Probiotics Upregulate Trefoil Factors and Downregulate Pepsinogen in the Mouse Stomach

 , ,
, , 
Abstract
:1. Introduction
2. Results
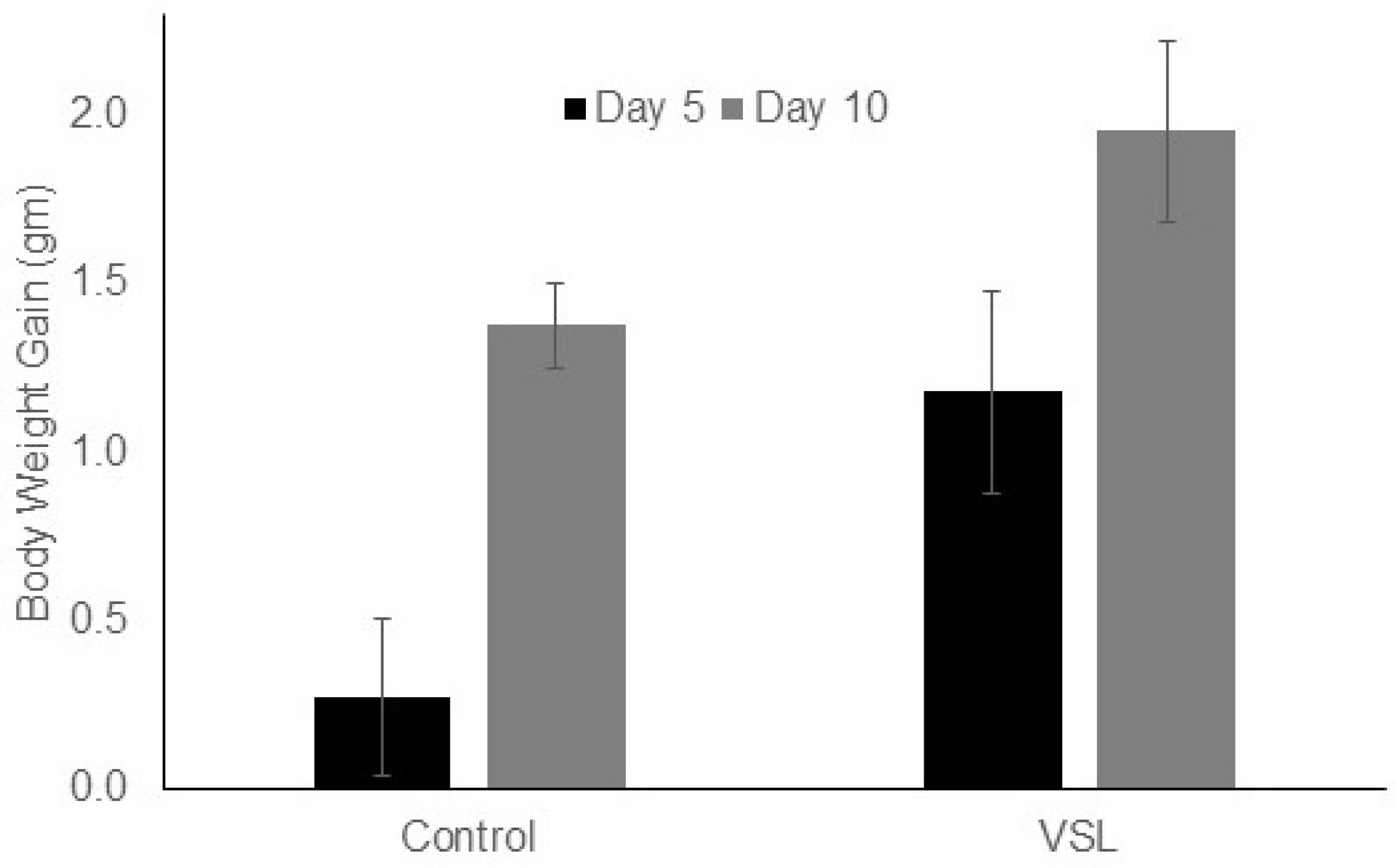
2.1. VSL Administration Has No Significant Effect on Mouse Bodyweight and Food Intake
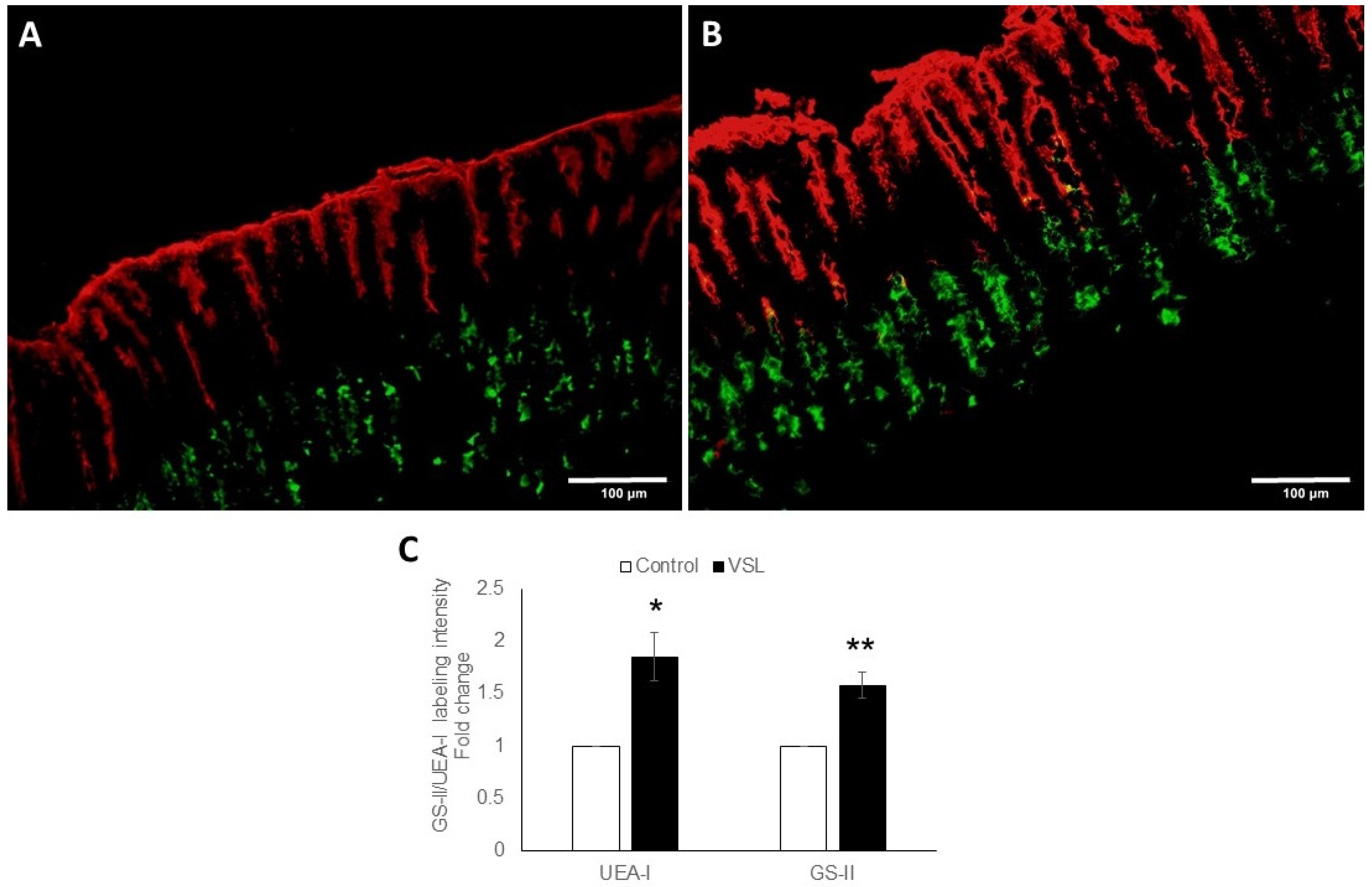
2.2. VSL Enhances Mucous Production in the Mouse Gastric Mucosa
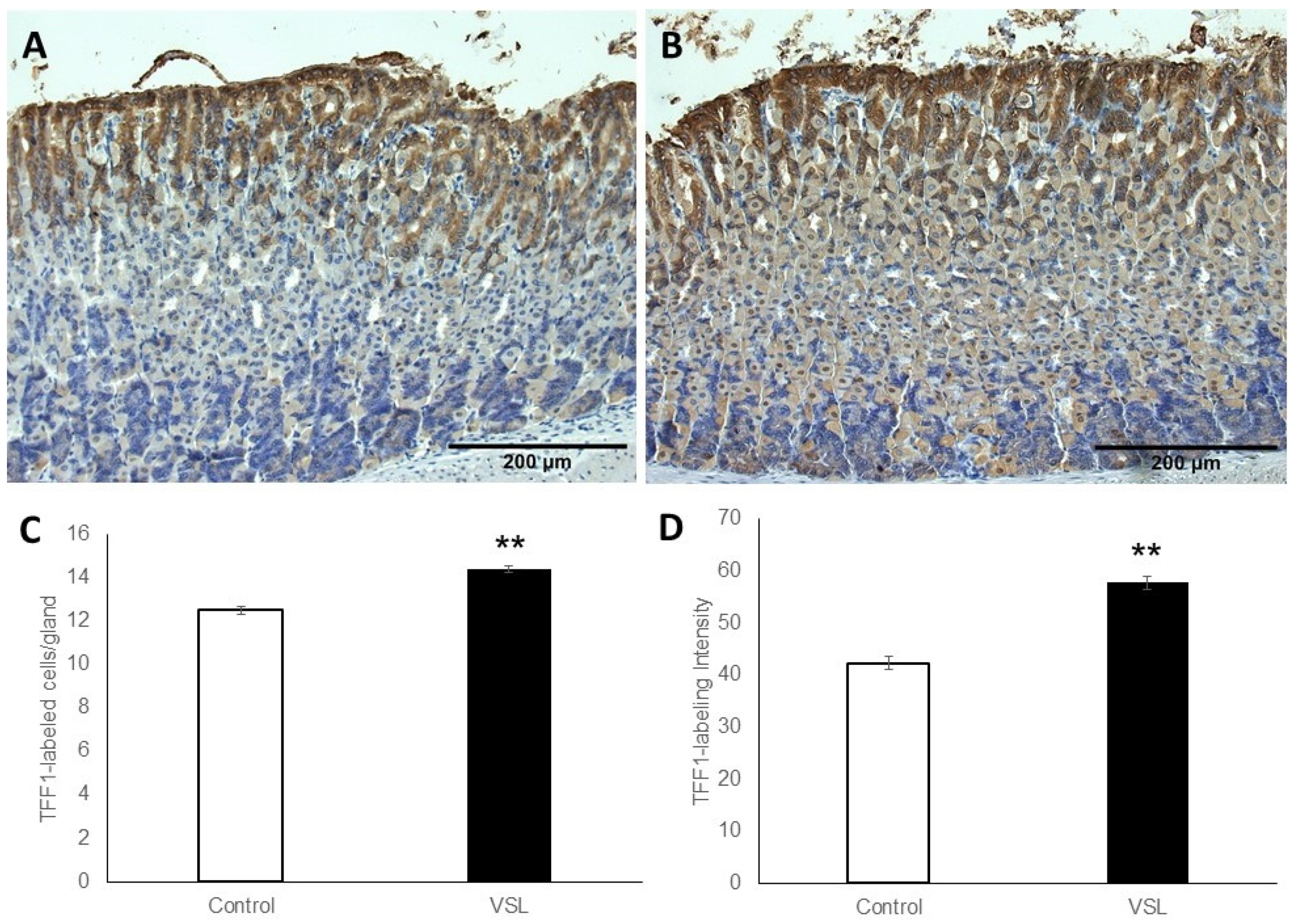
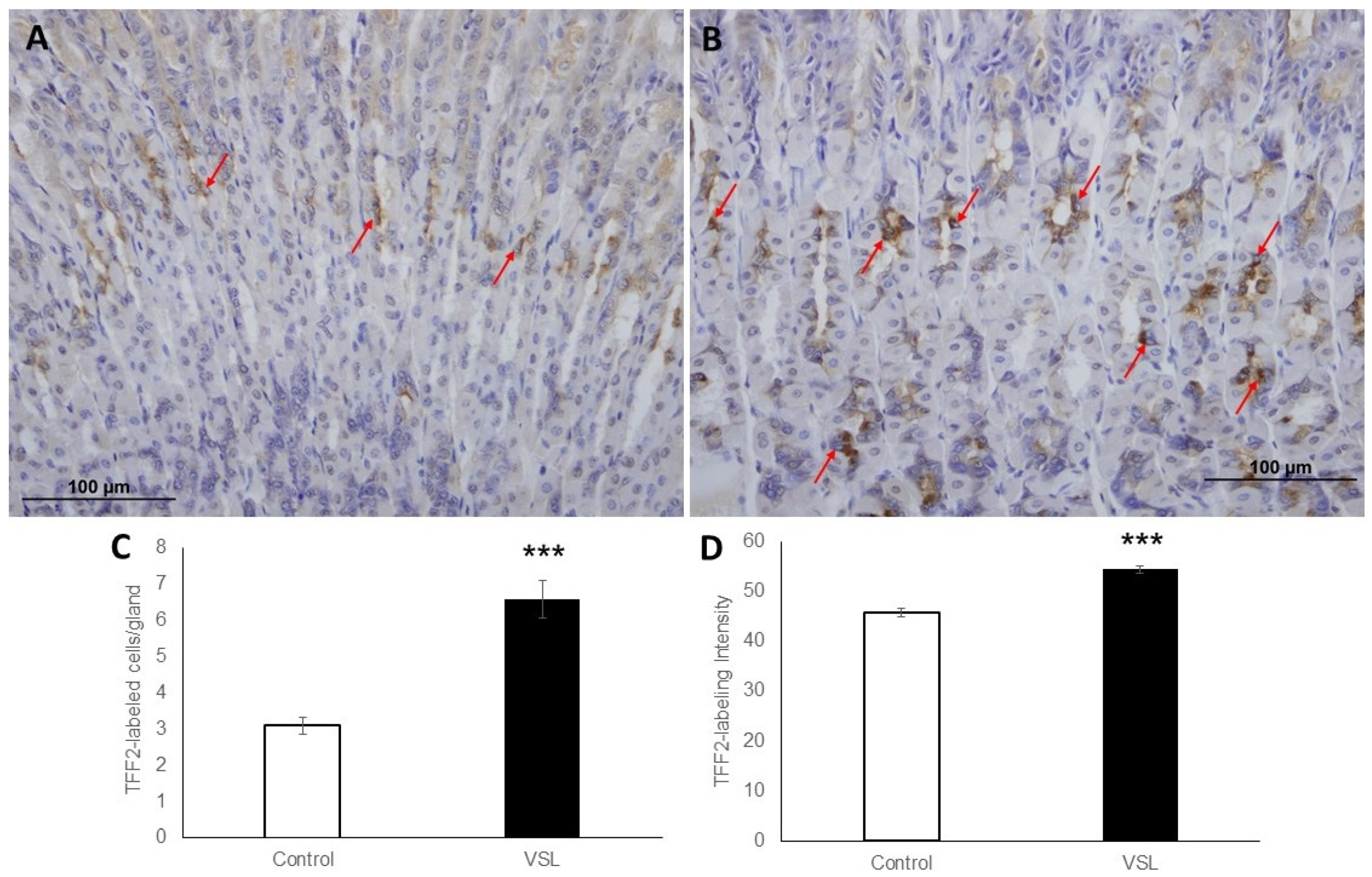
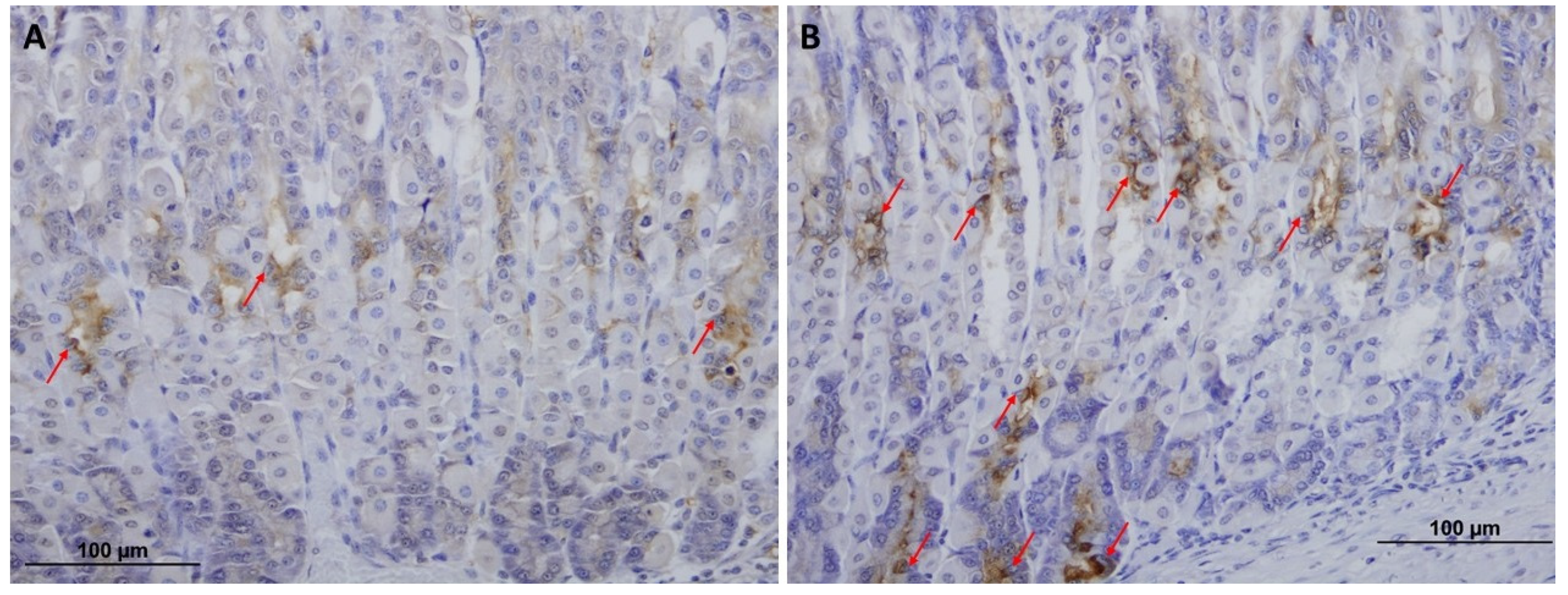
2.3. VSL Enhances TFF1 and TFF2 Production by Gastric Mucous Cells
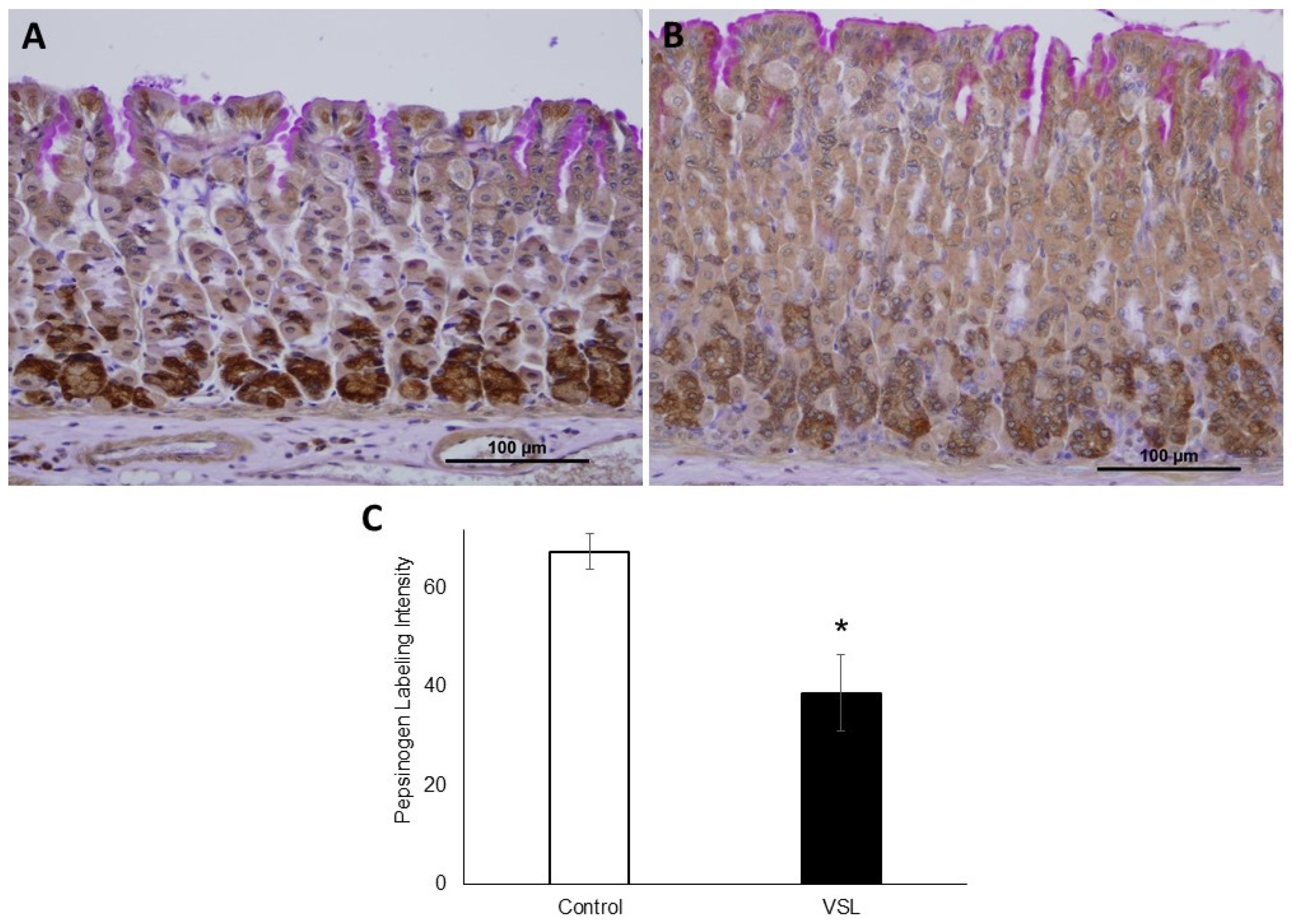
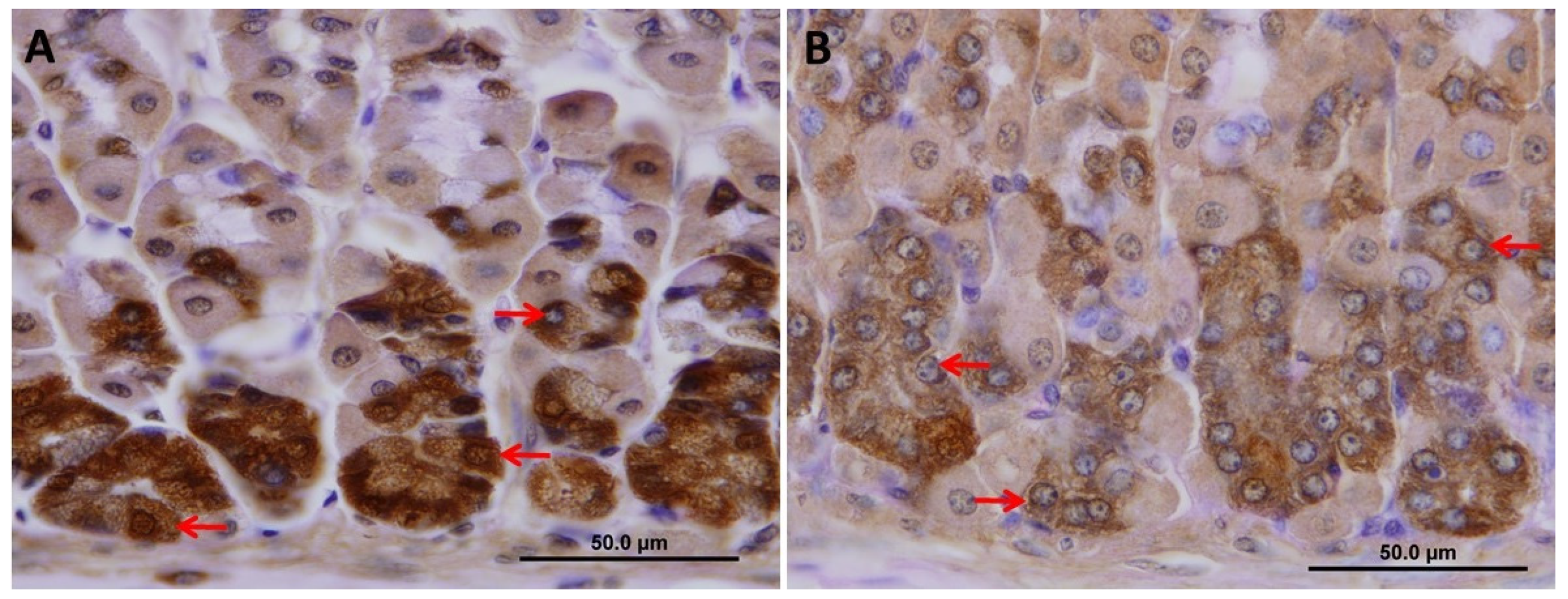
2.4. VSL Inhibits Pepsinogen Production by Chief Cells
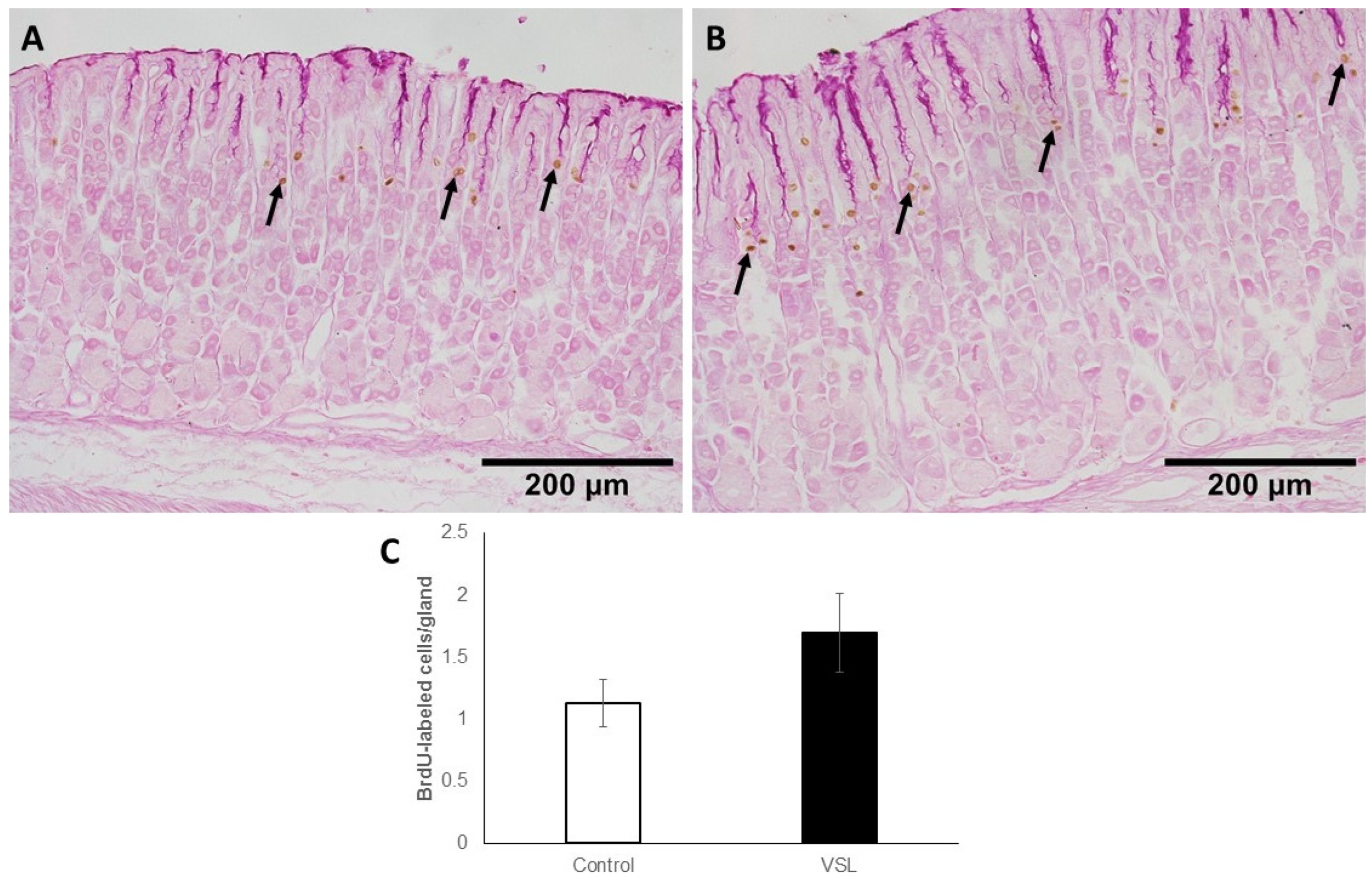
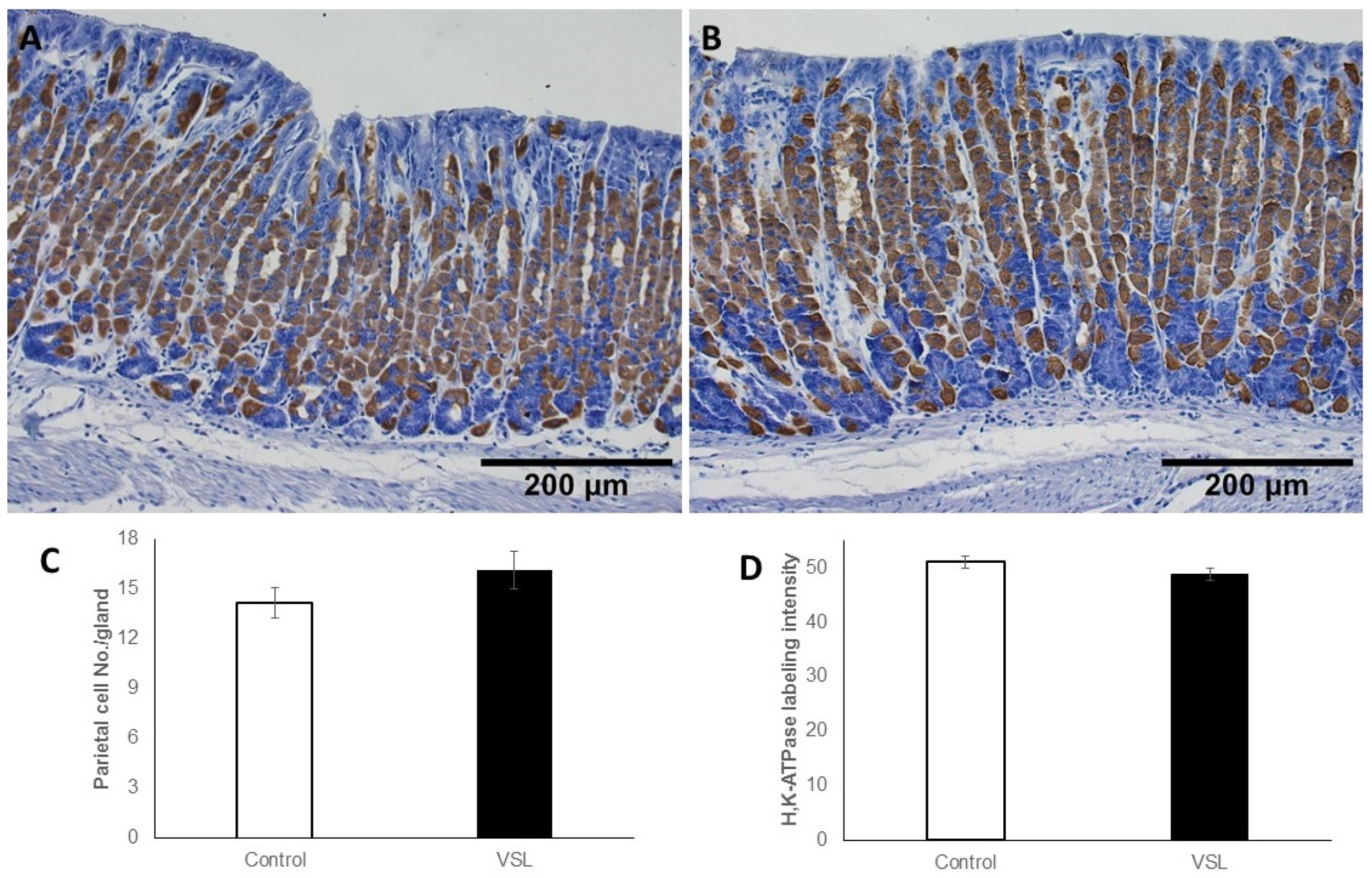
2.5. VSL Treatment neither Affect Cell Proliferation nor H+,K+-ATPase Immunolabeling
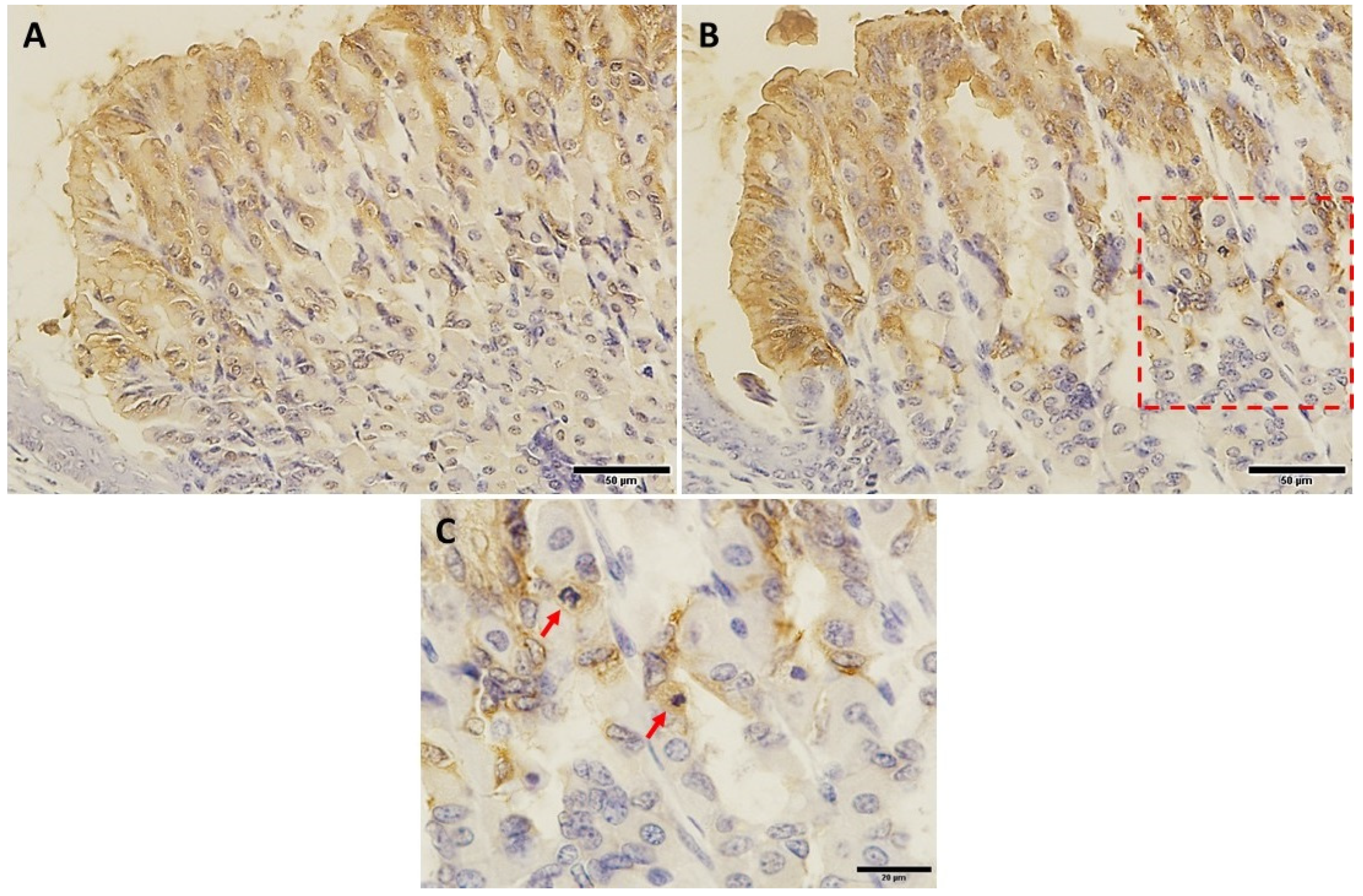
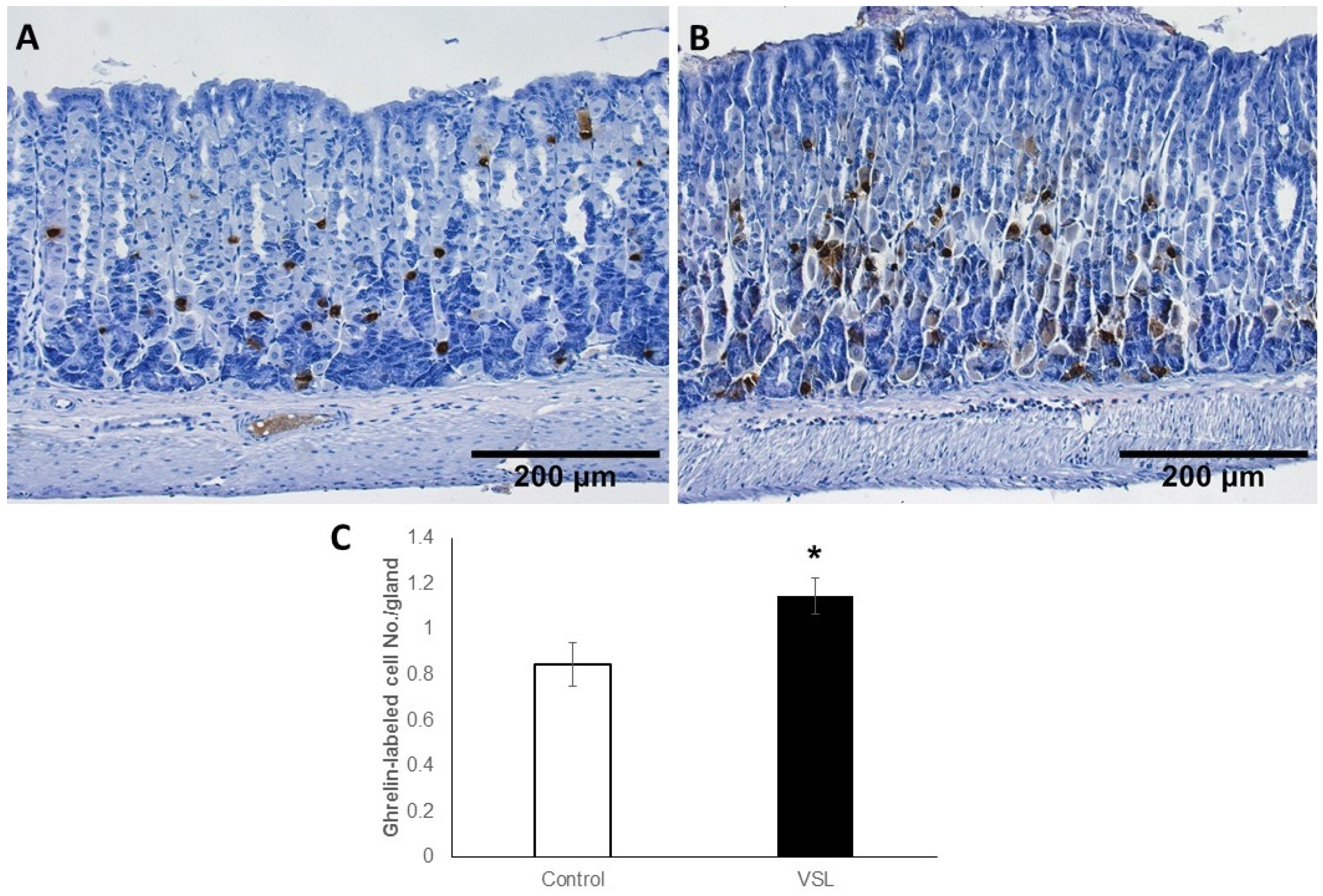
2.6. VSL Treated Tissues Showed an Increase in Ghrelin-Secreting Cells
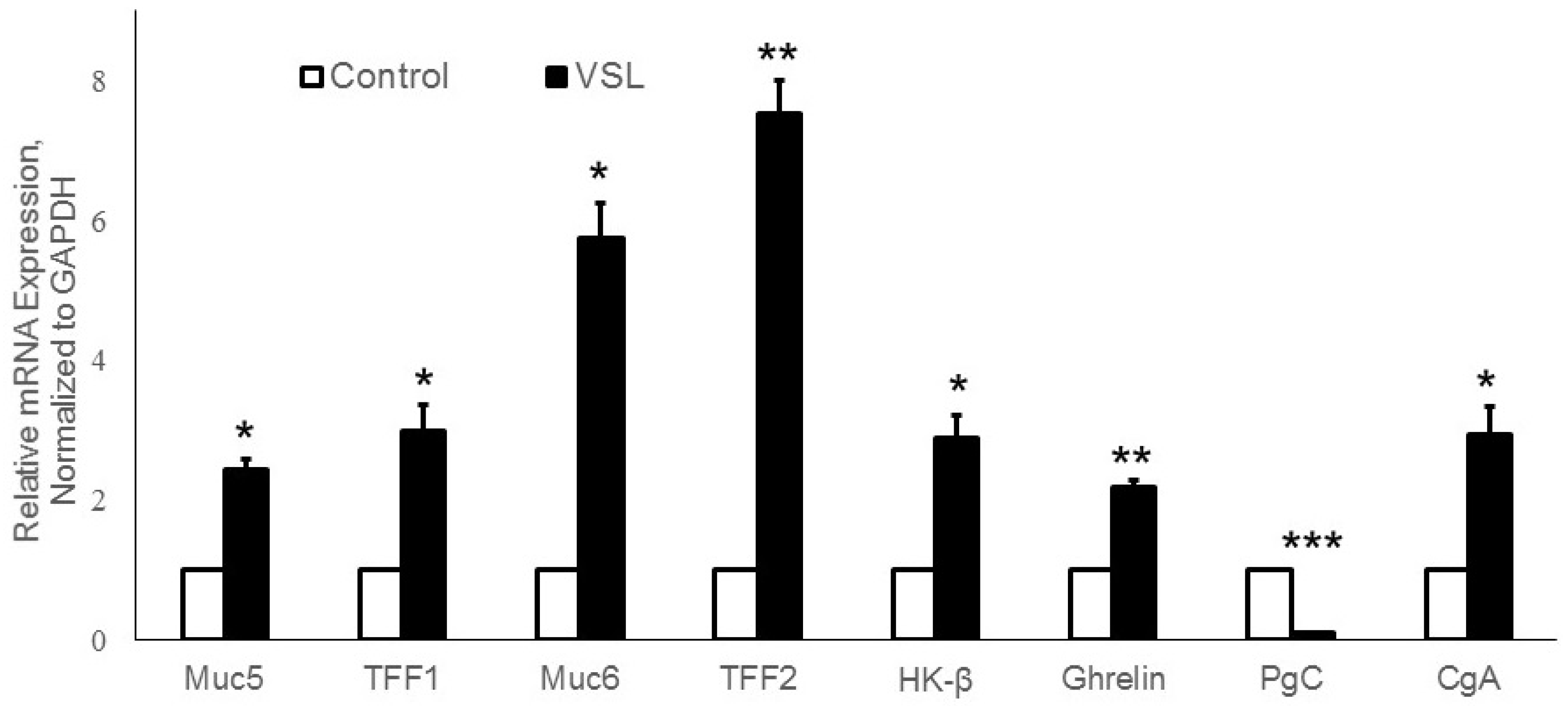
2.7. Effects of VSL Treatment on the Expression of Various Gastric Cell Lineage-Specific Genes
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Histological, Lectin Histochemistry, and Immunohistochemical Analysis
4.4. Immunohistochemical Quantification
4.5. RT-qPCR Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 119, 506–514. [Google Scholar] [CrossRef]
- Didari, T.; Solki, S.; Mozaffari, S.; Nikfar, S.; Abdollahi, M. A systematic review of the safety of probiotics. Expert Opin. Drug Saf. 2014, 13, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Fooks, L.; Gibson, G. Probiotics as modulators of the gut flora. Br. J. Nutr. 2002, 88, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, J. Clinical applications of probiotic agents. Am. J. Clin. Nutr. 2001, 73, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.; Guarner, F.; Guerrant, R.; Holt, P.; Quigley, E.; Sartor, R.; Sherman, P.; Mayer, E. An update on the use and investigation of probiotics in health and disease. Gut 2013, 62, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Isolauri, E. Probiotics in human disease. Am. J. Clin. Nutr. 2001, 73, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T.; Otoshi, T.; Hazama, D.; Kiriu, T.; Umezawa, K.; Katsurada, N.; Nishimura, Y. Novel cancer therapy targeting microbiome. OncoTargets Ther. 2019, 12, 3619–3624. [Google Scholar] [CrossRef] [PubMed]
- Pala, V.; Sieri, S.; Berrino, F.; Vineis, P.; Sacerdote, C.; Palli, D.; Masala, G.; Panico, S.; Mattiello, A.; Tumino, R.; et al. Yogurt consumption and risk of colorectal cancer in the Italian European prospective investigation into cancer and nutrition cohort. Int. J. Cancer 2011, 129, 2712–2719. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Li, W.B.; Wang, H.Y.; Ma, Y.M.; Zhao, X.H.; Yang, H.; Qian, J.M.; Li, J.N. VSL#3 can prevent ulcerative colitis-associated carcinogenesis in mice. World J. Gastroenterol. 2018, 24, 4254–4262. [Google Scholar]
- Javanmard, A.; Ashtari, S.; Sabet, B.; Davoodi, S.H.; Rostami-Nejad, M.; Esmaeil Akbari, M.; Niaz, A.; Mortazavian, A.M. Probiotics and their role in gastrointestinal cancers prevention and treatment; an overview. Gastroenterol. Hepatol. Bed Bench. 2018, 11, 284–295. [Google Scholar]
- Khoder, G.; Al Menhali, A.; Al-Yassir, F.; Karam, S. Potential role of probiotics in the management of gastric ulcer. Exp. Ther. Med. 2016, 12, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Gomez-Llorente, C.; Fontana, L.; Gil, A. Modulation of immunity and inflammatory gene expression in the gut, in inflammatory diseases of the gut and in the liver by probiotics. World J. Gastroenterol. 2014, 20, 15632–15649. [Google Scholar] [CrossRef] [PubMed]
- Delia, P.; Sansotta, G.; Donato, V.; Messina, G.; Frosinta, P.; Pergolixxi, S.; De Renzis, C.; Famularo, G. Prevention of radiation-induced diarrhea with the use of VSL#3, a new high potency probiotic preparation. Am. J. Gastroenterol. 2002, 97, 2150–2152. [Google Scholar] [PubMed]
- Mennigen, R.; Nolte, K.; Rijcken, E.; Utech, M.; Loeffler, B.; Senninger, N.; Bruewer, M. Probiotic mixture VSL#3 protects the epithelial barrier by maintaining tight junction protein expression and preventing apoptosis in a murine model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, 1140–1149. [Google Scholar]
- Suo, H.; Zhao, X.; Qian, Y.; Sun, P.; Zhu, K.; Li, J.; Sun, B. Lactobacillus fermentum Suo attenuates HCL/Ethanol induced gastric injury in mice through its antioxidant effects. Nutrients 2016, 8, 155. [Google Scholar] [CrossRef]
- Singh, P.; Deol, P.; Kaur, I. Entrapment of Lactobacillus acidophilus into alginate beads for the effective treatment of cold restraint stress induced gastric ulcer. Food Funct. 2012, 3, 83–90. [Google Scholar] [CrossRef]
- De Simone, C. Letter: What gastroenterologists should know about VSL#3. Aliment Pharmacol Ther. 2018, 47, 698–699. [Google Scholar]
- Ki Cha, B.; Mun Jung, S.; Hwan Choi, C.; Song, I.; Woong Lee, H.; Joon Kim, H.; Hyuk, J.; Kyung Chang, S.; Kim, K.; Chung, W.; et al. The effect of a multispecies probiotic mixture on the symptoms and fecal microbiota in diarrhea-dominant irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. J. Clin. Gastroenterol. 2012, 46, 220–227. [Google Scholar] [CrossRef]
- Dharmani, P.; De Simone, C.; Chadee, K. The probiotic mixture VSL#3 accelerates gastric ulcer healing by stimulating vascular endothelial growth factor. PLoS ONE 2013, 8, 58671. [Google Scholar]
- Dai, C.; Zhao, D.; Jiang, M. VSL#3 probiotics regulate the intestinal epithelial barrier in vivo and in vitro via the p38 and ERK signaling pathways. Int. J. Mol. Med. 2012, 29, 202–208. [Google Scholar]
- Meng, X.; Wang, Y.; Yan, P.; Li, Y.; Wang, H.; Qian, J.; Li, J. Effect of VSL#3 and S. Bouldarii on intestinal microbiota in mice with acute colitis. Zhonghua Yi Xue Za Zhi 2019, 99, 1735–1742. [Google Scholar]
- Kim, H.J.; Vazquez Roque, M.I.; Camilleri, M.; Stephens, D.; Burton, D.D.; Baxter, K.; Thomforde, G.; Zinsmeister, A.R. A randomized controlled trial of a probiotic combination VSL# 3 and placebo in irritable bowel syndrome with bloating. Neurogastroenterol. Motil. 2005, 17, 687–696. [Google Scholar]
- Mimura, T.; Rizzello, F.; Helwig, U.; Poggioli, G.; Schreiber, S.; Talbot, I.C.; Nicholls, R.J.; Gionchetti, P.; Campieri, M.; Kamm, M.A. Once daily high dose probiotic therapy (VSL#3) for maintaining remission in recurrent or refractory pouchitis. Gut 2004, 53, 108–114. [Google Scholar] [Green Version]
- Fedorak, R.; Feagan, B.; Hotte, N.; Leddin, D.; Dieleman, L.; Petrunia, D.; Enns, R.; Bitton, A.; Chiba, N.; Pare, P.; et al. The probiotic VSL#3 has anti-inflammatory effects and could reduce endoscopic recurrence after surgery for Crohn’s disease clinical gastroenterology and hepatology: The official clinical practice. J. Am. Gastroenterol. Assoc. 2015, 13, 928–935. [Google Scholar]
- Mardini, H.; Grigorian, A. Probiotic mix VSL#3 is effective adjunctive therapy for mild to moderately active ulcerative colitis: A meta-analysis. Inflamm. Bowel Dis. 2014, 20, 1562–1567. [Google Scholar]
- Bibiloni, R.; Fedorak, R.N.; Tannock, G.W.; Madsen, K.L.; Gionchetti, P.; Campieri, M.; De Simone, C.; Sartor, R.B. VSL#3 probiotic-mixture induces remission in patients with active ulcerative colitis. Am. J. Gastroenterol. 2005, 100, 1539–1546. [Google Scholar]
- Linsalata, M.; Russo, F.; Berloco, P.; Valentini, A.; Caruso, M.; De Simone, C.; Barone, M.; Polimeno, L.; Di Leo, A. Effects of Probiotics Bacteria (VSL#3) on the Polyamine Biosynthesis and Cell Proliferation of Normal Colonic Mucosa of Rats. In Vivo 2005, 19, 989–996. [Google Scholar]
- Sherman, P.; Ossa, C.; Johnson-Henry, K. Unraveling mechanisms of action of Probiotics. Nutr. Clin. Pract. 2009, 24, 10–14. [Google Scholar] [CrossRef]
- Roller, M.; Rechkemmer, G.; Watzi, B. Prebiotic inulin with oligofructose in combination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis modulates intestinal immune functions in rats. J. Nutr. 2004, 134, 153–156. [Google Scholar] [CrossRef]
- Wan, L.; Chen, Z.; Shah, N.; El-Nezami, H. Modulation of intestinal epithelial defense responses by probiotic bacteria. Crit. Rev. Food Sci. Nutr. 2016, 56, 2628–2641. [Google Scholar] [CrossRef]
- Khailova, L.; Dvorak, K.; Arganbright, K.; Halpern, M.; Kinouchi, T.; Yajima, M.; Dvorak, B. Bifidobacterium bifidum improves intestinal integrity in a rat model of necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, 940–949. [Google Scholar] [CrossRef]
- Moon, G.; Myung, S.; Jeong, J.; Yang, S.; Cho, Y.; Lee, S.; Chang, H.; Byeon, J.; Lee, Y.; Lee, G.; et al. Prophylactic effect of Lactobacillus GG in animal colitis and its effect on cytokine secretion and mucin gene expressions. Korean J. Gastroenterol. 2004, 43, 234–245. [Google Scholar]
- Mattar, A.F.; Teitelbaum, D.H.; Drongowski, R.A.; Yongyi, F.; Harmon, C.M.; Coran, A.G. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr. Surg. Int. 2002, 18, 586–590. [Google Scholar]
- Otte, J.M.; Podolsky, D.K. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G613–G626. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G315–G322. [Google Scholar]
- Karam, S.M.; Straiton, T.; Hassan, W.M.; Leblond, C.P. Defining epithelial cell progenitors in the human oxyntic mucosa. Stem Cells 2003, 21, 322–336. [Google Scholar] [CrossRef]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on Gut Microbiota in Patients with Lactose Intolerance and Persisting Functional Gastrointestinal Symptoms: A Randomised, Double-Blind, Cross-Over Study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Liu, Y.; Shi, W.; Zhang, J.; Dong, E.; Zhang, X.; Tao, Y.; Zhong, J. Lactobacillus plantarum PFM 105 Promotes Intestinal Development Through Modulation of Gut Microbiota in Weaning Piglets. Front. Microbiol. 2019, 10, 90. [Google Scholar] [CrossRef]
- Russo, F.; Linsalata, M.; Orlando, A. Probiotics against neoplastic transformation of gastric mucosa: Effects on cell proliferation and polyamine metabolism. World J. Gastroenterol. 2014, 20, 13258–13272. [Google Scholar] [CrossRef]
- Lam, K.; Tai, K.; Koo, W.; Wong, P.; Wu, K.; Yu, L.; So, H.; Woo, C.; Cho, H. Enhancement of gastric mucosal integrity by Lactobacillus rhamnosus GG. Life Sci. 2007, 80, 2128–2136. [Google Scholar] [CrossRef]
- Rasouli, B.S.; Ghadimi-Darsajini, A.; Nekouian, R.; Iragian, G.R. In vitro activity of probiotic Lactobacillus reuteri against gastric cancer progression by downregulation of urokinase plasminogen activator/urokinase plasminogen activator receptor gene expression. J. Cancer Res. Ther. 2017, 13, 246–251. [Google Scholar]
- Orlando, A.; Refolo, M.G.; Messa, C.; Amati, L.; Lavermicocca, P.; Guerra, V.; Russo, F. Antiproliferative and proapoptotic effects of viable or heat-killed Lactobacillus paracasei IMPC2.1 and Lactobacillus rhamnosus GG in HGC-27 gastric and DLD-1 colon cell lines. Nutr. Cancer. 2012, 64, 1103–1111. [Google Scholar] [CrossRef]
- Babińska, I.; Rotkiewicz, T.; Otrocka-Domagała, I. The effect of Lactobacillus acidophilus and Bifidobacterium spp. administration on the morphology of the gastrointestinal tract, liver and pancreas in piglets. Pol. J. Vet. Sci. 2005, 8, 29–35. [Google Scholar]
- Rodriguez, C.; Medici, M.; Mozzi, F.; Font de Valdez, G. Therapeutic effect of Streptococcus thermophilus CRL1190-fermented milk on chronic gastritis. World J. Gastroenterol. 2010, 16, 1622–1630. [Google Scholar] [CrossRef]
- Gomi, A.; Harima Mizusawa, N.; Shibahara Sone, H.; Kano, M.; Miyazaki, K.; Ishikawa, F. Effect of Bifidobacterium bifidum BF 1 on gastric protection and mucin production in an acute gastric injury rat model. J. Dairy Sci. 2013, 96, 832–837. [Google Scholar] [CrossRef]
- Vinderola, G.; Matar, C.; Perdigon, G. Milk fermentation products of L. helveticus R389 activate calcinurin as a signal to promote gut mucosal immunity. BMC Immunol. 2007, 8, 19. [Google Scholar] [CrossRef]
- Gaudier, E.; Michel, C.; Segain, J.; Cherbut, C.; Hoebler, C. The VSL#3 probiotic mixture modifies microflora but does not heal chronic dextran sodium sulphate induced colitis or reinforce the mucus barrier in mice. J. Nutr. 2005, 135, 2753–2761. [Google Scholar]
- Hoffmann, W. TFF2, a MUC6-binding lectin stabilizing the gastric mucus barrier and more. Int. J. Oncol. 2015, 47, 806–816. [Google Scholar] [CrossRef]
- Kjellev, S. The trefoil factor family-small peptides with multiple functionalities. Cell Mol. Life Sci. 2009, 66, 1350–1369. [Google Scholar] [CrossRef]
- Stürmer, R.; Müller, S.; Hanisch, F.; Hoffmann, W. Porcine Gastric TFF2 is a Mucus Constituent and Differs from Pancreatic TFF2. Cell. Physiol. Biochem. 2014, 33, 895–904. [Google Scholar] [CrossRef]
- Braga Emidio, N.; Hoffmann, W.; Brierley, S.M.; Muttenthaler, M. Trefoil Factor Family: Unresolved Questions and Clinical Perspectives. Trends Biochem. Sci. 2019, 44, 387–390. [Google Scholar] [CrossRef]
- Stürmer, R.; Harder, S.; Schlüter, H.; Hoffmann, W. Commercial pocrine gastric mucin preparations, also used as artificial saliva, are a rich source for the lectin TFF2: In vitro binding studies. Chem. Biochem. 2018, 19, 2598–2608. [Google Scholar]
- Reeves, E.; Ali, T.; Leonard, P.; Hearty, S.; O’Kennedy, R.; May, F.; Westley, B.; Josenhans, C.; Rust, M.; Suerbaum, S.; et al. Helicobacter pylori lipopolysaccharide interacts with TFF1 in pH-dependent manner. Gastroenterology 2008, 135, 2043–2054. [Google Scholar] [CrossRef]
- Farrell, J.; Taupin, D.; Koh, T.; Chen, D.; Zhao, C.; Podolsky, D.; Wang, T. TFF2/SP-deficient mice show decreased gastric proliferation, increased acid secretion, and increased susceptibility to NSAID injury. J. Clin. Investig. 2002, 109, 193–204. [Google Scholar] [CrossRef]
- Tomasetto, C.; Rio, M.C.; Gautier, C.; Wolf, C.; Hareuveni, M.; Chambon, P.; Lathe, R. hSP, the domain-duplicated homolog of pS2 protein, is co-expressed with pS2 in stomach but not in breast carcinoma. EMBO J. 1990, 9, 407–414. [Google Scholar] [CrossRef]
- Hoffmann, W.; Jagla, W.; Wiede, A. Molecular medicine of TFF-peptides: From gut to brain. Histol. Histopathol. 2001, 16, 319–334. [Google Scholar]
- Karam, S.; Tomasetto, C.; Rio, M. Trefoil Factor 1 is required for the commitment program of mouse oxyntic epithelial progenitors. Gut 2004, 53, 1408–1415. [Google Scholar] [CrossRef]
- Aihara, E.; Engevik, K.A.; Montrose, M.H. Trefoil Factor Peptides and Gastrointestinal Function. Annu. Rev. Physiol. 2017, 79, 357–380. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, W. Trefoil factors TFF (trefoil factor family) peptide-triggered signals promoting mucosal restitution. Cell. Mol. Life Sci. 2005, 62, 2932–2938. [Google Scholar] [CrossRef]
- Oretel, M.; Graness, A.; Thim, L.; Bühling, F.; Kalbacher, H.; Hoffmann, W. Trefoil factor family-peptides promote migration of human bronchial epithelial cells: Synergistic effect with epidermal growth factor. Am. J. Respir. Cell Mol. Biol. 2001, 25, 418–424. [Google Scholar] [CrossRef]
- Bossenmeyer-Pourie, C.; Kannan, R.; Ribieras, S.; Wendling, C.; Stoll, I.; Thim, L.; Tomasetto, C.; Rio, M. The trefoil factor 1 participates in gastrointestinal cell differentiation by delaying G1-S phase transition and reducing apoptosis. J. Cell Biol. 2002, 157, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Ulaganathan, M.; Familari, M.; Yeomans, N.; Giraud, A.; Cook, G. Spatio-temporal expression of trefoil peptide following severe gastric ulceration in the rat implicates it in late-stage repair processes. J. Gastroenterol. Hepatol. 2001, 16, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.; Dünker, N. Trefoil factor family peptides—Friends or foes? BioMol. Concepts. 2015, 6, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, K.; Hans, W.; Van Huysse, J.; Neirynck, S.; Demetter, P.; Remaut, E.; Rottiers, P.; Steidler, L. Active delivery of trefoil factors by genetically modified Lactococcus lactis prevents and heals acute colitis in mice. Gastroenterology 2004, 127, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kirpich, I.; Liu, Y.; Ma, Z.; Barve, S.; McClain, C.J.; Feng, W. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am. J. Pathol. 2011, 179, 2866–2875. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Y.; Sidhu, A.; Ma, Z.; McClain, C.; Feng, W. Lactobacillus rhamnosus GG culture supernatant ameliorates acute alcohol-induced intestinal permeability and liver injury. Am. J. Physiol. Gastrointest Liver Physiol. 2012, 303, G32–G41. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Przybyszewski, J.; Mitra, D.; Becker, C.; Brehm-Stecher, B.; Tentinger, A.; MacDonald, R.S. Soy protein diet, but not Lactobacillus rhamnosus GG, decreases mucin-1, trefoil factor-3, and tumor necrosis factor-α in colon of dextran sodium sulfate-treated C57BL/6 mice. J. Nutr. 2011, 141, 1239–1246. [Google Scholar] [CrossRef]
- Schmitz, S.; Glanemann, B.; Garden, O.A.; Brooks, H.; Chang, Y.M.; Werling, D.; Allenspach, K. A prospective, randomized, blinded, placebo-controlled pilot study on the effect of Enterococcus faecium on clinical activity and intestinal gene expression in canine food-responsive chronic enteropathy. J. Vet. Intern. Med. 2015, 29, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Scholven, J.; Taras, D.; Sharbati, S.; Schön, J.; Gabler, C.; Huber, O.; Meyer zum Büschenfelde, D.; Blin, N.; Einspanier, R. Intestinal expression of TFF and related genes during postnatal development in a piglet probiotic trial. Cell. Physiol. Biochem. 2009, 23, 143–156. [Google Scholar] [CrossRef]
- Miki, K.; Urita, Y.; Ishikawa, F.; Iino, T.; Shibahara-Sone, H.; Akahoshi, R.; Mizusawa, S.; Nose, A.; Nozaki, D.; Hirano, K.; et al. Effect of Bifidobacterium bifidum fermented milk on Helicobacter pylori and serum pepsinogen levels in humans. J. Dairy Sci. 2007, 90, 2630–2640. [Google Scholar] [CrossRef]
- Cinque, B.; La Torre, C.; Lombardi, F.; Palumbo, P.; Van der Rest, M.; Cifone, M.G. Production Conditions Affect the In Vitro Anti-Tumoral Effects of a High Concentration Multi-Strain Probiotic Preparation. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ekmekciu, I.; von Klitzing, E.; Fiebiger, U.; Neumann, C.; Bacher, P.; Scheffold, A.; Bereswill, S.; Heimesaat, M.M. The Probiotic Compound VSL#3 Modulates Mucosal, Peripheral, and Systemic Immunity Following Murine Broad-Spectrum Antibiotic Treatment. Front. Cell Infect. Microbiol. 2017, 7, 167. [Google Scholar] [PubMed]
- Gioacchini, G.; Ciani, E.; Pessina, A.; Cecchini, C.; Silvi, S.; Rodiles, A.; Merrifield, D.; Olivotto, I.; Carnevali, O. Effects of lactogen 13, a new probiotic preparation on gut microbiota and endocrine signals controlling growth and appetite of Oreochromis niloticus Juveniles. Microb. Ecol. 2018, 76, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Nakakita, Y.; Segawa, S.; Tsuchiya, Y. Oral administration of heat-killed Lactobacillus brevis SBC8803 elevates the ratio of acyl/des-acyl ghrelin in blood and increases short-term food intake. Benef. Microbes. 2019, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Warzechai, Z.; Dembinski, A.; Ceranowicz, P.; Dembinski, M.; Cieszkowski, J.; Konturek, S.; Polus, A.; Pawlik, W.; Kuwahara, A.; Kato, I.; et al. Influence of ghrelin on gastric and duodenal growth and expression of digestive enzymes in young mature rats. J. Physiol. Pharmacol. 2006, 57, 425–437. [Google Scholar]
- Plovier, H.; Cani, P.D. Enteroendocrine Cells: Metabolic Relays between Microbes and Their Host. Endocr. Dev. 2017, 32, 139–164. [Google Scholar] [PubMed]
- Latorre, R.; Sternini, C.; De Giorgio, R.; Greenwood-Van Meerveld, B. Enteroendocrine Cells: A Review of Their Role in Brain-Gut Communication. Neurogastroenterol. Motil. 2016, 28, 620–630. [Google Scholar] [CrossRef]
- Paim, F.; Langel, S.; Fischer, D.; Kandasamy, S.; Shao, L.; Alhamo, M.; Huang, H.; Kumar, A.; Rajashekara, G.; Saif, L.; et al. Effects of Escherichia coli Nissle 1917 and Ciprofloxacin on small intestinal epithelial cell mRNA expression in the neonatal piglet model of human rotavirus infection. Gut Pathog. 2016, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Yadav, H.; Lee, J.H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| GAPDH | Forward | TCAAGAAGGTGGTGAAGCAGG |
| Reverse | TATTATGGGGGTCTGGGATGG | |
| Muc5 | Forward | AGG GCC CAG TGA GCA TCT CCTA |
| Reverse | CAT CAT CGC AGC GCA GAG TCA | |
| TFF2 | Forward | GCA GTG CTT TGA TCT TGG ATG C |
| Reverse | TCA GGT TGG AAA AGC AGC AGTT | |
| HK-β | Forward | AAC AGA ATT GTC AAG TTC CTC |
| Reverse | AGA CTG AAG GTG CCA TTG | |
| Ghrl | Forward | AGGAATCCAAGAAGCCACCAGCTA |
| Reverse | ATGCCAACATCGAAGGGAGCATTG | |
| Muc6 | Forward | CTC ACC TTC TAC CCC AGT ATC A |
| Reverse | GGC AAC GAG TTA GAG TCA CAT T | |
| TFF1 | Forward | GGCCCAGGAAGAAACATGTATC |
| Reverse | ACTGCTGGGCGGTGACA | |
| PgC | Forward | AAACCGGCATCATGAAGTGGATGG |
| Reverse | TTGTTCCTTCATGGTCTCCCGGAT | |
| CgA | Forward | GCA GCA TCC AGT TCC CAC TTC C |
| Reverse | TCC CCA TCT TCC TCC TGC TGA G |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoder, G.; Al-Yassir, F.; Al Menhali, A.; Saseedharan, P.; Sugathan, S.; Tomasetto, C.; Karam, S.M. Probiotics Upregulate Trefoil Factors and Downregulate Pepsinogen in the Mouse Stomach. Int. J. Mol. Sci. 2019, 20, 3901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20163901
Khoder G, Al-Yassir F, Al Menhali A, Saseedharan P, Sugathan S, Tomasetto C, Karam SM. Probiotics Upregulate Trefoil Factors and Downregulate Pepsinogen in the Mouse Stomach. International Journal of Molecular Sciences. 2019; 20(16):3901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20163901
Chicago/Turabian StyleKhoder, Ghalia, Farah Al-Yassir, Asma Al Menhali, Prashanth Saseedharan, Subi Sugathan, Catherine Tomasetto, and Sherif M. Karam. 2019. "Probiotics Upregulate Trefoil Factors and Downregulate Pepsinogen in the Mouse Stomach" International Journal of Molecular Sciences 20, no. 16: 3901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20163901