Sex-Dimorphic Behavioral Alterations and Altered Neurogenesis in U12 Intron Splicing-Defective Zrsr1 Mutant Mice

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
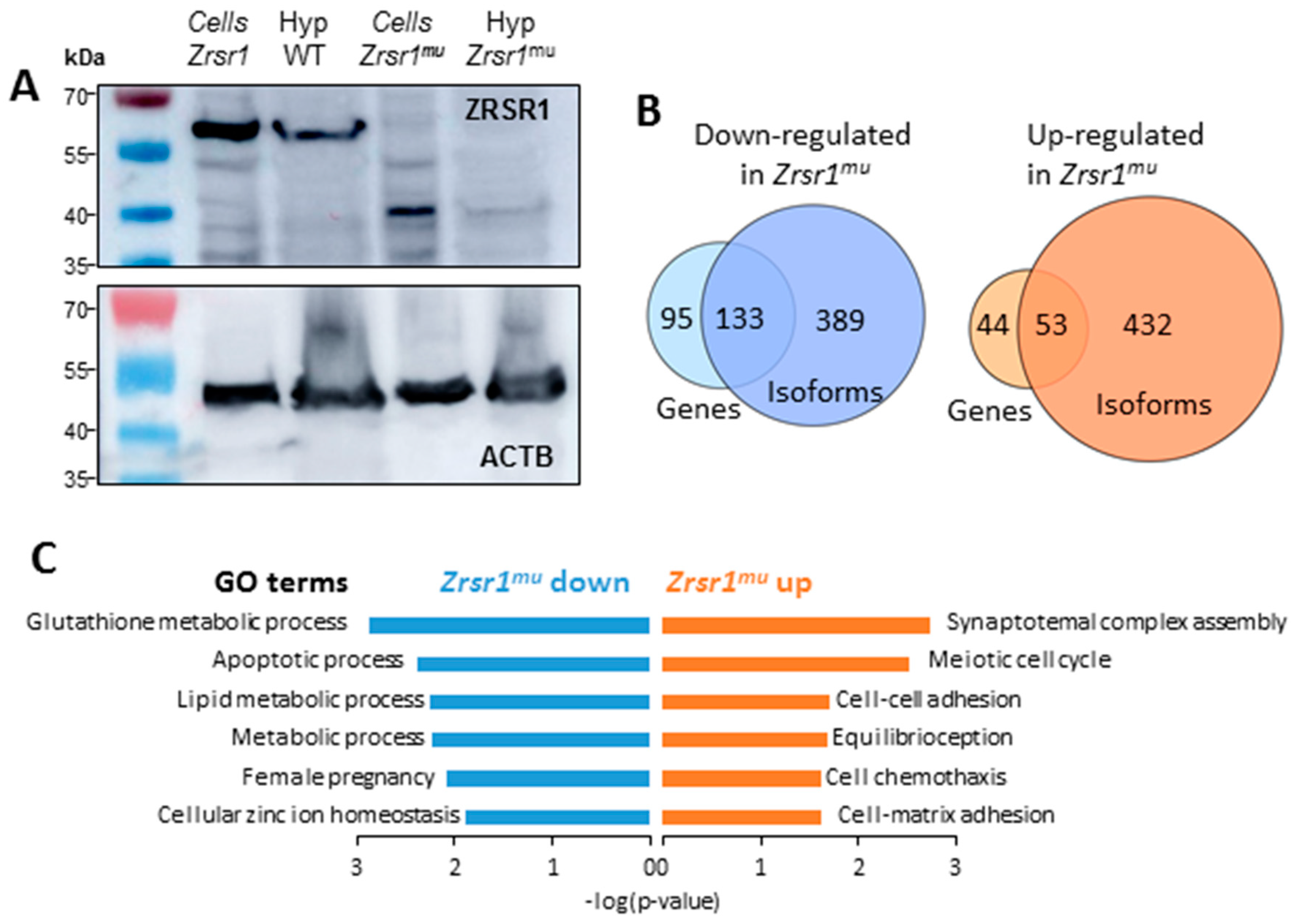
2.1. Expression of ZRSR1mu in the Hypothalamus
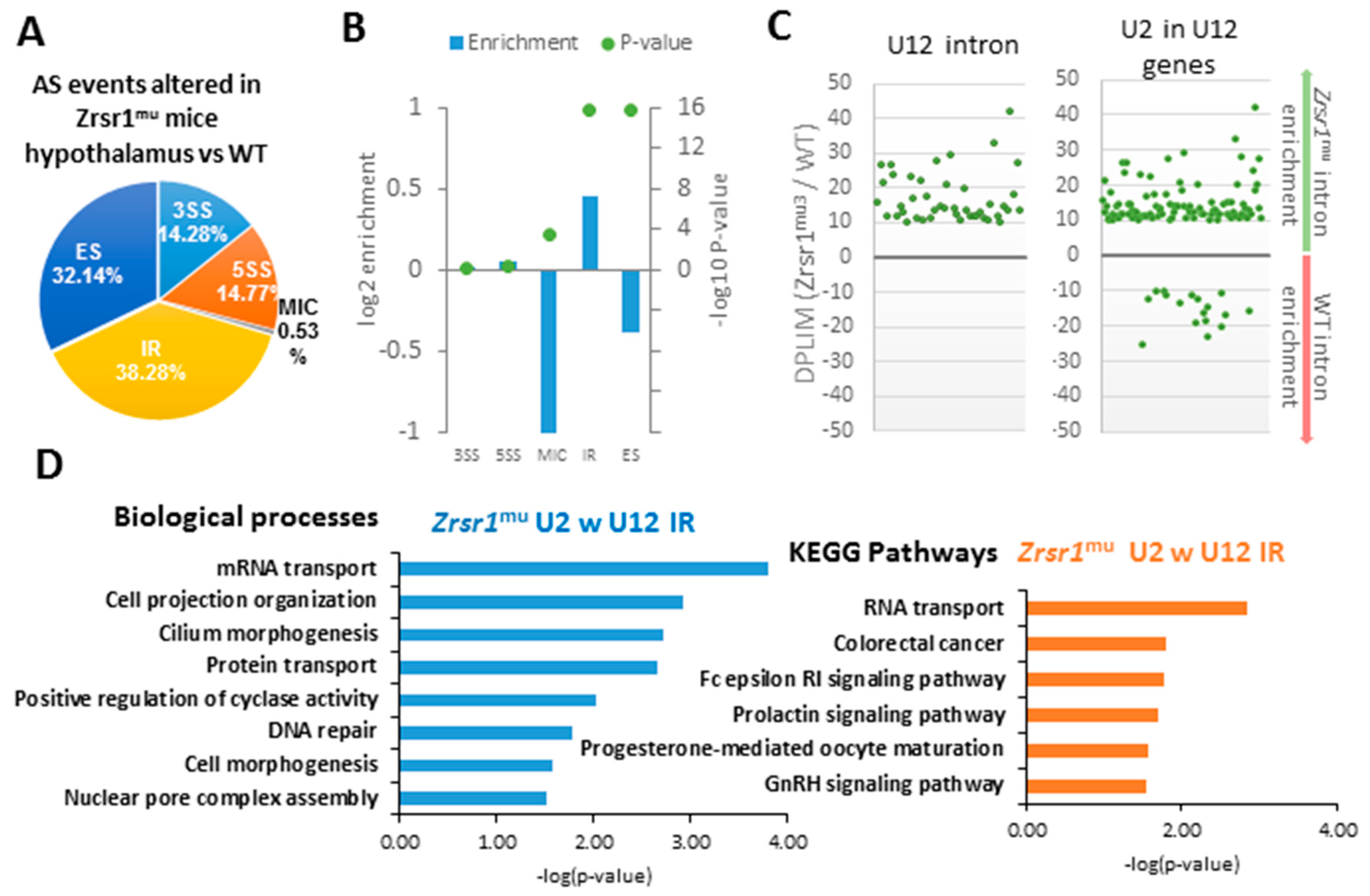
2.2. RNA-seq Analysis of Hypothalami from WT and Zrsr1mu Mice
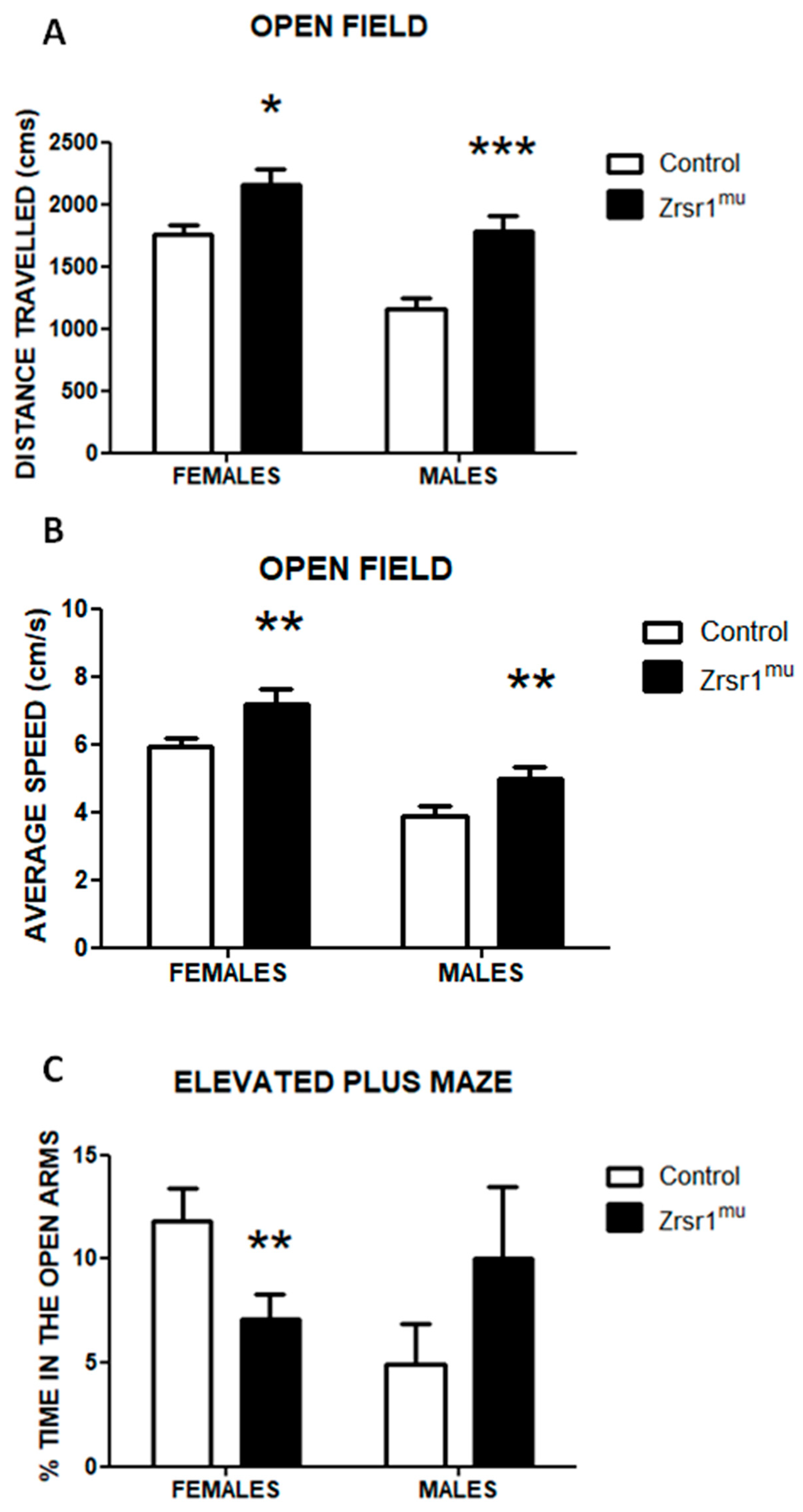
2.3. Spontaneous and Social Behaviors
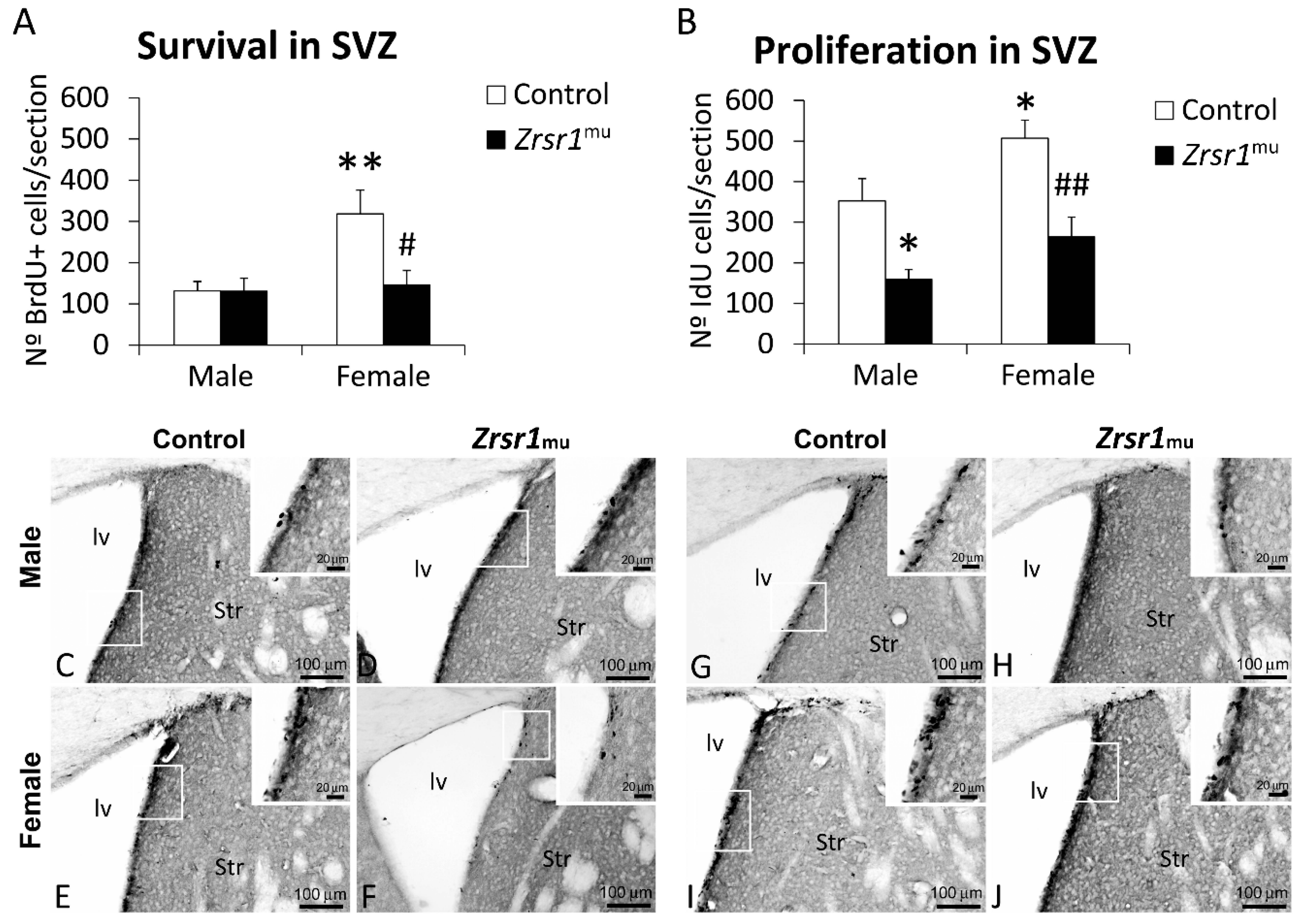
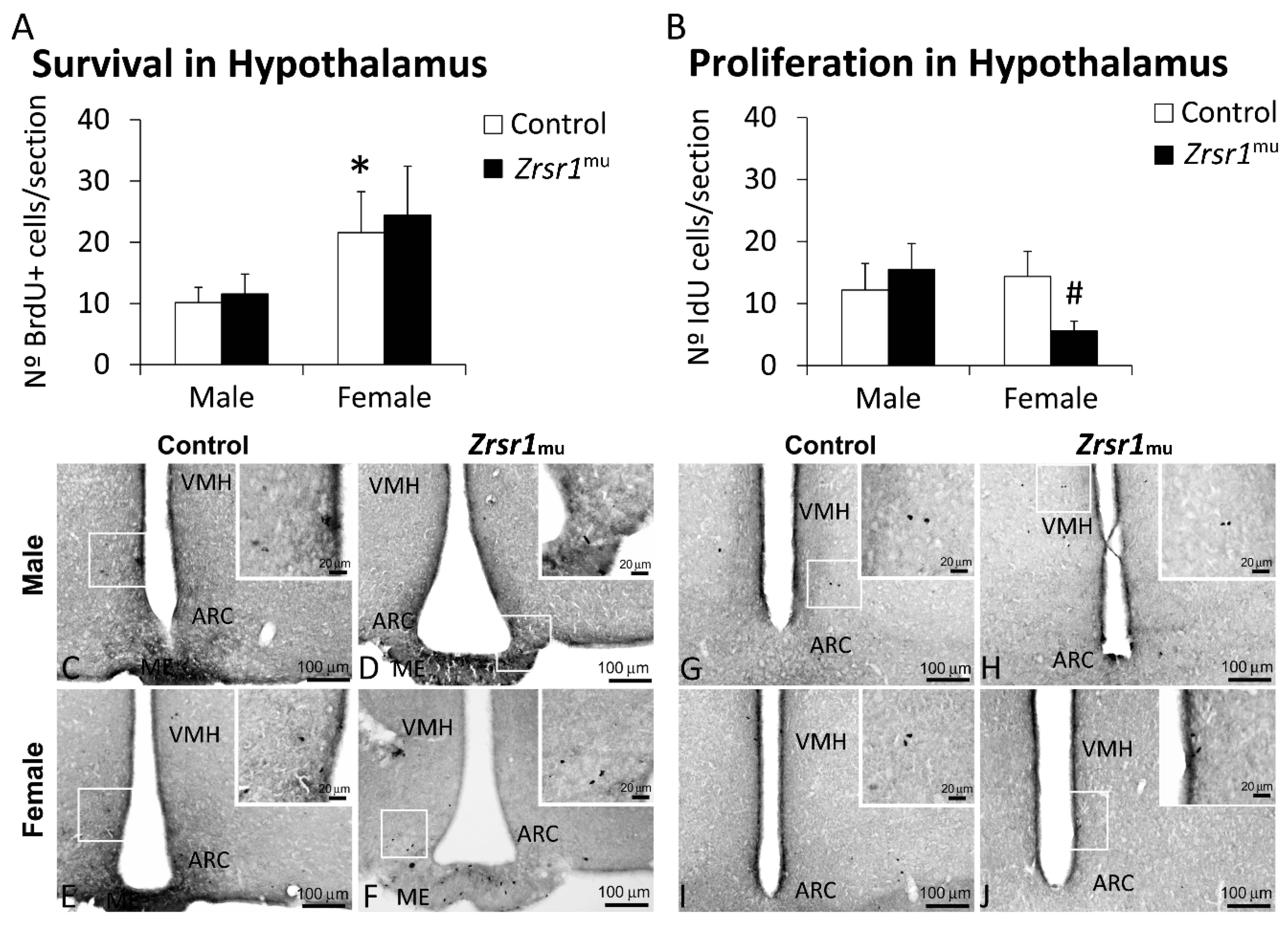
2.4. Cell Proliferation and Survival
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. RNA Extraction and RNA-seq Analysis
4.3. Differential Gene Expression Analysis
4.4. Differential Isoform Expression Analysis
4.5. Differential Alternative Splicing Analysis and GO Enrichment Analysis
4.6. Western Blotting
4.7. Open Field
4.8. Elevated Plus Maze
4.9. Social Behavior
4.10. Proliferation and Survival Analysis in the Hypothalamus and Subventricular Zone
4.10.1. Brain Collection
4.10.2. Immunohistochemistry
4.10.3. Quantification of BrdU- and IdU-Immunoreactive Cells
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Horiuchi, K.; Perez-Cerezales, S.; Papasaikas, P.; Ramos-Ibeas, P.; Lopez-Cardona, A.P.; Laguna-Barraza, R.; Fonseca Balvis, N.; Pericuesta, E.; Fernandez-Gonzalez, R.; Planells, B.; et al. Impaired Spermatogenesis, Muscle, and Erythrocyte Function in U12 Intron Splicing-Defective Zrsr1 Mutant Mice. Cell Rep. 2018, 23, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermejo-Alvarez, P.; Rizos, D.; Rath, D.; Lonergan, P.; Gutierrez-Adan, A. Sex determines the expression level of one third of the actively expressed genes in bovine blastocysts. Proc. Natl. Acad. Sci. USA 2010, 107, 3394–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Zheng, X.; Luecke, S.; Green, M.R. The U2AF35-related protein Urp contacts the 3’ splice site to promote U12-type intron splicing and the second step of U2-type intron splicing. Genes Dev. 2010, 24, 2389–2394. [Google Scholar] [CrossRef] [PubMed]
- Burge, C.B.; Padgett, R.A.; Sharp, P.A. Evolutionary fates and origins of U12-type introns. Mol. Cell 1998, 2, 773–785. [Google Scholar] [CrossRef]
- Sunahara, S.; Nakamura, K.; Nakao, K.; Gondo, Y.; Nagata, Y.; Katsuki, M. The oocyte-specific methylated region of the U2afbp-rs/U2af1-rs1 gene is dispensable for its imprinted methylation. Biochem. Biophys. Res. Commun. 2000, 268, 590–595. [Google Scholar] [CrossRef]
- Wang, Y.; Joh, K.; Masuko, S.; Yatsuki, H.; Soejima, H.; Nabetani, A.; Beechey, C.V.; Okinami, S.; Mukai, T. The mouse Murr1 gene is imprinted in the adult brain, presumably due to transcriptional interference by the antisense-oriented U2af1-rs1 gene. Mol. Cell Biol. 2004, 24, 270–279. [Google Scholar] [CrossRef]
- Hasin-Brumshtein, Y.; Khan, A.H.; Hormozdiari, F.; Pan, C.; Parks, B.W.; Petyuk, V.A.; Piehowski, P.D.; Brummer, A.; Pellegrini, M.; Xiao, X.; et al. Hypothalamic transcriptomes of 99 mouse strains reveal trans eQTL hotspots, splicing QTLs and novel non-coding genes. Elife 2016, 5, e15614. [Google Scholar] [CrossRef]
- Merico, D.; Roifman, M.; Braunschweig, U.; Yuen, R.K.; Alexandrova, R.; Bates, A.; Reid, B.; Nalpathamkalam, T.; Wang, Z.; Thiruvahindrapuram, B.; et al. Compound heterozygous mutations in the noncoding RNU4ATAC cause Roifman Syndrome by disrupting minor intron splicing. Nat. Commun. 2015, 6, 8718. [Google Scholar] [CrossRef]
- Turunen, J.J.; Niemela, E.H.; Verma, B.; Frilander, M.J. The significant other: Splicing by the minor spliceosome. Wiley Interdiscip. Rev. RNA 2013, 4, 61–76. [Google Scholar] [CrossRef]
- Chang, W.C.; Chen, Y.C.; Lee, K.M.; Tarn, W.Y. Alternative splicing and bioinformatic analysis of human U12-type introns. Nucleic Acids Res. 2007, 35, 1833–1841. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Liyanarachchi, S.; Akagi, K.; Nagy, R.; Li, J.; Dietrich, R.C.; Li, W.; Sebastian, N.; Wen, B.; Xin, B.; et al. Mutations in U4atac snRNA, a component of the minor spliceosome, in the developmental disorder MOPD I. Science 2011, 332, 238–240. [Google Scholar] [CrossRef]
- Elsaid, M.F.; Chalhoub, N.; Ben-Omran, T.; Kumar, P.; Kamel, H.; Ibrahim, K.; Mohamoud, Y.; Al-Dous, E.; Al-Azwani, I.; Malek, J.A.; et al. Mutation in noncoding RNA RNU12 causes early onset cerebellar ataxia. Ann. Neurol. 2017, 81, 68–78. [Google Scholar] [CrossRef]
- Edery, P.; Marcaillou, C.; Sahbatou, M.; Labalme, A.; Chastang, J.; Touraine, R.; Tubacher, E.; Senni, F.; Bober, M.B.; Nampoothiri, S.; et al. Association of TALS developmental disorder with defect in minor splicing component U4atac snRNA. Science 2011, 332, 240–243. [Google Scholar] [CrossRef]
- Doktor, T.K.; Hua, Y.; Andersen, H.S.; Broner, S.; Liu, Y.H.; Wieckowska, A.; Dembic, M.; Bruun, G.H.; Krainer, A.R.; Andresen, B.S. RNA-sequencing of a mouse-model of spinal muscular atrophy reveals tissue-wide changes in splicing of U12-dependent introns. Nucleic Acids Res. 2017, 45, 395–416. [Google Scholar] [CrossRef]
- Madan, V.; Kanojia, D.; Li, J.; Okamoto, R.; Sato-Otsubo, A.; Kohlmann, A.; Sanada, M.; Grossmann, V.; Sundaresan, J.; Shiraishi, Y.; et al. Aberrant splicing of U12-type introns is the hallmark of ZRSR2 mutant myelodysplastic syndrome. Nat. Commun. 2015, 6, 6042. [Google Scholar] [CrossRef]
- Xu, T.; Kim, B.M.; Kwak, K.J.; Jung, H.J.; Kang, H. The Arabidopsis homolog of human minor spliceosomal protein U11-48K plays a crucial role in U12 intron splicing and plant development. J. Exp. Bot. 2016, 67, 3397–3406. [Google Scholar] [CrossRef]
- Gault, C.M.; Martin, F.; Mei, W.; Bai, F.; Black, J.B.; Barbazuk, W.B.; Settles, A.M. Aberrant splicing in maize rough endosperm3 reveals a conserved role for U12 splicing in eukaryotic multicellular development. Proc. Natl. Acad. Sci. USA 2017, 114, E2195–E2204. [Google Scholar] [CrossRef]
- Otake, L.R.; Scamborova, P.; Hashimoto, C.; Steitz, J.A. The divergent U12-type spliceosome is required for pre-mRNA splicing and is essential for development in Drosophila. Mol. Cell 2002, 9, 439–446. [Google Scholar] [CrossRef]
- Markmiller, S.; Cloonan, N.; Lardelli, R.M.; Doggett, K.; Keightley, M.C.; Boglev, Y.; Trotter, A.J.; Ng, A.Y.; Wilkins, S.J.; Verkade, H.; et al. Minor class splicing shapes the zebrafish transcriptome during development. Proc. Natl. Acad. Sci. USA 2014, 111, 3062–3067. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, M.; Lemoine, C.; Al Seesi, S.; Karunakaran, D.K.; Sturrock, N.; Banday, A.R.; Kilcollins, A.M.; Mandoiu, I.; Kanadia, R.N. Minor splicing snRNAs are enriched in the developing mouse CNS and are crucial for survival of differentiating retinal neurons. Dev. Neurobiol. 2015, 75, 895–907. [Google Scholar] [CrossRef]
- Ohya, S.; Kito, H.; Hatano, N.; Muraki, K. Recent advances in therapeutic strategies that focus on the regulation of ion channel expression. Pharm. Ther. 2016, 160, 11–43. [Google Scholar] [CrossRef]
- Xie, J. Control of alternative pre-mRNA splicing by Ca(++) signals. Biochim. Biophys. Acta 2008, 1779, 438–452. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Miczek, K.A.; Maxson, S.C.; Fish, E.W.; Faccidomo, S. Aggressive behavioral phenotypes in mice. Behav. Brain Res. 2001, 125, 167–181. [Google Scholar] [CrossRef]
- Biagioli, M.; Pinto, M.; Cesselli, D.; Zaninello, M.; Lazarevic, D.; Roncaglia, P.; Simone, R.; Vlachouli, C.; Plessy, C.; Bertin, N.; et al. Unexpected expression of alpha- and beta-globin in mesencephalic dopaminergic neurons and glial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15454–15459. [Google Scholar] [CrossRef]
- Trarbach, E.B.; Silveira, L.G.; Latronico, A.C. Genetic insights into human isolated gonadotropin deficiency. Pituitary 2007, 10, 381–391. [Google Scholar] [CrossRef]
- Umathe, S.N.; Bhutada, P.S.; Jain, N.S.; Shukla, N.R.; Mundhada, Y.R.; Dixit, P.V. Gonadotropin-releasing hormone agonist blocks anxiogenic-like and depressant-like effect of corticotrophin-releasing hormone in mice. Neuropeptides 2008, 42, 399–410. [Google Scholar] [CrossRef]
- Bayerl, D.S.; Klampfl, S.M.; Bosch, O.J. More than reproduction: Central gonadotropin-releasing hormone antagonism decreases maternal aggression in lactating rats. J. Neuroendocr. 2019, e12709. [Google Scholar] [CrossRef]
- Floody, O.R.; Pfaff, D.W. Steroid hormones and aggressive behavior: Approaches to the study of hormone-sensitive brain mechanisms for behavior. Res. Publ. Assoc. Res. Nerv. Ment. Dis. 1974, 52, 149–185. [Google Scholar]
- Lammers, J.H.; Kruk, M.R.; Meelis, W.; van der Poel, A.M. Hypothalamic substrates for brain stimulation-induced attack, teeth-chattering and social grooming in the rat. Brain Res. 1988, 449, 311–327. [Google Scholar] [CrossRef]
- Falkner, A.L.; Grosenick, L.; Davidson, T.J.; Deisseroth, K.; Lin, D. Hypothalamic control of male aggression-seeking behavior. Nat. Neurosci. 2016, 19, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Yang, C.F.; Chizari, M.D.; Maheswaranathan, N.; Burke, K.J., Jr.; Borius, M.; Inoue, S.; Chiang, M.C.; Bender, K.J.; Ganguli, S.; et al. Social Control of Hypothalamus-Mediated Male Aggression. Neuron 2017, 95, 955–970.e4. [Google Scholar] [CrossRef]
- Trainor, B.C.; Greiwe, K.M.; Nelson, R.J. Individual differences in estrogen receptor alpha in select brain nuclei are associated with individual differences in aggression. Horm. Behav. 2006, 50, 338–345. [Google Scholar] [CrossRef]
- Sano, K.; Tsuda, M.C.; Musatov, S.; Sakamoto, T.; Ogawa, S. Differential effects of site-specific knockdown of estrogen receptor alpha in the medial amygdala, medial pre-optic area, and ventromedial nucleus of the hypothalamus on sexual and aggressive behavior of male mice. Eur. J. Neurosci. 2013, 37, 1308–1319. [Google Scholar] [CrossRef]
- Wu, M.V.; Manoli, D.S.; Fraser, E.J.; Coats, J.K.; Tollkuhn, J.; Honda, S.; Harada, N.; Shah, N.M. Estrogen masculinizes neural pathways and sex-specific behaviors. Cell 2009, 139, 61–72. [Google Scholar] [CrossRef]
- Rainville, J.R.; Weiss, G.L.; Evanson, N.; Herman, J.P.; Vasudevan, N.; Tasker, J.G. Membrane-initiated nuclear trafficking of the glucocorticoid receptor in hypothalamic neurons. Steroids 2019, 142, 55–64. [Google Scholar] [CrossRef]
- D’Anna, K.L.; Stevenson, S.A.; Gammie, S.C. Urocortin 1 and 3 impair maternal defense behavior in mice. Behav. Neurosci. 2005, 119, 1061–1071. [Google Scholar] [CrossRef]
- Trainor, B.C.; Crean, K.K.; Fry, W.H.; Sweeney, C. Activation of extracellular signal-regulated kinases in social behavior circuits during resident-intruder aggression tests. Neuroscience 2010, 165, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, A.L.; Shen, F.; Taravella, A.M.; Emery, S.; Veeramah, K.R.; Boyko, A.R.; Kidd, J.M. Comparison of village dog and wolf genomes highlights the role of the neural crest in dog domestication. BMC Biol. 2018, 16, 64. [Google Scholar] [CrossRef]
- Baumgartner, M.; Olthof, A.M.; Aquino, G.S.; Hyatt, K.C.; Lemoine, C.; Drake, K.; Sturrock, N.; Nguyen, N.; Al Seesi, S.; Kanadia, R.N. Minor spliceosome inactivation causes microcephaly, owing to cell cycle defects and death of self-amplifying radial glial cells. Development 2018, 145, dev166322. [Google Scholar] [CrossRef]
- Yoo, S.; Blackshaw, S. Regulation and function of neurogenesis in the adult mammalian hypothalamus. Prog. Neurobiol. 2018, 170, 53–66. [Google Scholar] [CrossRef]
- Mak, G.K.; Enwere, E.K.; Gregg, C.; Pakarainen, T.; Poutanen, M.; Huhtaniemi, I.; Weiss, S. Male pheromone-stimulated neurogenesis in the adult female brain: Possible role in mating behavior. Nat. Neurosci. 2007, 10, 1003–1011. [Google Scholar] [CrossRef]
- Su, C.H.; Dhananjaya, D.; Tarn, W.Y. Alternative Splicing in Neurogenesis and Brain Development. Front. Mol. Biosci. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Blekhman, R.; Marioni, J.C.; Zumbo, P.; Stephens, M.; Gilad, Y. Sex-specific and lineage-specific alternative splicing in primates. Genome Res. 2010, 20, 180–189. [Google Scholar] [CrossRef]
- Planells, B.; Gomez-Redondo, I.; Pericuesta, E.; Lonergan, P.; Gutierrez-Adan, A. Differential isoform expression and alternative splicing in sex determination in mice. BMC Genom. 2019, 20, 202. [Google Scholar] [CrossRef]
- Trabzuni, D.; Ramasamy, A.; Imran, S.; Walker, R.; Smith, C.; Weale, M.E.; Hardy, J.; Ryten, M. Widespread sex differences in gene expression and splicing in the adult human brain. Nat. Commun. 2013, 4, 2771. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Fernandez Banet, J.; Billis, K.; Garcia Giron, C.; Hourlier, T.; et al. The Ensembl gene annotation system. Database 2016, 2016, baw093. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Gould, T.D.; Dao, D.T.; Kovacsics, C.E. The Open Field Test. In Mood and Anxiety Related Phenotypes in Mice; Springer: Berlin, Germany, 2009; pp. 1–20. [Google Scholar]
- Fernandez-Gonzalez, R.; Moreira, P.; Bilbao, A.; Jimenez, A.; Perez-Crespo, M.; Ramirez, M.A.; Rodriguez De Fonseca, F.; Pintado, B.; Gutierrez-Adan, A. Long-term effect of in vitro culture of mouse embryos with serum on mRNA expression of imprinting genes, development, and behavior. Proc. Natl. Acad. Sci. USA 2004, 101, 5880–5885. [Google Scholar] [CrossRef]
- Navarro, J.F.; Rivera, A.; Maldonado, E.; Cavas, M.; de la Calle, A. Anxiogenic-like activity of 3,4-methylenedioxy-methamphetamine (“Ecstasy”) in the social interaction test is accompanied by an increase of c-fos expression in mice amygdala. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 249–254. [Google Scholar] [CrossRef]
- Blumstein, D.; Daniel, J.; Evans, C. JWatcher 1.0; San Francisco 2006. Available online: www.jwatcher.ucla.edu (accessed on 4 June 2019).
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Franklin, K.B.J., Paxinos, G., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands; London, UK, 2008. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Event Type | Zrsr1mu Up | Zrsr1mu Down | Total Events (Vast-Tools) | Enrichment (p-Value) |
|---|---|---|---|---|
| 3SS | 165 | 186 | 7048 | 0.0906 |
| 5SS | 182 | 181 | 7101 | 0.517 |
| ES | 444 | 346 | 20,935 | 2.20 × 10−16 |
| MIC | 8 | 5 | 643 | 3.82 × 10−4 |
| IR (U12) | 665 (98) | 276 (17) | 13,903 | 2.20 × 10−16 |
| Total | 1464 | 994 | 49,630 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alén, F.; Gómez-Redondo, I.; Rivera, P.; Suárez, J.; Ramos-Ibeas, P.; Pericuesta, E.; Fernández-González, R.; Perez-Cerezales, S.; Horiuchi, K.; Orio, L.; et al. Sex-Dimorphic Behavioral Alterations and Altered Neurogenesis in U12 Intron Splicing-Defective Zrsr1 Mutant Mice. Int. J. Mol. Sci. 2019, 20, 3543. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143543
Alén F, Gómez-Redondo I, Rivera P, Suárez J, Ramos-Ibeas P, Pericuesta E, Fernández-González R, Perez-Cerezales S, Horiuchi K, Orio L, et al. Sex-Dimorphic Behavioral Alterations and Altered Neurogenesis in U12 Intron Splicing-Defective Zrsr1 Mutant Mice. International Journal of Molecular Sciences. 2019; 20(14):3543. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143543
Chicago/Turabian StyleAlén, Francisco, Isabel Gómez-Redondo, Patricia Rivera, Juan Suárez, Priscila Ramos-Ibeas, Eva Pericuesta, Raul Fernández-González, Serafín Perez-Cerezales, Keiko Horiuchi, Laura Orio, and et al. 2019. "Sex-Dimorphic Behavioral Alterations and Altered Neurogenesis in U12 Intron Splicing-Defective Zrsr1 Mutant Mice" International Journal of Molecular Sciences 20, no. 14: 3543. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143543