Roles of Natural Killer T Cells and Natural Killer Cells in Kidney Injury

 , , and
, , and
Abstract
:1. Introduction
2. Mouse Natural Killer T (NKT) Cells and Natural Killer (NK) Cells in the Kidney under Normal and Activated Conditions
3. Functions and Roles of Mouse NKT Cells in Renal Diseases and Pathological Conditions
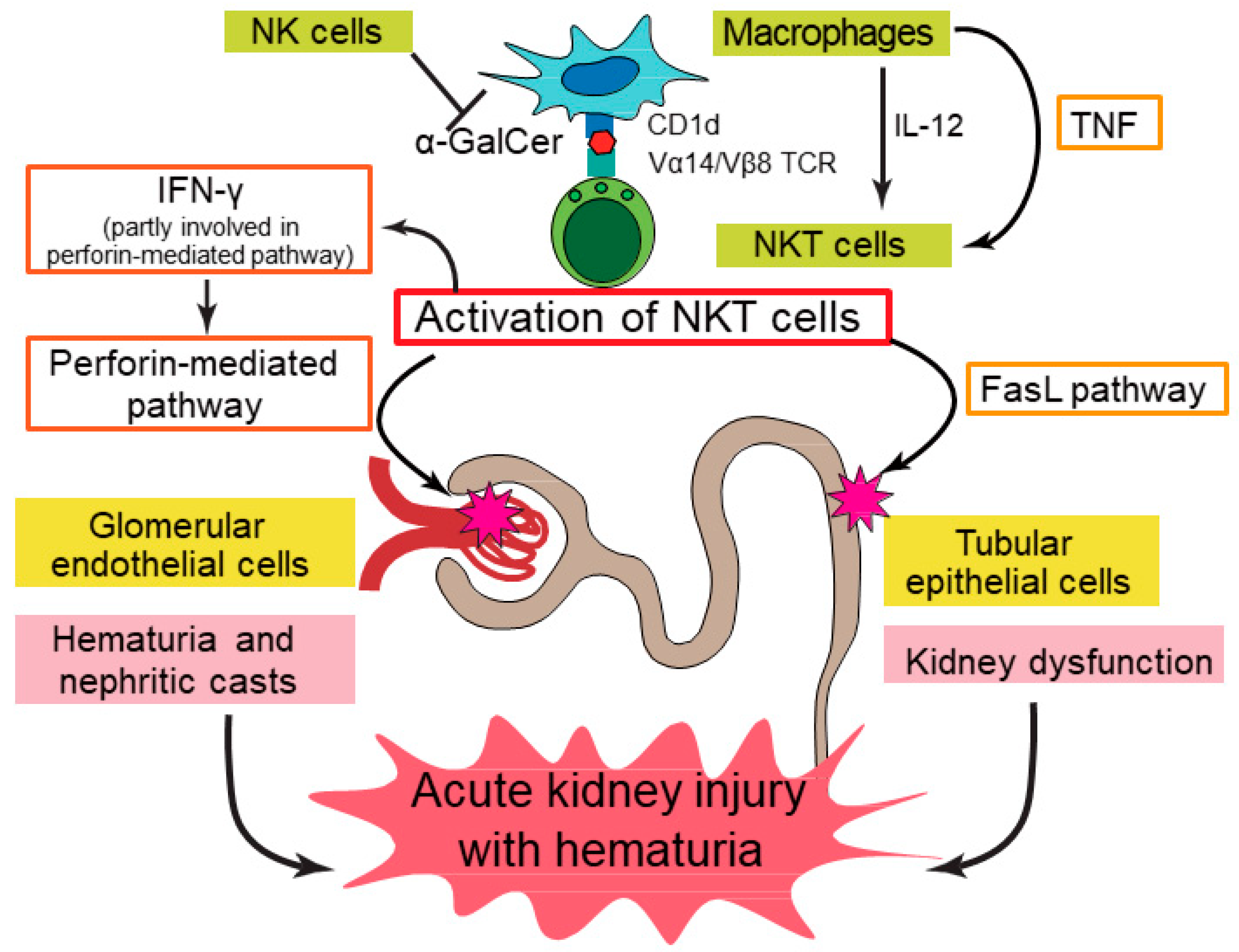
3.1. Acute Kidney Injury (AKI)
3.2. Lupus Nephritis
3.3. Other Renal Diseases
3.4. Renal Transplantation
4. Role of Mouse NK Cells in Kidney Injury
5. CD56+ T Cells Act as Human NKT Cells in Kidney Injury
6. Human CD56+ NK Cells in Kidney Injury
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seki, S.; Habu, Y.; Kawamura, T.; Takeda, K.; Dobashi, H.; Ohkawa, T.; Hiraide, H. The liver as a crucial organ in the first line of host defense: The roles of Kupffer cells, natural killer (NK) cells and NK1.1 Ag+ T cells in T helper 1 immune responses. Immunol. Rev. 2000, 174, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Motoki, K.; Akimoto, K.; Natori, T.; Sakai, T.; Sawa, E.; Yamaji, K.; Koezuka, Y.; Kobayashi, E.; Fukushima, H. Structure-activity relationship of alpha-galactosylceramides against B16-bearing mice. J. Med. Chem. 1995, 38, 2176–2187. [Google Scholar] [CrossRef]
- Kawano, T.; Cui, J.; Koezuka, Y.; Toura, I.; Kaneko, Y.; Motoki, K.; Ueno, H.; Nakagawa, R.; Sato, H.; Kondo, E.; et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science 1997, 278, 1626–1629. [Google Scholar] [CrossRef]
- Nakagawa, R.; Nagafune, I.; Tazunoki, Y.; Ehara, H.; Tomura, H.; Iijima, R.; Motoki, K.; Kamishohara, M.; Seki, S. Mechanisms of the antimetastatic effect in the liver and of the hepatocyte injury induced by alpha-galactosylceramide in mice. J. Immunol. 2001, 166, 6578–6584. [Google Scholar] [CrossRef] [PubMed]
- Inui, T.; Nakashima, H.; Habu, Y.; Nakagawa, R.; Fukasawa, M.; Kinoshita, M.; Shinomiya, N.; Seki, S. Neutralization of tumor necrosis factor abrogates hepatic failure induced by alpha-galactosylceramide without attenuating its antitumor effect in aged mice. J. Hepatol. 2005, 43, 670–678. [Google Scholar] [CrossRef]
- Kronenberg, M. Toward an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 2005, 23, 877–900. [Google Scholar] [CrossRef] [PubMed]
- Kawarabayashi, N.; Seki, S.; Hatsuse, K.; Ohkawa, T.; Koike, Y.; Aihara, T.; Habu, Y.; Nakagawa, R.; Ami, K.; Hiraide, H.; et al. Decrease of CD56(+)T cells and natural killer cells in cirrhotic livers with hepatitis C may be involved in their susceptibility to hepatocellular carcinoma. Hepatology 2000, 32, 962–969. [Google Scholar] [CrossRef]
- Takayama, E.; Koike, Y.; Ohkawa, T.; Majima, T.; Fukasawa, M.; Shinomiya, N.; Yamaguchi, T.; Konishi, M.; Hiraide, H.; Tadakuma, T.; et al. Functional and Vbeta repertoire characterization of human CD8+ T-cell subsets with natural killer cell markers, CD56+ CD57- T cells, CD56+ CD57+ T cells and CD56- CD57+ T cells. Immunology 2003, 108, 211–219. [Google Scholar] [CrossRef]
- Seki, S.; Nakashima, H.; Nakashima, M.; Kinoshita, M. Antitumor immunity produced by the liver Kupffer cells, NK cells, NKT cells, and CD8 CD122 T cells. Clin. Dev. Immunol. 2011, 2011, 868345. [Google Scholar] [CrossRef] [PubMed]
- Doherty, D.G.; O’Farrelly, C. Innate and adaptive lymphoid cells in the human liver. Immunol. Rev. 2000, 174, 5–20. [Google Scholar] [CrossRef]
- Kumar, V. NKT-cell subsets: Promoters and protectors in inflammatory liver disease. J. Hepatol. 2013, 59, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Alamartine, E.; Videcoq, C.; Saby, P.; Sabido, O.; Berthoux, F. T lymphocytes expressing NK antigens: Kinetics after renal transplantation. Transpl. Proc. 2000, 32, 419–420. [Google Scholar] [CrossRef]
- Ascon, D.B.; Ascon, M.; Satpute, S.; Lopez-Briones, S.; Racusen, L.; Colvin, R.B.; Soloski, M.J.; Rabb, H. Normal mouse kidneys contain activated and CD3+CD4− CD8− double-negative T lymphocytes with a distinct TCR repertoire. J. Leukoc. Biol. 2008, 84, 1400–1409. [Google Scholar] [CrossRef]
- Uchida, T.; Nakashima, H.; Ito, S.; Ishikiriyama, T.; Nakashima, M.; Seki, S.; Kumagai, H.; Oshima, N. Activated natural killer T cells in mice induce acute kidney injury with hematuria through possibly common mechanisms shared by human CD56(+) T cells. Am. J. Physiol. Ren. Physiol. 2018, 315, F618–F627. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.E.; Becker, M.; Mittrucker, H.W.; Panzer, U. Tissue-Resident Lymphocytes in the Kidney. J. Am. Soc. Nephrol. 2018, 29, 389–399. [Google Scholar] [CrossRef]
- Kinjo, Y.; Wu, D.; Kim, G.; Xing, G.W.; Poles, M.A.; Ho, D.D.; Tsuji, M.; Kawahara, K.; Wong, C.H.; Kronenberg, M. Recognition of bacterial glycosphingolipids by natural killer T cells. Nature 2005, 434, 520–525. [Google Scholar] [CrossRef]
- Mattner, J.; Debord, K.L.; Ismail, N.; Goff, R.D.; Cantu, C., 3rd; Zhou, D.; Saint-Mezard, P.; Wang, V.; Gao, Y.; Yin, N.; et al. Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature 2005, 434, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, T.; Kinoshita, M.; Inatsu, A.; Habu, Y.; Nakashima, H.; Shinomiya, N.; Seki, S. Functional alterations of liver innate immunity of mice with aging in response to CpG-oligodeoxynucleotide. Hepatology 2008, 48, 1586–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, K.; Takeda, K.; Hashimoto, W.; Satoh, M.; Okuyama, R.; Yanai, N.; Obinata, M.; Kumagai, K.; Takada, H.; Hiraide, H.; et al. Involvement of NK1+ T cells and their IFN-gamma production in the generalized Shwartzman reaction. J. Immunol. 1998, 160, 3522–3527. [Google Scholar]
- Sato, K.; Kinoshita, M.; Motegi, A.; Habu, Y.; Takayama, E.; Nonoyama, S.; Hiraide, H.; Seki, S. Critical role of the liver CD8+ CD122+ T cells in the generalized Shwartzman reaction of mice. Eur. J. Immunol. 2005, 35, 593–602. [Google Scholar] [CrossRef]
- Shono, S.; Habu, Y.; Nakashima, M.; Sato, A.; Nakashima, H.; Miyazaki, H.; Kinoshita, M.; Tsumatori, G.; Shinomiya, N.; Seki, S. The immunologic outcome of enhanced function of mouse liver lymphocytes and Kupffer cells by high-fat and high-cholesterol diet. Shock 2011, 36, 484–493. [Google Scholar] [CrossRef]
- Osman, Y.; Kawamura, T.; Naito, T.; Takeda, K.; Van Kaer, L.; Okumura, K.; Abo, T. Activation of hepatic NKT cells and subsequent liver injury following administration of alpha-galactosylceramide. Eur. J. Immunol. 2000, 30, 1919–1928. [Google Scholar] [CrossRef]
- Li, L.; Huang, L.; Sung, S.S.; Lobo, P.I.; Brown, M.G.; Gregg, R.K.; Engelhard, V.H.; Okusa, M.D. NKT cell activation mediates neutrophil IFN-gamma production and renal ischemia-reperfusion injury. J. Immunol. 2007, 178, 5899–5911. [Google Scholar] [CrossRef] [PubMed]
- Ferhat, M.; Robin, A.; Giraud, S.; Sena, S.; Goujon, J.M.; Touchard, G.; Hauet, T.; Girard, J.P.; Gombert, J.M.; Herbelin, A.; et al. Endogenous IL-33 Contributes to Kidney Ischemia-Reperfusion Injury as an Alarmin. J. Am. Soc. Nephrol. 2018, 29, 1272–1288. [Google Scholar] [CrossRef]
- Zhang, J.; Han, C.; Dai, H.; Hou, J.; Dong, Y.; Cui, X.; Xu, L.; Zhang, M.; Xia, Q. Hypoxia-Inducible Factor-2alpha Limits Natural Killer T Cell Cytotoxicity in Renal Ischemia/Reperfusion Injury. J. Am. Soc. Nephrol. 2016, 27, 92–106. [Google Scholar] [CrossRef]
- Yang, S.H.; Lee, J.P.; Jang, H.R.; Cha, R.H.; Han, S.S.; Jeon, U.S.; Kim, D.K.; Song, J.; Lee, D.S.; Kim, Y.S. Sulfatide-reactive natural killer T cells abrogate ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2011, 22, 1305–1314. [Google Scholar] [CrossRef]
- Nakashima, H.; Inui, T.; Habu, Y.; Kinoshita, M.; Nagao, S.; Kawaguchi, A.; Miura, S.; Shinomiya, N.; Yagita, H.; Seki, S. Activation of mouse natural killer T cells accelerates liver regeneration after partial hepatectomy. Gastroenterology 2006, 131, 1573–1583. [Google Scholar] [CrossRef]
- Yang, J.; Liang, D.; Zhang, H.; Liu, Z.; Le, W.; Zhou, M.; Hu, W.; Zeng, C.; Liu, Z. Long-term renal outcomes in a cohort of 1814 Chinese patients with biopsy-proven lupus nehritis. Lupus 2015, 24, 1468–1478. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Z.; Lee, K.; Wei, C.; Liu, Z.; Zhang, M.; Zhou, M.; Cai, M.; Zhang, W.; Chuang, P.Y.; et al. Transcriptomic analysis uncovers novel synergistic mechanisms in combination therapy for lupus nephritis. Kidney Int. 2018, 93, 416–429. [Google Scholar] [CrossRef]
- Morshed, S.R.; Mannoor, K.; Halder, R.C.; Kawamura, H.; Bannai, M.; Sekikawa, H.; Watanabe, H.; Abo, T. Tissue-specific expansion of NKT and CD5+B cells at the onset of autoimmune disease in (NZBxNZW)F1 mice. Eur. J. Immunol. 2002, 32, 2551–2561. [Google Scholar] [CrossRef]
- Green, M.R.; Kennell, A.S.; Larche, M.J.; Seifert, M.H.; Isenberg, D.A.; Salaman, M.R. Natural killer T cells in families of patients with systemic lupus erythematosus: Their possible role in regulation of IGG production. Arthritis Rheumatol. 2007, 56, 303–310. [Google Scholar] [CrossRef]
- Godo, M.; Sessler, T.; Hamar, P. Role of invariant natural killer T (iNKT) cells in systemic lupus erythematosus. Curr. Med. Chem. 2008, 15, 1778–1787. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, M.; Wang, J.; Li, X. Immunoregulation of NKT Cells in Systemic Lupus Erythematosus. J. Immunol. Res 2015, 2015, 206731. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Q.; Saxena, V.; Xu, H.; Van Kaer, L.; Wang, C.R.; Singh, R.R. Repeated alpha-galactosylceramide administration results in expansion of NK T cells and alleviates inflammatory dermatitis in MRL-lpr/lpr mice. J. Immunol. 2003, 171, 4439–4446. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Yang, J.Q.; Parekh, V.V.; Wei, J.; Wang, C.R.; Joyce, S.; Singh, R.R.; van Kaer, L. The natural killer T cell ligand alpha-galactosylceramide prevents or promotes pristane-induced lupus in mice. Eur. J. Immunol. 2005, 35, 1143–1154. [Google Scholar] [CrossRef]
- Peng, S.L. Experimental use of mouse models of systemic lupus erythematosus. Methods Mol. Biol. 2012, 900, 135–168. [Google Scholar]
- Postol, E.; Meyer, A.; Cardillo, F.; de Alencar, R.; Pessina, D.; Nihei, J.; Mariano, M.; Mengel, J. Long-term administration of IgG2a anti-NK1.1 monoclonal antibody ameliorates lupus-like disease in NZB/W mice in spite of an early worsening induced by an IgG2a-dependent BAFF/BLyS production. Immunology 2008, 125, 184–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, D.; Liu, Y.; Sidobre, S.; Kronenberg, M.; Strober, S. Activation of natural killer T cells in NZB/W mice induces Th1-type immune responses exacerbating lupus. J. Clin. Investig. 2003, 112, 1211–1222. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Q.; Kim, P.J.; Singh, R.R. Brief treatment with iNKT cell ligand alpha-galactosylceramide confers a long-term protection against lupus. J. Clin. Immunol. 2012, 32, 106–113. [Google Scholar] [CrossRef]
- Uldrich, A.P.; Crowe, N.Y.; Kyparissoudis, K.; Pellicci, D.G.; Zhan, Y.; Lew, A.M.; Bouillet, P.; Strasser, A.; Smyth, M.J.; Godfrey, D.I. NKT cell stimulation with glycolipid antigen in vivo: Costimulation-dependent expansion, Bim-dependent contraction, and hyporesponsiveness to further antigenic challenge. J. Immunol. 2005, 175, 3092–3101. [Google Scholar] [CrossRef]
- Uchida, T.; Nakashima, H.; Yamagata, A.; Ito, S.; Ishikiriyama, T.; Nakashima, M.; Seki, S.; Kumagai, H.; Oshima, N. Repeated administration of alpha-galactosylceramide ameliorates experimental lupus nephritis in mice. Sci. Rep. 2018, 8, 8225. [Google Scholar] [CrossRef]
- Takahashi, T.; Strober, S. Natural killer T cells and innate immune B cells from lupus-prone NZB/W mice interact to generate IgM and IgG autoantibodies. Eur. J. Immunol. 2008, 38, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Zhang, B.; Jarrell, J.A.; Price, J.V.; Dai, H.; Utz, P.J.; Strober, S. Ly108 expression distinguishes subsets of invariant NKT cells that help autoantibody production and secrete IL-21 from those that secrete IL-17 in lupus prone NZB/W mice. J. Autoimmun. 2014, 50, 87–98. [Google Scholar] [CrossRef]
- Singh, N.; Hong, S.; Scherer, D.C.; Serizawa, I.; Burdin, N.; Kronenberg, M.; Koezuka, Y.; van Kaer, L. Cutting edge: Activation of NK T cells by CD1d and alpha-galactosylceramide directs conventional T cells to the acquisition of a Th2 phenotype. J. Immunol. 1999, 163, 2373–2377. [Google Scholar]
- Hong, S.; Wilson, M.T.; Serizawa, I.; Wu, L.; Singh, N.; Naidenko, O.V.; Miura, T.; Haba, T.; Scherer, D.C.; Wei, J.; et al. The natural killer T-cell ligand alpha-galactosylceramide prevents autoimmune diabetes in non-obese diabetic mice. Nat. Med. 2001, 7, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Sharif, S.; Arreaza, G.A.; Zucker, P.; Mi, Q.S.; Sondhi, J.; Naidenko, O.V.; Kronenberg, M.; Koezuka, Y.; Delovitch, T.L.; Gombert, J.M.; et al. Activation of natural killer T cells by alpha-galactosylceramide treatment prevents the onset and recurrence of autoimmune Type 1 diabetes. Nat. Med. 2001, 7, 1057–1062. [Google Scholar] [CrossRef]
- Mizuno, M.; Masumura, M.; Tomi, C.; Chiba, A.; Oki, S.; Yamamura, T.; Miyake, S. Synthetic glycolipid OCH prevents insulitis and diabetes in NOD mice. J. Autoimmun. 2004, 23, 293–300. [Google Scholar] [CrossRef]
- Pereira, R.L.; Reis, V.O.; Semedo, P.; Buscariollo, B.N.; Donizetti-Oliveira, C.; Cenedeze, M.A.; Soares, M.F.; Pacheco-Silva, A.; Savage, P.B.; Camara, N.O.; et al. Invariant natural killer T cell agonist modulates experimental focal and segmental glomerulosclerosis. PLoS ONE 2012, 7, e32454. [Google Scholar] [CrossRef]
- Yang, S.H.; Kim, S.J.; Kim, N.; Oh, J.E.; Lee, J.G.; Chung, N.H.; Kim, S.; Kim, Y.S. NKT cells inhibit the development of experimental crescentic glomerulonephritis. J. Am. Soc. Nephrol. 2008, 19, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Mesnard, L.; Keller, A.C.; Michel, M.L.; Vandermeersch, S.; Rafat, C.; Letavernier, E.; Tillet, Y.; Rondeau, E.; Leite-de-Moraes, M.C. Invariant natural killer T cells and TGF-beta attenuate anti-GBM glomerulonephritis. J. Am. Soc. Nephrol. 2009, 20, 1282–1292. [Google Scholar] [CrossRef] [PubMed]
- Riedel, J.H.; Paust, H.J.; Turner, J.E.; Tittel, A.P.; Krebs, C.; Disteldorf, E.; Wegscheid, C.; Tiegs, G.; Velden, J.; Mittrucker, H.W.; et al. Immature renal dendritic cells recruit regulatory CXCR6(+) invariant natural killer T cells to attenuate crescentic GN. J. Am. Soc. Nephrol. 2012, 23, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, C.F.; Naffah-de-Souza, C.; Castoldi, A.; Correa-Costa, M.; Braga, T.T.; Naka, E.L.; Amano, M.T.; Abate, D.T.; Hiyane, M.I.; Cenedeze, M.A.; et al. Administration of alpha-Galactosylceramide Improves Adenine-Induced Renal Injury. Mol. Med. 2015, 21, 553–562. [Google Scholar] [CrossRef]
- Huang, H.; Lu, Y.; Zhou, T.; Gu, G.; Xia, Q. Innate Immune Cells in Immune Tolerance After Liver Transplantation. Front. Immunol. 2018, 9, 2401. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, A.; Yasunami, Y.; Nabeyama, K.; Nakano, M.; Satoh, M.; Matsuoka, N.; Ono, J.; Nakayama, T.; Taniguchi, M.; Tanaka, M.; et al. Natural killer T-cells participate in rejection of islet allografts in the liver of mice. Diabetes 2006, 55, 34–39. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Wang, S.; Huang, X.; Min, W.P.; Sun, H.; Liu, W.; Garcia, B.; Jevnikar, A.M. NK cells induce apoptosis in tubular epithelial cells and contribute to renal ischemia-reperfusion injury. J. Immunol. 2008, 181, 7489–7498. [Google Scholar] [CrossRef]
- Spada, R.; Rojas, J.M.; Barber, D.F. Recent findings on the role of natural killer cells in the pathogenesis of systemic lupus erythematosus. J. Leukoc. Biol. 2015, 98, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Spada, R.; Rojas, J.M.; Perez-Yague, S.; Mulens, V.; Cannata-Ortiz, P.; Bragado, R.; Barber, D.F. NKG2D ligand overexpression in lupus nephritis correlates with increased NK cell activity and differentiation in kidneys but not in the periphery. J. Leukoc. Biol. 2015, 97, 583–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Zheng, L.; Wang, Y.; Wu, H.; Kairaitis, L.; Zhang, C.; Tay, Y.C.; Wang, Y.; Alexander, S.I.; Harris, D.C. NK cells do not mediate renal injury in murine adriamycin nephropathy. Kidney Int. 2006, 69, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.X.; Huang, X.; Jiang, J.; Lau, A.; Yin, Z.; Liu, W.; Haig, A.; Jevnikar, A.M. Natural Killer Cells Mediate Long-term Kidney Allograft Injury. Transplantation 2015, 99, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Halloran, P.F.; Famulski, K.S.; Reeve, J. Molecular assessment of disease states in kidney transplant biopsy samples. Nat. Rev. 2016, 12, 534–548. [Google Scholar] [CrossRef]
- Motegi, A.; Kinoshita, M.; Sato, K.; Shinomiya, N.; Ono, S.; Nonoyama, S.; Hiraide, H.; Seki, S. An in vitro Shwartzman reaction-like response is augmented age-dependently in human peripheral blood mononuclear cells. J. Leukoc. Biol. 2006, 79, 463–472. [Google Scholar] [CrossRef]
- Ohkawa, T.; Seki, S.; Dobashi, H.; Koike, Y.; Habu, Y.; Ami, K.; Hiraide, H.; Sekine, I. Systematic characterization of human CD8+ T cells with natural killer cell markers in comparison with natural killer cells and normal CD8+ T cells. Immunology 2001, 103, 281–290. [Google Scholar] [CrossRef]
- Law, B.M.; Wilkinson, R.; Wang, X.; Kildey, K.; Lindner, M.; Beagley, K.; Healy, H.; Kassianos, A.J. Effector gammadelta T cells in human renal fibrosis and chronic kidney disease. Nephrol. Dial. Transplant. 2019, 34, 40–48. [Google Scholar] [CrossRef]
- Chen, H.; You, H.; Wang, L.; Zhang, X.; Zhang, J.; He, W. Chaperonin-containing T-complex Protein 1 Subunit zeta Serves as an Autoantigen Recognized by Human Vdelta2 gammadelta T Cells in Autoimmune Diseases. J. Biol. Chem. 2016, 291, 19985–19993. [Google Scholar] [CrossRef]
- Drugarin, D.; Tatu, C.; Noveanu, L.; Paunescu, V. Identification of a novel subset of T lymphocytes in patients with Balkanic nephropathy. Autoimmunity 1996, 23, 119–126. [Google Scholar] [CrossRef]
- Mendoza-Alvarez, S.A.; Rodriguez-Davila, F.M.; Moranchel-Garcia, L.; Soto, V.; Quisped, N. Extranodal NK-T-cell lymphoma, nasal type in granulomatosis with polyangiitis. A case report. Rev. Med. Inst. Mex Seguro Soc. 2017, 55, 394–398. [Google Scholar] [PubMed]
- Wilson, F.P.; Nasr, S.H.; Markowitz, G.S.; Naylor, E.C.; Sterman, P.L.; D’Agati, V.D. A destructive nasal lesion and glomerulonephritis. Kidney Int. 2006, 69, 1699–1703. [Google Scholar] [CrossRef] [Green Version]
- Bachetoni, A.; Lionetti, P.; Cinti, P.; Alo, P.; Molajoni, E.R.; Di Tondo, U.; Barnaba, V.; Alfani, D.; Cortesini, R. Homing of CD4+CD56+ T lymphocytes into kidney allografts during tubular necrosis or rejection. Clin. Transplant. 1995, 9, 433–437. [Google Scholar] [PubMed]
- Zhu, L.; Aly, M.; Wang, H.; Karakizlis, H.; Weimer, R.; Morath, C.; Kuon, R.J.; Toth, B.; Opelz, G.; Daniel, V. Decreased NK cell immunity in kidney transplant recipients late post-transplant and increased NK-cell immunity in patients with recurrent miscarriage. PLoS ONE 2017, 12, e0186349. [Google Scholar] [CrossRef] [PubMed]
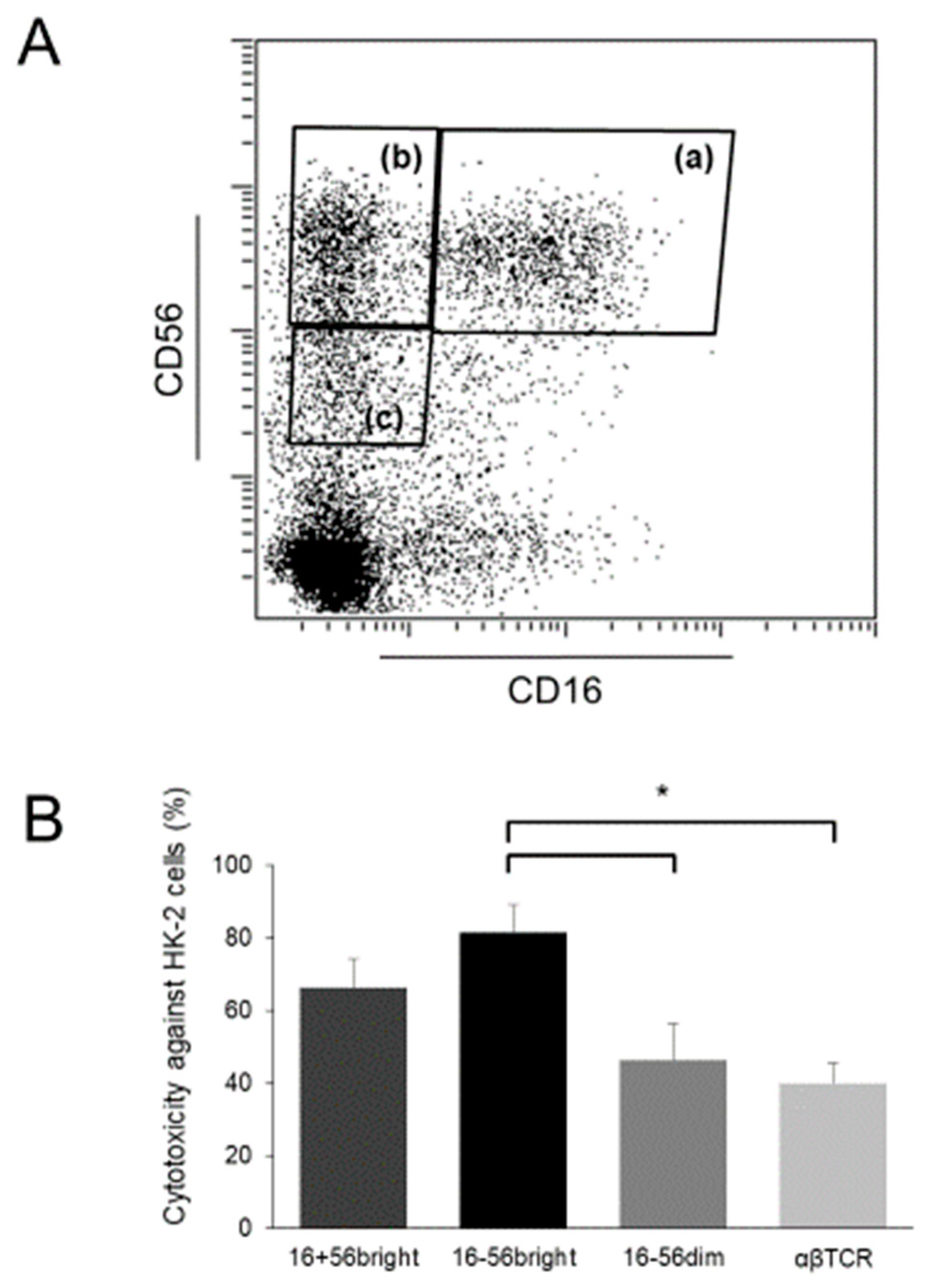
- Takahashi, E.; Kuranaga, N.; Satoh, K.; Habu, Y.; Shinomiya, N.; Asano, T.; Seki, S.; Hayakawa, M. Induction of CD16+ CD56bright NK cells with antitumour cytotoxicity not only from CD16− CD56bright NK Cells but also from CD16− CD56dim NK cells. Scand. J. Immunol. 2007, 65, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Jennette, J.C.; Falk, R.J.; Bacon, P.A.; Basu, N.; Cid, M.C.; Ferrario, F.; Flores-Suarez, L.F.; Gross, W.L.; Guillevin, L.; Hagen, E.C.; et al. 2012 revised International Chapel Hill Consensus Conference Nomenclature of Vasculitides. Arthritis Rheum. 2013, 65, 1–11. [Google Scholar] [CrossRef]
- Law, B.M.P.; Wilkinson, R.; Wang, X.; Kildey, K.; Lindner, M.; Rist, M.J.; Beagley, K.; Healy, H.; Kassianos, A.J. Interferon-gamma production by tubulointerstitial human CD56bright natural killer cells contributes to renal fibrosis and chronic kidney disease progression. Kidney Int. 2017, 92, 79–88. [Google Scholar] [CrossRef]
- Hotta, O.; Taguma, Y.; Yusa, N.; Ooyama, M. Analysis of mononuclear cells in urine using flow cytometry in glomerular diseases. Kidney Int. Suppl. 1994, 47, S117–S121. [Google Scholar]
- Iwatani, H.; Nagasawa, Y.; Yamamoto, R.; Iio, K.; Mizui, M.; Horii, A.; Kitahara, T.; Inohara, H.; Kumanogoh, A.; Imai, E.; et al. CD16+CD56+ cells are a potential culprit for hematuria in IgA nephropathy. Clin. Exp. Nephrol. 2015, 19, 216–224. [Google Scholar] [CrossRef]
- Hotta, O.; Taguma, Y.; Ooyama, M.; Yusa, N.; Nagura, H. Analysis of CD14+ cells and CD56+ cells in urine using flow cytometry: A useful tool for monitoring disease activity of IgA nephropathy. Clin. Nephrol. 1993, 39, 289–294. [Google Scholar]
- Cox, S.N.; Sallustio, F.; Serino, G.; Loverre, A.; Pesce, F.; Gigante, M.; Zaza, G.; Stifanelli, P.F.; Ancona, N.; Schena, F.P. Activated innate immunity and the involvement of CX3CR1-fractalkine in promoting hematuria in patients with IgA nephropathy. Kidney Int. 2012, 82, 548–560. [Google Scholar] [CrossRef]
- Risti, M.; Bicalho, M.D. MICA and NKG2D: Is There an Impact on Kidney Transplant Outcome? Front. Immunol. 2017, 8, 179. [Google Scholar] [CrossRef]
- Peraldi, M.N.; Berrou, J.; Dulphy, N.; Seidowsky, A.; Haas, P.; Boissel, N.; Metivier, F.; Randoux, C.; Kossari, N.; Guerin, A.; et al. Oxidative stress mediates a reduced expression of the activating receptor NKG2D in NK cells from end-stage renal disease patients. J. Immunol. 2009, 182, 1696–1705. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Irradiation (−) | Irradiation (+) | |
|---|---|---|
| Number of mice | 3 | 3 |
| Blood urea nitrogen (mg/dL) 1 | 27.4 ± 3.5 | 31.0 ± 3.5 |
| Alanine aminotransferase (IU/L) 2 | 208.0 ± 45.2 | 231.0 ± 29.1 |
| IL-4 (ng/mL) 3 | 3.1 ± 0.5 | 2.9 ± 0.3 |
| IFN-γ (ng/mL) 4 | 4.3 ± 1.2 | 19.7 ± 1.4 * |
| Strain (Model), Age | Treatment | Outcomes | Ref. |
|---|---|---|---|
| NZB/NZW F1 (BWF1) mice | None | Expansion of NKT cells in association with the onset of the disease | [30] |
| MRL/lpr mice, 2 months of age | 6 μg of alpha-galactosylceramide (α-GalCer) twice a week for 5 months | Improvement in inflammatory dermatitis without affecting renal disease | [34] |
| BALB/c and SJL mice (pristane-induced) | 6 μg of α-GalCer twice a week for 1 month | Suppression of nephritis (BALB/c mice): exacerbation of nephritis (SJL mice) | [35] |
| BWF1 young mice | 0.5 mg of anti-NK1.1 antibody three times a week for long periods | Amelioration of nephritis in late disease phase (worsening in early phase) | [37] |
| BWF1 mice, 20 weeks of age | 4 μg of α-GalCer twice a week for 2 weeks | Enhancement of Th1 immune responses and exacerbation of nephritis | [38] |
| BWF1 mice, 7 weeks of age | 4 μg of α-GalCer twice at a 3-day interval | Suppression of IL-10 production and reduction of severe proteinuria | [39] |
| BWF1 mice, 24 weeks of age | 2 μg of α-GalCer once a week for 4 weeks | Suppression of Th2 immune responses and amelioration of nephritis | [41] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchida, T.; Ito, S.; Kumagai, H.; Oda, T.; Nakashima, H.; Seki, S. Roles of Natural Killer T Cells and Natural Killer Cells in Kidney Injury. Int. J. Mol. Sci. 2019, 20, 2487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102487
Uchida T, Ito S, Kumagai H, Oda T, Nakashima H, Seki S. Roles of Natural Killer T Cells and Natural Killer Cells in Kidney Injury. International Journal of Molecular Sciences. 2019; 20(10):2487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102487
Chicago/Turabian StyleUchida, Takahiro, Seigo Ito, Hiroo Kumagai, Takashi Oda, Hiroyuki Nakashima, and Shuhji Seki. 2019. "Roles of Natural Killer T Cells and Natural Killer Cells in Kidney Injury" International Journal of Molecular Sciences 20, no. 10: 2487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102487