Genome-Wide Analysis of Glycoside Hydrolase Family 1 β-glucosidase Genes in Brassica rapa and Their Potential Role in Pollen Development


Abstract
:1. Introduction
2. Results
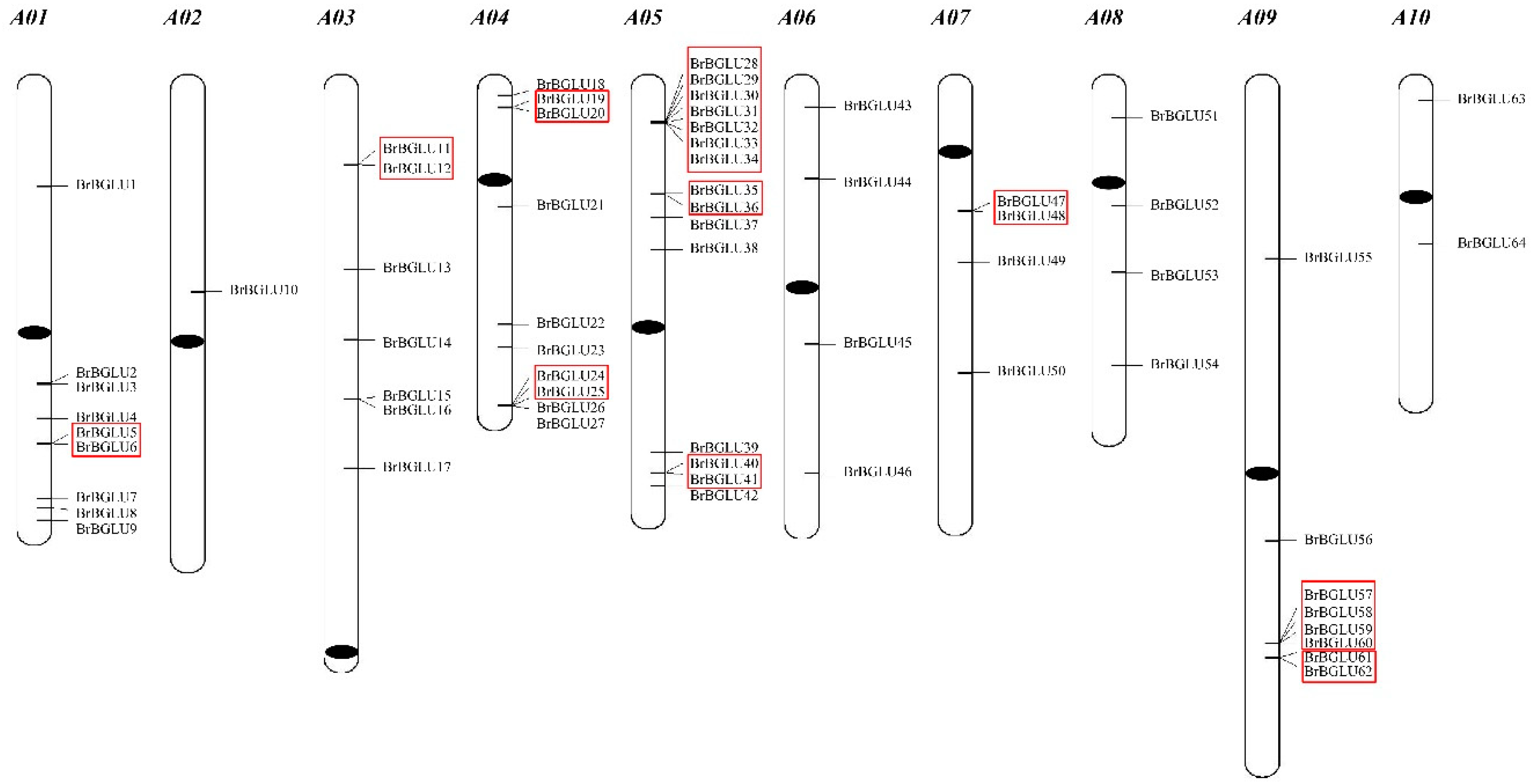
2.1. Identification and Chromosomal Distribution of BrBGLUs
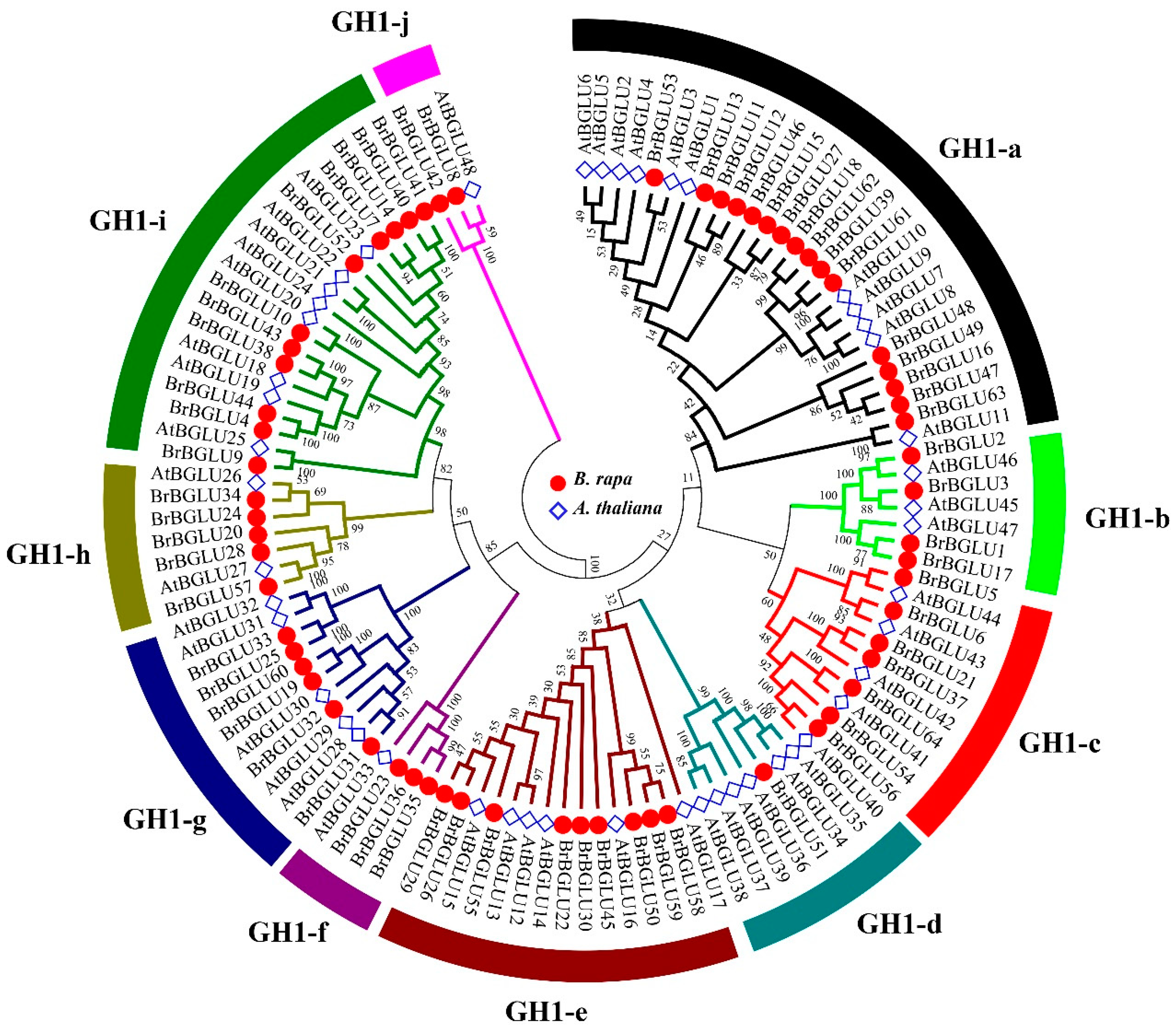
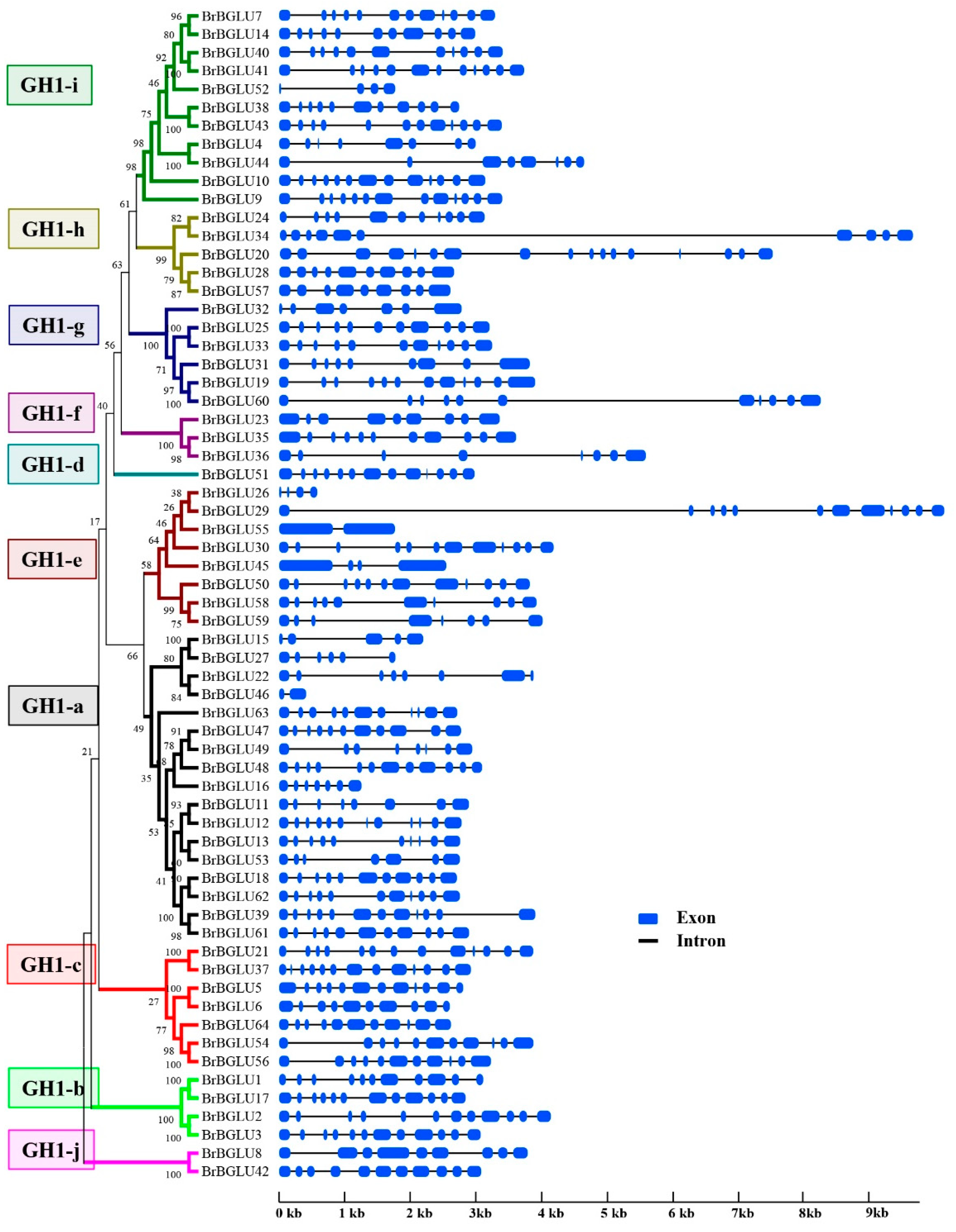
2.2. Phylogenetic and Gene Structure Analysis of BrBGLUs
2.3. Identification of BrBGLU Genes Involved in Pollen Development
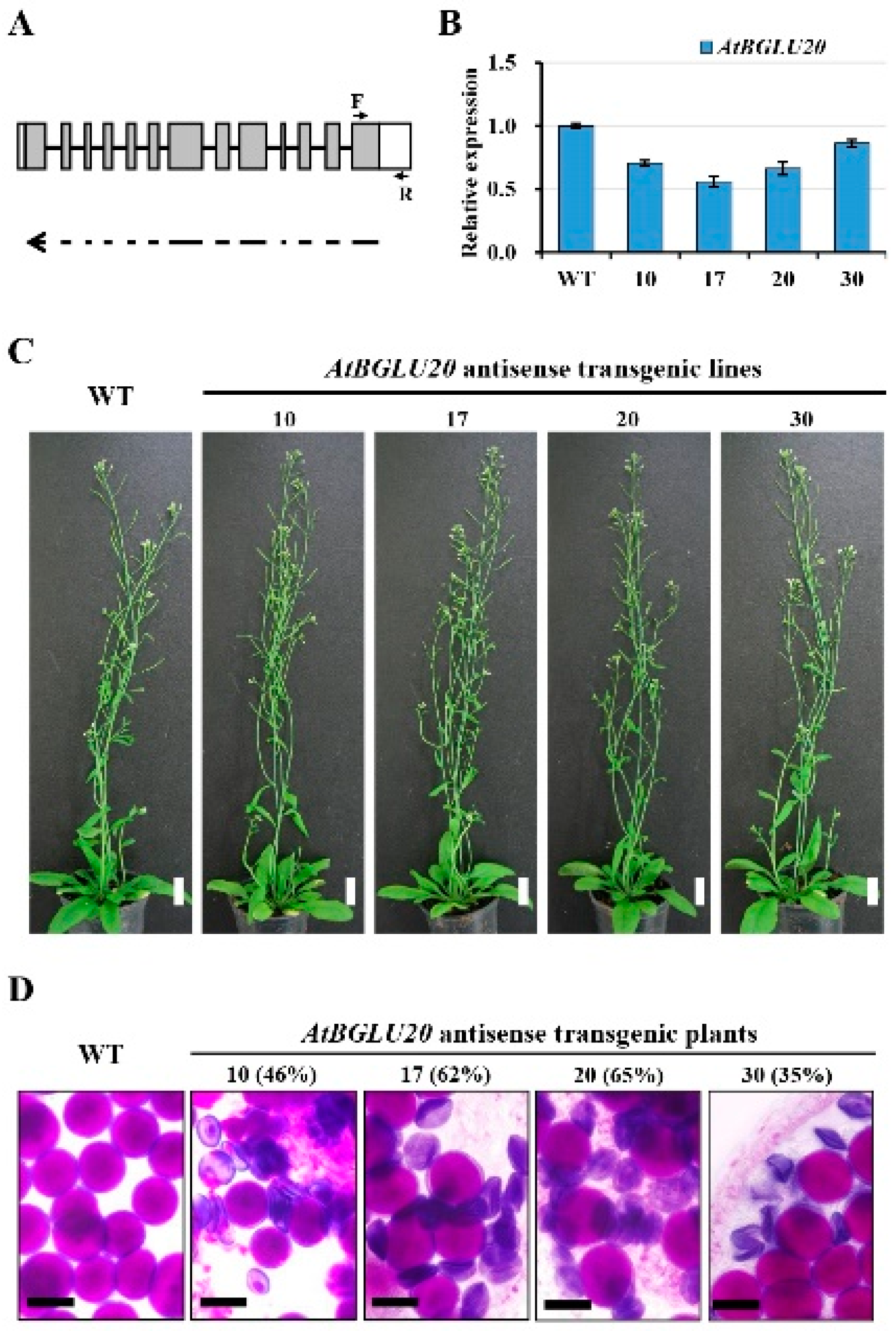
2.4. Analysis of the Putative Functions of BrBGLU10/AtBGLU20 in Pollen Development
3. Discussion
3.1. Identification and Analysis of BrBGLUs
3.2. The Potential Functions of BrBGLUs During Pollen Development
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Antisense Constructs and Plant Transformation
4.3. Reverse Transcription PCR and qRT-PCR
4.4. Pollen Viability
4.5. Identification of BrBGLUs and Phylogenetic Tree Construction
4.6. Chromosomal Location, Nomenclature, and Gene Duplication of BrBGLUs
4.7. Co-Expression and Gene Ontology Enrichment Analysis
4.8. Microarray Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| GH1 | Glycoside hydrolase family 1 |
| BGLUs | β-glycosidase genes |
| BrBGLUs | Brassica rapa β-glycosidase genes |
| ABA | abscisic acid |
| OsTDR | Tapetum Degeneration Retardation |
| HMM | Hidden Markov Model |
| GO | Gene Ontology |
| CDS | coding sequence |
References
- Chandrasekar, B.; Colby, T.; Emran Khan Emon, A.; Jiang, J.; Hong, T.N.; Villamor, J.G.; Harzen, A.; Overkleeft, H.S.; van der Hoorn, R.A. Broad-range glycosidase activity profiling. Mol. Cell. Proteomics 2014, 13, 2787–2800. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The carbohydrate-active enzymes database (cazy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (cazy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991, 280, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opassiri, R.; Pomthong, B.; Onkoksoong, T.; Akiyama, T.; Esen, A.; Ketudat Cairns, J.R. Analysis of rice glycosyl hydrolase family 1 and expression of os4bglu12 beta-glucosidase. BMC Plant Biol. 2006, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Escamilla-Trevino, L.; Zeng, L.; Lalgondar, M.; Bevan, D.; Winkel, B.; Mohamed, A.; Cheng, C.L.; Shih, M.C.; Poulton, J.; et al. Functional genomic analysis of arabidopsis thaliana glycoside hydrolase family 1. Plant Mol. Biol. 2004, 55, 343–367. [Google Scholar] [CrossRef]
- Gomez-Anduro, G.; Ceniceros-Ojeda, E.A.; Casados-Vazquez, L.E.; Bencivenni, C.; Sierra-Beltran, A.; Murillo-Amador, B.; Tiessen, A. Genome-wide analysis of the beta-glucosidase gene family in maize (zea mays l. Var b73). Plant Mol. Biol. 2011, 77, 159–183. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, T.; An, X.; Gu, R. Evolution and expression analysis of the β-glucosidase (glu) encoding gene subfamily in maize. Genes Genom. 2012, 34, 179–187. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Yang, J.F.; Liu, T.Y.; Su, Z.F.; Zhu, F.Y.; Chen, M.X.; Fan, T.; Ye, N.H.; Feng, Z.; Wang, L.J.; et al. A phylogenetically informed comparison of gh1 hydrolases between arabidopsis and rice response to stressors. Front Plant Sci. 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Roepke, J.; Bozzo, G.G. Arabidopsis thaliana β-glucosidase bglu15 attacks flavonol 3-O-β-glucoside-7-O-α-rhamnosides. Phytochemistry 2015, 109, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, H.; Tohge, T.; Viehöver, P.; Fernie, A.R.; Weisshaar, B.; Stracke, R. Natural variation in flavonol accumulation in arabidopsis is determined by the flavonol glucosyltransferase bglu6. J. Exp. Bot. 2016, 67, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, T.; Sakiyama, R.; Ozeki, Y.; Sasaki, N. Acyl-glucose-dependent glucosyltransferase catalyzes the final step of anthocyanin formation in arabidopsis. J. Plant Physiol. 2013, 170, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Roepke, J.; Gordon, H.O.W.; Neil, K.J.A.; Gidda, S.; Mullen, R.T.; Freixas Coutin, J.A.; Bray-Stone, D.; Bozzo, G.G. An apoplastic β-glucosidase is essential for the degradation of flavonol 3-O-β-glucoside-7-O-α-rhamnosides in arabidopsis. Plant Cell Physiol. 2017, 58, 1030–1047. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Jander, G. Arabidopsis myrosinases tgg1 and tgg2 have redundant function in glucosinolate breakdown and insect defense. Plant J. 2006, 46, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Burow, M. Glucosinolate breakdown in arabidopsis: Mechanism, regulation and biological significance. Arabidopsis Book 2010, 8, e0134. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Tokuhisa, J.G.; Bevan, D.R.; Esen, A. Properties of β-thioglucoside hydrolases (tgg1 and tgg2) from leaves of arabidopsis thaliana. Plant Sci. 2012, 191–192, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Hai, L.P.; Kim, H.Y.; Sang, M.C.; Fan, J.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.J.; Hwang, I. Activation of glucosidase via stress-induced polymerization rapidly increases active pools of abscisic acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.Y.; Lee, K.H.; Dong, T.; Jeong, J.C.; Jin, J.B.; Kanno, Y.; Kim, D.H.; Kim, S.Y.; Seo, M.; Bressan, R.A.; et al. A vacuolar beta-glucosidase homolog that possesses glucose-conjugated abscisic acid hydrolyzing activity plays an important role in osmotic stress responses in arabidopsis. Plant Cell 2012, 24, 2184–2199. [Google Scholar] [CrossRef] [PubMed]
- Costet, L.; Fritig, B.; Kauffmann, S. Scopoletin expression in elicitor-treated and tobacco mosaic virus-infected tobacco plants. Physiol. Plant 2002, 115, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Fabienne, B.; Patrice, D.R.; Serge, K. Molecular cloning and biological activity of alpha-, beta-, and gamma-megaspermin, three elicitins secreted by phytophthora megasperma h20. Plant Physiol. 2003, 131, 155–166. [Google Scholar]
- Hino, F.; Okazaki, M.; Miura, Y. Effect of 2,4-dichlorophenoxyacetic acid on glucosylation of scopoletin to scopolin in tobacco tissue culture. Plant Physiol. 1982, 69, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.O.; Shimizu, B.; Sakata, K.; Gantulga, D.; Zhou, Z.; Bevan, D.R.; Esen, A. Scopolin hydrolyzing beta-glucosidases in roots of arabidopsis. Plant Cell Physiol. 2010, 51, 132. [Google Scholar] [CrossRef] [PubMed]
- Chapelle, A.; Morreel, K.; Vanholme, R.; Le-Bris, P.; Morin, H.; Lapierre, C.; Boerjan, W.; Jouanin, L.; Demont-Caulet, N. Impact of the absence of stem-specific beta-glucosidases on lignin and monolignols. Plant Physiol. 2012, 160, 1204–1217. [Google Scholar] [CrossRef] [PubMed]
- Zamioudis, C.; Hanson, J.; Pieterse, C.M. Beta-glucosidase bglu42 is a myb72-dependent key regulator of rhizobacteria-induced systemic resistance and modulates iron deficiency responses in arabidopsis roots. New Phytol. 2014, 204, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Piffanelli, P.; Ross, J.H.E.; Murphy, D.J. Biogenesis and function of the lipidic structures of pollen grains. Sex. Plant Reprod. 1998, 11, 65–80. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, Z.; Cao, J. Pollen wall development: The associated enzymes and metabolic pathways. Plant Biol. 2013, 15, 249–263. [Google Scholar] [CrossRef]
- Minic, Z.; Jouanin, L. Plant glycoside hydrolases involved in cell wall polysaccharide degradation. Plant Physiol. Biochem. 2006, 44, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.S.; Liang, W.Q.; Yuan, Z.; Li, N.; Shi, J.; Wang, J.; Liu, Y.M.; Yu, W.J.; Zhang, D.B. Tapetum degeneration retardation is critical for aliphatic metabolism and gene regulation during rice pollen development. Mol. Plant 2008, 1, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Feng, H.; Xu, M.; Lee, J.; Kim, Y.K.; Lim, Y.P.; Piao, Z.; Park, Y.D.; Ma, H.; Hur, Y. Comprehensive analysis of genic male sterility-related genes in brassica rapa using a newly developed br300k oligomeric chip. PLoS ONE 2013, 8, e72178. [Google Scholar] [CrossRef]
- Singh, A.K.; Sharma, V.; Pal, A.K.; Acharya, V.; Ahuja, P.S. Genome-wide organization and expression profiling of the nac transcription factor family in potato (Solanum tuberosum L.). DNA Res. 2013, 20, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal w and clustal x version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Thorlby, G.; Fourrier, N.; Warren, G. The sensitive to freezing2 gene, required for freezing tolerance in arabidopsis thaliana, encodes a beta-glucosidase. Plant Cell 2004, 16, 2192–2203. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. Gsds 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Ng, C.Y.; Wickneswari, R.; Choong, C.Y. Identification of floral genes for sex determination in Calamus palustris griff. By using suppression subtractive hybridization. Genet. Mol. Res. 2014, 13, 6037–6049. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef]
- Rubinelli, P.; Hu, Y.; Ma, H. Identification, sequence analysis and expression studies of novel anther-specific genes of Arabidopsis thaliana. Plant Mol. Biol. 1998, 37, 607–619. [Google Scholar] [CrossRef]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “electronic fluorescent pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Smyth, D.R.; Bowman, J.L.; Meyerowitz, E.M. Early flower development in arabidopsis. Plant Cell 1990, 2, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Ma, H. Molecular genetic analyses of microsporogenesis and microgametogenesis in flowering plants. Annu. Rev. Plant Biol. 2005, 56, 393–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, H.; Hua, H.; Wang, L.; Song, C.-P. A vacuole localized β-glucosidase contributes to drought tolerance in arabidopsis. Chin. Sci. Bull. 2011, 56, 3538–3546. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of nac transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Fourrier, N.; Bedard, J.; Lopez-Juez, E.; Barbrook, A.; Bowyer, J.; Jarvis, P.; Warren, G.; Thorlby, G. A role for sensitive to freezing2 in protecting chloroplasts against freeze-induced damage in arabidopsis. Plant J. 2008, 55, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Moellering, E.R.; Bagyalakshmi, M.; Christoph, B. Freezing tolerance in plants requires lipid remodeling at the outer chloroplast membrane. Science 2010, 330, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Andreasson, E.; Bolt Jorgensen, L.; Hoglund, A.S.; Rask, L.; Meijer, J. Different myrosinase and idioblast distribution in arabidopsis and brassica napus. Plant Physiol. 2001, 127, 1750–1763. [Google Scholar] [CrossRef] [PubMed]
- Talalay, P.; Fahey, J.W. Phytochemicals from cruciferous plants protect against cancer by modulating carcinogen metabolism. J. Nutr. 2001, 131, 3027S–3033S. [Google Scholar] [CrossRef]
- Jeffery, E.H.; Araya, M. Physiological effects of broccoli consumption. Phytochem. Rev. 2009, 8, 283–298. [Google Scholar] [CrossRef]
- Borpatragohain, P.; Rose, T.J.; King, G.J. Fire and brimstone: Molecular interactions between sulfur and glucosinolate biosynthesis in model and crop brassicaceae. Front. Plant Sci. 2016, 7, 1735. [Google Scholar] [CrossRef] [PubMed]
- Baskar, V.; Gururani, M.A.; Yu, J.W.; Park, S.W. Engineering glucosinolates in plants: Current knowledge and potential uses. Appl. Biochem. Biotechnol. 2012, 168, 1694–1717. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.M.; Jeffery, E.H.; Juvik, J.A. Proposed method for estimating health-promoting glucosinolates and hydrolysis products in broccoli (Brassica oleracea var. Italica) using relative transcript abundance. J. Agric. Food Chem. 2017, 65, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Shimizu, B.; Mizutani, M.; Watanabe, K.; Sakata, K. Accumulation of coumarins in arabidopsis thaliana. Phytochemistry 2006, 67, 379–386. [Google Scholar] [CrossRef]
- Baiya, S.; Mahong, B.; Lee, S.; Jeon, J.S.; Cairns, J.R.K. Demonstration of monolignol β-glucosidase activity of rice os4bglu14, os4bglu16 and os4bglu18 in arabidopsis thaliana bglu 45 mutant. Plant Physiol. Biochem. 2018, 127, 223. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, H.; Li, H.; Gao, J.F.; Jiang, H.; Wang, C.; Guan, Y.F.; Yang, Z.N. Defective in tapetal development and function 1 is essential for anther development and tapetal function for microspore maturation in arabidopsis. Plant J. 2008, 55, 266–277. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Correction: Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef]
- Usadel, B.; Obayashi, T.; Mutwil, M.; Giorgi, F.M.; Bassel, G.W.; Tanimoto, M.; Chow, A.; Steinhauser, D.; Persson, S.; Provart, N.J. Co-expression tools for plant biology: Opportunities for hypothesis generation and caveats. Plant Cell Environ. 2009, 32, 1633–1651. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, J.; Pavlidis, P. “Guilt by association” is the exception rather than the rule in gene networks. PLoS Comput. Biol. 2012, 8, e1002444. [Google Scholar] [CrossRef] [PubMed]
- van Dam, S.; Vosa, U.; van der Graaf, A.; Franke, L.; de Magalhaes, J.P. Gene co-expression analysis for functional classification and gene-disease predictions. Brief Bioinf. 2018, 19, 575–592. [Google Scholar] [CrossRef]
- Zheng, J.; He, C.; Qin, Y.; Lin, G.; Park, W.D.; Sun, M.; Li, J.; Lu, X.; Zhang, C.; Yeh, C.T.; et al. Co-expression analysis aids in the identification of genes in the cuticular wax pathway in maize. Plant J. 2019, 97, 530–542. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for agrobacterium-mediated transformation of arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Peterson, R.; Slovin, J.P.; Chen, C. A simplified method for differential staining of aborted and non-aborted pollen grains. Int. J. Plant Biol. 2010, 1, e13. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. Mcscanx: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Toufighi, K.; Brady, S.M.; Austin, R.; Ly, E.; Provart, N.J. The botany array resource: E-northerns, expression angling, and promoter analyses. Plant J. 2005, 43, 153–163. [Google Scholar] [CrossRef]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. Agrigo: A go analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, W64–W70. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus ID | Gene Name | CDS Length (bp) | Protein Length (aa) | Chromosome | Gene Start | Gene End | gDNA Length (bp) | No. of Exons | Best Hit to Arabidopsis (BLASTP) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | Gene Name | E-Value | |||||||||
| BraA01g012490.3C | BrBGLU1 | 1290 | 430 | Chr 01 | 6,516,500 | 6,519,450 | 2950 | 11 | AT4G21760 | BGLU47 | 0 |
| BraA01g029610.3C | BrBGLU2 | 1452 | 484 | Chr 01 | 19,673,693 | 19,677,620 | 3927 | 12 | AT1G61820 | BGLU46 | 0 |
| BraA01g029670.3C | BrBGLU3 | 1551 | 517 | Chr 01 | 19,772,218 | 19,775,132 | 2914 | 12 | AT1G61810 | BGLU45 | 0 |
| BraA01g032340.3C | BrBGLU4 | 873 | 291 | Chr 01 | 22,083,906 | 22,086,749 | 2843 | 8 | AT1G52400 | BGLU18 | 6.38 × 10−85 |
| BraA01g034680.3C | BrBGLU5 | 1545 | 515 | Chr 01 | 23,747,770 | 23,750,431 | 2661 | 12 | AT3G18080 | BGLU44 | 0 |
| BraA01g034690.3C | BrBGLU6 | 1464 | 488 | Chr 01 | 23,754,677 | 23,757,145 | 2468 | 10 | AT3G18070 | BGLU43 | 0 |
| BraA01g040820.3C | BrBGLU7 | 1374 | 458 | Chr 01 | 27,455,063 | 27,458,182 | 3119 | 12 | AT3G09260 | BGLU23 | 0 |
| BraA01g041990.3C | BrBGLU8 | 1926 | 642 | Chr 01 | 28,048,456 | 28,052,048 | 3592 | 9 | AT3G06510 | BGLU48 | 0 |
| BraA01g043570.3C | BrBGLU9 | 1566 | 522 | Chr 01 | 28,909,991 | 28,913,218 | 3227 | 13 | AT3G03640 | BGLU25 | 0 |
| BraA02g023150.3C | BrBGLU10 | 1653 | 551 | Chr 02 | 13,570,012 | 13,572,993 | 2981 | 13 | AT1G75940 | BGLU20 | 0 |
| BraA03g011770.3C | BrBGLU11 | 894 | 298 | Chr 03 | 5,059,601 | 5,062,343 | 2742 | 8 | AT1G45191 | BGLU1 | 2.53 × 10−60 |
| BraA03g011780.3C | BrBGLU12 | 1023 | 341 | Chr 03 | 5,063,808 | 5,066,446 | 2638 | 12 | AT1G60090 | BGLU4 | 9.15 × 10−82 |
| BraA03g024570.3C | BrBGLU13 | 846 | 282 | Chr 03 | 12,073,161 | 12,075,780 | 2619 | 10 | AT4G22100 | BGLU3 | 5.01 × 10−68 |
| BraA03g033950.3C | BrBGLU14 | 1398 | 466 | Chr 03 | 16,798,347 | 16,801,182 | 2835 | 11 | AT3G09260 | BGLU23 | 0 |
| BraA03g041420.3C | BrBGLU15 | 729 | 243 | Chr 03 | 20,778,980 | 20,781,062 | 2082 | 5 | AT4G22100 | BGLU3 | 2.54 × 10−61 |
| BraA03g041430.3C | BrBGLU16 | 669 | 223 | Chr 03 | 20,781,085 | 20,782,278 | 1193 | 7 | AT1G60090 | BGLU4 | 3.4 × 10−100 |
| BraA03g049730.3C | BrBGLU17 | 1563 | 521 | Chr 03 | 25,428,252 | 25,430,947 | 2695 | 12 | AT4G21760 | BGLU47 | 0 |
| BraA04g000610.3C | BrBGLU18 | 1431 | 477 | Chr 04 | 408,734 | 411,303 | 2569 | 11 | AT4G27830 | BGLU10 | 0 |
| BraA04g002030.3C | BrBGLU19 | 1497 | 499 | Chr 04 | 1,226,615 | 1,230,317 | 3702 | 12 | AT3G60140 | BGLU30 | 0 |
| BraA04g002040.3C | BrBGLU20 | 2058 | 686 | Chr 04 | 1,238,401 | 1,245,729 | 7328 | 18 | AT3G60120 | BGLU27 | 2.2 × 10−149 |
| BraA04g010020.3C | BrBGLU21 | 1341 | 447 | Chr 04 | 7,880,007 | 7,883,680 | 3673 | 13 | AT5G36890 | BGLU42 | 0 |
| BraA04g020960.3C | BrBGLU22 | 891 | 297 | Chr 04 | 15,776,965 | 15,780,643 | 3678 | 8 | AT5G44640 | BGLU13 | 1.5 × 10−105 |
| BraA04g023640.3C | BrBGLU23 | 1638 | 546 | Chr 04 | 17,341,351 | 17,344,539 | 3188 | 9 | AT2G32860 | BGLU33 | 0 |
| BraA04g031090.3C | BrBGLU24 | 1233 | 411 | Chr 04 | 21,218,491 | 21,221,708 | 3217 | 12 | AT3G60120 | BGLU27 | 0 |
| BraA04g031100.3C | BrBGLU25 | 1380 | 460 | Chr 04 | 21,229,282 | 21,232,323 | 3041 | 11 | AT5G24550 | BGLU32 | 0 |
| BraA04g031130.3C | BrBGLU26 | 267 | 89 | Chr 04 | 21,248,071 | 21,248,622 | 551 | 4 | AT2G44450 | BGLU15 | 5.66 × 10−85 |
| BraA04g031140.3C | BrBGLU27 | 525 | 175 | Chr 04 | 21,249,497 | 21,251,177 | 1680 | 6 | AT2G44450 | BGLU15 | 1.4 × 10−110 |
| BraA05g004330.3C | BrBGLU28 | 1623 | 541 | Chr 05 | 2,194,953 | 2,197,671 | 2718 | 11 | AT3G60120 | BGLU27 | 0 |
| BraA05g004340.3C | BrBGLU29 | 1527 | 509 | Chr 05 | 2,201,548 | 2,211,165 | 9617 | 12 | AT2G44450 | BGLU15 | 0 |
| BraA05g004350.3C | BrBGLU30 | 1518 | 506 | Chr 05 | 2,216,705 | 2,220,674 | 3969 | 12 | AT5G44640 | BGLU13 | 0 |
| BraA05g004360.3C | BrBGLU31 | 1326 | 442 | Chr 05 | 2,223,901 | 2,227,525 | 3624 | 9 | AT2G44460 | BGLU28 | 2.7 × 10−131 |
| BraA05g004370.3C | BrBGLU32 | 1155 | 385 | Chr 05 | 2,245,448 | 2,248,087 | 2639 | 7 | AT3G60140 | BGLU30 | 2.1 × 10−140 |
| BraA05g004380.3C | BrBGLU33 | 1281 | 427 | Chr 05 | 2,255,905 | 2,258,985 | 3080 | 11 | AT5G24540 | BGLU31 | 5.1 × 10−145 |
| BraA05g004390.3C | BrBGLU34 | 1545 | 515 | Chr 05 | 2,261,685 | 2,270,997 | 9312 | 11 | AT2G44490 | BGLU26 | 0 |
| BraA05g012860.3C | BrBGLU35 | 1536 | 512 | Chr 05 | 7,011,962 | 7,015,388 | 3426 | 11 | AT2G32860 | BGLU33 | 2.3 × 10−162 |
| BraA05g012870.3C | BrBGLU36 | 957 | 319 | Chr 05 | 7,023,185 | 7,028,485 | 5300 | 8 | AT2G32860 | BGLU33 | 4.3 × 10−102 |
| BraA05g015060.3C | BrBGLU37 | 1461 | 487 | Chr 05 | 8,601,511 | 8,604,284 | 2773 | 13 | AT5G36890 | BGLU42 | 0 |
| BraA05g017770.3C | BrBGLU38 | 1278 | 426 | Chr 05 | 10,758,114 | 10,760,718 | 2604 | 11 | AT1G52400 | BGLU18 | 0 |
| BraA05g033960.3C | BrBGLU39 | 1434 | 478 | Chr 05 | 24,329,347 | 24,333,054 | 3707 | 12 | AT4G27830 | BGLU10 | 0 |
| BraA05g037140.3C | BrBGLU40 | 1332 | 444 | Chr 05 | 25,685,745 | 25,688,976 | 3231 | 11 | AT3G09260 | BGLU23 | 0 |
| BraA05g037150.3C | BrBGLU41 | 1374 | 458 | Chr 05 | 25,691,345 | 25,694,889 | 3544 | 12 | AT3G09260 | BGLU23 | 0 |
| BraA05g038920.3C | BrBGLU42 | 1782 | 594 | Chr 05 | 26,547,600 | 26,550,524 | 2924 | 11 | AT3G06510 | BGLU48 | 0 |
| BraA06g002000.3C | BrBGLU43 | 1347 | 449 | Chr 06 | 1,220,707 | 1,223,925 | 3218 | 12 | AT1G52400 | BGLU18 | 1.5 × 10−173 |
| BraA06g011040.3C | BrBGLU44 | 1080 | 360 | Chr 06 | 5,995,931 | 6,000,341 | 4410 | 8 | AT3G21370 | BGLU19 | 0 |
| BraA06g024630.3C | BrBGLU45 | 1599 | 533 | Chr 06 | 17,098,530 | 17,100,946 | 2416 | 4 | AT5G44640 | BGLU13 | 0 |
| BraA06g038720.3C | BrBGLU46 | 312 | 104 | Chr 06 | 25,758,774 | 25,759,164 | 390 | 2 | AT4G22100 | BGLU3 | 8 × 10−52 |
| BraA07g008030.3C | BrBGLU47 | 1434 | 478 | Chr 07 | 8,145,282 | 8,147,911 | 2629 | 11 | AT1G60090 | BGLU4 | 0 |
| BraA07g008050.3C | BrBGLU48 | 1428 | 476 | Chr 07 | 8,161,734 | 8,164,666 | 2932 | 12 | AT4G22100 | BGLU3 | 0 |
| BraA07g011940.3C | BrBGLU49 | 765 | 255 | Chr 07 | 11,620,825 | 11,623,618 | 2793 | 8 | AT3G62750 | BGLU8 | 3.75 × 10−29 |
| BraA07g024150.3C | BrBGLU50 | 1545 | 515 | Chr 07 | 18,998,283 | 19,001,907 | 3624 | 12 | AT3G60130 | BGLU16 | 0 |
| BraA08g002600.3C | BrBGLU51 | 1515 | 505 | Chr 08 | 1,915,015 | 1,917,839 | 2824 | 13 | AT1G47600 | BGLU34 | 0 |
| BraA08g008860.3C | BrBGLU52 | 408 | 136 | Chr 08 | 7,848,512 | 7,850,189 | 1677 | 4 | AT3G09260 | BGLU23 | 1.03 × 10−75 |
| BraA08g014870.3C | BrBGLU53 | 930 | 310 | Chr 08 | 12,301,355 | 12,303,970 | 2615 | 7 | AT4G22100 | BGLU3 | 3.9 × 10−127 |
| BraA08g025770.3C | BrBGLU54 | 1506 | 502 | Chr 08 | 18,552,796 | 18,556,470 | 3674 | 11 | AT1G26560 | BGLU40 | 0 |
| BraA09g018020.3C | BrBGLU55 | 1524 | 508 | Chr 09 | 11,385,273 | 11,386,948 | 1675 | 2 | AT5G44640 | BGLU13 | 0 |
| BraA09g038410.3C | BrBGLU56 | 1527 | 509 | Chr 09 | 30,292,880 | 30,295,941 | 3061 | 11 | AT1G26560 | BGLU40 | 0 |
| BraA09g049950.3C | BrBGLU57 | 1542 | 514 | Chr 09 | 37,157,612 | 37,160,275 | 2663 | 10 | AT3G60120 | BGLU27 | 0 |
| BraA09g049960.3C | BrBGLU58 | 1248 | 416 | Chr 09 | 37,164,182 | 37,167,906 | 3724 | 10 | AT3G60130 | BGLU16 | 2.2 × 10−165 |
| BraA09g049970.3C | BrBGLU59 | 1047 | 349 | Chr 09 | 37,169,340 | 37,173,150 | 3810 | 8 | AT3G60130 | BGLU16 | 3.4 × 10−165 |
| BraA09g049980.3C | BrBGLU60 | 1326 | 442 | Chr 09 | 37,178,403 | 37,186,233 | 7830 | 11 | AT3G60140 | BGLU30 | 2.6 × 10−179 |
| BraA09g052040.3C | BrBGLU61 | 1461 | 487 | Chr 09 | 38,112,498 | 38,115,245 | 2747 | 11 | AT4G27830 | BGLU10 | 0 |
| BraA09g052050.3C | BrBGLU62 | 1176 | 392 | Chr 09 | 38,116,332 | 38,118,944 | 2612 | 11 | AT4G27830 | BGLU10 | 9.8 × 10−140 |
| BraA10g001490.3C | BrBGLU63 | 1281 | 427 | Chr 10 | 776,354 | 778,932 | 2578 | 11 | AT1G02850 | BGLU11 | 0 |
| BraA10g012660.3C | BrBGLU64 | 1569 | 523 | Chr 10 | 10,414,966 | 10,417,449 | 2483 | 11 | AT5G54570 | BGLU41 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; Jiang, Y.; Hur, Y. Genome-Wide Analysis of Glycoside Hydrolase Family 1 β-glucosidase Genes in Brassica rapa and Their Potential Role in Pollen Development. Int. J. Mol. Sci. 2019, 20, 1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071663
Dong X, Jiang Y, Hur Y. Genome-Wide Analysis of Glycoside Hydrolase Family 1 β-glucosidase Genes in Brassica rapa and Their Potential Role in Pollen Development. International Journal of Molecular Sciences. 2019; 20(7):1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071663
Chicago/Turabian StyleDong, Xiangshu, Yuan Jiang, and Yoonkang Hur. 2019. "Genome-Wide Analysis of Glycoside Hydrolase Family 1 β-glucosidase Genes in Brassica rapa and Their Potential Role in Pollen Development" International Journal of Molecular Sciences 20, no. 7: 1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071663