Altered Promoter and G-Box Binding Factor for 1-Deoxy-d-Xylulose-5-Phosphate Synthase Gene Grown from Poa pratensis Seeds after Spaceflight


Abstract
:1. Introduction
2. Result
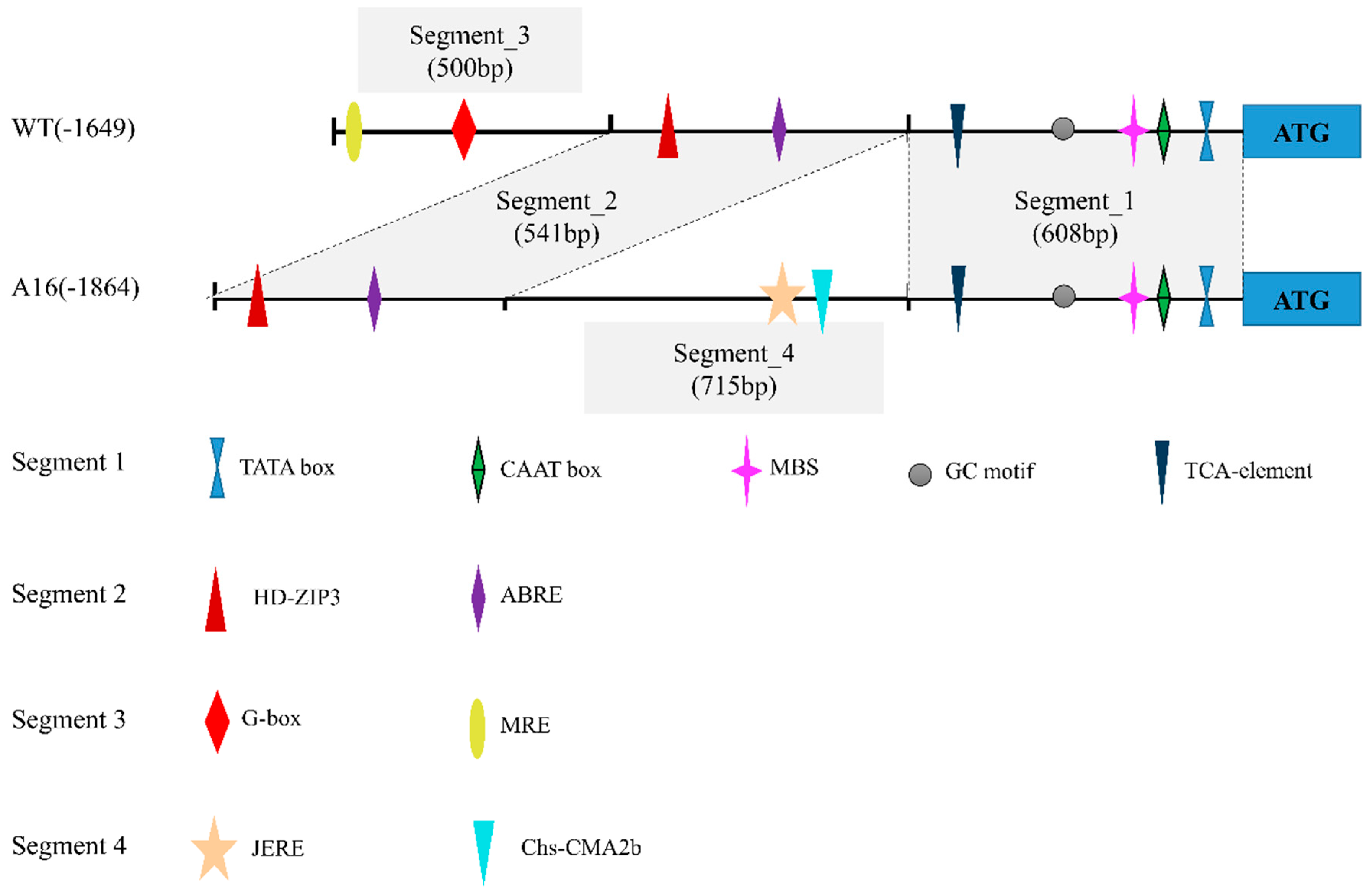
2.1. Isolation of Upstream Regulatory Region of PpDXS1
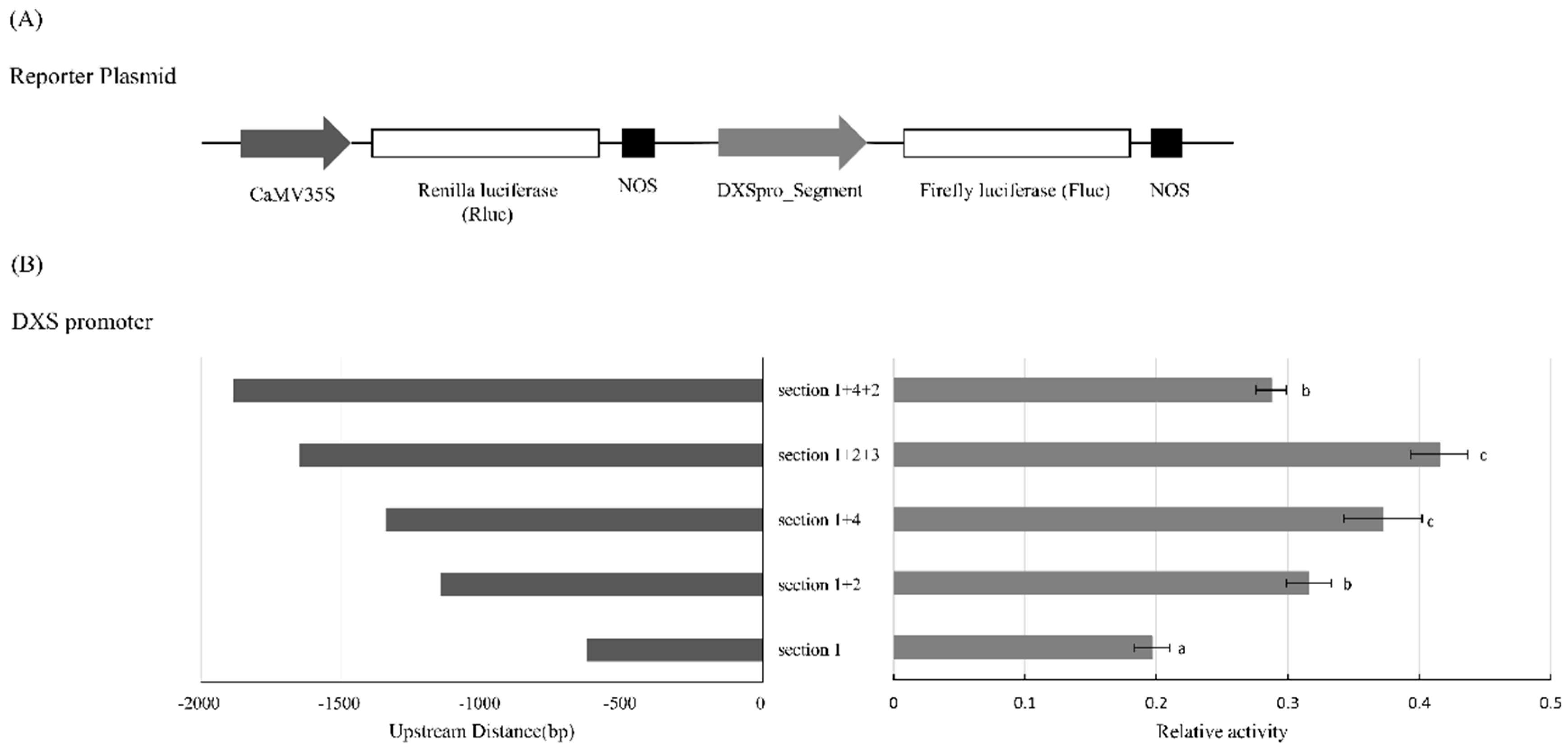
2.2. Activity of PpDXS1 Promoter in the Dual Luciferase Transient Protoplast Assay
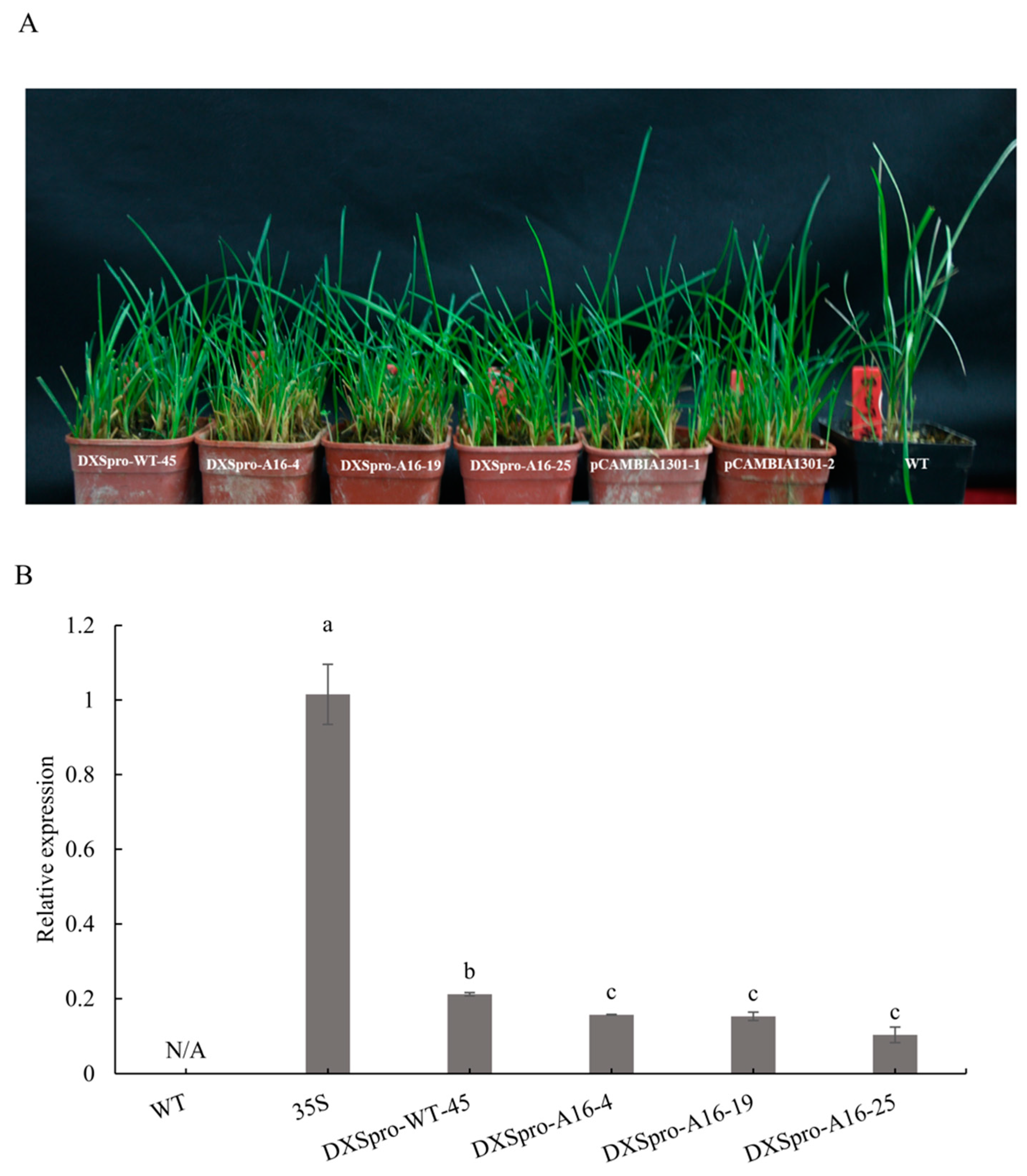
2.3. Promoter Activity in Poa pratensis Plants
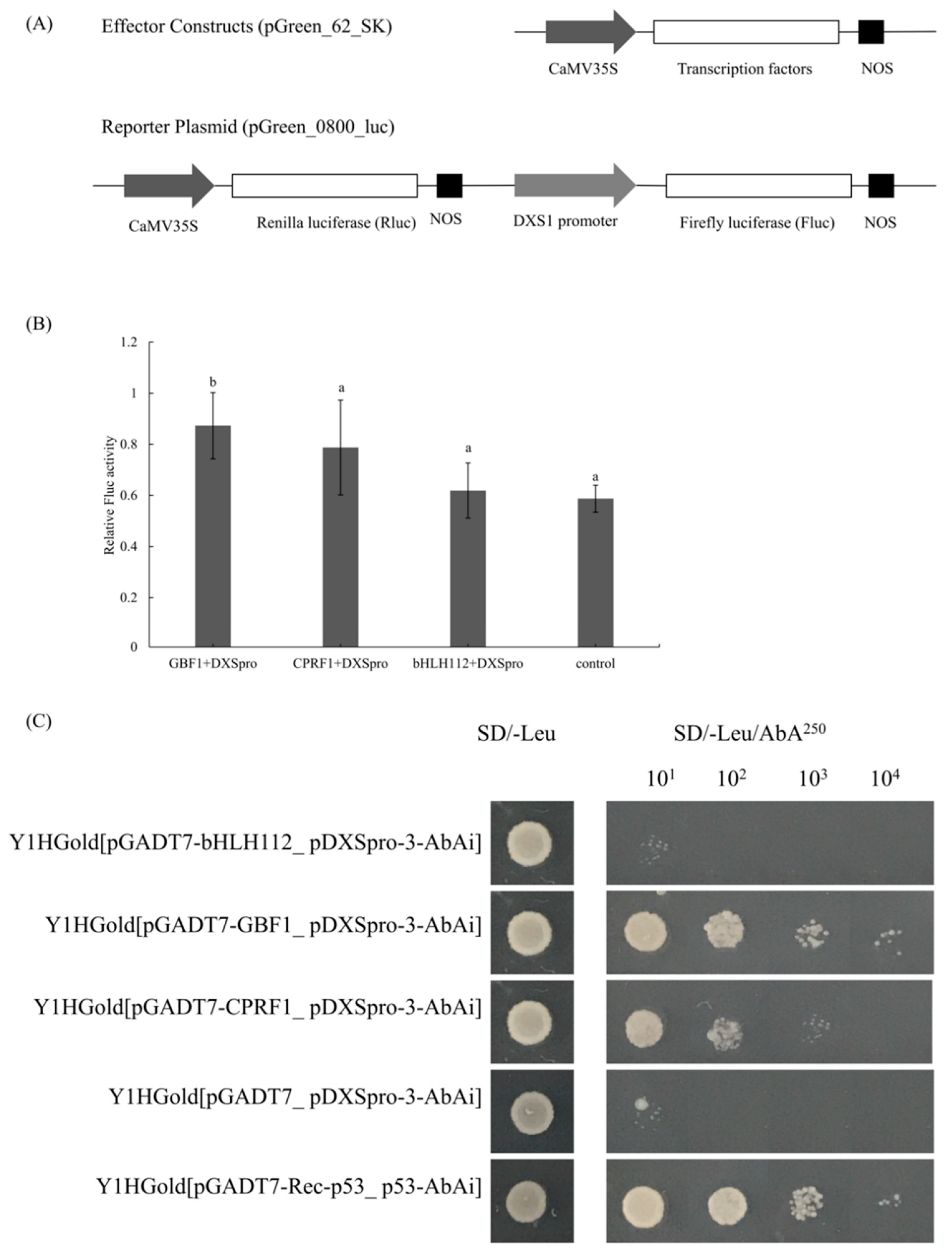
2.4. Enhanced Activity by Addition of GBF1 to DXS1 Promoter
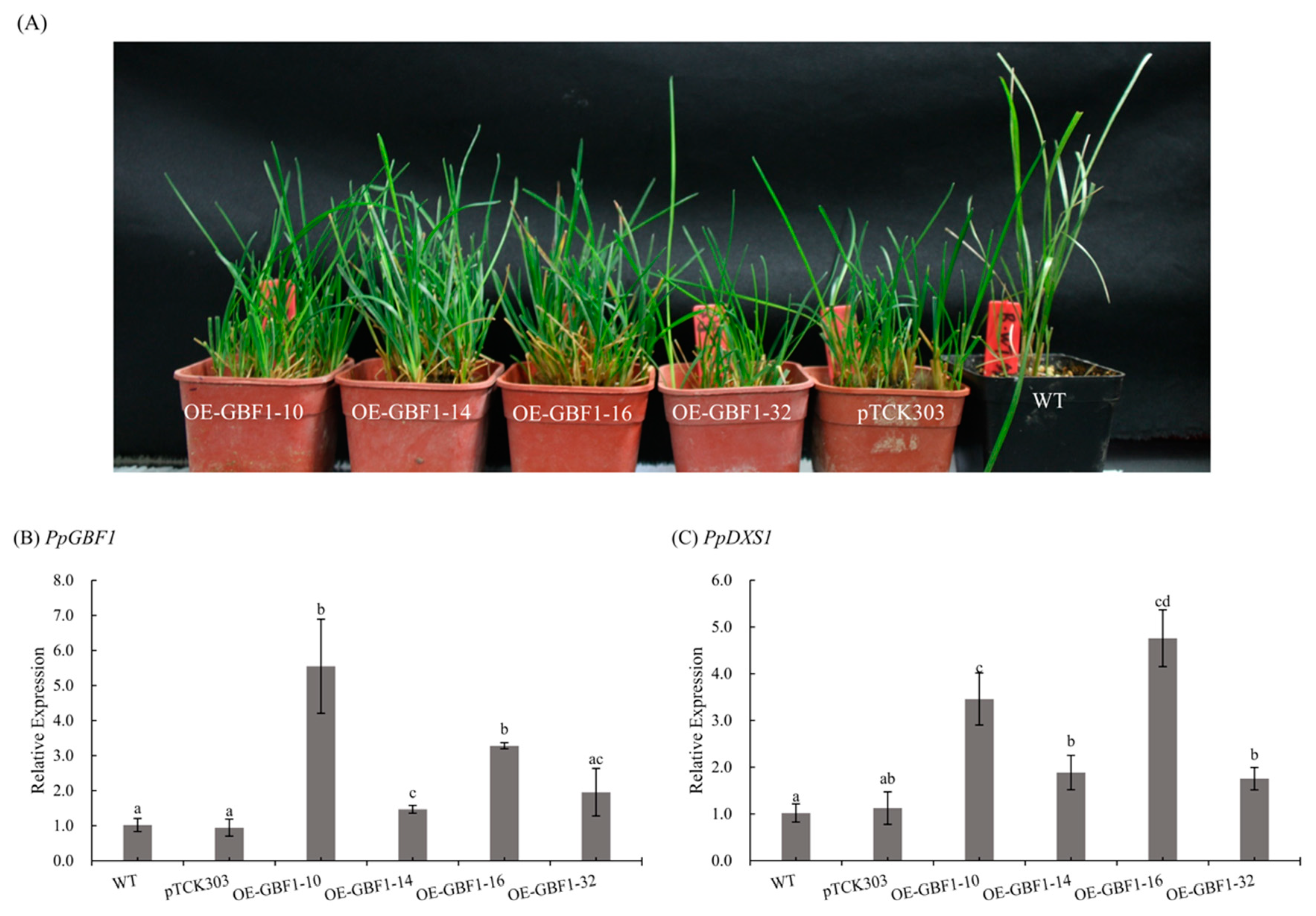
2.5. Overexpression of GBF1 Gene in Poa pratensis
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Isolation and Analysis of PpDXS1 Promoter Region
4.3. Dual Luciferase Transient Protoplast Assay Using Poa pratensis Protoplast
4.4. Yeast One-Hybrid Assay
4.5. Transformation of Poa pratensis and GUS Histochemical Assay
4.6. Expression Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bach, T.J. Some new aspects of isoprenoid biosynthesis in plants—A review. Lipids 1995, 30, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Concepción, M.; Boronat, A. Breaking new ground in the regulation of the early steps of plant isoprenoid biosynthesis. Curr. Opin. Plant Biol. 2015, 25, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Structure and dynamics of the isoprenoid pathway network. Mol. Plant 2012, 5, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Srinath, M.; Shailaja, A.; Bhavani, B.; Bindu, V.; Giri, C.C. Characterization of 1-deoxy-d-xylulose 5-phosphate synthase (DXS) protein in Andrographis paniculata (Burm. f.) Wall. ex. Nees: A in silico appraisal. Ann. Phytomed. Int. J. 2017, 6, 63–73. [Google Scholar] [CrossRef]
- Estevez, J.M.; Cantero, A.; Reindl, A.; Reichler, S.; Leon, P. 1-Deoxy-d-xylulose-5-phosphate synthase, a limiting enzyme for plastidic isoprenoid biosynthesis in plants. J. Biol. Chem. 2001, 276, 22901–22909. [Google Scholar] [CrossRef]
- Cordoba, E.; Porta, H.; Arroyo, A.; Román, C.S.; Medina, L.; Rodríguez-Concepción, M.; León, P. Functional characterization of the three genes encoding 1-deoxy-d-xylulose 5-phosphate synthase in maize. J. Exp. Bot. 2011, 62, 2023–2038. [Google Scholar] [CrossRef]
- Walter, M.H.; Hans, J.; Strack, D. Two distantly related genes encoding 1-deoxy-d-xylulose 5-phosphate synthases: Differential regulation in shoots and apocarotenoid-accumulating mycorrhizal roots. Plant J. 2002, 31, 243–254. [Google Scholar] [CrossRef]
- Kim, B.-R.; Kim, S.-U.; Chang, Y.-J. Differential expression of three 1-deoxy-d-xylulose-5-phosphate synthase genes in rice. Biotechnol. Lett. 2005, 27, 997–1001. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, X.; Li, S.; Chen, W.; Li, J. Gene cloning and functional characterization of three 1-deoxy-d-xylulose 5-phosphate synthases in simao pine. Bioresources 2018, 13, 6370–6382. [Google Scholar]
- Okada, A.; Shimizu, T.; Okada, K.; Kuzuyama, T.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. Elicitor induced activation of the methylerythritol phosphate pathway toward phytoalexins biosynthesis in rice. Plant Mol. Biol. 2007, 65, 177–187. [Google Scholar] [CrossRef]
- Kim, D.S.; Kim, J.B.; Goh, E.J.; Kim, W.J.; Kim, S.H.; Seo, Y.W.; Jang, C.S.; Kang, S.Y. Antioxidant response of Arabidopsis plants to gamma irradiation: Genome-wide expression profiling of the ROS scavenging and signal transduction pathways. J. Plant Physiol. 2011, 168, 1960–1971. [Google Scholar] [CrossRef]
- De Micco, V.; Arena, C.; Pignalosa, D.; Durante, M. Effects of sparsely and densely ionizing radiation on plants. Radiat. Environ. Biophys. 2011, 50, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Arena, C.; de Micco, V.; Macaeva, E.; Quintens, R. Space radiation effects on plant and mammalian cells. Acta Astronaut. 2014, 104, 419–431. [Google Scholar] [CrossRef]
- Wels, M.; Overmars, L.; Francke, C.; Kleerebezem, M.; Siezen, R.J. Reconstruction of the regulatory network of Lactobacillus plantarum WCFS1 on basis of correlated gene expression and conserved regulatory motifs. Microb. Biotechnol. 2011, 4, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M.; Makarevitch, I.; Waters, A.J.; West, P.T.; Stitzer, M.; Hirsch, C.N.; Ross-Ibarra, J.; Springer, N.M. Transposable elements contribute to activation of maize genes in response to abiotic stress. PLoS Genet. 2015, 11. [Google Scholar] [CrossRef]
- Mernke, D.; Dahm, S.; Walker, A.S.; Lalève, A.; Fillinger, S.; Leroch, M.; Hahn, M. Two promoter rearrangements in a drug efflux transporter gene are responsible for the appearance and spread of multidrug resistance phenotype MDR2 in Botrytis cinerea isolates in French and German vineyards. Phytopathology 2011, 101, 1176. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Brendolise, C.; Chagne, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef]
- Mansouri, H.; Asrar, Z.; Mehrabani, M. Effects of gibberellic acid on primary terpenoids and Δ9-tetrahydrocannabinol in Cannabis sativa at flowering stage. J. Integr. Plant Biol. 2009, 51, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Huang, X.H.; Li, L.L.; Deng, G.; Cheng, H.; Rong, X.F.; Li, J.B.; Cheng, S.Y. Molecular cloning and characterization of GbDXS and GbGGPPS gene promoters from Ginkgo biloba. Genet. Mol. Res. 2013, 12, 293–301. [Google Scholar] [CrossRef]
- Sun, J.; Chen, G.X.; Nai-Xing, Y.E.; Shi-Heng, L.; Liu, Z.Q.; Huang, W.; Lin, Z.D. Cloning and expression analysis of deoxyoxylulose-5-phosphate synthase gene related to aroma from Jasminum sambac and isolation of its promoter. Acta Hortic. Sin. 2014, 41, 1236–1244. [Google Scholar]
- Yatagai, F.; Saito, T.; Takahashi, A.; Fujie, A.; Nagaoka, S.; Sato, M.; Ohnishi, T. RPSL mutation induction after space flight on MIR. Mutat. Res. 2000, 453, 1–4. [Google Scholar] [CrossRef]
- Fukuda, T.; Fukuda, K.; Takahashi, A.; Ohnishi, T.; Nakano, T.; Sato, M.; Gunge, N. Analysis of deletion mutations of the rpsL gene in the yeast Saccharomyces cerevisiae detected after long-term flight on the Russian space station MIR. Mutat. Res. 2000, 470, 125–132. [Google Scholar] [CrossRef]
- Ou, X.F.; Long, L.K.; Zhang, Y.H.; Xue, Y.Q.; Liu, J.C.; Lin, X.Y.; Liu, B. Spaceflight induces both transient and heritable alterations in DNA methylation and gene expression in rice (Oryza sativa L.). Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2009, 662, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.A. Fundamental space radiobiology. Gravit. Space Biol. Bull. 2003, 16, 29–36. [Google Scholar] [PubMed]
- Patron, N.J.; Smith, A.M.; Fahy, B.F.; Hylton, C.M.; Naldrett, M.J.; Rossnagel, B.G.; Kay, D. The altered pattern of amylose accumulation in the endosperm of low-amylose barley cultivars is attributable to a single mutant allele of granule-bound starch synthase I with a deletion in the 5′-non-coding region. Plant Physiol. 2002, 130, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Lubliner, S.; Regev, I.; Lotan-Pompan, M.; Edelheit, S.; Weinberger, A.; Segal, E. Core promoter sequence in yeast is a major determinant of expression level. Genome Res. 2015, 25, 1008–1017. [Google Scholar] [CrossRef]
- Riewe, D.; Jeon, H.J.; Lisec, J.; Heuermann, M.C.; Schmeichel, J.; Seyfarth, M.; Meyer, R.C.; Willmitzer, L.; Altmann, T. A naturally occurring promoter polymorphism of the Arabidopsis FUM2 gene causes expression variation, and is associated with metabolic and growth traits. Plant J. 2016, 88, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jiang, Q.T.; Zhao, Q.Z.; Zhao, S.; Lan, X.J.; Dai, S.F.; Lu, Z.X.; Liu, C.J.; Wei, Y.M.; Zheng, Y.L. Characterization and expression analysis of waxy alleles in barley accessions. Genetica 2013, 141, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Midorikawa, T.; Narikawa, R.; Ikeuchi, M. A deletion mutation in the spacing within the psaA core promoter enhances transcription in a cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 2012, 53, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Yan, J.; Yan, H.; Wang, F. Characterization of a strong green tissue-specific motif in rice photosystem I gene promoter Ppsak. Plant Biotechnol. Rep. 2017, 11, 1–9. [Google Scholar] [CrossRef]
- Atanassova, R.; Flenet, M.; Gigot, C.; Chaubet, N. Functional analysis of the promoter region of a maize (Zea mays L.) H3 histone gene in transgenic Arabidopsis thaliana. Plant Mol. Biol. 1998, 37, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Murai, N. A distal promoter region of the rice seed storage protein glutelin gene enhanced quantitative gene expression. Plant Sci. 1997, 128, 59–65. [Google Scholar] [CrossRef]
- Sibéril, Y.; Doireau, P.; Gantet, P. Plant bZIP G-box binding factors. Eur. J. Biochem. 2001, 268, 5655–5666. [Google Scholar] [CrossRef] [PubMed]
- Chenge-Espinosa, M.; Cordoba, E.; Romero-Guido, C.; Toledo-Ortiz, G.; Leon, P. Shedding light on the methylerythritol phosphate (MEP)-pathway: Long hypocotyl 5 (HY5)/phytochrome-interacting factors (PIFs) transcription factors modulating key limiting steps. Plant J. 2018, 96, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.; Rohwer, J.M.; Ghirardo, A.; Hammerbacher, A.; Ortíz, M.; Raguschke, B.; Schnitzler, J.-P.; Gershenzon, J.; Phillips, M.A. 1-Deoxyxylulose 5-phosphate synthase controls flux through the 2-C-methylerythritol 4-phosphate pathway in Arabidopsis thaliana. Plant Physiol. 2014, 165, 1488–1504. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-G.; Chen, Y. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–650, 652, 654 passim. [Google Scholar] [CrossRef] [PubMed]
- Mathelier, A.; Fornes, O.; Arenillas, D.J.; Chen, C.Y.; Denay, G.; Lee, J.; Shi, W.; Shyr, C.; Tan, G.; Worsley-Hunt, R. JASPAR 2016: A major expansion and update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2016, 44, D110–D115. [Google Scholar] [CrossRef]
- Junker, B.; Zimny, J.; Lührs, R.; Lörz, H. Transient expression of chimaeric genes in dividing and non-dividing cereal protoplasts after PEG-induced DNA uptake. Plant Cell Rep. 1987, 6, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, J.; Duan, S.; Ao, Y.; Dai, J.; Liu, J.; Wang, P.; Li, Y.; Liu, B.; Feng, D.; et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods 2011, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.D.; Lemaux, P.G.; Cho, M.-J. Stable transformation of a recalcitrant Kentucky bluegrass (Poa pratensis L.) cultivar using mature seed-derived highly regenerative tissues. In Vitro Cell. Dev. Biol.-Plant 2001, 37, 6–11. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) |
|---|---|
| SP1 | CCGTCGGTCTGCCGCATCGTC |
| SP2 | ACGATGGACTCCAGTCGCTTGTCCTGAGGGGTGTTG |
| SP3 | TGAGGGACAGGTTCTTCATGTGGA |
| SP4 | CCTCTCCGTCAGCGACGCAGAAA |
| SP5 | ACGATGGACTCCAGATCTGGGGAGTGGAGCGGGA |
| SP6 | CGTGTGGCTGGTATTCGTTCG |
| SP7 | TCGGGACAGGGAGAGCGAAATGG |
| SP8 | ACGATGGACTCCAGAAAAGAGCAAAGGGGAACCG |
| SP9-WT | GAACAAAGTGGAATGTCGGCG |
| SP9-A16 | TGTGGAGGAGGTGGTGGTGCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, L.; Chao, Y.; Su, H.; Ren, Y.; Yin, S.; Han, L. Altered Promoter and G-Box Binding Factor for 1-Deoxy-d-Xylulose-5-Phosphate Synthase Gene Grown from Poa pratensis Seeds after Spaceflight. Int. J. Mol. Sci. 2019, 20, 1398. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061398
Gan L, Chao Y, Su H, Ren Y, Yin S, Han L. Altered Promoter and G-Box Binding Factor for 1-Deoxy-d-Xylulose-5-Phosphate Synthase Gene Grown from Poa pratensis Seeds after Spaceflight. International Journal of Molecular Sciences. 2019; 20(6):1398. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061398
Chicago/Turabian StyleGan, Lu, Yuehui Chao, Haotian Su, Yujing Ren, Shuxia Yin, and Liebao Han. 2019. "Altered Promoter and G-Box Binding Factor for 1-Deoxy-d-Xylulose-5-Phosphate Synthase Gene Grown from Poa pratensis Seeds after Spaceflight" International Journal of Molecular Sciences 20, no. 6: 1398. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061398