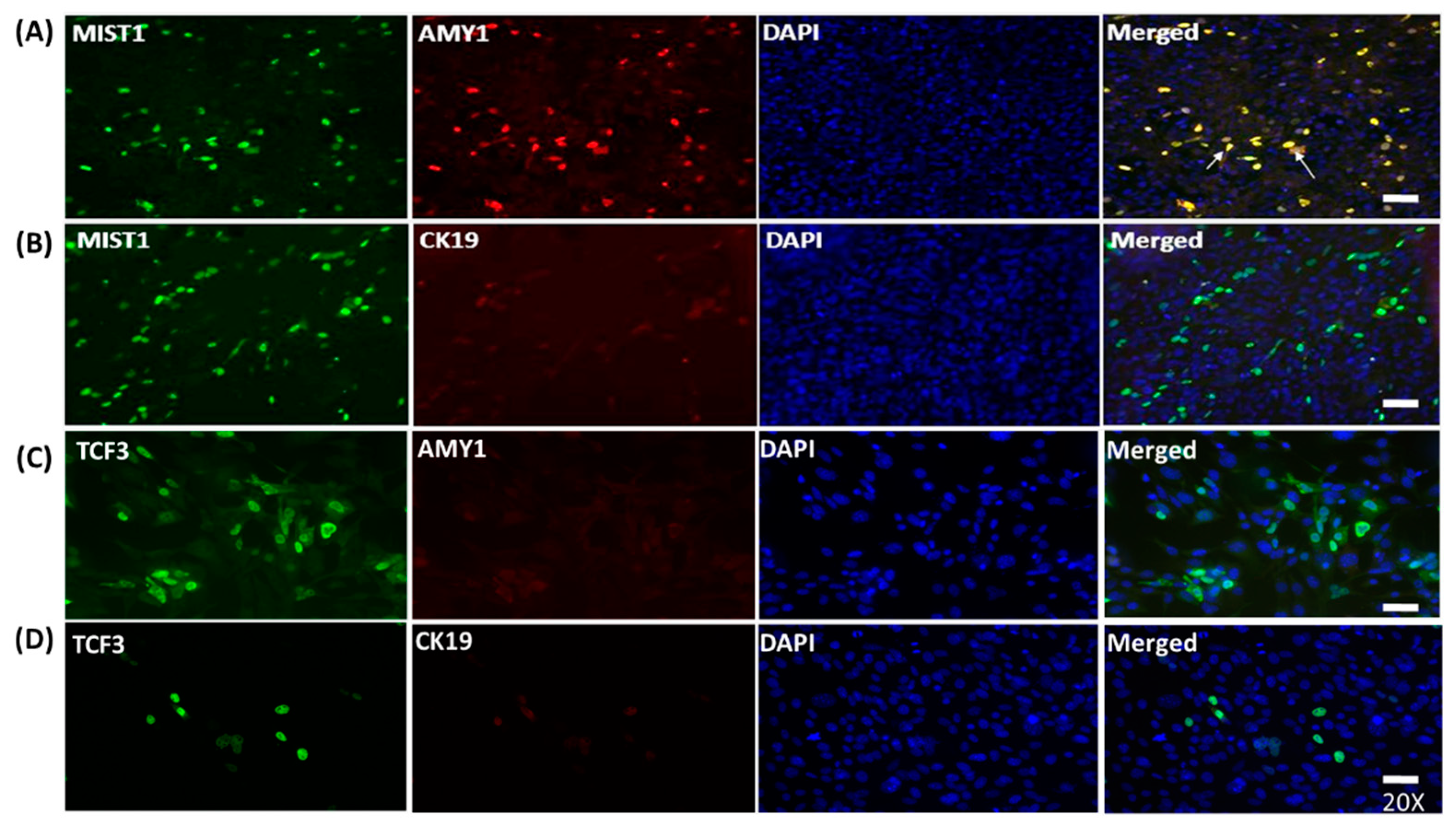
2.2. MIST1 Promotes AMY1 in mMSCs Whereas TCF3 Does not Induce its Expression
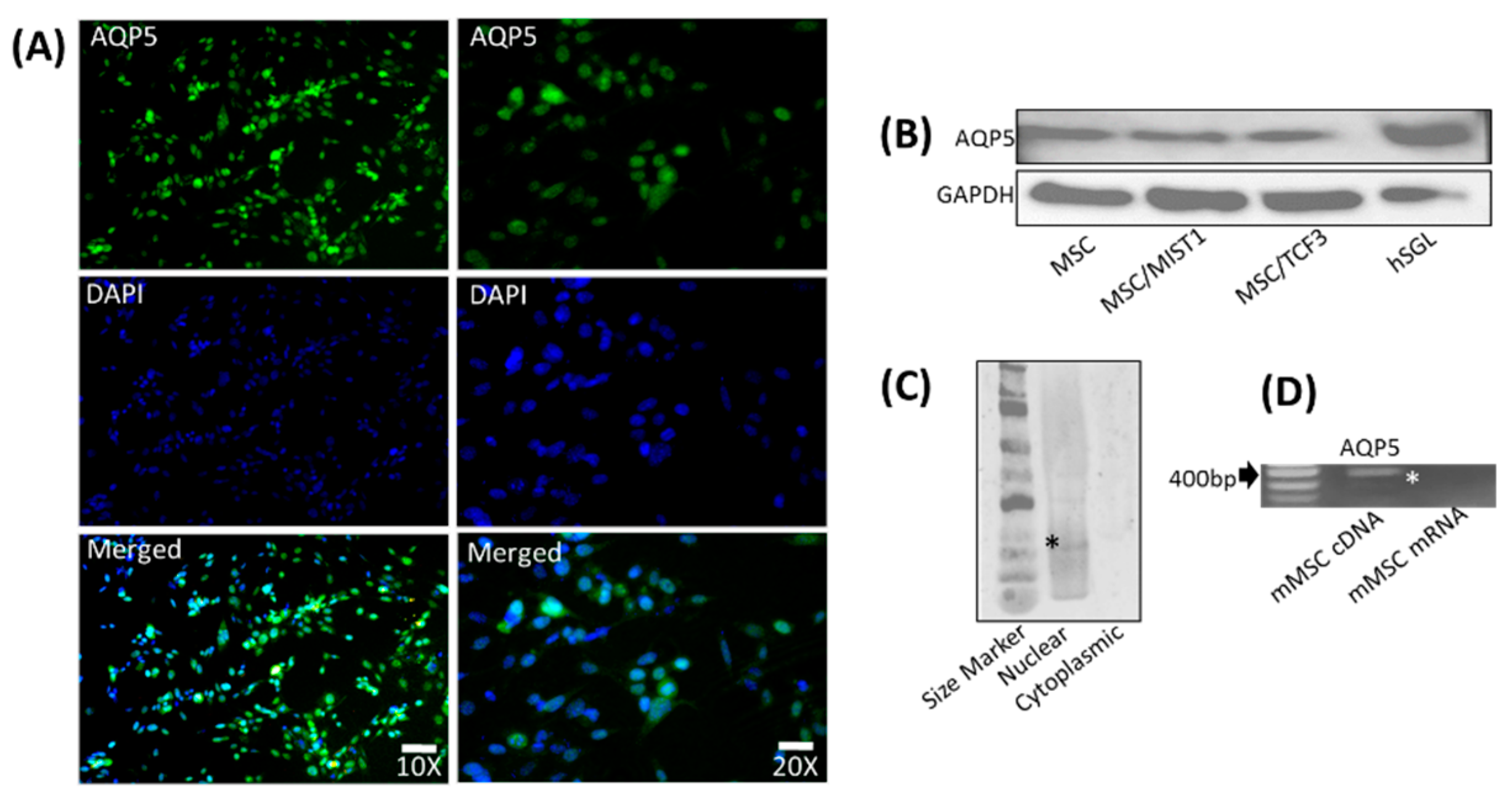
2.3. mMSCs Express AQP5, Which Is Localized to the Nucleus
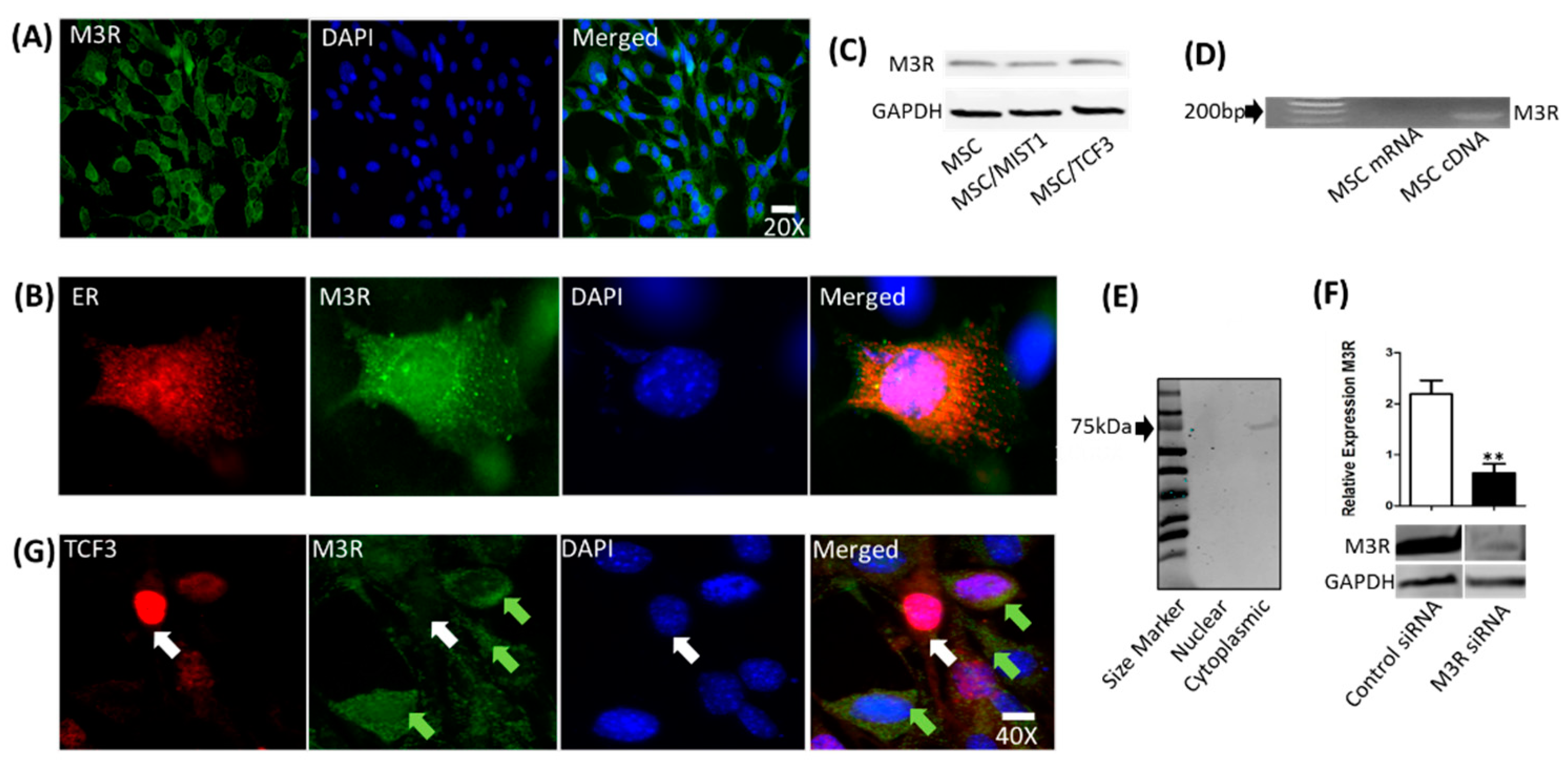
2.4. mMSCs Express M3R, Which Is Localized to the Endoplasmic Reticulum (ER)
3. Discussion
4. Limitations and Future Directions
5. Materials and Methods
5.1. Mesenchymal Stem Cell Culture
5.2. Recombinant MIST1 and TCF3 Expression
5.3. Transient Transfection
5.4. RNA Isolation, RT-PCR, and Quantitative Real-Time PCR (qRT-PCR)
5.5. Western Blot (WB) Analysis
5.6. Immunocytochemistry (ICC)
5.7. M3R Protein Knockdown by siRNA
5.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stem cells |
| SjS | Sjögren’s syndrome |
| mMSCs | mouse bone marrow-derived MSCs |
| TFs | transcription factors |
| AMY1 | alpha-salivary amylase 1 |
| CK19 | cytokeratin19 |
| MET, | mesenchymal-epithelial transition |
| MIST1 | muscle, intestine and stomach expression-1 |
| TCF3 | transcription factor E2a |
| PTF1α | pancreas-specific transcription factor 1 α |
| ASCL3 | achaete-scute complex homolog 3 |
| ANKRD56 | ankyrin repeat domain-containing protein 56 |
| HMG20B | high mobility group protein 20B |
| M3R | muscarinic type 3 receptor |
| AQP5 | aquaporin 5 |
| WB | western blot |
| ICC | Immunocytochemistry |
| RT-PCR | reverse transcription PCR |
| qRT-PCR | quantitative real-time PCR |
References
- Mavragani, C.P.; Moutsopoulos, H.M. Sjögren syndrome. CMAJ 2014, 186, E579–E586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.J.; Koh, J.; Kwon, J.T.; Park, Y.S.; Yang, L.; Cha, S. Uncovering stem cell differentiation factors for salivary gland regeneration by quantitative analysis of differential proteomes. PLoS ONE 2017, 12, e0169677. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Pal, M.; Rawat, S.; Grewal, M.S.; Garg, H.; Chauhan, D.; Ahlawat, P.; Tandon, S.; Khurana, R.; Pahuja, A.K.; et al. Radiation-induced dental caries, prevention and treatment—A systematic review. Natl. J. Maxillofac. Surg. 2015, 6, 160–166. [Google Scholar] [PubMed]
- Lee, B.H.; Gauna, A.E.; Perez, G.; Park, Y.J.; Pauley, K.M.; Kawai, T.; Cha, S. Autoantibodies against muscarinic type 3 receptor in Sjögren’s syndrome inhibit aquaporin 5 trafficking. PLoS ONE 2013, 8, e53113. [Google Scholar] [CrossRef] [PubMed]
- Proctor, G.B. The physiology of salivary secretion. Periodontol 2000, 2016, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Janeček, Š.; Svensson, B.; MacGregor, E.A. α-Amylase: An enzyme specificity found in various families of glycoside hydrolases. Cell Mol. Life Sci. 2014, 71, 1149–1170. [Google Scholar]
- Kho, H.S. Understanding of xerostomia and strategies for the development of artificial saliva. Chin. J. Dent. Res. 2014, 17, 75–83. [Google Scholar]
- Kramer, J.M. Current concepts in Sjögren’s syndrome and considerations for the dental practitioner. N. Y. State Dent. J. 2015, 81, 24–29. [Google Scholar]
- Cifuentes, M.; Del Barrio-Díaz, P.; Vera-Kellet, C. Pilocarpine and artificial saliva for the treatment of xerostomia and xerophthalmia in Sjögren syndrome: A double-blind randomized controlled trial. Br. J. Dermatol. 2018, 179, 1056–1061. [Google Scholar] [CrossRef]
- Cassolato, S.F.; Turnbull, R.S. Xerostomia: Clinical aspects and treatment. Gerodontology 2003, 20, 64–77. [Google Scholar] [CrossRef]
- Hanchanale, S.; Adkinson, L.; Daniel, S.; Fleming, M.; Oxberry, S.G. Systematic literature review: Xerostomia in advanced cancer patients. Support Care Cancer 2015, 23, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Jensen, D.H.; Oliveri, R.S.; Trojahn Kølle, S.F.; Fischer-Nielsen, A.; Specht, L.; Bardow, A.; Buchwald, C. Mesenchymal stem cell therapy for salivary gland dysfunction and xerostomia: A systematic review of preclinical studies. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. 2014, 117, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Baddour, J.A.; Sousounis, K.; Tsonis, P.A. Organ repair and regeneration: An overview. Birth Defects Res. C. Embryo. Today 2012, 96, 1–29. [Google Scholar] [CrossRef]
- Klassen, H. Stem cells in clinical trials for treatment of retinal degeneration. Expert Opin. Biol. Ther. 2016, 16, 7–14. [Google Scholar] [CrossRef]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.F.; Lian, Q. Paracrine mechanisms of mesenchymal stem cell-based therapy: Current status and perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Van Luijk, P.; Pringle, S.; Deasy, J.O.; Moiseenko, V.V.; Faber, H.; Hovan, A.; Baanstra, M.; van der Laan, H.P.; Kierkels, R.G.; van der Schaaf, A.; et al. Sparing the region of the salivary gland containing stem cells preserves saliva production after radiotherapy for head and neck cancer. Sci. Transl. Med. 2015, 7, 305. [Google Scholar] [CrossRef]
- Yoo, C.; Vines, J.B.; Alexander, G.; Murdock, K.; Hwang, P.; Jun, H.W. Adult stem cells and tissue engineering strategies for salivary gland regeneration: A review. Biomater. Res. 2014, 18, 9. [Google Scholar] [CrossRef]
- Lodi, D.; Iannitti, T.; Palmieri, B. Stem cells in clinical practice: Applications and warnings. J. Exp. Clin. Cancer Res. 2011, 30, 9. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Prockop, D.J. Marrow stromal cells as stem cells for continual renewal of nonhematopoietic tissues and as potential vectors for gene therapy. J. Cell. Biochem. 1998, 9, 284–285. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Haynesworth, S.E.; Goshima, J.; Goldberg, V.M.; Caplan, A.I. Characterization of cells with osteogenic potential from human marrow. Bone 1992, 13, 81–88. [Google Scholar] [CrossRef]
- Majumdar, M.K.; Thiede, M.A.; Mosca, J.D.; Moorman, M.; Gerson, S.L. Phenotypic and functional comparison of cultures of marrow-derived mesenchymal stem cells (MSCs) and stromal cells. J. Cell Physiol. 1998, 176, 57–66. [Google Scholar] [CrossRef]
- Aurich, H.; Sgodda, M.; Kaltwasser, P.; Vetter, M.; Weise, A.; Liehr, T.; Brulport, M.; Hengstler, J.G.; Dollinger, M.M.; Fleig, W.E.; et al. Hepatocyte differentiation of mesenchymal stem cells from human adipose tissue in vitro promotes hepatic integration in vivo. Gut 2009, 58, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Seaberg, R.M.; Smukler, S.R.; Kieffer, T.J.; Enikolopov, G.; Asghar, Z.; Wheeler, M.B.; Korbutt, G.; van der Kooy, D. Clonal identification of multipotent precursors from adult mouse pancreas that generate neural and pancreatic lineages. Nat. Biotechnol. 2004, 22, 1115–1124. [Google Scholar] [CrossRef]
- Katuchova, J.; Harvanova, D.; Spakova, T.; Kalanin, R.; Farkas, D.; Durny, P.; Rosocha, J.; Radonak, J.; Petrovic, D.; Siniscalco, D.; et al. Mesenchymal stem cells in the treatment of type 1 diabetes mellitus. Endocr. Pathol. 2015, 26, 95–103. [Google Scholar] [CrossRef]
- Park, Y.J.; Koh, J.; Gauna, A.E.; Chen, S.U.; Cha, S.H. Identification of Regulatory Factors for Mesenchymal Stem Cell-Derived Salivary Epithelial Cells in a Co-Culture System. PLoS ONE 2014, 9, e112158. [Google Scholar] [CrossRef]
- Wang, D.; Iwata, F.; Muraguchi, M.; Ooga, K.; Ohmoto, Y.; Takai, M.; Mori, T.; Ishikawa, Y. Correlation between salivary secretion and salivary AQP5 levels in health and disease. J. Med. Investig. 2009, 56, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Aure, M.H.; Ruus, A.K.; Galtung, H.K. Aquaporins in the adult mouse submandibular and sublingual salivary glands. J. Mol. Histol. 2014, 45, 69–80. [Google Scholar] [CrossRef]
- Proctor, G.B.; Carpenter, G.H. Regulation of salivary gland function by autonomic nerves. Auton Neurosci. 2007, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Gresz, V.; Kwon, T.H.; Hurley, P.T.; Varga, G.; Zelles, T.; Nielsen, S.; Case, R.M.; Steward, M.C. Identification and localization of aquaporin water channels in human salivary glands. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G247–G254. [Google Scholar] [CrossRef] [PubMed]
- Scarpero, H.M.; Dmochowski, R.R. Muscarinic receptors: What we know. Curr. Urol. Rep. 2003, 4, 421–428. [Google Scholar] [CrossRef]
- Sumida, T.; Tsuboi, H.; Iizuka, M.; Nakamura, Y.; Matsumoto, I. Functional role of M3 muscarinic acetylcholine receptor (M3R) reactive T cells and anti-M3R autoantibodies in patients with Sjögren’s syndrome. Autoimmun. Rev. 2010, 9, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Aure, M.H.; Konieczny, S.F.; Ovitt, C.E. Salivary gland homeostasis is maintained through acinar cell self-duplication. Dev. Cell 2015, 33, 231–237. [Google Scholar] [CrossRef]
- Direnzo, D.; Hess, D.A.; Damsz, B.; Hallett, J.E.; Marshall, B.; Goswami, C.; Liu, Y.; Deering, T.; Macdonald, R.J.; Konieczny, S.F. Induced Mist1 expression promotes remodeling of mouse pancreatic acinar cells. Gastroenterology 2012, 143, 469–480. [Google Scholar] [CrossRef]
- Yamagishi, R.; Wakayama, T.; Nakata, H.; Adthapanyawanich, K.; Kumchantuek, T.; Yamamoto, M.; Iseki, S. Expression and Localization of α-amylase in the Submandibular and Sublingual Glands of Mice. Acta Histochem. Cytochem. 2014, 47, 95–102. [Google Scholar] [CrossRef]
- Kawaguchi, Y.; Cooper, B.; Gannon, M.; Ray, M.; MacDonald, R.J.; Wright, C.V. The role of the transcriptional regulator Ptf1a in converting intestinal to pancreatic progenitors. Nat. Genet. 2002, 32, 128–134. [Google Scholar] [CrossRef]
- Obata, J.; Yano, M.; Mimura, H.; Goto, T.; Nakayama, R.; Mibu, Y.; Oka, C.; Kawaichi, M. p48 subunit of mouse PTF1 binds to RBP-Jkappa/CBF-1, the intracellular mediator of Notch signalling, and is expressed in the neural tube of early stage embryos. Genes Cells 2001, 6, 345–360. [Google Scholar] [CrossRef] [Green Version]
- Pin, C.L.; Rukstalis, J.M.; Johnson, C.; Konieczny, S.F. The bHLH transcription factor Mist1 is required to maintain exocrine pancreas cell organization and acinar cell identity. J. Cell. Biol. 2001, 155, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.; Jia, D.; Sun, Y.; Konieczny, S.F. The bHLH domain of Mistl is sufficient to activate gene transcription. Gene Expr. 2007, 13, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Azevedo-Pouly, A.; Deering, T.G.; Hoang, C.Q.; DiRenzo, D.; Hess, D.A.; Konieczny, S.F.; Swift, G.H.; MacDonald, R.J. MIST1 and PTF1 Collaborate in Feed-forward Regulatory Loops that Maintain the Pancreatic Acinar Phenotype in Adult Mice. Mol. Cell. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Barndt, R.J.; Dai, M.; Zhuang, Y. Functions of E2A-HEB heterodimers in T-cell development revealed by a dominant negative mutation of HEB. J. Exp. Med. 2005, 202, 1669–1677. [Google Scholar] [CrossRef]
- Hanrahan, J.; Pan, L.; Greenbaum, S.; Bradney, C.; Hjelmeland, M.; Dai, M.; Zhuang, Y. A genetic investigation of E2A function in lymphocyte development. Immunol. Res. 2000, 22, 211–222. [Google Scholar] [CrossRef]
- Borghesi, L.; Aites, J.; Nelson, S.; Lefterov, P.; James, P.; Gerstein, R. E47 is required for V(D)J recombinase activity in common lymphoid progenitors. J. Exp. Med. 2005, 202, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lahmy, R.; Riha, C.; Yang, C.; Jakubison, B.L.; van Niekerk, J.; Staub, C.; Wu, Y.; Gates, K.; Dong, D.S.; et al. The basic helix-loop-helix transcription factor E47 reprograms human pancreatic cancer cells to a quiescent acinar state with reduced tumorigenic potential. Pancreas 2015, 44, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Beroukas, D.; Goodfellow, R.; Hiscock, J.; Jonsson, R.; Gordon, T.P.; Waterman, S.A. Up-regulation of M3-muscarinic receptors in labial salivary gland acini in primary Sjögren’s syndrome. Lab. Investig. 2002, 82, 203–210. [Google Scholar] [CrossRef]
- Qin, K.; Dong, C.; Wu, G.; Lambert, N.A. Inactive-state preassembly of G(q)-coupled receptors and G(q) heterotrimers. Nat. Chem. Biol. 2011, 7, 740–747. [Google Scholar] [CrossRef]
- Nakamura, T.; Matsui, M.; Uchida, K.; Futatsugi, A.; Kusakawa, S.; Matsumoto, N.; Nakamura, K.; Manabe, T.; Taketo, M.M.; Mikoshiba, K. M(3) muscarinic acetylcholine receptor plays a critical role in parasympathetic control of salivation in mice. J. Physiol. 2004, 558, 561–575. [Google Scholar] [CrossRef]
- Piovesana, R.; Melfi, S.; Fiore, M.; Magnaghi, V.; Tata, A.M. M2 muscarinic receptor activation inhibits cell proliferation and migration of rat adipose-mesenchymal stem cells. J. Cell Physiol. 2018, 233, 5348–5360. [Google Scholar] [CrossRef]
- Kotton, D.N.; Ma, B.Y.; Cardoso, W.V.; Sanderson, E.A.; Summer, R.S.; Williams, M.C.; Fine, A. Bone marrow-derived cells as progenitors of lung alveolar epithelium. Development. Development 2001, 128, 5181–5188. [Google Scholar]
- Zhang, H.; Fang, J.; Wu, Y.; Mai, Y.; Lai, W.; Su, H. Mesenchymal stem cells protect against neonatal rat hyperoxic lung injury. Expert Opin. Biol. Ther. 2013, 13, 817–829. [Google Scholar] [CrossRef]
- Yi, F.; Khan, M.; Gao, H.; Hao, F.; Sun, M.; Zhong, L.; Lu, C.; Feng, X.; Ma, T. Increased differentiation capacity of bone marrow-derived mesenchymal stem cells in aquaporin-5 deficiency. Stem Cells Dev. 2012, 21, 2495–2507. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Eguchi, T.; Skowronski, M.T.; Ishida, H. Acetylcholine acts on M3 muscarinic receptors and induces the translocation of aquaporin5 water channel via cytosolic Ca2+ elevation in rat parotid glands. Biochem. Biophys. Res. Commun. 1998, 245, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, Y.; Skowronski, M.T.; Ishida, H. Persistent increase in the amount of aquaporin-5 in the apical plasma membrane of rat parotid acinar cells induced by a muscarinic agonist SNI-2011. FEBS Lett. 2000, 477, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Cho, G.; Bragiel, A.M.; Wang, D.; Pieczonka, T.D.; Skowronski, M.T.; Shono, M.; Nielsen, S.; Ishikawa, Y. Activation of muscarinic receptors in rat parotid acinar cells induces AQP5 trafficking to nuclei and apical plasma membrane. Biochim. Biophys. Acta. 2015, 1850, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mona, M.; Miller, R.; Li, H.; Park, Y.-J.; Zaman, R.; Yang, L.-J.; Cha, S. MIST1, an Inductive Signal for Salivary Amylase in Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 767. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030767
Mona M, Miller R, Li H, Park Y-J, Zaman R, Yang L-J, Cha S. MIST1, an Inductive Signal for Salivary Amylase in Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(3):767. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030767
Chicago/Turabian StyleMona, Mahmoud, Rehae Miller, Hui Li, Yun-Jong Park, Raafi Zaman, Li-Jun Yang, and Seunghee Cha. 2019. "MIST1, an Inductive Signal for Salivary Amylase in Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 3: 767. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030767