Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview


1
Microbiology Section, Department of Biology, Healthcare and Environment, Faculty of Pharmacy and Food Sciences, Universitat de Barcelona (UB), Av Joan XXIII, 27-31, 08028 Barcelona, Catalonia, Spain
2
Institut de Recerca en Nutrició i Seguretat Alimentària de la UB (INSA-UB), Universitat de Barcelona, 08921 Barcelona, Catalonia, Spain
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(10), 2534; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102534
Submission received: 25 April 2019
/
Revised: 17 May 2019
/
Accepted: 21 May 2019
/
Published: 23 May 2019
(This article belongs to the Section Bioactives and Nutraceuticals)
Abstract
:Nowadays, the oral use of probiotics is widespread. However, the safety profile with the use of live probiotics is still a matter of debate. Main risks include: Cases of systemic infections due to translocation, particularly in vulnerable patients and pediatric populations; acquisition of antibiotic resistance genes; or interference with gut colonization in neonates. To avoid these risks, there is an increasing interest in non-viable microorganisms or microbial cell extracts to be used as probiotics, mainly heat-killed (including tyndallized) probiotic bacteria (lactic acid bacteria and bifidobacteria). Heat-treated probiotic cells, cell-free supernatants, and purified key components are able to confer beneficial effects, mainly immunomodulatory effects, protection against enteropathogens, and maintenance of intestinal barrier integrity. At the clinical level, products containing tyndallized probiotic strains have had a role in gastrointestinal diseases, including bloating and infantile coli—in combination with mucosal protectors—and diarrhea. Heat-inactivated probiotics could also have a role in the management of dermatological or respiratory allergic diseases. The reviewed data indicate that heat-killed bacteria or their fractions or purified components have key probiotic effects, with advantages versus live probiotics (mainly their safety profile), positioning them as interesting strategies for the management of common prevalent conditions in a wide variety of patients´ characteristics.
1. Introduction
Currently, there is no doubt that the influence of the intestinal state on the health is gaining great interest, not only among the scientific community [1,2,3,4], but also for patients and consumers who are willing to adapt their diet habits to achieve the best well-being and health, besides other animal- or environment-related motives [5].
In this regard, gut microbiota, defined as a complex and dynamic microbiome containing more than 1000 different species, characteristic for each gastrointestinal (GI) tract segment, is recognized to be indispensable for homeostatic physiological functions in human health [1,3] at both the intestinal and extra-intestinal levels.
With the advent of new sequencing technologies, mainly based on the 16S ribosomal RNA genes, and the development of sophisticated bioinformatic tools, characterization of gut microbiota is being advanced, leading to the understanding of the composition and function of bacterial populations throughout the intestine and to the influence of fluctuations in the diversity of gut bacterial populations (known as dysbiosis) in the development of diseases [1,4,6,7].
This knowledge has been translated in a great interest in those therapeutical strategies to directly or indirectly influence gut microbiota to obtain clinical benefits, such as the use of probiotics, prebiotics, and other food supplements or fecal transplantations [8,9,10].
Probiotics, according to the revised definition of Food and Agriculture Organization (FAO)/World Health Organization (WHO, are considered as non-pathogenic live microorganisms that, when administered in adequate amounts, confer a health benefit on the host. Now probiotics are widely used in many countries in clinical practice and, frequently, are acquired by consumers with or without prescription [9,11,12,13,14]. In most cases, probiotics, mainly strains of Bifidobacterium or Lactobacillus species, come from the gut microbiota of healthy humans or from dairy products [15]. Probiotics also include species from the genera Streptococcus, Bacillus, and Enterococcus and the yeast Saccharomyces, which has been used as probiotics for many years [11,16,17].
Among the main effects of probiotics at the intestinal level, the following are noteworthy: Balancing and restoration of the gut microbiota, protection against pathogens, immunomodulation, and maintenance of intestinal barrier integrity [18]. Probiotics are widely used in dietary supplements, food, infant formula formulations, and medical devices [19,20]. They have demonstrated significant potential as therapeutic options for a variety of diseases, mainly gastrointestinal diseases (including acute infectious diarrhea, antibiotic-associated diarrhea, ulcerative colitis, irritable bowel syndrome, functional gastrointestinal disorders, or necrotizing enterocolitis), but also extra-intestinal disorders, such as hepatic encephalopathy [9,21].
However, there are still many issues on the table, for example, safety issues with the use of live microorganisms particularly in vulnerable populations [14,22,23,24], the lack of clear clinical recommendations in each specific clinical situation [9,21], the lack of compelling evidence from clinical trials for certain indications [8], the limited regulation of probiotics [20], or the lack of studies assessing the viability of microorganisms once in the intestine and the differences between viable or non-viable microorganisms [25].
In a recent survey on probiotic-prescribing practices among health care providers and review of current guidelines and published large clinical trials, it was concluded that recommendations appear to be inconsistent, non-specific, and, frequently, upon patient request. In a significant proportion, the choice of probiotic was left to the patient or the pharmacist. The three most common clinical indications for probiotics were prevention and treatment of antibiotic-related side effects and irritable bowel syndrome [21].
Moreover, safety issues with the use of live strains have been arisen in certain patient groups, such as neonates [11,26] and vulnerable patients [23], particularly due to translocation of bacteria from the gut to the systemic circulation, leading to an increased interest to use non-viable heat-killed probiotics [14,22,27].
There is considerable published evidence that preparations containing dead cells and their metabolites can also exert relevant biological responses, restoring the normal intestinal homeostasis, in many cases similar to that seen with live cells, although with potential differences [22,27]. After inactivation of bacteria, mainly by heat treatment, dead cells can release bacterial components with key immunomodulating effects and with antagonizing properties against pathogens. Different bacterial components, such as lipoteichoic acids, peptidoglycans, or exopolysaccharides (EPS), have been proposed to be mainly involved in these properties in preparations containing heat-killed bacteria [27,28,29].
Favorable properties of heat-killed bacteria have been observed in in vitro, animal models [27], and clinical trials [30,31], which have demonstrated their benefits in different indications, for example in neonates, without incurring the risks associated with live microorganisms [14,32], and with pharmaceutical advantages in terms of transport and storage (Table 1).
The objective of this article is to review the recent published studies indicating that heat-killed bacteria with health benefits can be a safe and feasible strategy for the management of different diseases, mainly gastrointestinal disorders, focusing on the possible mechanisms of action involved, in comparison with live strains.
2. Beneficial Effects of Probiotics
The most extensively-studied and widely-used probiotic bacteria are Lactobacillus and Bifidobacterium [14,33,34,35]. Species of these two genera (including Bifidobacterium breve, Bifidobacterium longum, Lactobactillus fermentum, Lactobacillus plantarum, Lactobacillus casei or Lactobacillus rhamnosus) naturally inhabit the human gastrointestinal tract, and are thought to play pivotal roles in maintaining human health [34,36]. Therefore, the oral administration of probiotics is thought to reinforce the physiological functions of gut microbiota at the intestinal level.
Probiotics are able to fight against pathogens by producing antimicrobial compounds and decreasing pH (with lactic acid production), and competing with pathogens for adhesion and colonization, and for nutrients and other growth factors in the gut [35], and suppressing the growth of pathogenic bacteria by directly binding to Gram-negative bacteria [9,19] (Figure 1).
Moreover, a body of evidence demonstrates that probiotic bacteria have immunomodulating properties, regulate inflammation in a number of ways, and enhance the epithelial barrier function to prevent chronic inflammation in the gut [19,35] (Figure 1). At the intestinal level, probiotics can also upregulate the intestinal electrolyte absorption and exert an effect on gut motility and constipation [9,37].
2.1. Immunomodulating Effects of Probiotics
There is evidence indicating that probiotics have immunomodulatory properties and protect against infection [33,38]. Probiotics, in strain-specific and dose-dependent manner, can modulate nonspecific cellular immune responses mediated by macrophages, natural killer (NK) cells, antigen-specific cytotoxic T-lymphocytes, and the release of cytokines [33].
Enhancement of innate immune responses has also been described for most probiotic strains, with IL-12 production by antigen-presenting cells (APCs), and the subsequent activation of host NK cells and promotion of type 1 helper T (Th1) cell responses [33,38].
Probiotics also enhance acquired immunity and induce IgA secretion in the intestine [38] by different mechanisms [39], with the activation of B lymphocytes and T cells [33,39].
However, the precise molecular mechanisms underlying the effects of each probiotic strain and different combinations of probiotic bacteria at the different immune pathways have not been fully resolved [38]. It should be also taken into account that, in formulations containing different live probiotics, certain species could inhibit the stimulatory effect of others [33,40].
Differences in the degree of stimulation of the defenses have been reported among different probiotic strains and different probiotic combinations, thus supporting the research on the active bacterial cellular components causing the variable immune stimulation [40], and identifying the levels at the intestinal level at which they can interact (outer, inner mucus layer, epithelium, immune cells).
2.1.1. Anti-Inflammatory Responses
Probiotic bacteria confer anti-inflammatory responses by modulating different signaling pathways [35,41]. Different anti-inflammatory effects at the intestinal level have been described with probiotics, for example, enhancement of the epithelial barrier function in the gut [19,35,42]; attenuation of barrier dysfunction due to pro-inflammatory cytokines [43]; or modulation of intestinal anti-inflammatory responses such as the expansion of the T-regulatory response, which may be relevant for its use in chronic inflammatory disease [41,44].
Probiotic lactobacilli and bifidobacteria have been shown to down-regulate the production of inflammatory mediators (such as IL-6 and TNF-α) upon exposure to pro-inflammatory compounds such as lipopolysaccharide (LPS) in intestinal epithelial cells [35,45,46] and also beyond the gut (reduction of pro-inflammatory biomarkers in plasma from patients treated with Bifidobacterium infantis) [47].
2.1.2. Enhancement of the Epithelial Barrier Integrity
The intestinal barrier is a major defense mechanism, including the mucous layer, antimicrobial peptides, secretory IgA, and the epithelial junction adhesion complex, with the aim to maintain epithelial integrity and to protect the organism from the environment, including pathogens [19].
Probiotic bacteria have been studied for their involvement in reinforcing the intestinal barrier, although the mechanisms involved are not fully elucidated [19,48]. Probiotics may initiate repair of the barrier function after damage induced by different pathological conditions, such as E. coli-induced mucosal disruption [19,49]. Probiotics can also prevent the cytokine-induced epithelial damage, which is characteristic of inflammatory bowel disease [19,50].
On the intestinal barrier, some strains can also block pathogen entry into the epithelial cells by increasing the mucus barrier, by stimulating the release of mucin granules from Goblet cells; and by maintaining the intestinal permeability, by increasing the intercellular integrity of apical tight junctions [19,51,52].
2.1.3. TLR-2 Receptor
Toll-like receptors (TLR) are a family of 11 transmembrane proteins (TLR-1-TLR11) expressed on various immune and non-immune cells that recognize specific patterns of microbial components and regulate the activation of both innate and adaptive immunity [19]. The existence of several TLRs enables the innate immunity to recognize different groups of pathogens, while initiating appropriate and distinct immunological responses [53]. In humans, TLR1, TLR2, TLR4, TLR5, TLR6, and TLR10 primarily respond to bacterial surface-associated pathogen-associated molecular patterns (PAMPs) [19].
Several studies have demonstrated that Toll-like receptor-2 (TLR2) is required for probiotic strains to exert their immunomodulatory and anti-inflammatory effects [19], while stimulation of TLR4 can induce potent inflammatory responses [53].
Stimulation of TLR2 is particularly important for regulating inflammatory signaling pathways for Gram-positive bacteria [35,45] and has an important role in enhancing transepithelial resistance to invading bacteria [19,54]. Conversely, mutations in the TLR2 gene appear to be associated to severe inflammatory bowel disease [54,55]. While TLR4 recognizes Gram-negative bacteria components, such as LPS, TLR2 recognizes cell wall components of Gram-positive bacteria, including probiotic bacteria [54].
Therefore, stimulation of TLR2 by probiotic bacteria can be one of the keys for the favorable effects of probiotics at the intestinal level, leading to anti-inflammatory states that enhance the intestinal barrier integrity.
2.1.4. NLRP3 Inflammasome
Nucleotide-binding oligomerization domain-containing protein (NOD)-like receptors (NLRs) are also known to transmit signals upon interaction with gut microbiota [19]. In particular, NLRP3 inflammasome is important to the maintenance of epithelial integrity [56] and the defense against pathogen infection in the intestine [57].
2.2. Protective Effects against Pathogenic Bacteria
Gut microbiota has a relevant role fortifying the epithelial barrier against enteric pathogens [58]. Probiotics including Lactobacillus and other lactic acid bacteria, such as Streptococcus thermophilus, and bifidobacteria have been shown to inhibit a broad range of enteropathogens, including E. coli, Salmonella, Helicobacter pylori, Listeria monocytogenes, and rotavirus [19,59,60,61].
Multiple direct anti-pathogen effects have been described including inhibition of pathogens growth with the production of antimicrobial compounds, resource competition, counteracting of toxin effects, inhibition of virulence, anti-adhesive and anti-invasive effects, and competitive exclusion by competition for binding sites or stimulation of epithelial barrier function [19,62]. Probiotic strains of Lactobacillus species have also been shown to reduce the biofilm formation in pathogenic bacteria, such as Listeria monocytogenes, through competition, exclusion, and displacement [63].
Competition for binding sites on host cells is common between lactobacilli/bifidobacterial and some enteropathogens, since they share carbohydrate-binding specificities. Steric hindrance at the intestinal level is an anti-attachment mechanism described in probiotic strains against pathogenic bacteria [19,59].
Probiotics can produce a wide range of antimicrobial substances, including lactic and acetic acids [19,39,64], ethanol [61], bacteriocins [62,63], and other antimicrobial compounds, such as reuterin [61].
Probiotics also have a role against viral pathogens [39,65]. It has been shown that Bifidobacterium breve and different Lactobacillus species can inhibit the absorption of the virus to the intestinal cells [39], mainly by steric hindrance or fortifying the mucosal epithelial barrier [39] or by competition for viral receptors on enterocytes [39,66]. Probiotics have also anti-fungal properties, for example Lactobacillus reuteri against Candida growth [61,67].
2.3 Other Activities
Other related activities have been reported in different probiotic strains, including antioxidant activity, anticarcinogenic properties, inhibition of α-glucosidase, or cholesterol lowering effects [46,62,68], due to the potential of probiotics to biosynthesize health-promoting compounds, such as vitamins (B vitamins), gamma-aminobutyric acid (GABA), bioactive peptides, or conjugated linoleic acid [15].
3. Safety Issues Regarding the Use of Live Probiotics
Despite their widespread use and the large body of evidence supporting the use of probiotic supplementation in different conditions, several concerns have been raised about the possibility of adverse events associated with the use of live strains, particularly in the pediatric populations and in adults with underlying diseases [32,36,69,70].
One of the main concerns about the use of live probiotics is that live bacteria may translocate from the intestine to the locally-draining tissues and blood, thereby causing bacteremia, particularly in immunocompromised, critically-ill subjects and in the pediatric populations [32,52].
Other concerns with the use of live probiotics are the possible acquisition/transmission of antibiotic resistance genes by the probiotic strains via horizontal gene transfer in the human digestive tract [14,71,72], the presence of deleterious metabolic activities, and the excessive immune stimulation in susceptible individuals [69]. Strict assessment of the probiotic strains before marketing of the product should be performed, including genome strain characterization, to assure, among others, the absence of resistance determinants [73]. A recent study has shown the ability of food-borne Lactobacillus in diffusing their antibiotic resistance traits to food pathogens under in vitro and in vivo conditions, thus raising concern of their use as probiotics or food supplements [74].
In the case of neonates, there is also concern that live probiotic strains may form a persistent colony that could prevent normal colonization with other microbiota or with the normal core microbiome in the GI tract, with subsequent alteration of normal immune system development [32,75]. In this regard, a combination of probiotic strains instead of a single strain has been proposed in neonates, taking into account the complexity of gut microbiome and the pathogenesis of certain diseases in preterm infants, such as necrotizing enterocolitis (NEC) [76]. The use of heat-killed probiotics (S. thermophilus) in enteral formula in pre-term infants has also been proposed to avoid interference with gut colonization [77].
While adverse events associated with the use of live probiotics are mainly described in case reports, in the design of randomized controlled trials key safety parameters often lack, as concluded in a recent review of 384 randomized controlled trials assessing probiotics, prebiotics, or symbiotics, recommending that an evaluation of the benefit–risk balance should always be included [24].
This benefit–risk balance is particularly important in vulnerable patients, as concluded in a systematic review of randomized controlled trials in which probiotics were used for the prevention of Clostridium difficile-associated diarrhea (CDAD) in adults and children. The short-term administration of probiotics appeared to be safe and effective in combination with antibiotics in patients who were not immunocompromised or severely debilitated, thus concluding that vulnerable patients should be informed of the potential benefits and risks of probiotics [23].
One important concern of safety of probiotic products is the risk of translocation and the subsequent bacteremia and sepsis. Some strains have good adherence properties on the intestinal mucosa, a mechanism associated with higher probability of bacterial translocation from gut to blood and other tissues, particularly in patients with epithelial barrier dysfunction [11,68]. This risk is of particular concern in neonates, particularly in critically ill and/or extremely preterm neonates with potentially compromised gut integrity, as described in case reports [14,26,78,79], and animal models in which the presence of immune deficiency in neonates may put them at particularly high risk of probiotic sepsis [11,80].
Although none of the randomized clinical trials have reported probiotic sepsis, there are case reports of serious infections such as septicemia, pneumonia, meningitis, endocarditis, and abscess in patients treated with different probiotics, including Lactobacillus, Bifidobacterium, Bacillus, and Streptococcus, particularly in children and adults with underlying diseases [14,69].
Since bacteremia due to probiotics usually occurs in intensive care settings, hand hygiene is recommended when manipulating central venous catheters and handling probiotic preparations [69].
In this regard, the most common adverse event associated with probiotics is fungemia in patients treated with yeast preparations (containing Saccharomyces cerevisiae/Saccharomyces cerevisiae boulardii), particularly in critically ill patients, with severe systemic gastrointestinal disease or immunosuppressed [70,81,82].
Recently, two case reports of fungemia after probiotic treatment with yeast probiotics have been published [70,83].
An eight-year-old patient in a pediatric surgical intensive care unit developed S. cerevisiae fungemia following treatment containing the yeast [83] and a case of fungemia due to Saccharomyces cerevisiae var. boulardii has been reported in an immunocompromised 73-year-old patient on chemotherapy and on treatment with a probiotic product (Floratil®, containing 0.5 × 109 cells of Saccharomyces cerevisiae var. boulardii/capsule) for the management of antibiotic-associated pseudomembranous colitis [70]. Translocation of the yeast from the gastrointestinal tract to the blood was proposed as the most likely mechanism [70].
Based on this information, safety issues with the use of live probiotics, including yeasts, should always be in mind in the clinical practice, particularly in neonates, and critically ill or immunosuppressed patients [39,51], and clinical guidelines should also include safety considerations. Published safety data regarding the broad range of probiotic strains added to food or feed in food are periodically compiled by the European Food Safety Authority (EFSA) (EFSA Scientific Opinion, 2016).
4. Characteristics of Heat-Killed Bacteria with Health Benefits, Including Tyndallized Bacteria
Inactivation of probiotics can be achieved by different methods, including heat, chemicals (e.g., formalin), gamma or ultraviolet rays, and sonication, with heat treatment being the method of choice for inactivation of probiotic strains in most cases [14,27,84].
Different methods of inactivation may affect structural components of the cell differently, and influence their biological activities [14,27].
As reviewed in this article, after heat treatment, industrially-grown probiotic bacteria, including bacterial extracts and supernatants in most cases, maintain their main probiotic properties at the intestinal level, thus allowing the development of safer preparations with more optimal pharmaceutical properties (long shelf-lives, etc.) [27,59,85].
Heat treatments of bacterial suspensions can use a range of temperatures between 70 and 100 °C and in some cases, inactivation is obtained with the combination of heat treatments with incubation periods at lower temperatures (ambient temperatures, cooling or freezing temperatures), a process known as tyndallization, due to the similarities with the method of sterilization to remove spores based on repeating boiling and incubation, developed by the physicist Dr John Tyndall during the nineteenth century [86,87].
A modified tyndallization process has been used to produce heat-treated industrially-grown bacteria for different uses [30,85,86]. In most of cases, the tyndallized product contains cell fractions and supernatants [85], thus taking profit of both cell structures and excreted bacterial factors. Research studies are necessary to assess the influence of the tyndallization process on the bacterial cells, since the cell structure and cell components can be disrupted/graded to different extents. In L. rhamnosus strains, it has been reported that the tyndallization process altered the cell form, with the presence of shrunk and fragmented cells (Figure 2) [85]. Moreover, tyndallization and other heat-treatments can lead to rupture of cell walls, with the release of cytoplasmic contents (bacterial lysates), such as DNA; and cell wall components, such as peptidoglycans, lipoteichoic acids, or heat labile pili. The released bacterial components play key immunomodulating roles [27] and can also have a role in the inhibition of pathogens [28,29].
To date; however, there is limited research on the effects that different types of inactivation treatments have on bacterial structure and components and on maintenance of probiotic properties, both qualitatively and quantitatively [27,88].
At the clinical level, there is currently increasing interest in the use of heat-killed preparations of different probiotic strains, from lactic acid bacteria and Bifidobacterium, in the management of a variety of diseases [84], mainly intestinal [30], but also for other diseases, for example, as support in Helicobacter therapy [59,89], allergic respiratory diseases [90], or topical diseases [85].
To date; however, the use of products containing heat-killed bacteria with health benefits is not completely widespread. Medical devices containing different tyndallized strains in combination with mucosal protectors, such as xyloglucan or gelatin tannate, are being recently marketed for the treatment of colic in children and adults (for example, xyloglucan plus tyndallized L. reuteri and B. breve strains) and for the treatment of diarrhea and for the prevention of gut dysbiosis associated to diarrhea (gelatin tannate plus tyndallized Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus casei, Lactobacillus rhamnosus, Bifidobacterium bifidum and Streptococcus thermophilus). In these products, synergism between mucosal protectors and probiotic strains are sought in terms of immunomodulation, cell barrier integrity, and competition against pathogens. Tyndallized Lactobacillus acidophilus HA122 (2 × 109 CFU/2 mL), in combination with extracts of Matricaria chamomilla and Melissa officinalis, is also marketed for the treatment of infantile colic.
5. Bacterial Cell Lysis as a Pre-Requisite for the Physiological Effects of Probiotics
Contrary to what is commonly believed, bacterial viability or bacterial cell wall integrity is not an essential condition for the intestinal effects of probiotics, as reviewed in the next section of this manuscript. In fact, key molecules from gut bacteria, including LPS or peptidoglycan, interact with eukaryotic receptors when they are released into the environment from disrupted or completely-lysed cells or during the bacterial growth process [91,92,93]. It has been recently shown that the degradation and lysis of bacteria by lysozyme enhance the release of bacterial products, including peptidoglycan, that activate pattern recognition receptors in host cells, this being the process important for the resolution of inflammation at mucosal sites [93].
This is also supported by the localization of gut microbiota in the colon in the absence of mucosal damage, mainly present in the outer mucus layer, which offer nutrients, and distanced from enterocytes by a firmer inner structure, which is almost devoid of bacteria and confers protection to the host [94] (Figure 3). Only certain types of bacteria, for example, Proteobacteria (including enterobacteria), are able to penetrate the mucus layers and reside in close proximity to the host cells [94]. Therefore, in this scenario, one can speculate that the probiotic effects, exerted by both gut microbiota in normal conditions or by probiotics taken from supplements, are mainly derived from the release of bacterial products, which can pass through the mucus and stimulate the epithelial cells more directly than whole cells can [68]. Therefore, in comparison with live bacteria, the use of heat-killed bacteria, providing disrupted cells and released bacterial components, could better reproduce in vivo the physiological conditions in the gut lumen and outer mucus layer, with key components reaching eukaryotic cells and enhancing the mucosal integrity.
Additionally, taking into account the gut biogeography of gut microbiota, with three defined levels (lumen, outer mucus, and inner mucus) (Figure 3), the passage of active components from heat-killed probiotics to reach the epithelium seems to be a gradual process, where not all molecules would reach the eukaryotic receptors in vivo. Therefore, one can speculate that the benefits would concentrate in the apical side of the mucosa, maintaining its integrity. Moreover, the immunomodulatory properties of probiotics observed in in vitro models using different immune cells would probably be reduced in the human intestine in vivo, in which the mucus layers in the colon create a boundary between the gut lumen and the host tissue [94].
6. Effects of Probiotics as Heat-Killed Bacteria
Different strains, including lactic acid bacteria and bifidobacteria, are able to produce beneficial effects in their heat-inactivated form [14]. There is also considerable data showing that not only dead cells, but also metabolites, cell fractions, and culture supernatants of probiotic bacteria can exert biological effects [22,27,32,95]. The use of them is based on the evidence suggesting that individual effector molecules interacting with host cells may underlie probiotic effects [35,96,97]. Although similar benefits can be obtained with the different strategies—live, heat-inactivated, or different fractions [98]—relevant differences could exist among all of them [27].
For example, while live probiotics can have difficulties in attaching to intestinal epithelial cells to modulate immune responses, due to the mucous layer that avoids direct contact between bacteria and epithelial cells, microbial products can pass through the mucus and stimulate epithelial cells more directly [68]. In any case, the mechanisms by which non-viable bacteria and different bacteria fractions can exert their effects need further research.
Various microbiological components, such as cell-free supernatants [68], exopolysaccharides (EPS) [99], teichoic and lipoteichoic acids [35,100], peptidoglycans, LPS [91], and metabolites (De Marco et al., 2018) have anti-inflammatory and immunomodulating activities, through stimulating the innate immune system (Adams, 2010), the adaptive responses [101] and through their effect on the integrity of the intestinal mucous membrane [19,35]. Heat-killed probiotics are also able to antagonize pathogens (with antimicrobial compounds and by competition with pathogens for adhesion and colonization) [35,99].
These specific components are usually active on Toll-like and other signal transduction receptors in the intestinal epithelium, dendritic cells, and other immune intestinal cells [32].
In this section we review the immunomodulating effects and competition activities against pathogens of both heat-killed preparations of beneficial bacteria and purified cell-wall components, such as lipoteichoic acids, peptidoglycan, or EPS. The main effects of heat-killed probiotics and supernatant fractions are summarized in Table 2 and Table 3.
6.1. Immunomodulating Effects of Heat-Killed Probiotics and Purified Components
6.1.1. Heat-Killed Bacteria
A body of evidence indicates that inactivated bacteria have immunomodulatory effects, which can be similar to that observed with live bacteria [27]. Interestingly, inactivation, with the subsequent loss of viability and cell lysis, can produce further and more complex immunomodulation than expected [27].
Lactic Acid Bacteria
Lactic acid bacteria can modulate immune responses, with the induction of IL-12 secretion that enhance the innate immunity [38].
In a recent study in mice, immune responses induced by different heat-killed Lactobacillus species were compared, indicating that L. paracasei had the highest capacity to induce IL-12 secretion in comparison with other Lactobacillus species, including L. reuteri, L. casei, and L. plantarum [38].
Combination of heat-killed multispecies of lactic acid bacteria have also been tested (including L. acidophilus, L. plantarum, L. fermentum, and Enterococcus faecium). Enhanced immunomodulatory activity in mouse macrophages was reported in comparison with the same combination containing live strains [102]. Heat-treatment at 100 °C for 30 min did not alter the capacity of these strains to adhere to Caco-2 cells, while treatment at 121 °C for 15 min reduced more than 50% of their adherent capacity [102].
Heat-killed probiotic strains also maintain their capacity to induce secretory IgA production, as demonstrated in fecal samples from pre-term infants treated with a formulation containing heat-killed S. thermophilus [77].
Heat-killed probiotic bacteria have also been shown to have an effect in the maintenance of barrier integrity. For example, heat-killed L. rhamnosus, strain OLL2838, has been shown to protect against mucosal barrier permeability defects in mice with induced colitis [103]. In Caco-2/TC7 cell monolayers infected with diarrheagenic, diffusely adhering Afa/Dr E. coli C1845, heat-killed L. acidophilus LB plus its culture medium counteracted E. coli-induced increase in paracellular permeability [104].
In a study in rats with acute alcohol intestinal injury, the administration of heat-killed bacteria of the probiotic product VSL≠3, containing B. breve, B. longum, B. infantis, L. acidophilus, L. plantarum, L. paracasei, L. bulgaricus, and S. thermophilus, significantly protected the cyto-architecture of the intestinal barrier, preventing passage of endotoxin and other bacterial products from the gut lumen into the portal circulation and down-regulating the expression of TNF-α [105].
Bifidobacterium
In a comparison between live and heat-killed B. breve M-16-V, both forms showed immunomodulating effects that suppressed pro-inflammatory cytokine production [106].
Heat inactivated B. bifidum OLB6378 can also act on sIgA production, as observed in a mouse intestinal explant model, being the result of a direct microbial effect on the intestinal epithelium [107].
6.1.2. Cell Wall Components
Currently, there is increasing interest to understand the biological activities of cell wall components of probiotic bacteria in the design of new advanced therapeutics and to avoid the use of live microorganisms [28]. In the case of the development of products containing heat-killed strains, the identification of key cell wall components is also necessary, together with the assurance that these molecules maintain their activity after the heat treatment.
Despite their biological importance, cell wall components of probiotics are poorly characterized [120]. Peptidoglycan and lipoteichoic acids are the major cell wall components of Gram-positive bacteria and can be considered the pivotal components for the immunomodulating effects of most probiotics [35,97,120]. While lipoteichoic acids and peptidoglycan from Lactobacillus species have been associated with immunomodulating effects in different models [35,41,121], in the case of bifidobacteria, the immunomodulating roles of these molecules have not yet been properly studied [28].
Lipoteichoic Acids
The role of lipoteichoic acids as IL-12 inducers, thus activating the innate immune functions, have been demonstrated in L. plantarum in cultures of mouse spleen cells and splenic dendritic cells [121]. Lipoteichoic acid from L. plantarum also confers anti-inflammatory responses, as observed in a study in porcine intestinal epithelial cell lines. Of note, lipoteichoic acids, suppressed poly I:C-induced IL-8 production, suggesting the capacity of these molecules to inhibit viral pathogen-induced inflammatory responses in intestinal epithelial models [35].
Peptidoglycans
Peptidoglycan from L. rhamnosus has been shown to improve innate immune responses in immunocompromised-malnourished mice after Streptococcus pneumoniae infection. Moreover, nasal administration of this molecule improved innate immune responses and induced respiratory and systemic adaptive human responses [122]. Peptidoglycans from different Lactobacillus species have also the capacity to inhibit the release of inflammatory cytokines in models of LPS-induced macrophage-like cells [123].
6.1.3. Exopolysaccharides and Surface-Layer Proteins
Exopolysaccharides
Exopolysaccharides (EPS) are secreted and extracellular surface carbohydrate polymers, which can be loosely attached to the bacterial cell surface or released into the surrounding cell environment [29,124]. Present in most bacteria, they act as a protective surface layer, and also interact with the surrounding environment [29], mainly in bacterial biofilm formation, in which the EPS can be produced within individual bacterial strains and also by different species [125].
A wide variety of EPS functions have been characterized in probiotic bacteria, including immunomodulating and pathogen protection properties [29,124]. Due to their biological functions and physicochemical properties, bacterial EPS are being extensively studied due to their potential applications at the industrial, food, cosmetic, or medical levels [29,126]. A growing number of studies are reporting in vivo and in vitro immunomodulating effects of EPS from strains of Bifidobacterium and Lactobacillus [29]. EPS has been suggested to be involved in the cross-talk between probiotic bacteria and host immune system, potentially playing a role in intestinal homeostasis via interaction with intestinal epithelial cells [28,127].
In the case of B. breve, the immunomodulating role of EPS has been demonstrated by comparison between EPS-positive and EPS-deficient strains [29,124]. In Lactobacillus species, different EPS have exhibited immunomodulatory effects in cultures of immune cells, while only limited studies have reported their interaction with intestinal epithelial cells [127].
Surface-Layer Proteins
Surface-layers are paracrystalline dimensional arrays of proteins and glycoproteins that overlay the cell surface of several genus and species of Bacteria and Archaea, forming a symmetric, porous layer that completely covers the cell surface [128].
6.1.4. Cell-Free Supernatants and Soluble Factors
Cell-free supernatants contain batch culture medium, metabolites, and other secreted products that can cross the mucus layer and reach the intestinal monolayer of epithelial cells and interact with mucosal immune cells [95,110]. Probiotic metabolites have anti-inflammatory and antioxidant activity, acting first on intestinal epithelial cells and then on immune cells, with differences depending on the probiotic strain [68]. Reduction of the production of pro-inflammatory mediators have been demonstrated in in vitro models of immune cells upon exposure to secreted products from Lactobacillus [68,108] and Bifidobacterium species [68,110].
In a study with different probiotic strains (including L. delbrueckii, L. paracasei, L. salivarius, L. reuteri, L. rhamnosus, L. acidophilus, L. plantarum, L. lactis, L. casei, S. thermophilus, B. breve, and B. longum) in peripheral blood mononuclear cells (PBMC), the anti-inflammatory immune responses observed were mediated by both metabolites and cell-surfaces of these bacteria [130]. In models of colon epithelial cells, soluble purified peptides secreted by L. rhamnosus GG have prevented cytokine-induced cell apoptosis, thus promoting intestinal epithelial homeostasis [19,109], and cell-free supernatants of L. acidophilus, L. casei, and L. reuteri, containing metabolites, were able to downregulate the expression of PGE-2 and IL-8 [68].
Identification of the key metabolites with immunomodulating effects present in cell-free supernatants would deserve further research.
6.2. Protective Effects against Pathogens of Heat-Killed Probiotics and Purified Components
Protection against pathogens, by the production of substances (metabolites and bacteriocins), preventing pathogens adhesion and invasion, and also preventing biofilm formation by pathogenic bacteria, has also been described in heat-killed bacteria, in cell-free supernatants, and in purified compounds, particularly EPS [29,99], thus supporting their use as an alternative strategy to live probiotics.
6.2.1. Heat-Killed Probiotics
Competition for adhesion sites at th gastrointestinal level has been described between heat-killed cells/purified structures from Lactobacillus and gastrointestinal pathogens, such as diarrheagenic E. coli (ETEC) [111], Campylobacter [112], and H. pylori [59,89].
In a mice model of Salmonella infection, the combination of heat-killed multispecies of lactic acid bacteria (including L. acidophilus, L. plantarum, L. fermentum, and Enterococcus faecium) was able to reduce Salmonella invasion and the induced inflammation [102], this being the effect attributed to lipoteichoic acids and EPS [102]. Heat-killed L. plantarum also protected against Salmonella infection in mice and reduced translocation of this pathogen into different organs, such as spleen or liver, mainly by inhibiting pathogen adhesion and invasion [113].
Heat-killed lactobacilli has also exhibited activity against H. pylori. In vitro, heat-killed Lactobacillus johnsonii inhibited the growth of H. pylori. Moreover, the number of H. pylori in the infected stomach of germ-free mice was significantly decreased by the repeated oral administration of the heat-killed strain, with deformations in H. pylori cells being observed (disappearance of spiral, bending of cell body, coccoid formation, degradations, etc.) [89].
6.2.2. Cell Wall Components
Cell Wall Polysaccharides
6.2.3. Exopolysaccharides and Surface-Layer Proteins
EPS
Protection against pathogens has been described in purified EPS from lactic acid bacteria and bifidobacteria [28,29], through their anti-adhesive properties against pathogens (mainly enterobacteria) and also through the stimulation of the immune response against pathogens. EPS has also been shown to decrease the cytotoxic effects of bacterial toxins in Caco-2 cells [132]. In fact, some authors postulate that these protective actions of EPS-producing probiotics could be related to the formation of a protective film, preserving the host cells against injury, for example, by pathogens or their toxins [29]. Moreover, EPS from bifidobacteria has been shown to facilitate the growth of lactobacilli along with other anaerobic bacteria [28].
Bifidobacteria strains are popularly associated with EPS, with high structural diversity among strains. EPS form an interfacial layer separating the bacteria from its surrounding environment, considerably contributing to their anti-pathogenic activity [28]. In animal studies, the administration of B. breve, producing EPS, reduced colonization of Citrobacter rodentium, in comparison with the mutant strain [124,133]. EPS isolated from B. bifidum facilitated the growth of lactobacilli and other anaerobic bacteria and inhibited the growth of enterobacteria, enterococci, and Bacteroides fragilis. EPS from B. longum also inhibited pathogenic bacteria growth, including E. coli, Salmonella, S. aureus, B. subtilis, and B. cereus [134].
S. thermophilus CRL1190 strain reduced H. pylori adhesion and attenuated inflammatory response in AGS cells, being the first demonstration of the capacity of this strain to adhere to the stomach gastric mucosa, and to improve protection against H. pylori, being these effects attributed to the EPS [135]. These characteristics convert different EPS in promising candidates in developing functional food and medical devices for the management of different diseases [29,99].
The antagonistic effect of isolated EPS from lactobacilli has also been assessed in in vitro and in vivo studies [29]. Purified EPS from L. plantarum WLPL04, consisting of xylose, glucose and galactose, was able to inhibit the adhesion of E. coli O157:H7 to HT-29 cells in competition, replacement, and inhibition assays. Additionally, the EPS exhibited strong inhibition against biofilm formation by pathogenic bacteria, including Pseudomonas aeruginosa, E. coli O157:H7, Salmonella, and Staphylococcus aureus [99].
In fact, EPS molecules from probiotics would have structural and biological similarities to other non-bacterial polymers, for example, xyloglucan, a vegetal polymer (from the seeds of Tamarindus indica) contained in different medical devices and currently used in the management of different gastrointestinal diseases [4]. Since xyloglucan also has protective film-forming properties against E. coli or Salmonella [4], synergism can exist with heat-killed probiotics, thus supporting their combined use in gastrointestinal diseases, as is the case of medical devices containing heat-killed probiotics and mucosal protectors (xyloglucan and also gelatin tannate).
The difficulties in the purification of EPS from bacterial cells support the use of other polymers with similar properties, for example xyloglucan [4].
S-Layer Proteins
Although poorly understood, protective properties against pathogens have been described in the case of Lactobacillus S-layer proteins [128]. Surface-layer protein extracts from Lactobacillus helveticus, strain R0052, has prevented EHEC O157:H7 binding to epithelial cells in vitro [51,136]. Exposure of epithelial cells with S-layer protein extracts decreased E. coli O157:H7 adherence and attaching-effacing lesions and preserved the epithelial barrier function [136].
6.2.4. Cell-Free Supernatants
Cell-free supernatants from probiotic bacteria contain a wide range of compounds with anti-microbial properties, including organic acids, such as lactic acid, hydrogen peroxide, diacetyl, reuterin, and bacteriocins [137,138].
The production of organic acids by multiple probiotic strains, belonging both to lactic acid bacteria and bifidobacteria, is mainly responsible for the antimicrobial activity against Gram-negative pathogens [138]. Exposure of C. difficile to filtered supernatants from S. thermophilus has shown a dose-dependent, bactericidal effect due to lactic acid [139].
Reuterin (3-hydroxypropionaldehyde) is a well-known antimicrobial metabolite produced by L. reuteri, and thought to exert its effect by oxidizing thiol groups in the target gut pathogenic microorganisms [138,140].
Secreted Bacteriocins
Bacteriocins are antibacterial small heat-stable peptides that are able to inhibit the growth of other bacteria, including enteric pathogens [39,51], (Bactibase Database http://bactibase.hammamilab.org/main.php). Exceptionally, few bacteriocins, together with their native antibacterial property, also exhibit additional anti-viral and anti-fungal properties. Bacteriocins from Gram-positive bacteria, especially from lactic acid bacteria, have been thoroughly investigated considering their great biosafety and broad industrial applications [116].
Inhibition of the in vitro growth of a broad range of pathogens, including Clostridium, Bacillus, Listeria, Enterococcus and Staphylococcus, enterobacteria, and other Gram-negative bacteria and in vivo protection against infection has been described in different lactic acid bacteria [39,116,117].
Bifidobacteria release a wide diversity of bacteriocins, being considered the main factor responsible for the antimicrobial activity of the cell-free supernatants [28]. Bifidocins, isolated from different Bifidobacterium strains, have exhibited a wide range of bactericidal activity, against Gram-positive and Gram-negative bacteria and some yeasts, through cell lysis. Another bacteriocin produced by Bifidobacterium, acidocin, has been shown to inhibit Clostridium species in fermented food products [28,118,119].
Bacteriocins and other antimicrobial compounds can be present in the heat-inactivated probiotic products, since they can resist temperatures up to 100 °C [116]. Other interesting properties of bacteriocins to be considered good candidates as possible ingredients in new-generation probiotic products are their stability in a wide pH range of 3–10, and towards the action of weak organic solvents, refrigeration, freezing, and action of salts and enzymes [28].
Nevertheless, the presence and activity of antimicrobial compounds in products containing heat-inactivated bacteria and their culture medium deserve further research.
7. Protective Barrier Properties of Tyndallized Probiotics in Combination with Mucosal Protectors in Intestinal In Vitro Models
One common property among the different strains of probiotics is their capacity to fortify the intestinal mucosal barrier [19,48]. These effects have also been observed in heat-inactivated probiotics [103] and in purified components, such as EPS (Castro-Bravo et al., 2018), and also in the group of components with mucosal protective properties such as xyloglucan and gelatin tannate [4]. Synergism between tyndallized probiotic strains and mucosal protectors have been demonstrated in in vitro models of intestinal cells.
In in vitro models of intestinal mucosa (HT29-MTX cells), the combination of tyndallized strains, including L. acidophilus, L. plantarum, L. casei, L. rhamnosus, B. bifidum, and S. thermophilus, and gelatin tannate protected intestinal cells from E. coli infection by inhibiting the adhesion and internalization of bacteria, preventing the increase of paracellular permeability and modulating cytokine gene expression [52,141].
The same combination was also assessed in E. coli-infected CacoGoblet® cells, with an increase in the transepithelial electrical resistance (TEER) and a reduction in the paracellular flux, being these effects more important than those observed with the heat-killed probiotic mixture alone, S. boulardii or the anti-diarrheal agent diosmectite. These results highlight the synergism between a mucosal protector and heat-killed probiotics to protect the intestinal barrier integrity and to prevent enteropathogens adhesion and invasion. Synergism has also been proposed in terms of onset of action, in which the presence of the mucosal protector would produce a faster onset of action of the probiotic mixture [142].
The protective properties of tyndallized probiotics plus other mucosal protectors, such as xyloglucan, should deserve further research, in intestinal cells and also in other models, as nasal epithelial cells, based on the previous studies supporting the use of xyloglucan as protector of the nasal mucosal epithelial cells [143,144]. In fact, xyloglucan in nasal formulations is an innovative strategy for the management of nasal disorders, as rhinitis and rhinosinusitis, based on their protective properties on the nasal epithelial cells, maintaining the barrier integrity and allowing the avoidance of allergens and triggering factors, as demonstrated in MucilAir™Nasal cells [4,143,144] and in patients with rhinosinusitis [145].
Although experience with probiotics for the treatment of nasal disorders is limited, recent data from patient biopsy specimens also indicate that topical heat-killed probiotics can be a safe and feasible alternative treatment, through their anti-inflammatory properties [90]. Further research; however, is needed to assess the clinical effects of heat-killed bacteria in nasal disorders and also in combination with mucosal protectors such as xyloglucan.
8. Clinical Benefits of Tyndallized Bacteria as Probiotics in Gastrointestinal Diseases
8.1. Bloating
In a recent double-blind, multicenter, randomized clinical trial in adult subjects with a diagnosis of functional bloating, the administration of a medical device containing the mucosal protector xyloglucan plus tyndallized L. reuteri and B. breve, during 20 consecutive days, produced higher symptoms relief than simethicone, particularly regarding abdominal distension and flatulence. Of note, at baseline, all subjects had a diagnosis of small intestinal bacterial overgrowth (SIBO) confirmed by the hydrogen breath test, while at the end of treatment a reduction in hydrogen gas production was observed in both treatment arms [31].
SIBO is a common gastrointestinal dysbiosis that can be caused by the overuse of certain drugs such as proton pump inhibitors. The long-term reduction of gastric secretion creates favorable conditions for the colonization of various bacterial species in the upper gastrointestinal tract [146]. Moreover, it is also known that H. pylori infection can also alter the microbiota of the upper gastrointestinal tract, and active H. pylori infection has been found to be significantly associated with the presence of SIBO [146,147].
SIBO is due to the overgrowth of species that commonly colonize the colon, mainly Gram-negative, strict anaerobes, and Enterococci [148]. Interestingly, in children with SIBO, higher counts of Salmonella have been detected in fecal samples, leading to the assumption that individuals with SIBO possibly have dysbiosis in different intestinal segments and not only in the small intestine [149].
In this context, we can speculate that, while the effect of simethicone on SIBO is through its de-foaming properties, altering the elasticity of interfaces of mucus-embedded bubbles in the gastrointestinal tract [150], the effect of the medical device in reducing SIBO and the associated symptoms would be more associated with the protective effects against pathogens produced by the probiotic strains and the mucoadhesive properties of xyloglucan, with antiadhesive properties against enterobacteria, as already demonstrated in in vitro [151] and in vivo studies [152].
8.2. Pediatric Disorders
With the use of new sequencing techniques, gut microbiota and the characteristics of dysbiosis is currently being assessed in detail in pediatric populations, particularly in infants and preterm infants. Recent findings suggest that the immature intestinal mucosa and gut dysbiosis in infants precedes the development of relevant severe diseases, as late-onset sepsis [153] or necrotizing enterocolitis (NEC) [54], and also as less severe, but particularly stressful for parents, infantile colic [87]. It has been recently shown that at weaning the intestinal microbiota induces a vigorous immune response (the “weaning reaction”) that is programmed in time, and inhibition of this effect leads to pathological imprinting and increased susceptibility to colitis, allergic inflammation, and cancer later in life [154].
B. breve is the dominant species in the gut of breast-fed infants and it has also been isolated from human milk. For this reason, strains of B. breve are widely used in pediatrics, having antimicrobial activity against enteropathogens and immunomodulatory effects. Of note, it is devoid of transmissible antibiotic resistance traits and cytotoxicity [133].
Probiotic supplementation with strains of L. reuteri, originally cultured from mother´s breast milk, endowed with immunomodulating effects, have been shown to reduce the incidence and severity of severe infant diseases, such as NEC [54] or late-onset sepsis [155], and have also been tested in infantile colic [87].
Particularly in neonates; however, it is important not to alter the gut bacterial colonization [77], thus supporting the use of heat-killed strains, for example in neonates in enteral nutrition [77] or for the management of infantile colic [30,87,156].
Infantile Colic
Infantile colic is a common condition (20% of infants) occurring during the first four months of life, defined as infant irritability, fussing, or crying that occur without obvious cause, without evidence of infant failure to thrive, fever, or ill health, presenting with recurrent prolonged periods [87,156]. To date, infantile colic pathophysiology is poorly understood, with the presence of gut microbiota dysbiosis, barrier alterations, and mild chronic gastro-intestinal inflammation [87]. Gut dysbiosis in colicky infants is characterized by decreased levels of bifidobacteria, lactobacilli, and butyrate-producing species and increased levels of Proteobacteria, leading to a more pro-inflammatory environment [87]. Moreover, intestinal mucosal immaturity has also been reported, with the possible entry of toxic compounds from the gut lumen to the blood [87,157].
This knowledge and the dissatisfaction with conventional treatment options (for example, simethicone) is opening new therapeutic strategies for the management of the disease, particularly based on the use of probiotic heat-killed strains [87,156].
In a recent pilot study in 46 infants aged three to 16 weeks with infantile colic, the administration of xyloglucan plus tyndallized Lactobacillus reuteri SGL01 and Bifidobacterium breve SGB01, at 100 × 109 CFU/g, significantly decreased the mean duration of crying episodes, in comparison with a lactase dietary supplement. These results suggest a role of the combined use of xyloglucan plus tyndallized bacteria in the management of infantile colic, although further research in larger studies is needed [30].
Administration of tyndallized Lactobacillus acidophilus HA122 (2 × 109 CFU/2 mL), in combination with extracts of Matricaria chamomilla and Melissa officinalis produced a significant reduction of the mean daily crying time in comparison with simethicone, in a recent randomized open-label controlled clinical trial in children aged between two weeks and four months old [156].
Based on these results, the use of tyndallized bacteria in combination with mucosal protectors could also be considered in the prevention strategies of the disease.
8.3. Diarrhea
Heat-killed L. acidophilus LB has been tested in adult patients with chronic diarrhea, with marked improvements in the remission of clinical symptoms at the end of treatment in comparison with live lactobacilli [158].
Heat-killed bacteria have also been tested in children with diarrhea. Lyophilized, heat-killed L. acidophilus LB was tested vs. placebo in children with acute diarrhea as an adjunct to oral rehydration therapy. After 24 h of treatment, in the L. acidophilus LB group the number of rotavirus-positive children with watery stools was significantly lower, with a significant reduction in the mean duration of diarrhea vs. placebo [159].
In a randomized, double-blind, placebo-controlled clinical trial, in selected and controlled homogeneous groups of children with well-established, non-rotavirus diarrhea, adding lyophilized, heat-killed L. acidophilus LB bacteria plus their culture medium to a solution of oral rehydration solution shortened the recovery time by one day (i.e., the time until the first normal stool was passed) as compared with children who received placebo oral rehydration solution [104].
Based on the known synergism between tyndallized bacteria and mucosal protectors, such as xyloglucan or gelatin tannate, maintaining mucosal integrity and interfering with potential pathogenic bacteria [4,52], different clinical trials could be performed with the combination of different types of gastroenteritis, as for example in the prevention of antibiotic-associated diarrhea, in diarrhea in immunocompromised children, or in gastroenteritis produced by different bacterial species.
8.4. Extra-Intestinal Diseases
Research on the benefits of inactivated bacteria is being extended to a variety of extra-intestinal diseases [165,166].
The use of oral probiotics is an attractive option for the management of allergic diseases, particularly atopic dermatitis [85], based on the observations that infants who develop atopic dermatitis have fewer probiotic bacteria in the gut than healthy controls [85,167] and that modification and stabilization of gut microbiota with the use of probiotics could improve gastrointestinal dysbiosis [51,85].
Results about the use of different live probiotics, mainly L. rhamnosus GG and B. breve and B. longum, in atopic dermatitis have generated considerable controversy in children, adults, and also during pregnancy [32,168,169], due to contrasting efficacy results and, in some cases, due to the occurrence of adverse events [32].
A body of evidence indicates that the positive effects may be related to the type of probiotic strain, the method of administration, onset time, dose, and treatment duration [35,168]. Several studies in mice have demonstrated that tyndallized L. rhamnosus and L. brevis strains can prevent the development of atopic dermatitis [170,171]. Oral administration in mice of tyndallized L. rhamnosus at 108, 109, and 1010 CFU/mL produced dose-dependent improvement in signs and symptoms of the disease, thus indicating their potential for the management of the disease [85]. In a recent study in a murine model of atopic dermatitis, the oral administration of metabolites from lactic acid bacteria improved skin injury [165].
In a multicenter, randomized, double-blind controlled trial, the use of a milk formula containing heat-killed B. breve C50 and S. thermophilus 065 in children at high risk of atopy reduced the incidence of digestive and respiratory potentially allergic events [172,173].
In fact, topical application of heat-killed probiotics, purified compounds, and also in combination with mucosal protectors, is receiving special attention and deserves further studies. The use of mucosal protectors in dermatological diseases is based on their protective barrier properties to avoid skin damage and their role in skin regeneration [4,174].
9. Concluding Remarks
Probiotics are the focus of interest at multiple levels, including consumers, patients, clinicians, scientific community, and pharmaceutical companies, and there is increasing interest to improve probiotic products, making them safer and more specific for each intended condition. In this context, new-generation probiotic products, including heat-killed strains, key components, or compounds with similar effects to living probiotic cells are being developed and already marketed, for certain indications, particularly for gastrointestinal disorders.
To date; however, there is still a number of issues to be tackled for both live probiotics and for new-generation products containing inactivated cells, cell fractions, or purified components to develop rationally-designed beneficial therapies to provide enhanced protection against infections and other diseases [17]. In general, a better understanding of the complex probiotic–pathogen interactions in the real human intestine will help to develop more specific products for each condition and to know the extent to which the bacterial-derived components are active in vivo [17,62,94,175], with a better defined benefit–risk ratio, particularly in vulnerable groups [69,70].
Currently, the use of probiotics is framed within the strategies to avoid antimicrobial resistances [4,176,177] and the need to avoid chronic pharmacological treatments and their adverse effects [4]. In fact, in the current context of high levels of antibiotic resistances, acquisition and retransfer of resistance genes should be addressed in the safety evaluation of live probiotics [71,73], and should be considered in the development of future products [73,76]. In this regard, the use of inactivated bacteria can provide important benefits, decreasing the risk of transmission of antibiotic-resistant genes.
Based on the evidence from case reports, it is clear that standard safety evaluations have to be included in randomized clinical trials assessing probiotics [24], and safety issues also have to be transmitted to health care professionals, including pharmacists in the pharmaceutical offices, where recommendation of probiotics is widespread and often obtained without medical prescription. Comorbidities and vulnerable conditions can be frequent in patients taking probiotics and; therefore, information about the possible associated risks should be given. The risk of translocation with possible systemic infections should be taken into account in vulnerable patients, and also considering certain conditions that can favor translocation, such as the presence of dysbiosis and certain conditions altering gut microbiota (for example, immunosuppression) [178].
In this regard, increasing interest is being focused on new-era products, with the use of heat-inactivated strains and purified key components responsible for the beneficial effects [28]. Purified components, such as EPS, lipoteichoic acids, metabolites, and bacteriocins, might play an important role in replacing live probiotics. In this field, more research is needed in different aspects, for example, to identify specific strains for each condition; to assess the degree of bacterial cell disruption after heat treatments (and to identify the optimal conditions that can inactivated with maintenance of the cell structure); to identify the key components of the beneficial effect for a certain strain; and to test the synergism of different combinations, which could include different heat-inactivated strains and purified key components, as well as mucosal protectors, with protective barrier properties. Moreover, the in vitro results and animal models should be interpreted considering the particular conditions of the human intestine, particularly in the colon, with a stratified layer structure where gut microbiota is mainly present in the outer layer. The physiological effect that heat-killed strains and their release compounds can exert in vivo should be also taken into account, since a substantial presence of disrupted cells or released compounds in the outer mucus layers seems to be the most probable situation, rather than a predominant direct contact with the epithelial cells.
Anyway, results reviewed in this article have shown that tyndallized bacteria clearly have favorable effects at the clinical level in the management of different diseases, representing a new generation of safer and more stable products.
As we have reviewed, the presence of key structures in the cell or supernatant fractions is able to confer probiotic properties, mainly through immune-modulation, protection against pathogens, and fortifying the mucosal barrier integrity. For the next generation products, the purification of these components and quantification of these effects would probably allow more standardization, leading to high specific and safe products intended for patient-tailored therapies. To compare and standardize these products, common activities among probiotic strains could be assessed, for example, their capacity to maintain mucosal integrity.
Current existing evidence of heat-killed bacteria in relation to health benefits indicates that they can be safe alternatives to live probiotics in vulnerable populations, such as neonates [77], and also have a role in the management of gastrointestinal disorders in children and adults, including bloating and diarrhea [30,104,156,158,159]. The synergism between tyndallized bacteria and mucosal protectors has been demonstrated in patients with bloating [30,31], while the role of this combination in other intestinal diseases and also in extra-intestinal diseases could also be explored.
This is the case of topical diseases, such as atopic dermatitis, with a demonstrated relationship to environmental pollution, and related to skin barrier dysfunction [179]. The topical use of heat-killed probiotic bacteria and mucosal protectors could provide benefits for the management of this disease, taking into account that, to date, the benefits provided by some topical protection creams are under debate [180].
Topical application of heat-killed bacteria could have also a role in the management of allergic respiratory diseases, based on the favorable results obtained with mucosal protectors in nasal in vitro models [142,144].
Another field that could be explored is in urinary tract infections (UTIs), based on the evidence indicating that mucosal protectors can reduce the intestinal reservoirs of uropathogenic E. coli strains [4,151,181]. The results indicating that metabolites produced by lactobacilli (hydrogen peroxide and lactic acid) act cooperatively to kill uropathogenic organisms in vitro [182,183] could be the starting point for the development of products containing heat-killed bacteria for the management of UTIs.
Preliminary in vitro data have also been obtained in H. pylori infection models, thus suggesting that heat-killed bacteria could also have a role in the prevention and treatment of H. pylori infection [89]. Although more research is needed to assess the interaction between H. pylori and probiotic strains and the role that probiotics (live or inactivated) can play in the prevention and in the support of antibiotic treatment strategies [184].
Overall, the reviewed data are indicating that alternatives to live probiotics, including heat-killed bacteria or their fractions or purified components, have key beneficial effects. These types of products offer advantages in respect to the use of live probiotics, mainly their safety profile, positioning them as interesting strategies for the management of common prevalent conditions in a wide variety of patients´ characteristics.
Author Contributions
All the authors have contributed equally to writing the original draft preparation, review, and editing.
Funding
Noventure SL funded medical writing and editorial activities.
Acknowledgments
Emma Córcoles for editing assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| EPS | Exopolysaccharides |
| NK | Natural killer |
| APCs | Antigen-presenting cells |
| Th1 | Type 1 helper T |
| LPS | Lipopolysaccharide |
| TLR2 | Toll-like receptor-2 |
| NEC | Necrotizing enterocolitis |
| CDAD | C. difficile-associated diarrhea |
| TEER | Transepithelial electrical resistance |
| SIBO | Small intestinal bacterial overgrowth |
| UTIs | Urinary tract infections |
References
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the Gut Microbiome Using 16S or Shotgun Metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Palau, M.; Kulmann, M.; Ramírez-Lázaro, M.J.; Lario, S.; Quilez, M.E.; Campo, R.; Piqué, N.; Calvet, X.; Miñana-Galbis, D. Usefulness of Housekeeping Genes for the Diagnosis of Helicobacter pylori Infection, Strain Discrimination and Detection of Multiple Infection. Helicobacter 2016, 21, 481–487. [Google Scholar] [CrossRef]
- Zhao, Y.; Jaber, V.; Lukiw, W. Secretory products of the human GI tract microbiome and their potential impact on Alzheimer´s Disease (AD): Detection of Lipopolysaccharide (LPS) in AD hippocampus. Front. Cell. Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef]
- Piqué, N.; Gómez-Guillén, M.D.C.; Montero, M.P. Xyloglucan, a Plant Polymer with Barrier Protective Properties over the Mucous Membranes: An Overview. Int. J. Mol. Sci. 2018, 19, 673. [Google Scholar] [CrossRef]
- Janssen, M.; Busch, C.; Rödiger, M.; Hamm, U. Motives of consumers following a vegan diet and their attitudes towards animal agriculture. Appetite 2016, 105, 643–651. [Google Scholar] [CrossRef]
- Nagpal, R.; Yadav, H.; Marotta, F. Gut microbiota: The next-gen frontier in preventive and therapeutic medicine? Front. Med. 2014, 1, 15. [Google Scholar] [CrossRef]
- Mosca, A.; Leclerc, M.; Hugot, J.P. Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 2016, 7, 455. [Google Scholar] [CrossRef] [Green Version]
- Crow, J.R.; Davis, S.L.; Chaykosky, D.M.; Smith, T.T.; Smith, J.M. Probiotics and Fecal Microbiota Transplant for Primary and Secondary Prevention of Clostridium difficile Infection. Pharmacotherapy 2015, 35, 1016–1025. [Google Scholar] [CrossRef]
- Wilkins, T.; Sequoia, J. Probiotics for Gastrointestinal Conditions: A Summary of the Evidence. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar]
- Dronkers, T.M.G.; Krist, L.; Van Overveld, F.J.; Rijkers, G.T. The ascent of the blessed: Regulatory issues on health effects and health claims for probiotics in Europe and the rest of the world. Benef. Microbes 2018, 9, 717–723. [Google Scholar] [CrossRef]
- Boyle, R.J.; Robins-Browne, R.M.; Tang, M.L. Probiotic use in clinical practice: What are the risks? Am. J. Clin. Nutr. 2006, 83, 1256–1264. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef]
- Deshpande, G.; Athalye-Jape, G.; Patole, S. Para-probiotics for Preterm Neonates. Next. Front. Nutr. 2018, 10, E871. [Google Scholar]
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic acid bacteria and bifidobacteria with potential to design natural biofunctional health-promoting dairy foods. Front. Microbiol. 2017, 8, 846. [Google Scholar] [CrossRef]
- Guandalini, S. Probiotics for prevention and treatment of diarrhea. J. Clin. Gastroenterol. 2011, 45, S149–S153. [Google Scholar] [CrossRef]
- Mills, J.P.; Rao, K.; Young, V.B. Probiotics for prevention of Clostridium difficile infection. Curr. Opin. Gastroenterol. 2018, 34, 3–10. [Google Scholar] [CrossRef]
- Zyrek, A.A.; Cichon, C.; Helms, S.; Enders, C.; Sonnenborn, U.; Schmidt, M.A. Molecular mechanisms underlying the probiotic effects of Escherichia coli Nissle 1917 involve ZO-2 and PKCzeta redistribution resulting in tight junction and epithelial barrier repair. Cell. Microbiol. 2007, 9, 804–816. [Google Scholar] [CrossRef]
- Bermúdez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Pamer, E.G. Resurrecting the intestinal microbiota to combat antibiotic-resistant pathogens. Science 2016, 352, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Draper, K.; Ley, C.; Parsonnet, J. Probiotic guidelines and physician practice: A cross-sectional survey and overview of the literature. Benef. Microbes 2017, 8, 507–519. [Google Scholar] [CrossRef]
- Adams, C.A. The probiotic paradox: Live and dead cells are biological response modifiers. Nutr. Res. Rev. 2010, 23, 37–46. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Yap, C.; Lytvyn, L.; Lo, C.K.; Beardsley, J.; Mertz, D.; Johnston, B.C. Probiotics for the prevention of Clostridium difficile-associated diarrhea in adults and children. Cochrane Database Syst. Rev. 2017, 12, CD006095. [Google Scholar] [CrossRef]
- Bafeta, A.; Koh, M.; Riveros, C.; Ravaud, P. Harms Reporting in Randomized Controlled Trials of Interventions Aimed at Modifying Microbiota: A Systematic Review. Ann. Intern. Med. 2018, 169, 240–247. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Ouwehand, A.C.; Reid, G.; Salminen, S.; Cabana, M.D.; Paraskevakos, G.; Leyer, G. Probiotic use in at-risk populations. J. Am. Pharm. Assoc. 2016, 56, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Ohishi, A.; Takahashi, S.; Ito, Y.; Ohishi, Y.; Tsukamoto, K.; Nanba, Y.; Ito, N.; Kakiuchi, S.; Saitoh, A.; Morotomi, M.; et al. Bifidobacterium septicemia associated with postoperative probiotic therapy in a neonate with omphalocele. J. Pediatr. 2010, 156, 679–681. [Google Scholar] [CrossRef]
- Taverniti, V.; Guglielmetti, S. The immunomodulatory properties of probiotic microorganisms beyond their viability (ghost probiotics: Proposal of paraprobiotic concept). Genes Nutr. 2011, 6, 261–274. [Google Scholar] [CrossRef]
- Sarkar, A.; Mandal, S. Bifidobacteria-Insight into clinical outcomes and mechanisms of its probiotic action. Microbiol. Res. 2016, 192, 159–171. [Google Scholar] [CrossRef]
- Castro-Bravo, N.; Wells, J.M.; Margolles, A.; Ruas-Madiedo, P. Interactions of Surface Exopolysaccharides From Bifidobacterium and Lactobacillus Within the Intestinal Environment. Front. Microbiol. 2018, 9, 2426. [Google Scholar] [CrossRef] [Green Version]
- Vandenplas, Y.; Bacarea, A.; Marusteri, M.; Bacarea, V.; M, C.; Manolache, M. Efficacy and safety of APT198K for the treatment of infantile colic: A pilot study. J. Comp. Effect Res. 2017, 6, 137–144. [Google Scholar] [CrossRef]
- Burta, O.; Iacobescu, C.; Mateescu, R.B.; Nicolaie, T.; Tiuca, N.; Pop, C.S. Efficacy and safety of APT036 versus simethicone in the treatment of functional bloating: A multicentre, randomised, double-blind, parallel group, clinical study. Transl. Gastroenterol. Hepatol. 2018, 3, 72. [Google Scholar] [CrossRef]
- Kataria, J.; Li, N.; Wynn, J.L.; Neu, J. Probiotic microbes: Do they need to be alive to be beneficial? Nutr. Rev. 2009, 67, 546–550. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Immune system stimulation by probiotic microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Kato, K.; Odamaki, T.; Mitsuyama, E.; Sugahara, H.; Xiao, J.Z.; Osawa, R. Age-Related Changes in the Composition of Gut Bifidobacterium Species. Curr. Microbiol. 2017, 74, 987–995. [Google Scholar] [CrossRef]
- Kim, K.W.; Kang, S.S.; Woo, S.J.; Park, O.J.; Ahn, K.B.; Song, K.D.; Lee, H.K.; Yun, C.H.; Han, S.H. Lipoteichoic Acid of Probiotic Lactobacillus plantarum Attenuates Poly I:C-Induced IL-8 Production in Porcine Intestinal Epithelial Cells. Front. Microbiol. 2017, 8, 1827. [Google Scholar] [CrossRef]
- Goldstein, E.J.; Tyrrell, K.L.; Citron, D.M. Lactobacillus species: Taxonomic complexity and controversial susceptibilities. Clin. Infect. Dis. 2015, 60, s98–s107. [Google Scholar] [CrossRef]
- Dimidi, E.; Christodoulides, S.; Scott, S.M.; Whelan, K. Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation. Adv. Nutr. 2017, 8, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Arai, S.; Iwabuchi, N.; Takahashi, S.; Xiao, J.Z.; Abe, F.; Hachimura, S. Orally administered heat-killed Lactobacillus paracasei MCC1849 enhances antigen-specific IgA secretion and induces follicular helper T cells in mice. PLoS ONE 2018, 13, e0199018. [Google Scholar] [CrossRef]
- do Carmo, M.S.; Santos, C.I.D.; Araújo, M.C.; Girón, J.A.; Fernandes, E.S.; Monteiro-Neto, V. Probiotics, mechanisms of action, and clinical perspectives for diarrhea management in children. Food Funct. 2018, 9, 5074–5095. [Google Scholar] [CrossRef]
- Christensen, H.R.; Frokiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef]
- Thomas, C.M.; Versalovic, J. Probiotics-host communication: Modulation of signaling pathways in the intestine. Gut Microbes 2010, 1, 148–163. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef]
- Donato, K.A.; Gareau, M.G.; Wang, Y.J.; Sherman, P.M. Lactobacillus rhamnosus GG attenuates interferon-γ and tumour necrosis factor-α-induced barrier dysfunction and pro-inflammatory signalling. Microbiology 2010, 156, 3288–3297. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and Their Health-Promoting Effects. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Noh, S.Y.; Kang, S.S.; Yun, C.H.; Han, S.H. Lipoteichoic acid from Lactobacillus plantarum inhibits Pam2CSK4-induced IL-8 production in human intestinal epithelial cells. Mol. Immunol. 2015, 64, 183–189. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y. Probiotic and anti-inflammatory potential of Lactobacillus rhamnosus 4B15 and Lactobacillus gasseri 4M13 isolated from infant feces. PLoS ONE 2018, 13, e0192021. [Google Scholar] [CrossRef]
- Groeger, D.; O’Mahony, L.; Murphy, E.F.; Bourke, J.F.; Dinan, T.G.; Kiely, B.; Shanahan, F.; Quigley, E.M. Bifidobacterium infantis 35624 modulates host inflammatory processes beyond the gut. Gut Microbes 2013, 4, 325–339. [Google Scholar] [CrossRef]
- Mujagic, Z.; de Vos, P.; Boekschoten, M.V.; Govers, C.; Pieters, H.H.; de Wit, N.J.; Bron, P.A.; Masclee, A.A.; Troost, F.J. The effects of Lactobacillus plantarum on small intestinal barrier function and mucosal gene transcription; a randomized double-blind placebo controlled trial. Sci. Rep. 2017, 7, 40128. [Google Scholar] [CrossRef]
- Hou, Q.; Ye, L.; Liu, H.; Huang, L.; Yang, Q.; Turner, J.R.; Yu, Q. Lactobacillus accelerates ISCs regeneration to protect the integrity of intestinal mucosa through activation of STAT3 signaling pathway induced by LPLs secretion of IL-22. Cell Death Differ. 2018, 25, 1657–1670. [Google Scholar] [CrossRef] [Green Version]
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 390–407. [Google Scholar] [CrossRef]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Lopetuso, L.; Graziani, C.; Guarino, A.; Lamborghini, A.; Masi, S.; Stanghellini, V. Gelatin tannate and tyndallized probiotics: A novel approach for treatment of diarrhea. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 873–883. [Google Scholar]
- Soares, J.B.; Pimentel-Nunes, P.; Roncon-Albuquerque, R.; Leite-Moreira, A. The role of lipopolysaccharide/toll-like receptor 4 signaling in chronic liver diseases. Hepatol. Int. 2010, 4, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.K.; He, B.; Wang, T.; Tran, D.Q.; Rhoads, J.M.; Liu, Y. Protective effect of Lactobacillus reuteri DSM 17938 against experimental necrotizing enterocolitis is mediated by Toll-like receptor 2. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G231–G240. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Zaki, M.H.; Boyd, K.L.; Vogel, P.; Kastan, M.B.; Lamkanfi, M.; Kanneganti, T.D. The NLRP3 inflammasome protects against loss of epithelial integrity and mortality during experimental colitis. Immunity 2010, 32, 379–391. [Google Scholar] [CrossRef]
- Seo, S.U.; Kamada, N.; Muñoz-Planillo, R.; Kim, Y.G.; Kim, D.; Koizumi, Y.; Hasegawa, M.; Himpsl, S.D.; Browne, H.P.; Lawley, T.D.; et al. Distinct Commensals Induce Interleukin-1β via NLRP3 Inflammasome in Inflammatory Monocytes to Promote Intestinal Inflammation in Response to Injury. Immunity 2015, 42, 744–755. [Google Scholar] [CrossRef]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Cristea, V.C.; Bleotu, C.; Chifiriuc, M.C.; Bezirtzoglou, E.; Lazar, V. Antagonistic activities of some Bifidobacterium sp. strains isolated from resident infant gastrointestinal microbiota on Gram-negative enteric pathogens. Anaerobe 2016, 39, 39–44. [Google Scholar] [CrossRef]
- Canducci, F.; Armuzzi, A.; Cremonini, F.; Cammarota, G.; Bartolozzi, F.; Pola, P.; Gasbarrini, G.; Gasbarrini, A. A lyophilized and inactivated culture of Lactobacillus acidophilus increases Helicobacter pylori eradication rates. Aliment. Pharmacol. Ther. 2000, 14, 1625–1629. [Google Scholar] [CrossRef]
- Evivie, S.E.; Li, B.; Ding, X.; Meng, Y.; Yu, S.; Du, J.; Xu, M.; Li, W.; Jin, D.; Huo, G.; et al. Complete Genome Sequence of Streptococcus thermophilus KLDS 3.1003, A Strain with High Antimicrobial Potential against Foodborne and Vaginal Pathogens. Front. Microbiol. 2017, 8, 1238. [Google Scholar] [CrossRef]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef]
- Surendran Nair, M.; Amalaradjou, M.A.; Venkitanarayanan, K. Antivirulence Properties of Probiotics in Combating Microbial Pathogenesis. Adv. Appl. Microbiol. 2017, 98, 1–29. [Google Scholar]
- Woo, J.; Ahn, J. Probiotic-mediated competition, exclusion and displacement in biofilm formation by food-borne pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Zalan, Z.; Hudáček, J.; Štětina, J.; Chumchalová, J.; Halasz, A. Production of organic acids by Lactobacillus strains in three different media. Eur. Food. Res. Technol. 2010, 230, 395. [Google Scholar] [CrossRef]
- Gonzalez-Ochoa, G.; Flores-Mendoza, L.K.; Icedo-Garcia, R.; Gomez-Flores, R.; Tamez-Guerra, P. Modulation of rotavirus severe gastroenteritis by the combination of probiotics and prebiotics. Arch. Microbiol. 2017, 199, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, M.G.; Prado Acosta, M.; Candurra, N.A.; Ruzal, S.M. S-layer proteins of Lactobacillus acidophilus inhibits JUNV infection. Biochem. Biophys. Res. Commun. 2012, 422, 590–595. [Google Scholar] [CrossRef]
- Jorgensen, M.R.; Kragelund, C.; Jensen, P.O.; Keller, M.K.; Twetman, S. Probiotic Lactobacillus reuteri has antifungal effects on oral Candida species in vitro. J. Oral Microbiol. 2017, 9, 1274582. [Google Scholar] [CrossRef] [PubMed]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic Cell-Free Supernatants Exhibited Anti-Inflammatory and Antioxidant Activity on Human Gut Epithelial Cells and Macrophages Stimulated with LPS. Evid. Based Complement. Altern. Med. 2018, 1756308. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef]
- Appel-da-Silva, M.C.; Narvaez, G.A.; Perez, L.R.R.; Drehmer, L.; Lewgoy, J. Saccharomyces cerevisiae var. boulardii fungemia following probiotic treatment. Med. Mycol. Case. Rep. 2017, 18, 15–17. [Google Scholar] [CrossRef]
- Mater, D.D.; Langella, P.; Corthier, G.; Flores, M. A probiotic Lactobacillus strain can acquire vancomycin resistance during digestive transit in mice. J. Mol. Microbiol. Biotechnol. 2008, 14, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Snydman, D.R. The safety of probiotics. Clin. Infect. Dis. 2008, 46, S104–S111. [Google Scholar] [CrossRef]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: Reiterating what they are and what they are not. Front. Microbiol. 2019, 10, 424. [Google Scholar] [CrossRef]
- Thumu, S.C.R.; Halami, P. Conjugal transfer of ERM(B) and multiple tet genes from Lactobacillus spp. to bacterial pathogens in animal gut, in vitro and during food fermentation. Food Res. Int. 2019, 116, 1066–1075. [Google Scholar] [CrossRef]
- Neu, J. Perinatal and neonatal manipulation of the intestinal microbiome: A note of caution. Nutr. Rev. 2007, 65, 282–285. [Google Scholar] [CrossRef]
- Aceti, A.; Beghetti, I.; Maggio, L.; Martini, S.; Faldella, G.; Corvaglia, L. Filling the Gaps: Current Research Directions for a Rational Use of Probiotics in Preterm Infants. Nutrients 2018, 10, 1472. [Google Scholar] [CrossRef]
- Campeotto, F.; Suau, A.; Kapel, N.; Magne, F.; Viallon, V.; Ferraris, L.; Waligora-Dupriet, A.J.; Soulaines, P.; Leroux, B.; Kalach, N.; et al. A fermented formula in pre-term infants: Clinical tolerance, gut microbiota, down-regulation of faecal calprotectin and up-regulation of faecal secretory IgA. Br. J. Nutr. 2011, 105, 1843–1851. [Google Scholar] [CrossRef]
- Jenke, A.; Ruf, E.M.; Hoppe, T.; Heldmann, M.; Wirth, S. Bifidobacterium septicaemia in an extremely low-birthweight infant under probiotic therapy. Arch. Dis. Child Fetal. Neonatal. 2012, 97, F217–F218. [Google Scholar] [CrossRef] [PubMed]
- Dani, C.; Coviello, C.C.; Corsini, I.I.; Arena, F.; Antonelli, A.; Rossolini, G.M. Lactobacillus Sepsis and Probiotic Therapy in Newborns: Two New Cases and Literature. Rev. AJP Rep. 2016, 6, e25–e29. [Google Scholar]
- Wagner, R.D.; Warner, T.; Roberts, L.; Farmer, J.; Balish, E. Colonization of congenitally immunodeficient mice with probiotic bacteria. Infect. Immun. 1997, 65, 3345–3351. [Google Scholar] [Green Version]
- Lherm, T.; Monet, C.; Nougière, B.; Soulier, M.; Larbi, D.; Le Gall, C.; Caen, D.; Malbrunot, C. Seven cases of fungemia with Saccharomyces boulardii in critically ill patients. Intensive. Care Med. 2002, 28, 797–801. [Google Scholar] [CrossRef]
- Lolis, N.; Veldekis, D.; Moraitou, H.; Kanavaki, S.; Velegraki, A.; Triandafyllidis, C.; Tasioudis, C.; Pefanis, A.; Pneumatikos, I. Saccharomyces boulardii fungaemia in an intensive care unit patient treated with caspofungin. Crit. Care 2008, 12, 414. [Google Scholar]
- Atici, S.; Soysal, A.; Karadeniz Cerit, K.; Yilmaz, Ş.; Aksu, B.; Kiyan, G.; Bakir, M. Catheter-related Saccharomyces cerevisiae Fungemia Following Saccharomyces boulardii Probiotic Treatment: In a child in intensive care unit and review of the literature. Med. Mycol. Case Rep. 2017, 15, 33–35. [Google Scholar] [CrossRef]
- Zorzela, L.; Ardestani, S.K.; McFarland, L.V.; Vohra, S. Is there a role for modified probiotics as beneficial microbes: A systematic review of the literature. Benef. Microbes 2017, 8, 739–754. [Google Scholar] [CrossRef]
- Lee, S.H.; Yoon, J.M.; Kim, Y.H.; Jeong, D.G.; Park, S.; Kang, D.J. Therapeutic effect of tyndallized Lactobacillus rhamnosus IDCC 3201 on atopic dermatitis mediated by down-regulation of immunoglobulin E in NC/Nga mice. Microbiol. Immunol. 2016, 60, 468–476. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.; Bang, J.; Kim, Y.; Beuchat, L.R.; Ryu, J.H. Reduction of Bacillus cereus spores in sikhye, a traditional Korean rice beverage, by modified tyndallization processes with and without carbon dioxide injection. Lett. Appl. Microbiol. 2012, 55, 218–223. [Google Scholar] [CrossRef]
- Daelemans, S.; Peeters, L.; Hauser, B.; Vandenplas, Y. Recent advances in understanding and managing infantile colic. F1000Res 2018, 7, F1000. [Google Scholar] [CrossRef]
- Ananta, E.; Knorr, D. Comparison of inactivation pathways of thermal or high pressure inactivated Lactobacillusrhamnosus ATCC 53103 by flow cytometry analysis. Food. Microbiol. 2009, 26, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Aiba, Y.; Ishikawa, H.; Tokunaga, M.; Komatsu, Y. Anti-Helicobacter pylori activity of non-living, heat-killed form of lactobacilli including Lactobacillus johnsonii No.1088. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.S.; Peres, A.G.; Mfuna Endam, L.; Cousineau, B.; Madrenas, J.; Desrosiers, M. Topical probiotics as a therapeutic alternative for chronic rhinosinusitis: A preclinical proof of concept. Am. J. Rhinol. Allergy 2016, 30, 202–205. [Google Scholar] [CrossRef]
- Piqué, N.; Miñana-Galbis, D.; Merino, S.; Tomás, J.M. The lipopolysaccharide of Aeromonas spp: Structure-activity relationships. Curr. Top. Biochem. Res. 2013, 15, 41–56. [Google Scholar]
- Lenz, J.D.; Hackett, K.T.; Dillard, J.P. A Single Dual-Function Enzyme Controls the Production of Inflammatory NOD Agonist Peptidoglycan Fragments by Neisseria gonorrhoeae. MBio 2017, 8, e01464-17. [Google Scholar] [CrossRef]
- Ragland, SA.; Criss, A. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Liu, Y.; Gibson, G.R.; Walton, G.E. An In Vitro Approach to Study Effects of Prebiotics and Probiotics on the Faecal Microbiota and Selected Immune Parameters Relevant to the Elderly. PLoS ONE 2016, 11, e0162604. [Google Scholar] [CrossRef]
- Bron, P.A.; Tomita, S.; Mercenier, A.; Kleerebezem, M. Cell surface-associated compounds of probiotic lactobacilli sustain the strain-specificity dogma. Curr. Opin. Microbiol. 2013, 16, 262–269. [Google Scholar] [CrossRef]
- Lee, I.C.; Tomita, S.; Kleerebezem, M.; Bron, P.A. The quest for probiotic effector molecules--unraveling strain specificity at the molecular level. Pharmacol. Res. 2013, 69, 61–74. [Google Scholar] [CrossRef]
- Galdeano, C.M.; Perdigón, G. Role of viability of probiotic strains in their persistence in the gut and in mucosal immune stimulation. J. Appl. Microbiol. 2004, 97, 673–681. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Qiu, L.; Zhang, F.; Xu, X.; Wei, H.; Tao, X. Characterization and bioactivities of the exopolysaccharide from a probiotic strain of Lactobacillus plantarum WLPL04. J. Dairy Sci. 2017, 100, 6895–6905. [Google Scholar] [CrossRef]
- Matsuguchi, T.; Takagi, A.; Matsuzaki, T.; Nagaoka, M.; Ishikawa, K.; Yokokura, T.; Yoshikai, Y. Lipoteichoic acids from Lactobacillus strains elicit strong tumor necrosis factor alpha-inducing activities in macrophages through Toll-like receptor 2. Clin. Diagn. Lab. Immunol. 2003, 10, 259–266. [Google Scholar] [CrossRef]
- Hirose, Y.; Murosaki, S.; Yamamoto, Y.; Yoshikai, Y.; Tsuru, T. Daily intake of heat-killed Lactobacillus plantarum L-137 augments acquired immunity in healthy adults. J. Nutr. 2006, 136, 3069–3073. [Google Scholar] [CrossRef]
- Chen, C.Y.; Tsen, H.Y.; Lin, C.L.; Lin, C.K.; Chuang, L.T.; Chen, C.S.; Chiang, Y.C. Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J. Med. Microbiol. 2013, 62, 1657–1664. [Google Scholar] [CrossRef]
- Miyauchi, E.; Morita, H.; Tanabe, S. Lactobacillus rhamnosus alleviates intestinal barrier dysfunction in part by increasing expression of zonula occludens-1 and myosin light-chain kinase in vivo. J. Dairy Sci. 2009, 92, 2400–2408. [Google Scholar] [CrossRef]
- Liévin-Le Moal, V.; Sarrazin-Davila, L.E.; Servin, A.L. An experimental study and a randomized, double-blind, placebo-controlled clinical trial to evaluate the antisecretory activity of Lactobacillus acidophilus strain LB against nonrotavirus diarrhea. Pediatrics 2007, 120, e795–e803. [Google Scholar] [CrossRef]
- Chang, B.; Sang, L.; Wang, Y.; Tong, J.; Zhang, D.; Wang, B. The protective effect of VSL#3 on intestinal permeability in a rat model of alcoholic intestinal injury. BMC Gastroenterol. 2013, 13, 151. [Google Scholar]
- Sugahara, H.; Yao, R.; Odamaki, T.; Xiao, J.Z. Differences between live and heat-killed bifidobacteria in the regulation of immune function and the intestinal environment. Benef. Microbes 2017, 8, 463–472. [Google Scholar] [CrossRef]
- Nakamura, Y.; Terahara, M.; Iwamoto, T.; Yamada, K.; Asano, M.; Kakuta, S.; Iwakura, Y.; Totsuka, M. Upregulation of Polymeric Immunoglobulin Receptor Expression by the Heat-Inactivated Potential Probiotic Bifidobacterium bifidum OLB6378 in a Mouse Intestinal Explant. Model. Scand. J. Immunol. 2012, 75, 176–183. [Google Scholar] [CrossRef]
- Griet, M.; Zelaya, H.; Mateos, M.V.; Salva, S.; Juarez, G.E.; de Valdez, G.F.; Villena, J.; Salvador, G.A.; Rodriguez, A.V. Soluble factors from Lactobacillus reuteri CRL1098 have anti-inflammatory effects in acute lung injury induced by lipopolysa, E110027. PLoS ONE 2014, 9, e110027. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef]
- Hoarau, C.; Lagaraine, C.; Martin, L.; Velge-Roussel, F.; Lebranchu, Y. Supernatant of Bifidobacterium breve induces dendritic cell maturation, activation, and survival through a Toll-like receptor 2 pathway. J. Allergy Clin. Immunol. 2006, 117, 696–702. [Google Scholar] [CrossRef]
- Chauvière, G.; Coconnier, M.H.; Kerneis, S.; Darfeuille-Michaud, A.; Joly, B.; Servin, A.L. Competitive exclusion of diarrheagenic Escherichia coli (ETEC) from human enterocyte-like Caco-2 cells by heat-killed Lactobacillus. FEMS Microbiol. Lett. 1992, 70, 213–217. [Google Scholar] [CrossRef]
- Moyen, E.N.; Bonneville, F.; Fauchère, J.L. Modification of intestinal colonization and translocation of Campylobacter jejuni by erythromycin and an extract of Lactobacillus acidophilus in axenic mice. Ann. Inst. Pasteur. Microbiol. 1986, 137A, 199–207. [Google Scholar] [CrossRef]
- Ishikawa, H.; Kutsukake, E.; Fukui, T.; Sato, I.; Shirai, T.; Kurihara, T.; Okada, N.; Danbara, H.; Toba, M.; Kohda, N.; et al. Oral administration of heat-killed Lactobacillus plantarum strain b240 protected mice against Salmonella enterica Serovar Typhimurium. Biosci. Biotechnol. Biochem. 2010, 74, 1338–1342. [Google Scholar] [CrossRef]
- Shkarupeta, M.M.; Korshunov, V.M.; Savenkova, V.T.; Pinegin, B.V. Influence of the oral administration of indigenous microorganisms on the resistance of mice to Salmonella infection. Zh. Mikrobiol. Epidemiol. Immunobiol. 1988, 7, 46–50. [Google Scholar]
- Schwendicke, F.; Horb, K.; Kneist, S.; Dörfer, C.; Paris, S. Effects of heat-inactivated Bifidobacterium BB12 on cariogenicity of Streptococcus mutans in vitro. Arch. Oral Biol. 2014, 59, 1384–1390. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial production of bacteriocins: Latest research development and applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc. Natl Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef]
- Bali, V.; Panesar, P.S.; Bera, M.B. Trends in utilization of agro-industrial byproducts for production of bacteriocins and their biopreservative applications. Crit. Rev. Biotechnol. 2016, 36, 204–214. [Google Scholar] [CrossRef]
- Liu, G.; Ren, L.; Song, Z.; Wang, C.; Sun, B. Purification and characteristics of bifidocin A, a novel bacteriocin produced by Bifidobacterium animals BB04 from centenarians’ intestine. Food Control. 2015, 50, 889–895. [Google Scholar] [CrossRef]
- Vinogradov, E.; Sadovskaya, I.; Grard, T.; Chapot-Chartier, M.P. Structural studies of the rhamnose-rich cell wall polysaccharide of Lactobacillus casei BL23. Carbohydr. Res. 2016, 435, 156–161. [Google Scholar] [CrossRef]
- Hirose, Y.; Murosaki, S.; Fujiki, T.; Yamamoto, Y.; Yoshikai, Y.; Yamashita, M. Lipoteichoic acids on Lactobacillus plantarum cell surfaces correlate with induction of interleukin-12p40 production. Microbiol. Immunol. 2010, 54, 143–151. [Google Scholar] [CrossRef]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the immunomodulatory properties of Lactobacillus rhamnosus CRL1505 peptidoglycancommon for all Lactobacilli during respiratory infection in malnourished mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Pan, D.; Guo, Y.; Sun, Y.; Zeng, X. Peptidoglycan diversity and anti-inflammatory capacity in Lactobacillus strains. Carbohydr. Polym. 2015, 128, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Hall, L.J.; Cronin, M.; Zomer, A.; MacSharry, J.; Goulding, D.; Motherway, M.O.; Shanahan, F.; Nally, K.; Dougan, G.; et al. Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. USA 2012, 109, 2108–2113. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, G.B.; Marmont, L.S.; Howell, P.L. Enzymatic modifications of exopolysaccharides enhance bacterial persistence. Front. Microbiol. 2015, 6, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Fact. 2016, 15, 165. [Google Scholar] [CrossRef] [PubMed]
- Patten, D.A.; Leivers, S.; Chadha, M.J.; Maqsood, M.; Humphreys, P.N.; Laws, A.P.; Collett, A. The structure and immunomodulatory activity on intestinal epithelial cells of the EPSs isolated from Lactobacillus helveticus sp. Rosyjski and Lactobacillus acidophilus sp. 5e2. Carbohydr. Res. 2014, 384, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein A of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, R.; Vasiljevic, T.; Smith, S.C.; Donkor, O.N. Effect of cell-surface components and metabolites of lactic acid bacteria and probiotic organisms on cytokine production and induction of CD25 expression in human peripheral mononuclearcells. J Dairy Sci. 2014, 97, 2542–2558. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.; Muto, M.; Nomoto, K.; Matuzaki, T.; Watanabe, T.; Yokokura, T. Structure of polysaccharide-peptidoglycan complex from the cell wall of Lactobacillus caseiYIT9018. J. Biochem. 1990, 108, 568–571. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Medrano, M.; Salazar, N.; De Los Reyes-Gavilán, C.G.; Pérez, P.F.; Abraham, A.G. Exopolysaccharides produced by Lactobacillus and Bifidobacterium strains abrogate in vitro the cytotoxic effect of bacterial toxins on eukaryotic cells. J. Appl. Microbiol. 2010, 109, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Bozzi Cionci, N.; Baffoni, L.; Gaggìa, F.; Di Gioia, D. Therapeutic Microbiology: The Role of Bifidobacterium breve as Food Supplement for the Prevention/Treatment of Pediatric. Dis. Nutr. 2018, 10, E1723. [Google Scholar]
- Wu, M.H.; Pan, T.M.; Wu, Y.J.; Chang, S.J.; Chang, M.S.; Hu, C.Y. Exopolysaccharide activities from probiotic bifidobacterium: Immunomodulatory effects (on J774A.1 macrophages) and antimicrobial properties. Int. J. Food. Microbiol. 2010, 144, 104–110. [Google Scholar] [CrossRef]
- Marcial, G.; Villena, J.; Faller, G.; Hensel, A.; de Valdéz, G.F. Exopolysaccharide-producing Streptococcus thermophilus CRL1190 reduces the inflammatory response caused by Helicobacter pylori. Benef. Microbes 2017, 8, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Hagen, K.E.; Gordonpour, M.; Tompkins, T.A.; Sherman, P.M. Surface-layer protein extracts from Lactobacillus helveticus inhibit enterohaemorrhagic Escherichia coli O157:H7 adhesion to epithelial cells. Cell. Microbiol. 2007, 9, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Mariam, S.H.; Zegeye, N.; Tariku, T.; Andargie, E.; Endalafer, N.; Aseffa, A. Potential of cell-free supernatants from cultures of selected lactic acid bacteria and yeast obtained from local fermented foods as inhibitors of Listeria monocytogenes, Salmonella spp. and Staphylococcus aureus. BMC Res. Notes 2014, 7, 606. [Google Scholar] [CrossRef] [PubMed]
- Lukic, J.; Chen, V.; Strahinic, I.; Begovic, J.; Lev-Tov, H.; Davis, S.C.; Tomic-Canic, M.; Pastar, I. Probiotics or pro-healers: The role of beneficial bacteria in tissue repair. Wound. Repair. Regen. 2018, 25, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Kolling, G.L.; Wu, M.; Warren, C.A.; Durmaz, E.; Klaenhammer, T.R.; Timko, M.P.; Guerrant, R.L. Lactic acid production by Streptococcus thermophilus alters Clostridium difficile infection and in vitro Toxin A production. Gut Microbes 2012, 3, 523–529. [Google Scholar] [CrossRef]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The antimicrobial compound reuterin (3-hydroxypropionaldehyde) induces oxidative stress via interaction with thiol groups. Microbiology 2010, 156, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- De Servi, B.; Meloni, M. Antidiarrhoeal agents and paracellular permeability of E. coli-infected Caco-Goblet intestinal model. In Proceedings of the XXV Belgian Week of Gastroenterology, Ostend, Belgium, 9 February 2013. Abstract B22. [Google Scholar]
- Servi, D.B.; Ranzini, F. Protective efficacy of antidiarrheal agents in a permeability model of Escherichia coli-infected CacoGoblet® cells. Futur. Microbiol. 2017, 12, 1449–1455. [Google Scholar] [CrossRef]
- De Servi, B.; Ranzini, F.; Piqué, N. Protective barrier properties of Rhinosectan® spray (containing xyloglucan) on an organotypic 3D airway tissue model (MucilAir): Results of an in vitro study. Allergy Asthma. Clin. Immunol. 2017, 13, 37. [Google Scholar] [CrossRef]
- Piqué, N.; De Servi, B. Rhinosectan® spray (containing xyloglucan) on the ciliary function of the nasal respiratory epithelium; results of an in vitro study. Allergy Asthma Clin. Immunol. 2018, 14, 41. [Google Scholar] [CrossRef]
- Allegrini, A.; Pavone, D.; Carluccio, F.A. A randomized controlled trial comparing a xyloglucan-based nasal spray with saline in adults with symptoms of rhinosinusitis. Curr. Med. Res. Opin. 2018, 34, 377–385. [Google Scholar] [CrossRef]
- Konrad, P.; Chojnacki, J.; Gąsiorowska, A.; Rudnicki, C.; Kaczka, A.; Chojnacki, C. Therapeutic efficacy of amoxicillin and rifaximin in patients with small intestinal bacterial overgrowth and Helicobacter pylori infection. Prz. Gastroenterol. 2018, 13, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Enko, D.; Kriegshäuser, G. Functional 13C-urea and glucose hydrogen/methane breath tests reveal significant association of small intestinal bacterial overgrowth in individuals with active Helicobacter pylori infection. Clin. Biochem. 2017, 50, 46–49. [Google Scholar] [CrossRef]
- Grace, E.; Shaw, C.; Whelan, K.; Andreyev, H. Review article: Small intestinal bacterial overgrowth—Prevalence, clinical features, current and developing diagnostic tests, and treatment. Aliment. Pharmacol. Ther. 2013, 38, 674–688. [Google Scholar] [CrossRef]
- Mello, C.S.; Rodrigues, M.S.D.C.; Filho, H.B.A.; Melli, L.C.F.L.; Tahan, S.; Pignatari, A.C.C.; de Morais, M.B. Fecal microbiota analysis of children with small intestinal bacterial overgrowth among residents of an urban slum in Brazil. J. Pediatr. 2018, 94, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Azpiroz, F.; Serra, J. Treatment of Excessive Intestinal Gas. Curr. Treat. Options. Gastroenterol. 2004, 7, 299–305. [Google Scholar] [CrossRef]
- de Servi, B.; Ranzini, F.; Piqué, N. Effect of Utipro® (containing gelatin-xyloglucan) against Escherichia coli invasion of intestinal epithelial cells: Results of an in vitro study. Future Microbiol. 2016, 11, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Campolo, M.; Casili, G.; Lanza, M.; Franco, D.; Filippone, A.; Peritore, A.F.; Cuzzocrea, S. Protective Effects of Xyloglucan in Association with the Polysaccharide Gelose in an Experimental Model of Gastroenteritis and Urinary Tract Infections. Int. J. Mol. Sci. 2018, 19, 1844. [Google Scholar] [CrossRef]
- Graspeuntner, S.; Waschina, S.; Künzel, S.; Twisselmann, N.; Rausch, T.K.; Cloppenborg-Schmidt, K.; Zimmermann, J.; Viemann, D.; Herting, E.; Göpel, W.; et al. Gut dysbiosis with Bacilli dominance and accumulation of fermentation products precedes late-onset sepsis in preterm infants. Clin. Infect. Dis. 2018. [Google Scholar] [CrossRef]
- Al Nabhani, Z.; Dulauroy, S.; Marques, R.; Cousu, C.; Al Bounny, S.; Déjardin, F.; Sparwasser, T.; Bérard, M.; Cerf-Bensussan, N.; Eberl, G. A Weaning Reaction to Microbiota Is Required for Resistance to Immunopathologies in the Adult. Immunity 2019, S1074-7613(19)30081-0. [Google Scholar] [CrossRef]
- Athalye-Jape, G.; Rao, S.; Patole, S. Lactobacillus reuteri DSM 17938 as a Probiotic for Preterm Neonates: A Strain-Specific Systematic Review. JPEN. J. Parenter. Enteral. Nutr. 2016, 40, 783–794. [Google Scholar] [CrossRef]
- Martinelli, M.; Ummarino, D.; Giugliano, F.P.; Sciorio, E.; Tortora, C.; Bruzzese, D.; De Giovanni, D.; Rutigliano, I.; Valenti, S.; Romano, C.; et al. Efficacy of a standardized extract of Matricariae chamomilla L., Melissa officinalis L. and tyndallized Lactobacillus acidophilus (HA122) in infantile colic: An open randomized controlled trial. Neurogastroenterol. Motil. 2017, 29. [Google Scholar] [CrossRef]
- Halpern, M.D.; Denning, P.W. The role of intestinal epithelial barrier function in the development of NEC. Tissue Barriers 2015, 3, E1000707. [Google Scholar] [CrossRef]
- Xiao, S.D.; Zhang, D.Z.; Lu, H.; Jiang, S.H.; Liu, H.Y.; Wang, G.S.; Xu, G.M.; Zhang, Z.B.; Lin, G.J.; Wang, G.L. Multicenter, randomized, controlled trial of heat-killed Lactobacillus acidophilus LB in patients with chronic diarrhea. Adv. Ther. 2003, 20, 253–260. [Google Scholar] [CrossRef]
- Simakachorn, N.; Pichaipat, V.; Rithipornpaisarn, P.; Kongkaew, C.; Tongpradit, P.; Varavithya, W. Clinical evaluation of the addition of lyophilized, heat-killed Lactobacillus acidophilus LB to oral rehydration therapy in the treatment of acute diarrhea in children. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 68–72. [Google Scholar] [CrossRef]
- Eutamene, H.; Beaufrand, C.; Harkat, C.; Theodorou, V. The role of mucoprotectants in the management of gastrointestinal disorders. Expert. Rev. Gastroenterol. Hepatol. 2018, 12, 83–90. [Google Scholar] [CrossRef]
- Ruszczyński, M.; Urbańska, M.; Szajewska, H. Gelatin tannate for treating acute gastroenteritis: A systematic review. Ann. Gastroenterol. 2014, 27, 121–124. [Google Scholar]
- Gnessi, L.; Bacarea, V.; Marusteri, M.; Piqué, N. Xyloglucan for the treatment of acute diarrhea: Results of a randomized, controlled, open-label, parallel group, multicentre, national clinical trial. BMC Gastroenterol. 2015, 15, 153. [Google Scholar] [CrossRef]
- Plesea Condratovici, C.; Bacarea, V.; Piqué, N. Xyloglucan for the treatment of acute gastroenteritis in children: Results of a randomized controlled, clinical trial. Gastroenterol. Res. Pract. 2016, 2016, 6874207. [Google Scholar] [CrossRef]
- Kara, S.S.; Volkan, B.; Erten, İ. The therapeutic effect of gelatin tannate in acute diarrhea in children. Turk. J. Pediatr. 2017, 59, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Tokudome, Y. Influence of Oral Administration of Lactic Acid Bacteria Metabolites on Skin Barrier Function and Water Content in a Murine Model of Atopic Dermatitis. Nutrients 2018, 10, 1858. [Google Scholar] [CrossRef]
- Warda, A.; Rea, K.; Fitzgerald, P.; Hueston, C.; Gonzalez-Tortuero, E.; Dinan, T.; Hill, C. Heat-killed lactobacilli alter both microbiota composition and behaviour. Behav. Brain Res. 2019, 362, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Björkstén, B.; Sepp, E.; Julge, K.; Voor, T.; Mikelsaar, M. Allergy development and the intestinal microflora during the first year of life. J. Allergy Clin. Immunol. 2001, 108, 516–520. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Kumar, S.; Lim, J.; Paek, W.K.; Park, Y.H. Probiotics and Atopic Dermatitis: An Overview. Front. Microbiol. 2016, 7, 507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.; Ning, H.; Shen, M.; Li, J.; Zhang, J.; Chen, X. Probiotics for the Treatment of Atopic Dermatitis in Children: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Cell. Infect. Microbiol. 2017, 7, 392. [Google Scholar] [CrossRef]
- Sawada, J.; Morita, H.; Tanaka, A.; Salminen, S.; He, F.; Matsuda, H. Ingestion of heat-treated Lactobacillus rhamnosus GG prevents development of atopic dermatitis in NC/Nga mice. Clin. Exp. Allergy 2007, 37, 296–303. [Google Scholar] [CrossRef]
- Segawa, S.; Hayashi, A.; Nakakita, Y.; Kaneda, H.; Watari, J.; Yasui, H. Oral administration of heat-killed Lactobacillus brevis SBC8803 ameliorates the development of dermatitis and inhibits immunoglobulin E production in atopic dermatitis model NC/Nga mice. Biol. Pharm. Bull. 2008, 31, 884–889. [Google Scholar] [CrossRef]
- Morisset, M.; Aubert-Jacquin, C.; Soulaines, P.; Moneret-Vautrin, D.A.; Dupont, C. A non-hydrolyzed, fermented milk formula reduces digestive and respiratory events in infants at high risk of allergy. Eur. J. Clin. Nutr. 2011, 65, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Lau, S. Bacterial lysates in food allergy prevention. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 293–295. [Google Scholar] [CrossRef]
- Nie, W.; Deters, A.M. Tamarind Seed Xyloglucans Promote Proliferation and Migration of Human Skin Cells through Internalization via Stimulation of Proproliferative Signal Transduction Pathways. Dermatol. Res. Pract. 2013, 359756. [Google Scholar] [CrossRef]
- Keith, J.W.; Pamer, E.G. Enlisting commensal microbes to resist antibiotic-resistant pathogens. J. Exp. Med. 2018. [Google Scholar] [CrossRef]
- Costelloe, C.; Metcalfe, C.; Lovering, A.; Mant, D.; Hay, A.D. Effect of antibiotic prescribing in primary care on antimicrobial resistance in individual patients: Systematic review and meta-analysis. BMJ 2010, 349, c2096. [Google Scholar] [CrossRef]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Available online: http://www.who.int/medicines/publications/global-priority-list-antibiotic-resistant-bacteria/en/ (accessed on 20 July 2012).
- Zhang, Y.; Zhang, B.; Dong, L.; Chang, P. Potential of Omega-3 Polyunsaturated Fatty Acids in Managing Chemotherapy- or Radiotherapy-Related Intestinal Microbial Dysbiosis. Adv. Nutr. 2019, 10, 133–147. [Google Scholar] [CrossRef]
- David Boothe, W.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar] [PubMed]
- Schliemann, S.; Kleesz, P.; Elsner, P. Protective creams fail to prevent solvent-induced cumulative skin irritation—Results of a randomized double-blind study. Contact Dermat. 2013, 69, 363–371. [Google Scholar] [CrossRef]
- Fraile, B.; Alcover, J.; Royuela, M.; Rodríguez, D.; Chaves, C.; Palacios, R.; Piqué, N. Xyloglucan, hibiscus and propolis for the prevention of urinary tract infections: Results of in vitro studies. Futur. Microbiol. 2017, 12, 721–731. [Google Scholar] [CrossRef]
- Atassi, F.; Servin, A.L. Individual and co-operative roles of lactic acid and hydrogen peroxide in the killing activity of enteric strain Lactobacillus johnsonii NCC933 and vaginal strain Lactobacillus gasseri KS120.1 against enteric, uropathogenic and vaginosis-associated pathog. Microbiol. Lett. 2010, 304, 29–38. [Google Scholar] [CrossRef]
- Sihra, N.; Goodman, A.; Zakri, R.; Sahai, A.; Malde, S. Nonantibiotic prevention and management of recurrent urinary tract infection. Nat. Rev. Urol. 2018, 15, 750–776. [Google Scholar] [CrossRef]
- Kafshdooz, T.; Akbarzadeh, A.; Majdi Seghinsara, A.; Pourhassan, M.; Nasrabadi, H.T.; Milani, M. Role of Probiotics in Managing of Helicobacter pylori Infection: A Review. Drug Res. 2017, 67, 88–93. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Main mechanisms of action of probiotics [19].
Figure 1.
Main mechanisms of action of probiotics [19].

Figure 2.
Comparison with non-treated (left panel) and tyndallized L. rhamnosus (right panel) [85].
Figure 2.
Comparison with non-treated (left panel) and tyndallized L. rhamnosus (right panel) [85].

Figure 3.
Gut biogeography of the bacterial microbiota [94]. (a) A subset of species (green) is able to penetrate the inner mucus layer and enter crypt spaces. (b) Environmental challenges such as diet perturbations, antibiotic consumption or abnormalities in gastrointestinal motility massively alter the lumen community. However, the more stable mucosal environment and the crypts protect important bacterial species. (c) The crypts and mucosa serve as reservoirs to repopulate the lumen.
Figure 3.
Gut biogeography of the bacterial microbiota [94]. (a) A subset of species (green) is able to penetrate the inner mucus layer and enter crypt spaces. (b) Environmental challenges such as diet perturbations, antibiotic consumption or abnormalities in gastrointestinal motility massively alter the lumen community. However, the more stable mucosal environment and the crypts protect important bacterial species. (c) The crypts and mucosa serve as reservoirs to repopulate the lumen.


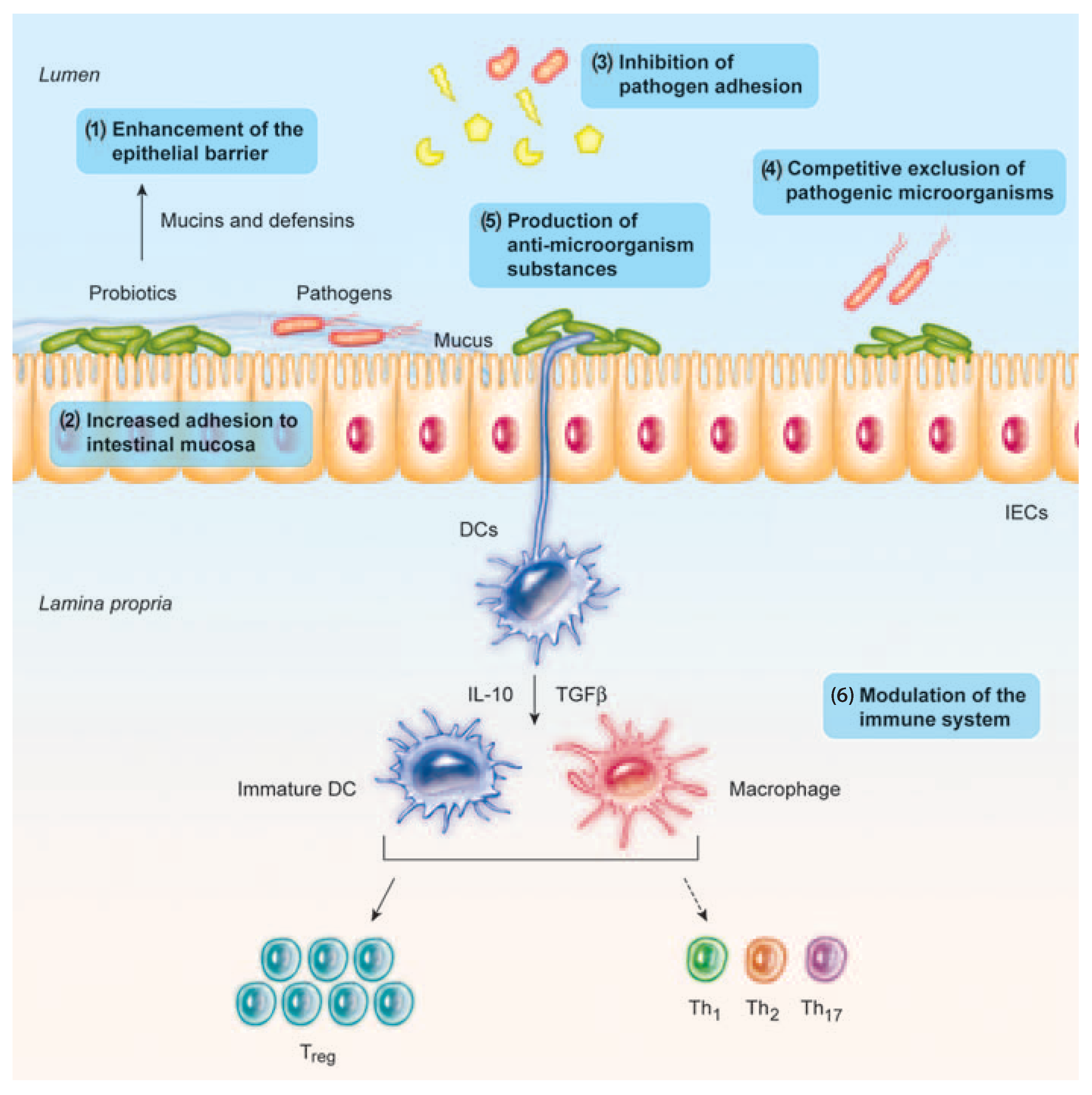{kind=link}
{kind=link}
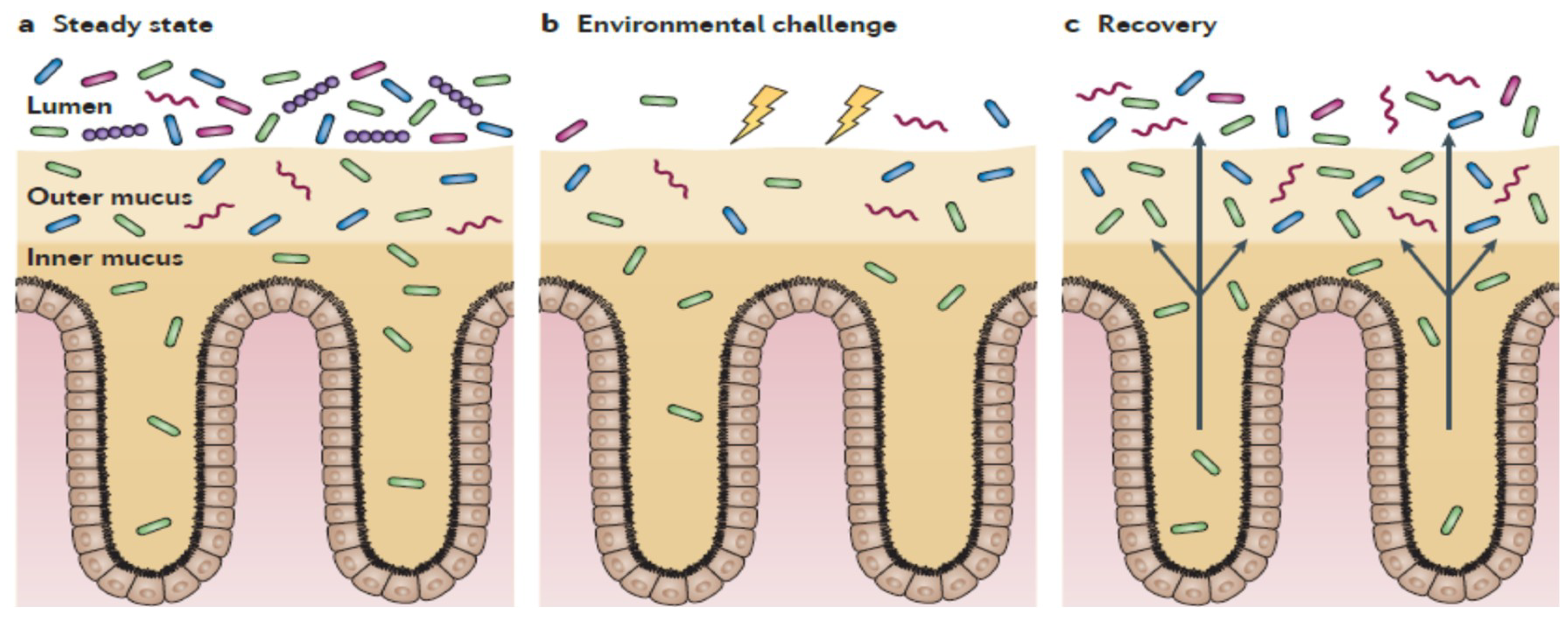{kind=link}
Table 1.
Advantages of inactivated bacteria and/or purified compounds in comparison with live probiotics.
Table 1.
Advantages of inactivated bacteria and/or purified compounds in comparison with live probiotics.
| Aspect | Advantages |
|---|---|
| Safety | No risk of translocation from gut lumen to blood, particularly in vulnerable subjects. No risk of acquisition and retransfer of antibiotic resistance genes. No risk of interference with normal colonization of gut microbiota in neonates. |
| Physiological effects | Release of active molecules from the disrupted inactivated cells, passing through the mucus layers and stimulating epithelial cells more directly. Loss of viability and cell lysis can produce further and more complex beneficial effects. |
| Pharmaceutical characteristics | Easier to standardize, transport, and store. |
Table 2.
Immunomodulating effects of heat-killed bacteria and cell-free supernatants.
| Immunomodulating Properties | |||
|---|---|---|---|
| Component/Fraction | Species | Effects | References |
| Heat-killed bacteria | L. paracasei, L. reuteri, L. casei, L. plantarum | Induction of IL-12 | [38] |
| Combination of L. acidophilus, L. plantarum, L. fermentum, and E. faecium | Enhanced immunomodulatory activity in comparison with live strains. Treatment at 100 °C for 30 min did not alter their adhesive capacity | [102] | |
| S. thermophilus | Production of IgA | [77] | |
| L. rhamnosus OLL2838 | Barrier protective properties in mice with induced colitis | [103] | |
| L. acidophilus LB + culture medium | Reduced paracellular permeability | [104] | |
| Product containing B. breve, B. longum, B. infantis, L. acidophilus, L. plantarum, L. paracasei, L. bulgaricus, and S. thermophilus | Protection of cyto-architecture of intestinal barrier; down-regulation of TNF-α expression | [105] | |
| B. breve M-16-V | Suppression of pro-inflammatory cytokine production | [106] | |
| B. bifidum OLB6378 | Increased expression of sIgA receptor | [107] | |
| Cell-free supernatants | L. acidophilus, L. casei, and L. reuteri | Down-regulation of PGE-2 and IL-8 expression | [68] |
| L. delbrueckii, L. paracasei, L. salivarius, L. reuteri, L. rhamnosus, L. acidophilus, L. plantarum, L. lactis, L. casei, S. thermophilus, B. breve, and B. longum | Anti-inflammatory responses mediated by metabolites and cell surfaces. Stimulation of cell-surface structures of PBMC similar to olive strains | [33] | |
| Soluble factors of L. reuteri CRL1098 | Anti-inflammatory responses | [68,108] | |
| Soluble peptides of L. rhamnosus GG | Prevention of cytokine-induced cell apoptosis | [19,109] | |
| Metabolites from B. breve | Immunomodulation in human dendritic cells | [68,110] | |
Table 3.
Protection against pathogens of heat-killed probiotic bacteria and cell-free supernatants.
| Protective Effects against Pathogens | |||
|---|---|---|---|
| Component/Fraction | Species | Effects | References |
| Heat-killed bacteria | Lactobacillus | Competition for adhesion sites (enterotoxigenic E. coli -ETEC-, Campylobacter, H. pylori | [59,89,111,112] |
| Combination of L. acidophilus, L. plantarum, L. fermentum, and E. faecium | Reduction of Salmonella invasion and the induced inflammation | [102] | |
| L. plantarum | Protection against Salmonella infection and reduction of translocation | [113] | |
| L. johnsonii | Inhibition of H. pylori growth | [89] | |
| Bifidobacteria | Resistance to Salmonella infection | [114] | |
| Bifidobacterium BB12 | Interference with S. mutans biofilm formation | [115] | |
| Cell-free supernatants | Lactic acid bacteria | Release of bacteriocins, inhibition of Gram-positive and Gram-negative bacteria | [39,116,117] |
| Bifidobacteria | Release of bacteriocins, against Gram-positive and Gram-negative bacteria and yeasts | [28,118,119] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102534
AMA Style
Piqué N, Berlanga M, Miñana-Galbis D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. International Journal of Molecular Sciences. 2019; 20(10):2534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102534
Chicago/Turabian StylePiqué, Núria, Mercedes Berlanga, and David Miñana-Galbis. 2019. "Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview" International Journal of Molecular Sciences 20, no. 10: 2534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102534
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.