Symmetrical Modification of Minimized Dermaseptins to Extend the Spectrum of Antimicrobials with Endotoxin Neutralization Potency

Abstract
:1. Introduction
2. Results and Discussion
2.1. Peptide Design and Characterization
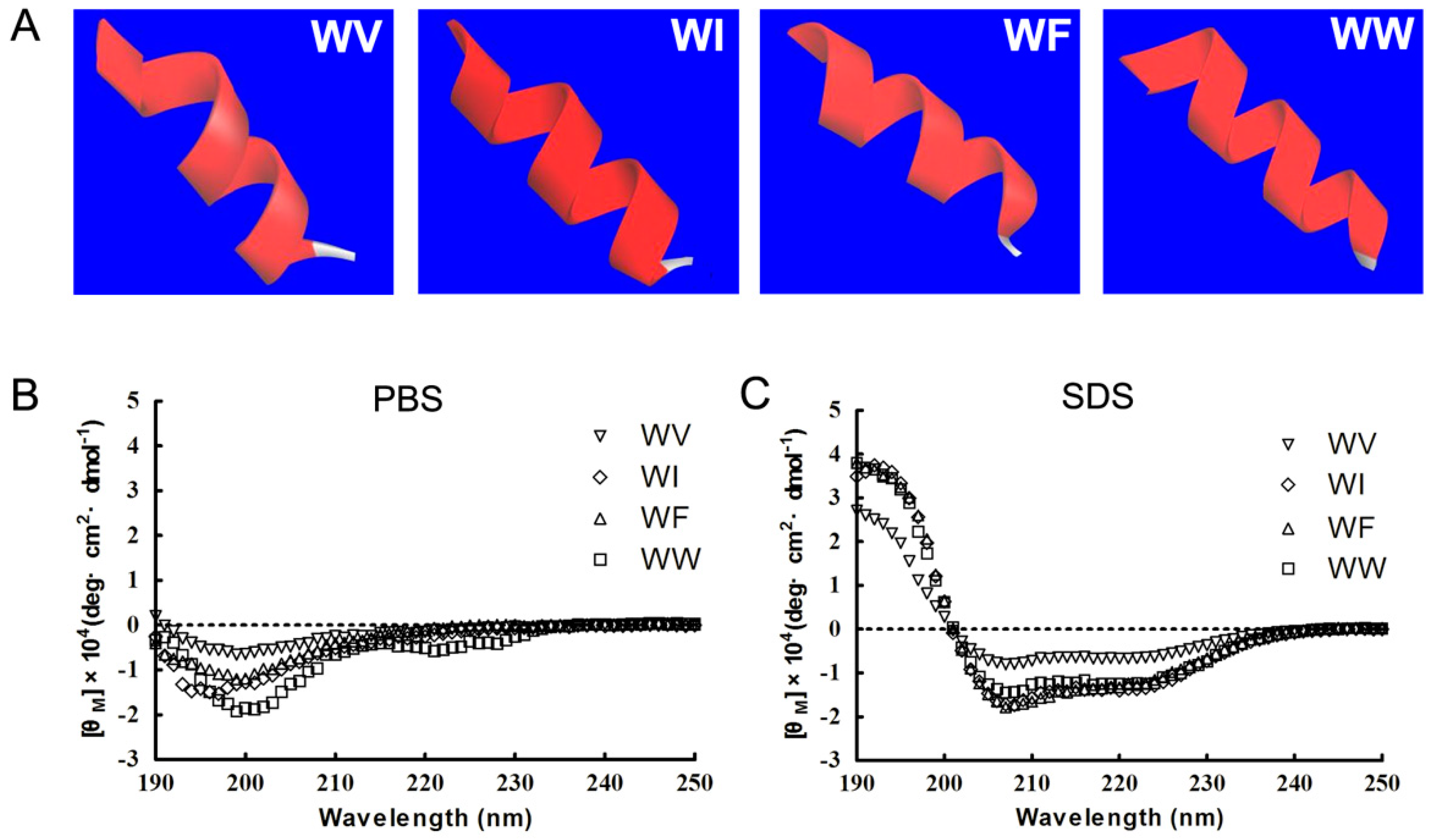
2.2. Structure Variability of the Peptides
2.3. Antimicrobial Activity
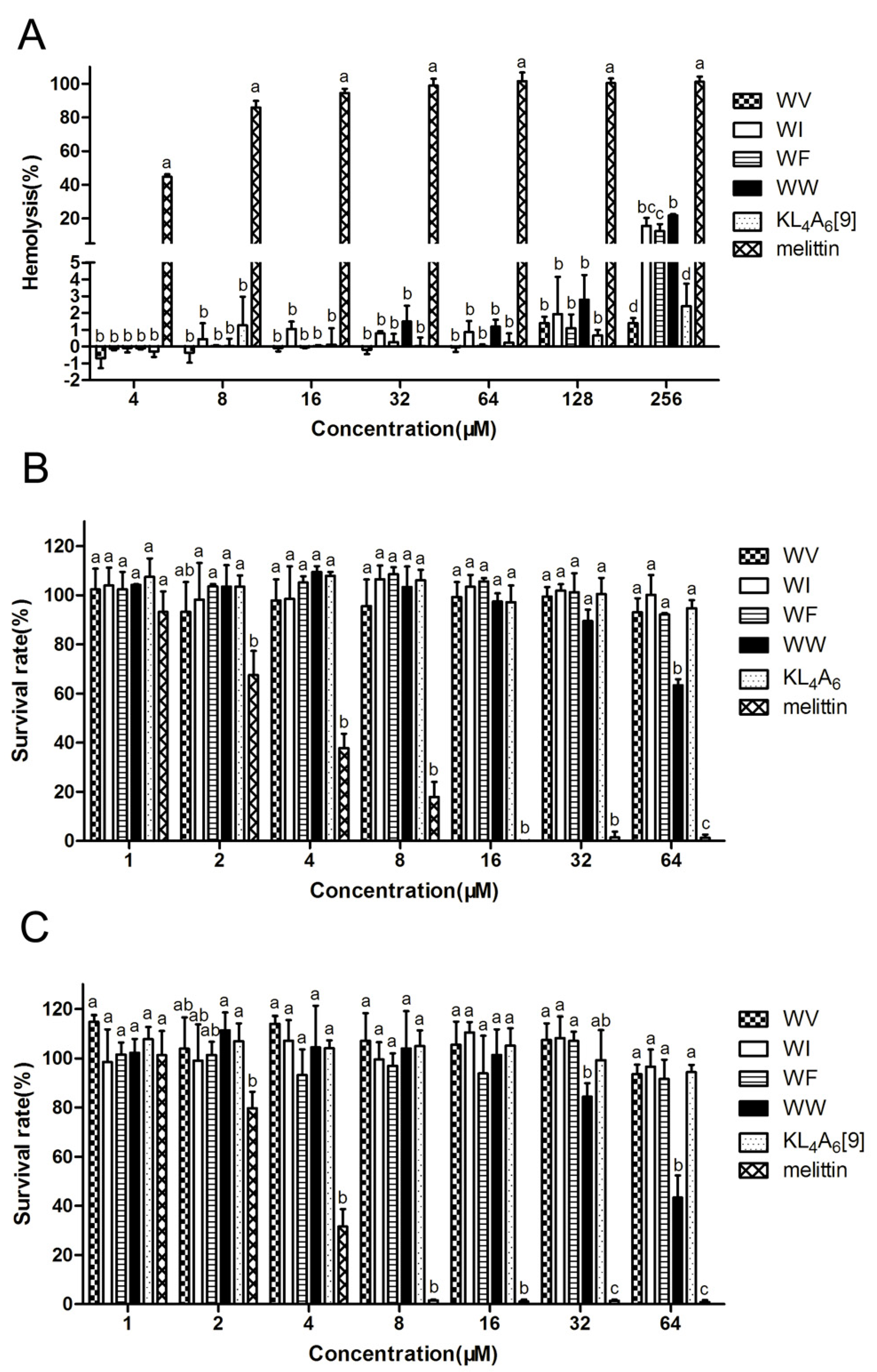
2.4. Cytotoxicity and Therapeutic Index
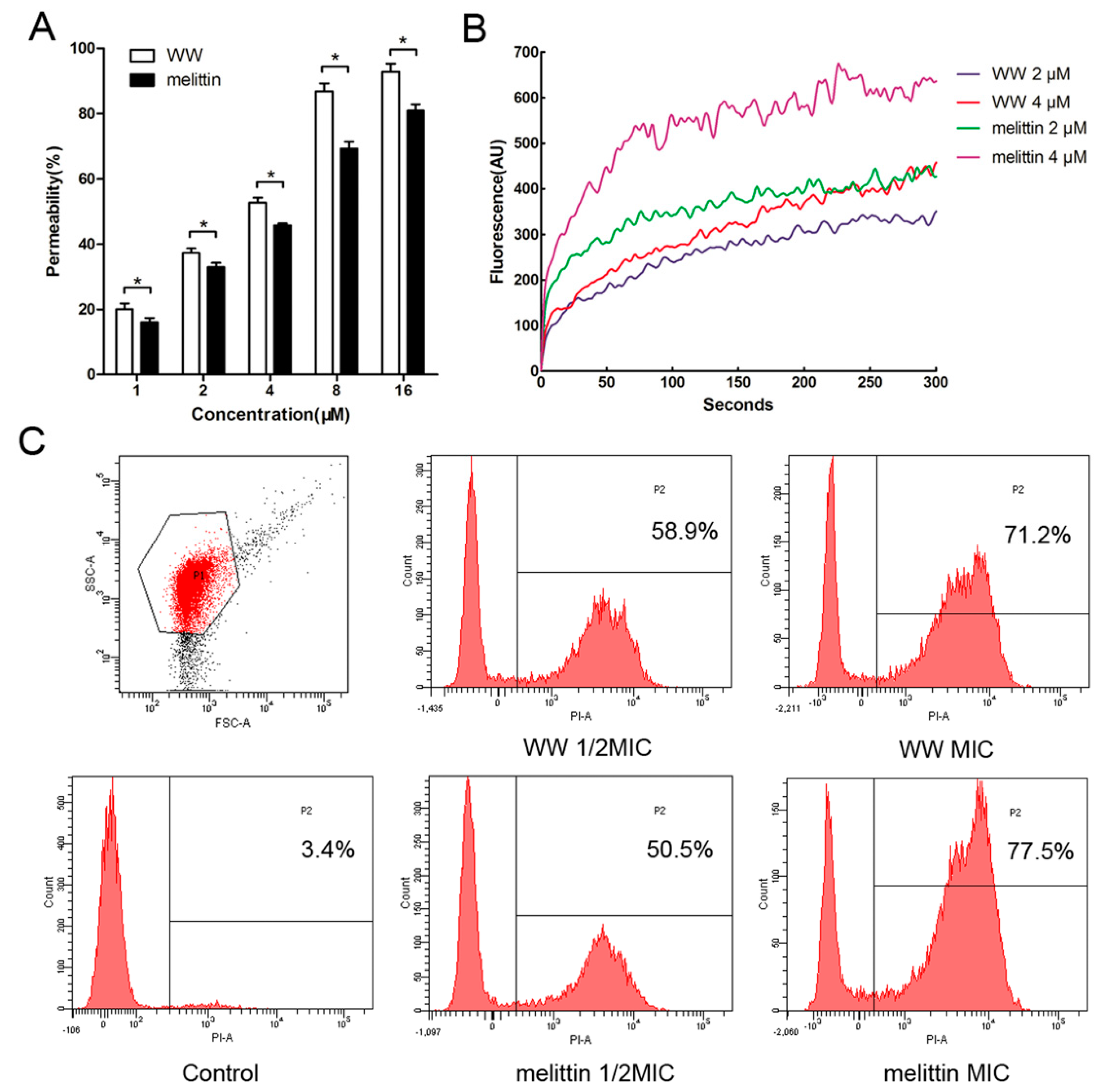
2.5. Mechanism of Action of the Peptides
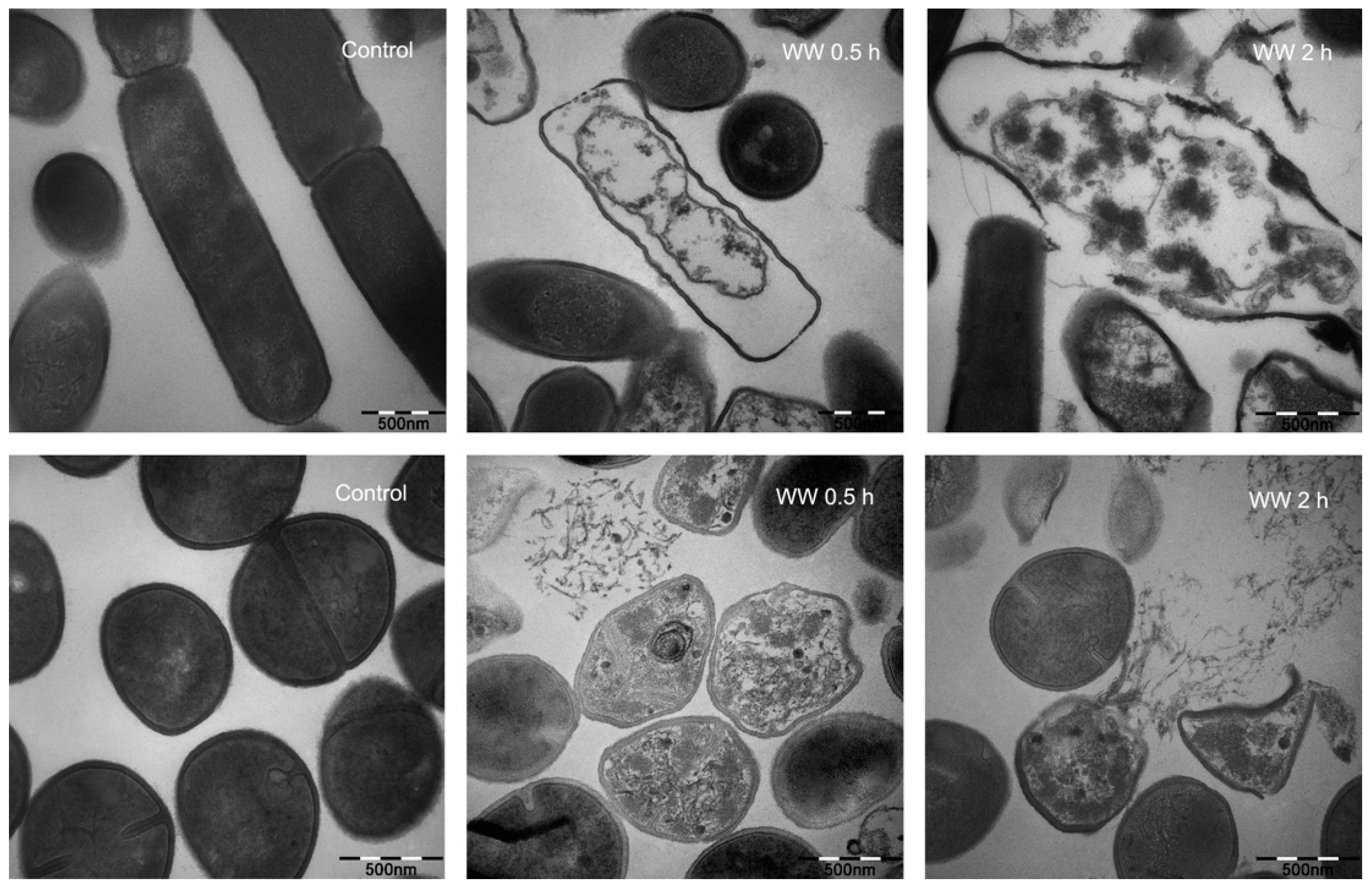
2.6. Membrane Morphological Analysis
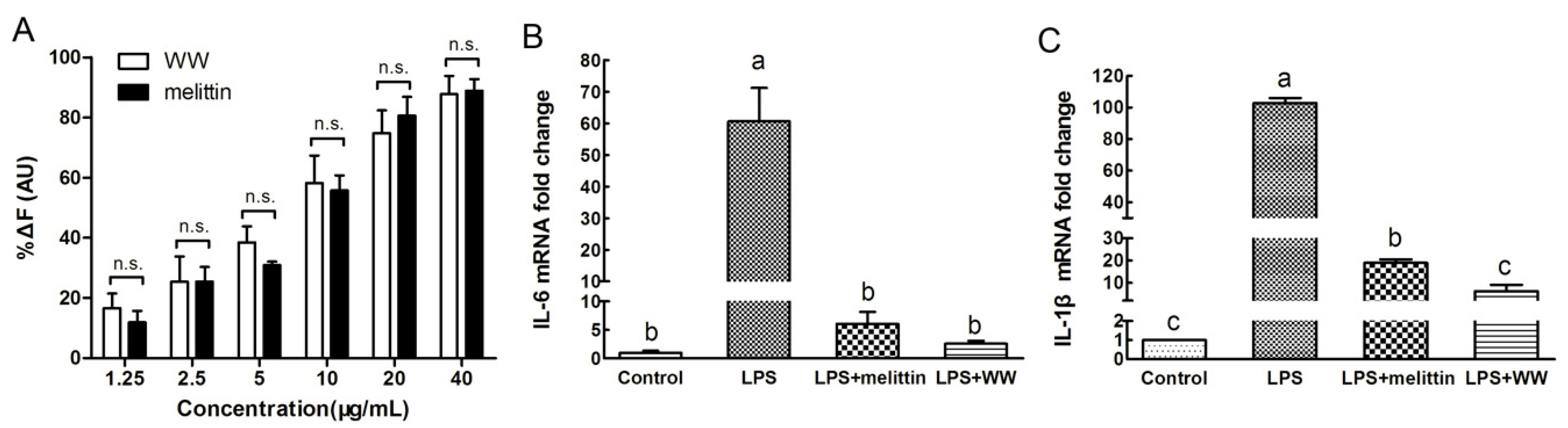
2.7. Endotoxin Neutralization Assay
3. Materials and Methods
3.1. Bacterial Strains and Mammalian Cells
3.2. Peptides Synthesis and Sequence Analysis
3.3. CD Spectroscopy
3.4. MIC Measurements
3.5. Hemolytic Activity Assay
3.6. Cytotoxicity Assay
3.7. Outer Membrane (OM) Permeability Assay
3.8. Cytoplasmic Membrane Depolarization Assay
3.9. FACScan Analysis
3.10. Membrane Morphological Observations
3.11. LPS Neutralization Assay
3.12. Cytokine Measurements
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sommer, M.O.A.; Munck, C.; Toft-Kehler, R.V.; Andersson, D.I. Prediction of antibiotic resistance: Time for a new preclinical paradigm? Nat. Rev. Microbiol. 2017, 15, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- Omardien, S.; Brul, S.; Zaat, S.A. Antimicrobial activity of cationic antimicrobial peptides against gram-positives: Current progress made in understanding the mode of action and the response of bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Gao, S.J.; Yang, Y.Y. Short syntheticβ-sheet forming peptide amphiphiles as broad spectrum antimicrobials with antibiofilm and endotoxin neutralizing capabilities. Adv. Funct. Mater. 2013, 23, 3682–3692. [Google Scholar] [CrossRef]
- Ong, Z.Y.; Wiradharma, N.; Yang, Y.Y. Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv. Drug Deliv. Rev. 2014, 78, 28–45. [Google Scholar] [CrossRef]
- Ma, Z.; Wei, D.; Yan, P.; Zhu, X.; Shan, A.; Bi, Z. Characterization of cell selectivity, physiological stability and endotoxin neutralization capabilities of alpha-helix-based peptide amphiphiles. Biomaterials 2015, 52, 517–530. [Google Scholar] [CrossRef]
- Lyu, Y.; Yang, Y.; Lyu, X.; Na, D.; Shan, A. Antimicrobial activity, improved cell selectivity and mode of action of short PMAP-36-derived peptides against bacteria and Candida. Sci. Rep. 2016, 6, 27258. [Google Scholar] [CrossRef]
- Dong, N.; Zhu, X.; Chou, S.; Shan, A.; Li, W.; Jiang, J. Antimicrobial potency and selectivity of simplified symmetric-end peptides. Biomaterials 2014, 35, 8028–8039. [Google Scholar] [CrossRef]
- Li, W.; Tan, T.; Xu, W.; Xu, L.; Dong, N.; Ma, D.; Shan, A. Rational design of mirror-like peptides with alanine regulation. Amino Acids 2016, 48, 403–417. [Google Scholar] [CrossRef]
- Fimland, G.; Eijsink, V.G.H.; Nissen-Meyer, J. Mutational analysis of the role of tryptophan residues in an antimicrobial peptide. Biochemistry 2002, 41, 9508–9515. [Google Scholar] [CrossRef]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan-and arginine-rich antimicrobial peptides: Structures and mechanisms of action. Biochim. Et Biophys. Acta (Bba) Biomembr. 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.T.; Shin, S.Y.; Hahm, K.S.; Kim, J.I. Design of perfectly symmetric Trp-rich peptides with potent and broad-spectrum antimicrobial activities. Int. J. Antimicrob. Agents 2006, 27, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Sung-Tae, Y.; Yub, S.S.; Jae Il, K. Interaction mode of a symmetric Trp-rich undeca peptide PST11-RK with lipid bilayers. Febs Lett. 2007, 581, 157–163. [Google Scholar]
- Podorieszach, A.P.; Huttunen-Hennelly, H.E. The effects of tryptophan and hydrophobicity on the structure and bioactivity of novel indolicidin derivatives with promising pharmaceutical potential. Org. Biomol. Chem. 2010, 8, 1679–1687. [Google Scholar] [CrossRef]
- Papanastasiou, E.A.; Hua, Q.; Sandouk, A.; Son, U.H.; Christenson, A.J.; Van Hoek, M.L.; Bishop, B.M. Role of acetylation and charge in antimicrobial peptides based on human β-defensin-3. Apmis 2009, 117, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, B.C.; Wells, J.A. Minimized proteins. Curr. Opin. Struct. Biol. 1997, 7, 457–462. [Google Scholar] [CrossRef]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural determinants of host defense peptides for antimicrobial activity and target cell selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef]
- Reisser, S.; Strandberg, E.; Steinbrecher, T.; Ulrich, A.S. 3D hydrophobic moment vectors as a tool to characterize the surface polarity of amphiphilic peptides. Biophys. J. 2014, 106, 2385–2394. [Google Scholar] [CrossRef]
- Wiradharma, N.; Khoe, U.; Hauser, C.A.; Seow, S.V.; Zhang, S.; Yang, Y.-Y. Synthetic cationic amphiphilic α-helical peptides as antimicrobial agents. Biomaterials 2011, 32, 2204–2212. [Google Scholar] [CrossRef]
- Mecke, A.; Lee, D.-K.; Ramamoorthy, A.; Orr, B.G.; Holl, M.M.B. Membrane thinning due to antimicrobial peptide binding: An atomic force microscopy study of MSI-78 in lipid bilayers. Biophys. J. 2005, 89, 4043–4050. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, C.; Liang, G.; Zhang, M.; Zheng, J. Engineering antimicrobial peptides with improved antimicrobial and hemolytic activities. J. Chem. Inf. Modeling 2013, 53, 3280–3296. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Et Biophys. Acta (Bba) Biomembr. 2016, 1858, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, J.; Gao, H.; Wang, Z.; Dong, N.; Ma, Q.; Shan, A. Antimicrobial properties and membrane-active mechanism of a potential alpha-helical antimicrobial derived from cathelicidin PMAP-36. PLoS ONE 2014, 9, e86364. [Google Scholar]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Zong, X.; Song, D.; Wang, T.; Xia, X.; Hu, W.; Han, F.; Wang, Y. LFP-20, a porcine lactoferrin peptide, ameliorates LPS-induced inflammation via the MyD88/NF-kappaB and MyD88/MAPK signaling pathways. Dev. Comp. Immunol. 2015, 52, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Yoo, H.; Kim, J.A.; Lee, S.; Jee, J.G.; Lee, M.Y.; Lee, Y.M.; Bae, J.S. Barrier protective effects of piperlonguminine in LPS-induced inflammation in vitro and in vivo. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 58, 149–157. [Google Scholar] [CrossRef] [PubMed]
- AMP Database. Available online: http://aps.unmc.edu/AP/main.php (accessed on 25 January 2019).
- I-TASSER. Available online: http://zhanglab.ccmb.med.umich.edu/I-TASSER/ (accessed on 12 December 2018).
- 3D-HM Calculator. Available online: http://www.ibg.kit.edu/HM/ (accessed on 25 January 2019).
- Shao, C.; Tian, H.; Wang, T.; Wang, Z.; Chou, S.; Shan, A.; Cheng, B. Central beta-turn increases the cell selectivity of imperfectly amphipathic alpha-helical peptides. Acta Biomater. 2018, 69, 243–255. [Google Scholar] [CrossRef]
- Tan, T.; Wu, D.; Li, W.; Zheng, X.; Li, W.; Shan, A. High Specific selectivity and membrane-active mechanism of synthetic cationic hybrid antimicrobial peptides based on the peptide FV7. Int. J. Mol. Sci. 2017, 18, 339. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Sequences | Molecular Mass | Net Charge | 3D Hydrophobic Moment Vectors (20.0) 1 | ||
|---|---|---|---|---|---|---|
| Calculated | Measured | Length (kTÅ/e) | Angle (°) | |||
| KL4A6 [9] | LLKAAAKAAAKLL-NH2 | 1280.69 | 1280.68 | +4 | 18.94 | 109.81 |
| WV | WVKAAAKAAAKVW-NH2 | 1398.74 | 1398.73 | +4 | 36.49 | 30.87 |
| WI | WIKAAAKAAAKIW-NH2 | 1426.79 | 1426.78 | +4 | 30.30 | 141.05 |
| WF | WFKAAAKAAAKFW-NH2 | 1494.83 | 1494.82 | +4 | 35.79 | 86.60 |
| WW | WWKAAAKAAAKWW-NH2 | 1572.87 | 1572.86 | +4 | 47.40 | 84.50 |
| MIC (μM) | KL4A6 [9] | WV | WI | WF | WW | Melittin |
|---|---|---|---|---|---|---|
| Gram-negative bacteria | ||||||
| E. coli ATCC25922 | 2 | 32 | 8 | 4 | 2 | 1 |
| E. coli UB1005 | 32 | >128 | 8 | 4 | 4 | 2 |
| P. aeruginosa ATCC 27853 | >128 | >128 | 32 | 16 | 8 | 2 |
| S. typhimurium ATCC 14028 | 128 | >128 | 32 | 32 | 8 | 4 |
| S. pullorum C79-13 | >128 | 64 | 8 | 8 | 2 | 1 |
| Gram-positive bacteria | ||||||
| S. aureus ATCC 29213 | >128 | >128 | 32 | 8 | 4 | 1 |
| S. epidermidis ATCC 12228 | 128 | >128 | 32 | 16 | 4 | 0.5 |
| S. faecalis ATCC 29212 | 16 | 64 | 32 | 32 | 4 | 1 |
| B. subtilis CMCC 63501 | 32 | >128 | 8 | 8 | 4 | 1 |
| MHC5(μM) 1 | >256 | >256 | 256 | 256 | 256 | 0.25 |
| GM(μM) 2 | 59.3 | 149.3 | 17.3 | 10.9 | 4.0 | 1.3 |
| Therapeutic index 3 | 8.6 | 3.4 | 14.8 | 23.5 | 64.0 | 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, C.; Li, W.; Tan, P.; Shan, A.; Dou, X.; Ma, D.; Liu, C. Symmetrical Modification of Minimized Dermaseptins to Extend the Spectrum of Antimicrobials with Endotoxin Neutralization Potency. Int. J. Mol. Sci. 2019, 20, 1417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061417
Shao C, Li W, Tan P, Shan A, Dou X, Ma D, Liu C. Symmetrical Modification of Minimized Dermaseptins to Extend the Spectrum of Antimicrobials with Endotoxin Neutralization Potency. International Journal of Molecular Sciences. 2019; 20(6):1417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061417
Chicago/Turabian StyleShao, Changxuan, Weizhong Li, Peng Tan, Anshan Shan, Xiujing Dou, Deying Ma, and Chunyu Liu. 2019. "Symmetrical Modification of Minimized Dermaseptins to Extend the Spectrum of Antimicrobials with Endotoxin Neutralization Potency" International Journal of Molecular Sciences 20, no. 6: 1417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061417