Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk

 ,
,
Abstract
:1. Introduction
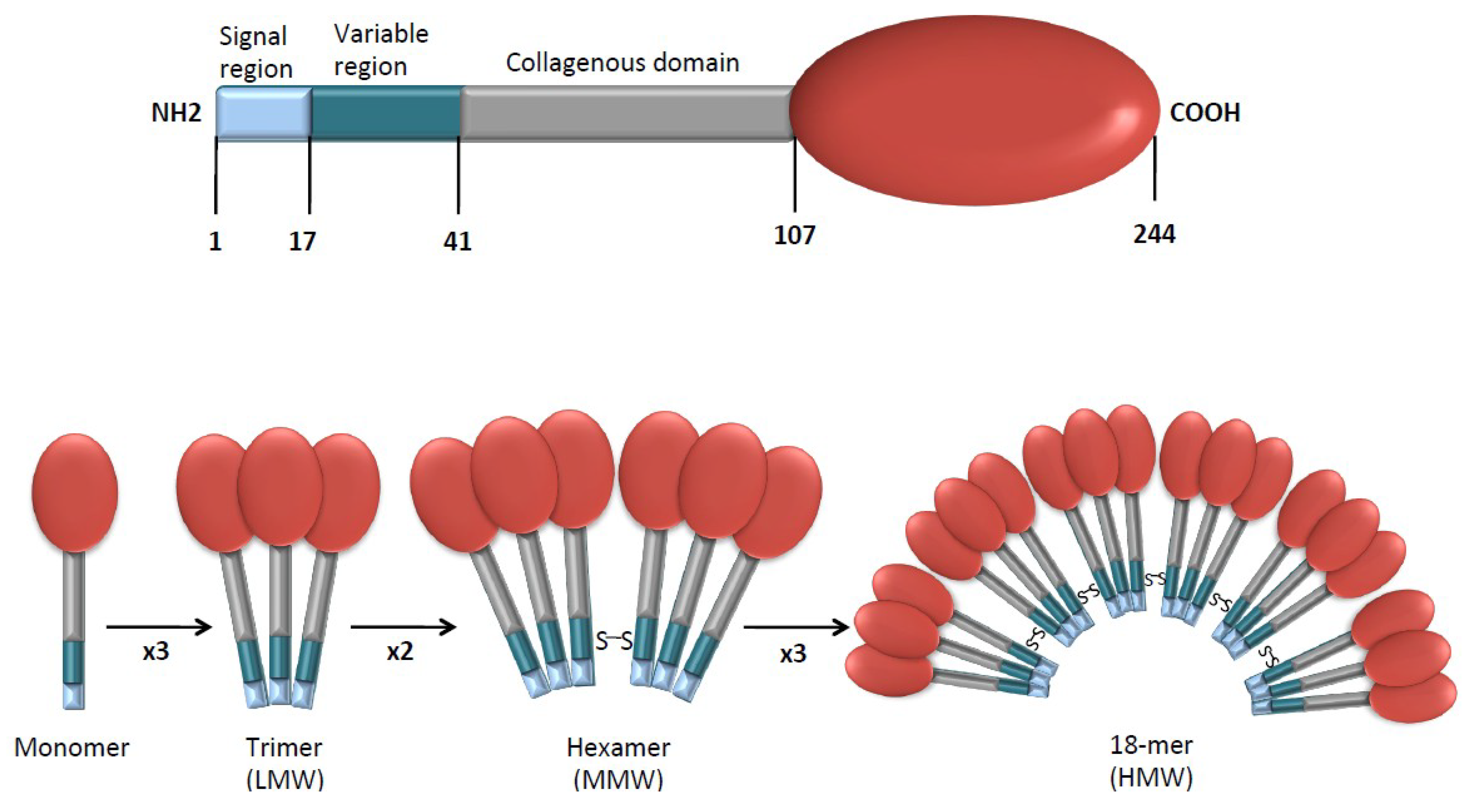
2. APN Structure and Receptors
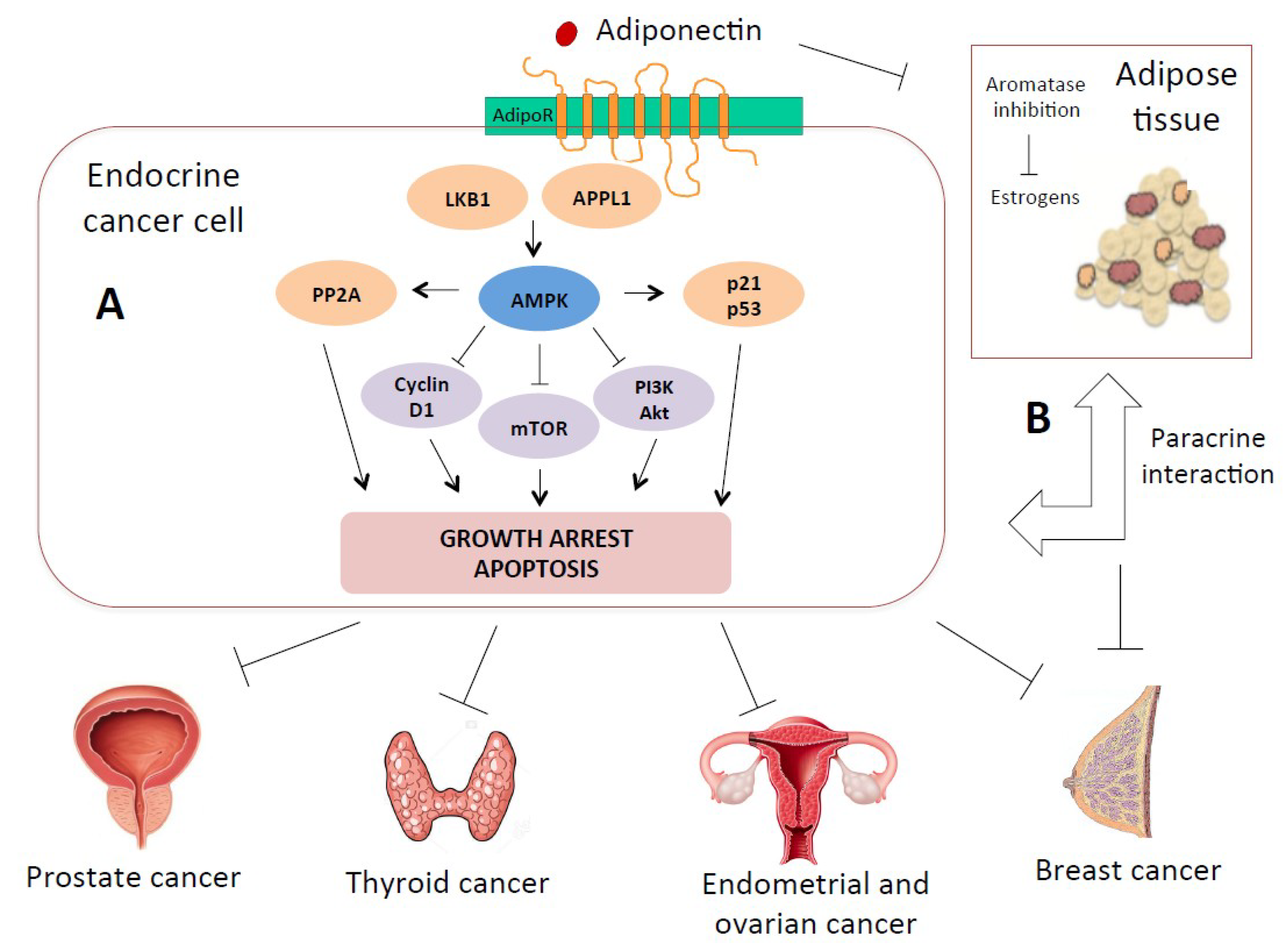
3. Adiponectin Signaling and Mechanisms of Carcinogenesis
3.1. Direct Mechanisms: Receptor-Mediated and Paracrine
3.2. Indirect Mechanisms: Insulin-Sensitizing, Immune-Related, Anti-Angiogenic Effects
4. Breast Cancer
5. Endometrial Cancer
6. Ovarian Cancer
7. Thyroid Cancer
8. Prostate Cancer
9. Adiponectin Role in Endocrine Cancer Metastasis
10. Future Perspectives and Therapeutic Implications
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APN | Adiponectin |
| AdipoR1/2 | Adiponectin receptor 1/2 |
| BMI | Body mass index |
| TNF | Tumor necrosis factor |
| LMW | Low molecular weight |
| MMW | Middle molecular weight |
| HMW | High molecular weight |
| AMPK | Adenosine monophosphate-activated protein kinase |
| LKB1 | Liver kinase B1 |
| ATP | Adenosine triphosphate |
| mTOR | Mammalian target of rapamycin |
| JAK | Janus kinase |
| IL-6 | Interleukin-6 |
| STAT3 | Signal transducer and activator of transcription 3 |
| ERK1/2 | Extracellular signal-regulated protein kinases 1 and 2 |
| PP2A | Protein phosphatase 2A |
| ERα | Estrogen receptor alpha |
| VEGF | Vascular endothelial growth factor |
| CRP | C-reactive protein |
| MAPK | Mitogen-activated protein kinase |
| IGF-I | Insulin-like growth factor I |
| SHBG | Sex hormones binding globulin |
| PPARγ | Proliferator-activated receptor-gamma |
| S6K | p70S6 kinase |
References
- NCD Risk Factor Collaboration (NCD-RisC). Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef]
- Global Burden of Metabolic Risk Factors for Chronic Diseases Collaboration (BMI Mediated Effects); Lu, Y.; Hajifathalian, K.; Ezzati, M.; Woodward, M.; Rimm, E.B.; Danaei, G. Metabolic mediators of the effects of body-mass index, overweight, and obesity on coronary heart disease and stroke: A pooled analysis of 97 prospective cohorts with 1.8 million participants. Lancet 2014, 383, 970–983. [Google Scholar] [CrossRef]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-mass index and incidence of cancer: A systematic review and meta-analysis of prospective observational studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Khandekar, M.J.; Cohen, P.; Spiegelman, B.M. Molecular mechanisms of cancer development in obesity. Nat. Rev. Cancer 2011, 11, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Global BMI Mortality Collaboration; Di Angelantonio, E.; Bhupathiraju Sh, N.; Wormser, D.; Gao, P.; Kaptoge, S.; Berrington de Gonzalez, A.; Cairns, B.J.; Huxley, R.; Jackson Ch, L.; et al. Body-mass index and all-cause mortality: Individual-participant-data meta-analysis of 239 prospective studies in four continents. Lancet 2016, 388, 776–786. [Google Scholar] [CrossRef]
- Sciacca, L.; Vigneri, R.; Tumminia, A.; Frasca, F.; Squatrito, S.; Frittitta, L.; Vigneri, P. Clinical and molecular mechanisms favoring cancer initiation and progression in diabetic patients. Nutr. Metab. Cardiov. Dis. 2013, 23, 808–815. [Google Scholar] [CrossRef]
- Vigneri, R.; Goldfine, I.D.; Frittitta, L. Insulin, insulin receptors, and cancer. J. Endocrinol. Invest. 2016, 39, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Vigneri, P.; Frasca, F.; Sciacca, L.; Frittitta, L.; Vigneri, R. Obesity and cancer. Nutr. Metab. Cardiov. Dis. 2006, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Magnuson, A.; Fouts, J.; Foster, M. Adipose tissue, obesity and adipokines: Role in cancer promotion. Hormone Mol. Biol. Clin. Invest. 2015, 21, 57–74. [Google Scholar] [CrossRef]
- Scherer, P.E. The Multifaceted Roles of Adipose Tissue-Therapeutic Targets for Diabetes and Beyond: The 2015 Banting Lecture. Diabetes 2016, 65, 1452–1461. [Google Scholar] [CrossRef] [PubMed]
- Di Zazzo, E.; Polito, R.; Bartollino, S.; Nigro, E.; Porcile, C.; Bianco, A.; Daniele, A.; Moncharmont, B. Adiponectin as Link Factor between Adipose Tissue and Cancer. Int. J. Mol. Sci. 2019, 20, 839. [Google Scholar] [CrossRef] [PubMed]
- Baratta, R.; Amato, S.; Degano, C.; Farina, M.G.; Patane, G.; Vigneri, R.; Frittitta, L. Adiponectin relationship with lipid metabolism is independent of body fat mass: Evidence from both cross-sectional and intervention studies. J. Clin. Endocrinol. Metab. 2004, 89, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Patane, G.; Caporarello, N.; Marchetti, P.; Parrino, C.; Sudano, D.; Marselli, L.; Vigneri, R.; Frittitta, L. Adiponectin increases glucose-induced insulin secretion through the activation of lipid oxidation. Acta Diabetol. 2013, 50, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Vansaun, M.N. Molecular pathways: Adiponectin and leptin signaling in cancer. Clin. Cancer Res. 2013, 19, 1926–1932. [Google Scholar] [CrossRef] [PubMed]
- Katira, A.; Tan, P.H. Evolving role of adiponectin in cancer-controversies and update. Cancer Biol. Med. 2016, 13, 101–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzaghi, C.; Trischitta, V. The Adiponectin Paradox for All-Cause and Cardiovascular Mortality. Diabetes 2018, 67, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.W.; Wang, J.; Hug, C.; Tsao, T.S.; Lodish, H.F. A family of Acrp30/adiponectin structural and functional paralogs. Proc. Natl. Acad. Sci. USA 2004, 101, 10302–10307. [Google Scholar] [CrossRef] [Green Version]
- Takemura, Y.; Ouchi, N.; Shibata, R.; Aprahamian, T.; Kirber, M.T.; Summer, R.S.; Kihara, S.; Walsh, K. Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies. J. Clin. Invest. 2007, 117, 375–386. [Google Scholar] [CrossRef]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.S.; Lodish, H.F. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebbard, L.; Ranscht, B. Multifaceted roles of adiponectin in cancer. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Diakopoulos, K.N.; Mantzoros, C.S. The role of adiponectin in cancer: A review of current evidence. Endocr. Rev. 2012, 33, 547–594. [Google Scholar] [CrossRef] [PubMed]
- Hefetz-Sela, S.; Scherer, P.E. Adipocytes: Impact on tumor growth and potential sites for therapeutic intervention. Pharmacol. Ther. 2013, 138, 197–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Housa, D.; Vernerova, Z.; Heracek, J.; Prochazka, B.; Cechak, P.; Kuncova, J.; Haluzik, M. Adiponectin as a potential marker of prostate cancer progression: Studies in organ-confined and locally advanced prostate cancer. Physiol. Res. 2008, 57, 451–458. [Google Scholar] [PubMed]
- Izadi, V.; Farabad, E.; Azadbakht, L. Serum adiponectin level and different kinds of cancer: A review of recent evidence. ISRN Oncol. 2012, 2012, 982769. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Kemp, B.E. AMPK in Health and Disease. Physiol. Rev. 2009, 89, 1025–1078. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, D.B.; Shaw, R.J. The LKB1-AMPK pathway: Metabolism and growth control in tumour suppression. Nat. Rev. Cancer 2009, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Taliaferro-Smith, L.; Nagalingam, A.; Zhong, D.; Zhou, W.; Saxena, N.K.; Sharma, D. LKB1 is required for adiponectin-mediated modulation of AMPK-S6K axis and inhibition of migration and invasion of breast cancer cells. Oncogene 2009, 28, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Manieri, E.; Herrera-Melle, L.; Mora, A.; Tomas-Loba, A.; Leiva-Vega, L.; Fernandez, D.I.; Rodriguez, E.; Moran, L.; Hernandez-Cosido, L.; Torres, J.L.; et al. Adiponectin accounts for gender differences in hepatocellular carcinoma incidence. J. Exp. Med. 2019, 216, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Saha, A.K.; Xiang, X.; Ruderman, N.B. AMPK, the metabolic syndrome and cancer. Trends Pharmacol. Sci. 2005, 26, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cong, L.; Gasser, J.; Zhao, J.; Chen, K.; Li, F. Mechanisms underlying the anti-proliferative actions of adiponectin in human breast cancer cells, MCF7-dependency on the cAMP/protein kinase-A pathway. Nutr. Cancer 2011, 63, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Korner, A.; Pazaitou-Panayiotou, K.; Kelesidis, T.; Kelesidis, I.; Williams, C.J.; Kaprara, A.; Bullen, J.; Neuwirth, A.; Tseleni, S.; Mitsiades, N.; et al. Total and high-molecular-weight adiponectin in breast cancer: In vitro and in vivo studies. J. Clin. Endocrinol. Metab. 2007, 92, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Zakikhani, M.; Dowling, R.J.; Sonenberg, N.; Pollak, M.N. The effects of adiponectin and metformin on prostate and colon neoplasia involve activation of AMP-activated protein kinase. Cancer Prev. Res. 2008, 1, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Barb, D.; Neuwirth, A.; Mantzoros, C.S.; Balk, S.P. Adiponectin signals in prostate cancer cells through Akt to activate the mammalian target of rapamycin pathway. Endocr. Relat. Cancer 2007, 14, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Gasser, J.; Zhao, J.; Yang, B.; Li, F.; Zhao, A.Z. Human adiponectin inhibits cell growth and induces apoptosis in human endometrial carcinoma cells, HEC-1-A and RL95 2. Endocr. Relat. Cancer 2007, 14, 713–720. [Google Scholar] [CrossRef]
- Sablina, A.A.; Hahn, W.C. The role of PP2A A subunits in tumor suppression. Cell Adhes. Migr. 2007, 1, 140–141. [Google Scholar] [CrossRef]
- Kim, K.Y.; Baek, A.; Hwang, J.E.; Choi, Y.A.; Jeong, J.; Lee, M.S.; Cho, D.H.; Lim, J.S.; Kim, K.I.; Yang, Y. Adiponectin-activated AMPK stimulates dephosphorylation of AKT through protein phosphatase 2A activation. Cancer Res. 2009, 69, 4018–4026. [Google Scholar] [CrossRef]
- Bowman, T.; Garcia, R.; Turkson, J.; Jove, R. STATs in oncogenesis. Oncogene 2000, 19, 2474–2488. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Euhus, D.M.; Scherer, P.E. Paracrine and endocrine effects of adipose tissue on cancer development and progression. Endocr. Rev. 2011, 32, 550–570. [Google Scholar] [CrossRef]
- Brown, K.A.; Simpson, E.R. Obesity and breast cancer: Progress to understanding the relationship. Cancer Res. 2010, 70, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Stephens, J.M.; Elks, C.M. Oncostatin M: Potential Implications for Malignancy and Metabolism. Curr. Pharm. Des. 2017, 23, 3645–3657. [Google Scholar] [CrossRef] [PubMed]
- Pollak, M. Insulin and insulin-like growth factor signalling in neoplasia. Nat. Rev. Cancer 2008, 8, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Sciacca, L.; Vella, V.; Frittitta, L.; Tumminia, A.; Manzella, L.; Squatrito, S.; Belfiore, A.; Vigneri, R. Long-acting insulin analogs and cancer. Nutr. Metab. Cardiov. Dis. 2018, 28, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Weyer, C.; Funahashi, T.; Tanaka, S.; Hotta, K.; Matsuzawa, Y.; Pratley, R.E.; Tataranni, P.A. Hypoadiponectinemia in obesity and type 2 diabetes: Close association with insulin resistance and hyperinsulinemia. J. Clin. Endocrinol. Metab. 2001, 86, 1930–1935. [Google Scholar] [CrossRef]
- Kelesidis, I.; Kelesidis, T.; Mantzoros, C.S. Adiponectin and cancer: A systematic review. Br. J. Cancer 2006, 94, 1221–1225. [Google Scholar] [CrossRef]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Mol. Cell 2010, 40, 310–322. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Stacker, S.A.; Williams, S.P.; Karnezis, T.; Shayan, R.; Fox, S.B.; Achen, M.G. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat. Rev. Cancer 2014, 14, 159–172. [Google Scholar] [CrossRef]
- Catalan, V.; Gomez-Ambrosi, J.; Rodriguez, A.; Fruhbeck, G. Adipose tissue immunity and cancer. Front. Physiol. 2013, 4, 275. [Google Scholar] [CrossRef] [Green Version]
- Mraz, M.; Haluzik, M. The role of adipose tissue immune cells in obesity and low-grade inflammation. J. Endocrinol. 2014, 222, R113–R127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoldi, A.; Naffah de Souza, C.; Camara, N.O.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2015, 6, 637. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New insight into adiponectin role in obesity and obesity-related diseases. BioMed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Yoshida, H. Beneficial Effects of Adiponectin on Glucose and Lipid Metabolism and Atherosclerotic Progression: Mechanisms and Perspectives. Int. J. Mol. Sci. 2019, 20, 1190. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Oritani, K.; Takahashi, I.; Ishikawa, J.; Matsuyama, A.; Ouchi, N.; Kihara, S.; Funahashi, T.; Tenner, A.J.; Tomiyama, Y.; et al. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000, 96, 1723–1732. [Google Scholar] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Brakenhielm, E.; Veitonmaki, N.; Cao, R.; Kihara, S.; Matsuzawa, Y.; Zhivotovsky, B.; Funahashi, T.; Cao, Y. Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 2476–2481. [Google Scholar] [CrossRef] [Green Version]
- Bergers, G.; Hanahan, D. Modes of resistance to anti-angiogenic therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef] [Green Version]
- Mahadev, K.; Wu, X.; Donnelly, S.; Ouedraogo, R.; Eckhart, A.D.; Goldstein, B.J. Adiponectin inhibits vascular endothelial growth factor-induced migration of human coronary artery endothelial cells. Cardiov. Res. 2008, 78, 376–384. [Google Scholar] [CrossRef]
- Hebbard, L.W.; Garlatti, M.; Young, L.J.; Cardiff, R.D.; Oshima, R.G.; Ranscht, B. T-cadherin supports angiogenesis and adiponectin association with the vasculature in a mouse mammary tumor model. Cancer Res. 2008, 68, 1407–1416. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Picon-Ruiz, M.; Morata-Tarifa, C.; Valle-Goffin, J.J.; Friedman, E.R.; Slingerland, J.M. Obesity and adverse breast cancer risk and outcome: Mechanistic insights and strategies for intervention. Cancer J. Clin. 2017, 67, 378–397. [Google Scholar] [CrossRef] [PubMed]
- Panis, C.; Herrera, A.; Aranome, A.M.F.; Victorino, V.J.; Michelleti, P.L.; Morimoto, H.K.; Cecchini, A.L.; Simao, A.N.C.; Cecchini, R. Clinical insights from adiponectin analysis in breast cancer patients reveal its anti-inflammatory properties in non-obese women. Mol. Cell. Endocrinol. 2014, 382, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Carroll, P.A.; Healy, L.; Lysaght, J.; Boyle, T.; Reynolds, J.V.; Kennedy, M.J.; Pidgeon, G.; Connolly, E.M. Influence of the metabolic syndrome on leptin and leptin receptor in breast cancer. Mol. Carcinog. 2011, 50, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.C.; Hosick, H.L.; Watkins, B.A. Growth of epithelium from a preneoplastic mammary outgrowth in response to mammary adipose tissue. In Vitro Cell. Dev. Biol. 1989, 25, 409–418. [Google Scholar] [CrossRef]
- Iyengar, P.; Combs, T.P.; Shah, S.J.; Gouon-Evans, V.; Pollard, J.W.; Albanese, C.; Flanagan, L.; Tenniswood, M.P.; Guha, C.; Lisanti, M.P.; et al. Adipocyte-secreted factors synergistically promote mammary tumorigenesis through induction of anti-apoptotic transcriptional programs and proto-oncogene stabilization. Oncogene 2003, 22, 6408–6423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, E.; Benaitreau, D.; Dieudonne, M.N.; Leneveu, M.C.; Serazin, V.; Giudicelli, Y.; Pecquery, R. Adiponectin mediates an antiproliferative response in human MDA-MB 231 breast cancer cells. Oncol. Rep. 2008, 20, 971–977. [Google Scholar] [PubMed]
- Arditi, J.D.; Venihaki, M.; Karalis, K.P.; Chrousos, G.P. Antiproliferative effect of adiponectin on MCF7 breast cancer cells: A potential hormonal link between obesity and cancer. Hormone Metab. Res. 2007, 39, 9–13. [Google Scholar] [CrossRef]
- Dieudonne, M.N.; Bussiere, M.; Dos Santos, E.; Leneveu, M.C.; Giudicelli, Y.; Pecquery, R. Adiponectin mediates antiproliferative and apoptotic responses in human MCF7 breast cancer cells. Biochem. Biophys. Res. Commun. 2006, 345, 271–279. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Funahashi, T.; Kihara, S.; Taguchi, T.; Tamaki, Y.; Matsuzawa, Y.; Noguchi, S. Association of serum adiponectin levels with breast cancer risk. Clin. Cancer Res. 2003, 9, 5699–5704. [Google Scholar]
- Macis, D.; Guerrieri-Gonzaga, A.; Gandini, S. Circulating adiponectin and breast cancer risk: A systematic review and meta-analysis. In. J. Epidemiol. 2014, 43, 1226–1236. [Google Scholar] [CrossRef]
- Tian, Y.F.; Chu, C.H.; Wu, M.H.; Chang, C.L.; Yang, T.; Chou, Y.C.; Hsu, G.C.; Yu, C.P.; Yu, J.C.; Sun, C.A. Anthropometric measures, plasma adiponectin, and breast cancer risk. Endocr. Cancer 2007, 14, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Jia, J.; Dong, S.; Zhang, C.; Yu, S.; Li, L.; Mao, C.; Wang, D.; Chen, J.; Yuan, G. Circulating adiponectin levels and the risk of breast cancer: A meta-analysis. Eur. J. Cancer Prev. 2014, 23, 158–165. [Google Scholar] [CrossRef]
- Mantzoros, C.; Petridou, E.; Dessypris, N.; Chavelas, C.; Dalamaga, M.; Alexe, D.M.; Papadiamantis, Y.; Markopoulos, C.; Spanos, E.; Chrousos, G.; et al. Adiponectin and breast cancer risk. J. Clin. Endocrinol. Metab. 2004, 89, 1102–1107. [Google Scholar] [CrossRef]
- Tworoger, S.S.; Eliassen, A.H.; Kelesidis, T.; Colditz, G.A.; Willett, W.C.; Mantzoros, C.S.; Hankinson, S.E. Plasma adiponectin concentrations and risk of incident breast cancer. J. Clin. Endocrinol. Metab. 2007, 92, 1510–1516. [Google Scholar] [CrossRef]
- Cust, A.E.; Stocks, T.; Lukanova, A.; Lundin, E.; Hallmans, G.; Kaaks, R.; Jonsson, H.; Stattin, P. The influence of overweight and insulin resistance on breast cancer risk and tumour stage at diagnosis: A prospective study. Breast Cancer Res. Treat. 2009, 113, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Hancke, K.; Grubeck, D.; Hauser, N.; Kreienberg, R.; Weiss, J.M. Adipocyte fatty acid-binding protein as a novel prognostic factor in obese breast cancer patients. Breast Cancer Res. Treat. 2010, 119, 367. [Google Scholar] [CrossRef] [PubMed]
- Macis, D.; Gandini, S.; Guerrieri-Gonzaga, A.; Johansson, H.; Magni, P.; Ruscica, M.; Lazzeroni, M.; Serrano, D.; Cazzaniga, M.; Mora, S.; et al. Prognostic effect of circulating adiponectin in a randomized 2 x 2 trial of low-dose tamoxifen and fenretinide in premenopausal women at risk for breast cancer. J. Clin. Oncol. 2012, 30, 151–157. [Google Scholar] [CrossRef]
- Gu, L.; Cao, C.; Fu, J.; Li, Q.; Li, D.H.; Chen, M.Y. Serum adiponectin in breast cancer: A meta-analysis. Medicine 2018, 97, e11433. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Bong, J.G.; Park, S.H.; Choi, J.H.; Oh, H.K. Expression of leptin, leptin receptor, adiponectin, and adiponectin receptor in ductal carcinoma in situ and invasive breast cancer. J. Breast Cancer 2011, 14, 96–103. [Google Scholar] [CrossRef]
- Yu, Z.; Tang, S.; Ma, H.; Duan, H.; Zeng, Y. Association of serum adiponectin with breast cancer: A meta-analysis of 27 case-control studies. Medicine 2019, 98, e14359. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Wang, M.; Ma, Z.B.; Yu, L.X.; Zhang, Q.; Gao, D.Z.; Wang, F.; Yu, Z.G. The role of adiponectin in breast cancer: A meta-analysis. PLoS ONE 2013, 8, e73183. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.M.; de Almeida, J.C.; Feoli, A.M. Effect of diet on adiponectin levels in blood. Nutr. Rev. 2011, 69, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.; Coleman, M.; Riis, S.; Favre, C.; O’Flanagan, C.H.; Zhdanov, A.V.; Papkovsky, D.B.; Hursting, S.D.; O’Connor, R. Insulin-like growth factor 1 signaling is essential for mitochondrial biogenesis and mitophagy in cancer cells. J. Biol. Chem. 2017, 292, 16983–16998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, A.; Sarcevic, B.; Prall, O.W.; Sutherland, R.L. Insulin/insulin-like growth factor-I and estrogen cooperate to stimulate cyclin E-Cdk2 activation and cell Cycle progression in MCF-7 breast cancer cells through differential regulation of cyclin E and p21(WAF1/Cip1). J. Biol. Chem. 2001, 276, 25823–25833. [Google Scholar] [CrossRef] [PubMed]
- Gavrila, A.; Chan, J.L.; Yiannakouris, N.; Kontogianni, M.; Miller, L.C.; Orlova, C.; Mantzoros, C.S. Serum adiponectin levels are inversely associated with overall and central fat distribution but are not directly regulated by acute fasting or leptin administration in humans: Cross-sectional and interventional studies. J. Clin. Endocrinol. Metab. 2003, 88, 4823–4831. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, G.P.; Provatopoulou, X.; Kalogera, E.; Siasos, G.; Menenakos, E.; Zografos, G.C.; Gounaris, A. Serum resistin is inversely related to breast cancer risk in premenopausal women. Breast 2016, 29, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.M.; Duan, X.N.; Cui, S.D.; Tian, F.G.; Cao, X.C.; Geng, C.Z.; Fan, Z.M.; Wang, X.; Wang, S.; Jiang, H.C.; et al. Circulating High-Molecular-Weight (HMW) Adiponectin Level Is Related with Breast Cancer Risk Better than Total Adiponectin: A Case-Control Study. PLoS ONE 2015, 10, e0129246. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lam, J.B.; Lam, K.S.; Liu, J.; Lam, M.C.; Hoo, R.L.; Wu, D.; Cooper, G.J.; Xu, A. Adiponectin modulates the glycogen synthase kinase-3beta/beta-catenin signaling pathway and attenuates mammary tumorigenesis of MDA-MB-231 cells in nude mice. Cancer Res. 2006, 66, 11462–11470. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, M.E.; Nkhata, K.J.; Mizuno, N.K.; Ray, A.; Cleary, M.P. Effects of adiponectin on breast cancer cell growth and signaling. Br. J. Cancer 2008, 98, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, S.; Miyoshi, Y.; Ishihara, H.; Noguchi, S. Growth-inhibitory effect of adiponectin via adiponectin receptor 1 on human breast cancer cells through inhibition of S-phase entry without inducing apoptosis. Breast Cancer Res. Treat. 2008, 112, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Mauro, L.; Pellegrino, M.; De Amicis, F.; Ricchio, E.; Giordano, F.; Rizza, P.; Catalano, S.; Bonofiglio, D.; Sisci, D.; Panno, M.L.; et al. Evidences that estrogen receptor alpha interferes with adiponectin effects on breast cancer cell growth. Cell Cycle 2014, 13, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Panno, M.L.; Naimo, G.D.; Spina, E.; Ando, S.; Mauro, L. Different molecular signaling sustaining adiponectin action in breast cancer. Curr. Opin. Pharmacol. 2016, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mauro, L.; Naimo, G.D.; Gelsomino, L.; Malivindi, R.; Bruno, L.; Pellegrino, M.; Tarallo, R.; Memoli, D.; Weisz, A.; Panno, M.L.; et al. Uncoupling effects of estrogen receptor alpha on LKB1/AMPK interaction upon adiponectin exposure in breast cancer. FASEB J. 2018, 32, 4343–4355. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lee, Y.Y.; Yu, B.Y.; Yang, B.S.; Cho, K.H.; Yoon, D.K.; Roh, Y.K. Adiponectin induces growth arrest and apoptosis of MDA-MB-231 breast cancer cell. Archives Pharm. Res. 2005, 28, 1263–1269. [Google Scholar] [CrossRef]
- Li, Z.J.; Yang, X.L.; Yao, Y.; Han, W.Q.; Li, B.O. Circulating adiponectin levels and risk of endometrial cancer: Systematic review and meta-analysis. Exp. Ther. Med. 2016, 11, 2305–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Zhao, X.; Kong, W.M. Association between adiponectin levels and endometrial carcinoma risk: Evidence from a dose-response meta-analysis. BMJ open 2015, 5, e008541. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Shi, J.; Long, Y.; Tian, H.; Li, X.; Zhao, A.Z.; Li, R.F.; Chen, T. Adiponectin and Endometrial Cancer: A Systematic Review and Meta-Analysis. Cell. Physiol. Biochem. 2015, 36, 1670–1678. [Google Scholar] [CrossRef]
- Zheng, Q.; Wu, H.; Cao, J. Circulating adiponectin and risk of endometrial cancer. PLoS ONE 2015, 10, e0129824. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.T.; Wu, Q.J.; Wang, Y.L.; Ma, X.X. Circulating adiponectin, leptin and adiponectin-leptin ratio and endometrial cancer risk: Evidence from a meta-analysis of epidemiologic studies. Int. J. Cancer 2015, 137, 1967–1978. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.H.; Kim, H.J.; Kim, C.Y.; Kim, Y.H.; Ju, W.; Kim, S.C. Association of plasma adiponectin and leptin levels with the development and progression of ovarian cancer. Obstetrics Gynecol. Sci. 2016, 59, 279–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossus, L.; Franceschi, S.; Biessy, C.; Navionis, A.S.; Travis, R.C.; Weiderpass, E.; Scalbert, A.; Romieu, I.; Tjonneland, A.; Olsen, A.; et al. Adipokines and inflammation markers and risk of differentiated thyroid carcinoma: The EPIC study. Int. J. Cancer 2018, 142, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Warakomski, J.; Romuk, E.; Jarzab, B.; Krajewska, J.; Sieminska, L. Concentrations of Selected Adipokines, Interleukin-6, and Vitamin D in Patients with Papillary Thyroid Carcinoma in Respect to Thyroid Cancer Stages. Int. J. Endocrinol. 2018, 2018, 4921803. [Google Scholar] [CrossRef] [PubMed]
- Abooshahab, R.; Yaghmaei, P.; Ghadaksaz, H.G.; Hedayati, M. Lack of Association between Serum Adiponectin/Leptin Levels and Medullary Thyroid Cancer. Asian Pac. J. Cancer Prev. 2016, 17, 3861–3864. [Google Scholar]
- Mitsiades, N.; Pazaitou-Panayiotou, K.; Aronis, K.N.; Moon, H.S.; Chamberland, J.P.; Liu, X.; Diakopoulos, K.N.; Kyttaris, V.; Panagiotou, V.; Mylvaganam, G.; et al. Circulating adiponectin is inversely associated with risk of thyroid cancer: In vivo and in vitro studies. J. Clin. Endocrinol. Metab. 2011, 96, E2023–E2028. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Long, C.; Deng, Z.; Bi, X.; Hu, J. The role of circulating adiponectin in prostate cancer: A meta-analysis. Int. J. Biol. Markers 2015, 30. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, A.; Nakagawa, T.; Kawai, K.; Onozawa, M.; Hayashi, T.; Matsushita, Y.; Tsutsumi, M.; Kojima, T.; Miyazaki, J.; Nishiyama, H. Serum adiponectin concentration in 2,939 Japanese men undergoing screening for prostate cancer. Prostate Inte. 2015, 3, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Stampfer, M.J.; Mucci, L.; Rifai, N.; Qiu, W.; Kurth, T.; Ma, J. A 25-year prospective study of plasma adiponectin and leptin concentrations and prostate cancer risk and survival. Clin. Chem. 2010, 56, 34–43. [Google Scholar] [CrossRef]
- Baillargeon, J.; Platz, E.A.; Rose, D.P.; Pollock, B.H.; Ankerst, D.P.; Haffner, S.; Higgins, B.; Lokshin, A.; Troyer, D.; Hernandez, J.; et al. Obesity, adipokines, and prostate cancer in a prospective population-based study. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Goktas, S.; Yilmaz, M.I.; Caglar, K.; Sonmez, A.; Kilic, S.; Bedir, S. Prostate cancer and adiponectin. Urology 2005, 65, 1168–1172. [Google Scholar] [CrossRef]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Orekoya, O.; Samson, M.E.; Trivedi, T.; Vyas, S.; Steck, S.E. The Impact of Obesity on Surgical Outcome in Endometrial Cancer Patients: A Systematic Review. J. Gynecol. Surg. 2016, 32, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Crosbie, E.J.; Zwahlen, M.; Kitchener, H.C.; Egger, M.; Renehan, A.G. Body mass index, hormone replacement therapy, and endometrial cancer risk: A meta-analysis. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 3119–3130. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, J.; Gamborg, M.; Ulrich, L.G.; Sorensen, T.I.; Baker, J.L. Childhood body mass index and height and risk of histologic subtypes of endometrial cancer. Int. J. Obesity 2016, 40, 1096–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kruijsdijk, R.C.; van der Wall, E.; Visseren, F.L. Obesity and cancer: The role of dysfunctional adipose tissue. Cancer Epidemiol. Biomarkers Prev. 2009, 18, 2569–2578. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, Z.; Zhang, Y.; Lu, B. Serum leptin, adiponectin and endometrial cancer risk in Chinese women. J. Gynecol. Oncol. 2013, 24, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wen, K.; Han, X.; Liu, R.; Qu, Q. Adiponectin mediates antiproliferative and apoptotic responses in endometrial carcinoma by the AdipoRs/AMPK pathway. Gynecol. Oncol. 2015, 137, 311–320. [Google Scholar] [CrossRef]
- Cust, A.E.; Kaaks, R.; Friedenreich, C.; Bonnet, F.; Laville, M.; Lukanova, A.; Rinaldi, S.; Dossus, L.; Slimani, N.; Lundin, E.; et al. Plasma adiponectin levels and endometrial cancer risk in pre- and postmenopausal women. J. Clin. Endocrinol. Metab. 2007, 92, 255–263. [Google Scholar] [CrossRef]
- Rzepka-Gorska, I.; Bedner, R.; Cymbaluk-Ploska, A.; Chudecka-Glaz, A. Serum adiponectin in relation to endometrial cancer and endometrial hyperplasia with atypia in obese women. Eur. J. Gynaecol. Oncol. 2008, 29, 594–597. [Google Scholar]
- Moon, H.S.; Chamberland, J.P.; Aronis, K.; Tseleni-Balafouta, S.; Mantzoros, C.S. Direct role of adiponectin and adiponectin receptors in endometrial cancer: In vitro and ex vivo studies in humans. Mol. Cancer Ther. 2011, 10, 2234–2243. [Google Scholar] [CrossRef]
- Yabushita, H.; Iwasaki, K.; Obayashi, Y.; Wakatsuki, A. Clinicopathological roles of adiponectin and leptin receptors in endometrial carcinoma. Oncol. Lett. 2014, 7, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, I.; Bast, R.C., Jr. Minireview: Human ovarian cancer: Biology, current management, and paths to personalizing therapy. Endocrinology 2012, 153, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Kalliala, I.; Markozannes, G.; Gunter, M.J.; Paraskevaidis, E.; Gabra, H.; Mitra, A.; Terzidou, V.; Bennett, P.; Martin-Hirsch, P.; Tsilidis, K.K.; et al. Obesity and gynaecological and obstetric conditions: Umbrella review of the literature. BMJ 2017, 359, j4511. [Google Scholar] [CrossRef] [PubMed]
- Diaz, E.S.; Karlan, B.Y.; Li, A.J. Obesity-associated adipokines correlate with survival in epithelial ovarian cancer. Gynecol. Oncol. 2013, 129, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Slomian, G.J.; Nowak, D.; Buczkowska, M.; Glogowska-Gruszka, A.; Slomian, S.P.; Roczniak, W.; Janyga, S.; Nowak, P. The role of adiponectin and leptin in the treatment of ovarian cancer patients. Endokr. Polska 2019, 70, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yu, Z.; Fang, L.; Liu, F.; Jiang, K. Expression of Adiponectin Receptor-1 and Prognosis of Epithelial Ovarian Cancer Patients. Med. Sci. Monit. 2017, 23, 1514–1521. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Gogola, J.; Ptak, A. Adiponectin Reverses the Proliferative Effects of Estradiol and IGF-1 in Human Epithelial Ovarian Cancer Cells by Downregulating the Expression of Their Receptors. Hormones Cancer 2018, 9, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid 2016, 26, 1–133. [Google Scholar] [CrossRef] [Green Version]
- Schmid, D.; Ricci, C.; Behrens, G.; Leitzmann, M.F. Adiposity and risk of thyroid cancer: A systematic review and meta-analysis. Obesity Rev. 2015, 16, 1042–1054. [Google Scholar] [CrossRef]
- Pappa, T.; Alevizaki, M. Obesity and thyroid cancer: A clinical update. Thyroid 2014, 24, 190–199. [Google Scholar] [CrossRef]
- Cheng, S.P.; Liu, C.L.; Hsu, Y.C.; Chang, Y.C.; Huang, S.Y.; Lee, J.J. Expression and biologic significance of adiponectin receptors in papillary thyroid carcinoma. Cell Biochem. Biophys. 2013, 65, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.B.; Liu, S.H.; Jiang, H.W.; Bai, P.D.; Ding, Q. Obesity affects the biopsy-mediated detection of prostate cancer, particularly high-grade prostate cancer: A dose-response meta-analysis of 29,464 patients. PLoS ONE 2014, 9, e106677. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.B.; Xu, H.; Bai, P.D.; Jiang, H.W.; Ding, Q. Obesity has multifaceted impact on biochemical recurrence of prostate cancer: A dose-response meta-analysis of 36,927 patients. Med. Oncol. 2014, 31, 829. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.B.; Xu, H.; Hu, J.M.; Zhu, W.H.; Yang, T.; Jiang, H.W.; Ding, Q. Genetic polymorphisms in leptin, adiponectin and their receptors affect risk and aggressiveness of prostate cancer: Evidence from a meta-analysis and pooled-review. Oncotarget 2016, 7, 81049–81061. [Google Scholar] [CrossRef] [PubMed]
- Michalakis, K.; Williams, C.J.; Mitsiades, N.; Blakeman, J.; Balafouta-Tselenis, S.; Giannopoulos, A.; Mantzoros, C.S. Serum adiponectin concentrations and tissue expression of adiponectin receptors are reduced in patients with prostate cancer: A case control study. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Wang, L.; Ma, Q.; Qi, M.; Lu, N.; Zhang, L.; Han, B. Adiponectin as a potential tumor suppressor inhibiting epithelial-to-mesenchymal transition but frequently silenced in prostate cancer by promoter methylation. Prostate 2015, 75, 1197–1205. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.P.; Hou, Z.F.; Duivenvoorden, W.C.; Whelan, K.; Honig, A.; Pinthus, J.H. Adiponectin inhibits oxidative stress in human prostate carcinoma cells. Prostate Cancer Prostatic Dis. 2012, 15, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Bub, J.D.; Miyazaki, T.; Iwamoto, Y. Adiponectin as a growth inhibitor in prostate cancer cells. Biochem. Biophys. Res. Commun. 2006, 340, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zheng, J.; Yao, X.; Peng, B. Adiponectin inhibits VEGF-A in prostate cancer cells. Tumour Biol. 2015, 36, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Yao, X.; Zheng, J. MiR-323 Inhibits Prostate Cancer Vascularization Through Adiponectin Receptor. Cellular Physiol. Biochem. 2015, 36, 1491–1498. [Google Scholar] [CrossRef]
- Rider, J.R.; Fiorentino, M.; Kelly, R.; Gerke, T.; Jordahl, K.; Sinnott, J.A.; Giovannucci, E.L.; Loda, M.; Mucci, L.A.; Finn, S.; et al. Tumor expression of adiponectin receptor 2 and lethal prostate cancer. Carcinogenesis 2015, 36, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.L.; Ma, J.; Chavarro, J.E.; Freedman, M.L.; Lis, R.; Fedele, G.; Fiore, C.; Qiu, W.; Fiorentino, M.; Finn, S.; et al. Fatty acid synthase polymorphisms, tumor expression, body mass index, prostate cancer risk, and survival. J.Clin. Oncol. 2010, 28, 3958–3964. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Qu, Y.; Zhang, G.; Sun, L.; Zhu, Y.; Ye, D. A single nucleotide polymorphism in ADIPOQ predicts biochemical recurrence after radical prostatectomy in localized prostate cancer. Oncotarget 2015, 6, 32205–32211. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.K.; Sharma, D. Metastasis suppression by adiponectin: LKB1 rises up to the challenge. Cell Adhes. Migr. 2010, 4, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yan, Q.; Zhang, Z.; Du, G.; Wan, X. Acrp30 inhibits leptin-induced metastasis by downregulating the JAK/STAT3 pathway via AMPK activation in aggressive SPEC-2 endometrial cancer cells. Oncol. Rep. 2012, 27, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Hadad, S.M.; Fleming, S.; Thompson, A.M. Targeting AMPK: A new therapeutic opportunity in breast cancer. Crit. Rev. Oncol. Hematol. 2008, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Otvos, L., Jr.; Haspinger, E.; La Russa, F.; Maspero, F.; Graziano, P.; Kovalszky, I.; Lovas, S.; Nama, K.; Hoffmann, R.; Knappe, D.; et al. Design and development of a peptide-based adiponectin receptor agonist for cancer treatment. BMC Biotech. 2011, 11, 90. [Google Scholar] [CrossRef]
- Otvos, L., Jr.; Kovalszky, I.; Olah, J.; Coroniti, R.; Knappe, D.; Nollmann, F.I.; Hoffmann, R.; Wade, J.D.; Lovas, S.; Surmacz, E. Optimization of adiponectin-derived peptides for inhibition of cancer cell growth and signaling. Biopolymers 2015, 104, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zang, Z.; Zhong, L.; Wu, M.; Su, Q.; Gao, X.; Zan, W.; Lin, D.; Zhao, Y.; Zhang, Z. Identification of adiponectin receptor agonist utilizing a fluorescence polarization based high throughput assay. PLoS ONE 2013, 8, e63354. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Takahashi, M.; Funahashi, T.; Kihara, S.; Nishizawa, H.; Kishida, K.; Nagaretani, H.; Matsuda, M.; Komuro, R.; Ouchi, N.; et al. PPARgamma ligands increase expression and plasma concentrations of adiponectin, an adipose-derived protein. Diabetes 2001, 50, 2094–2099. [Google Scholar] [CrossRef]
- Burstein, H.J.; Demetri, G.D.; Mueller, E.; Sarraf, P.; Spiegelman, B.M.; Winer, E.P. Use of the peroxisome proliferator-activated receptor (PPAR) gamma ligand troglitazone as treatment for refractory breast cancer: A phase II study. Breast Cancer Res. Treat. 2003, 79, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Williams, R. Discontinued in 2013: Oncology drugs. Exp. Opin. Invest. Drugs 2015, 24, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Ealey, K.N.; Kaludjerovic, J.; Archer, M.C.; Ward, W.E. Adiponectin is a negative regulator of bone mineral and bone strength in growing mice. Exper. Biol. Med. 2008, 233, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Scherer, P.E. Cell Biology. Ronning after the adiponectin receptors. Science 2013, 342, 1460–1461. [Google Scholar] [CrossRef] [PubMed]
- Kriketos, A.D.; Gan, S.K.; Poynten, A.M.; Furler, S.M.; Chisholm, D.J.; Campbell, L.V. Exercise increases adiponectin levels and insulin sensitivity in humans. Diabetes Care 2004, 27, 629–630. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Endocrine Cancer | APN Association to Cancer | Year | # of Studies | Ref. |
|---|---|---|---|---|
| Breast cancer | Inverse association. Low serum APN levels are associated with breast cancer in pre- and postmenopausal Asian women. | 2019 | 27 | [81] |
| Inverse association. APN is a biomarker of breast cancer risk in pre- and postmenopausal women, especially among Asians. | 2018 | 31 | [79] | |
| Inverse association. Low APN concentrations are associated with an increased risk of breast cancer. | 2014 | 15 | [71] | |
| Inverse association. Lower APN levels correlate with a higher risk of breast cancer in postmenopausal women. | 2014 | 8 | [73] | |
| Inverse association. High APN level might decrease the risk of postmenopausal breast cancer. | 2013 | 17 | [82] | |
| Endometrial cancer | Inverse association. Low APN level increases the risk of endometrial cancer. | 2016 | 18 | [96] |
| Inverse association. Higher APN levels might have a protective effect against endometrial cancer in postmenopausal women. | 2015 | 12 | [97] | |
| Inverse association. Higher serum APN concentrations are associated with a reduced risk of endometrial cancer, especially in postmenopausal women. | 2015 | 12 | [98] | |
| Inverse association. Each 1 µg/mL increase of APN level is associated with a 3% reduction in endometrial cancer risk. | 2015 | 12 | [99] | |
| Inverse association. Increased circulating APN and adiponectin/leptin ratio are associated with a decreased risk of endometrial cancer. | 2015 | 13 | [100] | |
| Ovarian cancer | Inverse association. The mean APN concentrations in patients with ovarian cancer are lower than those of the control group. | 2016 | 1 | [101] |
| Thyroid cancer | Inverse association. APN is inversely associated with thyroid cancer risk among women, but not among men. | 2018 | 1 | [102] |
| No association. No direct association between decreased levels of APN and papillary thyroid carcinoma size or stage was found. | 2018 | 1 | [103] | |
| No association. Serum APN levels are not significantly different between patients with or without medullary thyroid carcinomas. | 2016 | 1 | [104] | |
| Inverse association. Circulating APN is inversely associated with thyroid cancer risk. | 2011 | 1 | [105] | |
| Prostate cancer | Inverse association. Decreased concentration of APN is associated with a greater risk of prostate cancer. | 2015 | 11 | [106] |
| Direct association. The incidence of prostate cancer is increased in overweight men with high APN concentrations. | 2015 | 1 | [107] | |
| Inverse association. Higher APN levels reduce both the risk of developing high-grade prostate cancer and a risk of dying from the cancer. | 2010 | 1 | [108] | |
| Direct association. Serum APN levels are higher in advanced outside (relative to organ-confined) prostate cancers. | 2008 | 1 | [25] | |
| No association. APN was not of significantly associated with prostate cancer risk or high-grade disease. | 2006 | 1 | [109] | |
| Inverse association. APN levels are decreased in patients with prostate cancer and are also inversely associated with the histologic grade of the tumor. | 2005 | 1 | [110] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tumminia, A.; Vinciguerra, F.; Parisi, M.; Graziano, M.; Sciacca, L.; Baratta, R.; Frittitta, L. Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk. Int. J. Mol. Sci. 2019, 20, 2863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122863
Tumminia A, Vinciguerra F, Parisi M, Graziano M, Sciacca L, Baratta R, Frittitta L. Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk. International Journal of Molecular Sciences. 2019; 20(12):2863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122863
Chicago/Turabian StyleTumminia, Andrea, Federica Vinciguerra, Miriam Parisi, Marco Graziano, Laura Sciacca, Roberto Baratta, and Lucia Frittitta. 2019. "Adipose Tissue, Obesity and Adiponectin: Role in Endocrine Cancer Risk" International Journal of Molecular Sciences 20, no. 12: 2863. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122863