Human Circulating miRNAs Real-time qRT-PCR-based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization


Abstract
:1. Introduction
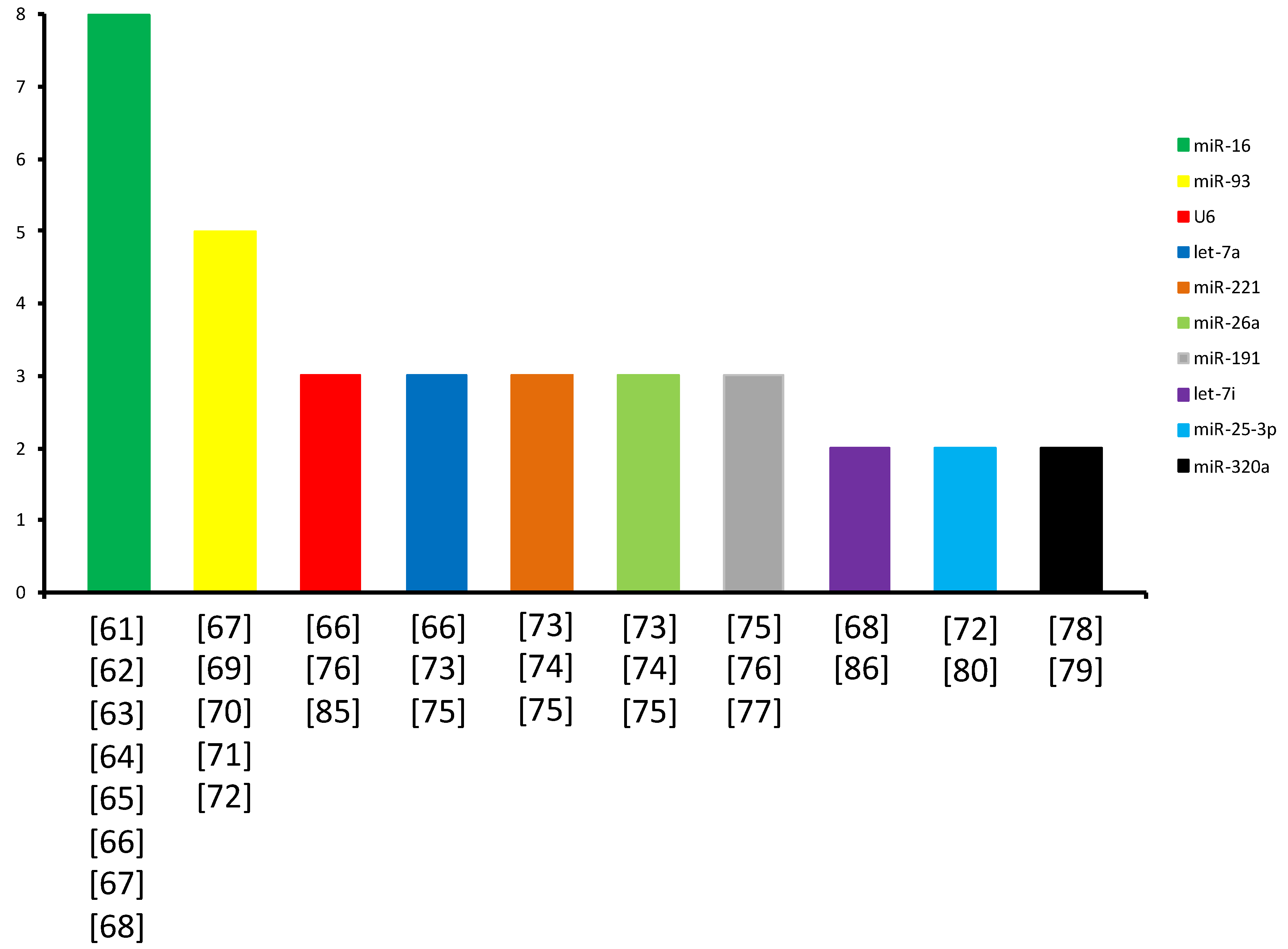
2. Best Identified miRNAs as Potential RGs
2.1. miR-16
2.2. miR-93
2.3. Combination of miR-221, miR-26a
2.4. miR-191
2.5. miR-320a
2.6. Other Genes Adopted as Reference
3. Discussion
Author Contributions
Conflicts of Interest
References
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynt, A.S.; Lai, E.C. Biological principles of microRNA-mediated regulation: Shared themes amid diversity. Nat. Rev. Genet. 2008, 9, 831–842. [Google Scholar] [CrossRef]
- Hsu, P.W.; Huang, H.D.; Hsu, S.D.; Lin, L.Z.; Tsou, A.P.; Tseng, C.P.; Stadler, P.F.; Washietl, S.; Hofacker, I.L. miRNAMap: Genomic maps of microRNA genes and their target genes in mammalian genomes. Nucleic Acids Res. 2006, 34, D135–D139. [Google Scholar] [CrossRef]
- Malan-Müller, S.; Hemmings, S.M.; Seedat, S. Big effects of small RNAs: A review of microRNAs in anxiety. Mol. Neurobiol. 2013, 47, 726–739. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell. 2016, 64, 320–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forman, J.J.; Legesse-Miller, A.; Coller, H.A. A search for conserved sequences in coding regions reveals that the let-7 microRNA targets Dicer within its coding sequence. Proc. Natl. Acad. Sci. USA 2008, 105, 14879–14884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, W.; Liu, Y.; Liu, T.; Li, C.; Wang, L. Oncogenic role of microRNA-532-5p in human colorectal cancer via targeting of the 5’UTR of RUNX3. Oncol. Lett. 2018, 15, 7215–7220. [Google Scholar] [CrossRef]
- Dharap, A.; Pokrzywa, C.; Murali, S.; Pandi, G.; Vemuganti, R. MicroRNA miR-324-3p induces promoter-mediated expression of RelA gene. PLoS ONE 2013, 8, e79467. [Google Scholar] [CrossRef]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Joshua-Tor, L. From guide to target: Molecular insights into eukaryotic RNA-interference machinery. Nat. Struct. Mol. Biol. 2015, 22, 20–28. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.; Plasterk, R.H.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.; Iwasaki, S.; Liu, Q.; Tomari, Y. ATP-dependent human RISC assembly pathways. Nat. Struct. Mol. Biol. 2010, 17, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Khvorova, A.; Reynolds, A.; Jayasena, S.D. Functional siRNAs and miRNAs exhibit strand bias. Cell 2003, 115, 209–216. [Google Scholar] [CrossRef]
- Reid, G.; Kirschner, M.B.; van Zandwijk, N. Circulating microRNAs: Association with disease and potential use as biomarkers. Crit. Rev. Oncol. Hematol. 2011, 80, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Chim, S.S.; Shing, T.K.; Hung, E.C.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.; Lo, Y.M. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Zhang, Y.; Chen, J.; Guo, X.; Li, Q.; et al. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Iftikhar, H.; Carney, G.E. Evidence and potential in vivo functions for biofluid miRNAs: From expression profiling to functional testing: Potential roles of extracellular miRNAs as indicators of physiological change and as agents of intercellular information exchange. Bioessays 2016, 38, 367–378. [Google Scholar] [CrossRef]
- Cogswell, J.P.; Ward, J.; Taylor, I.A.; Waters, M.; Shi, Y.; Cannon, B.; Cannon, B.; Kelnar, K.; Kemppainen, J.; Brown, D.; et al. Identification of miRNA changes in Alzheimer’s disease brain and CSF yields putative biomarkers and insights into disease pathways. J. Alzheimers Dis. 2008, 14, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, M.; Wang, X.; Li, Q.; Wang, T.; Zhu, Q.; Zhou, X.; Wang, X.; Gao, X.; Li, X. Immune-related microRNAs are abundant in breast milk exosomes. Int. J. Biol. Sci. 2012, 8, 118–123. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.C.; Veeramachaneni, D.N.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-secreted vesicles in equine ovarian follicular fluid contain miRNAs and proteins: A possible new form of cell communication within the ovarian follicle. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef]
- Tsui, N.B.; Ng, E.K.; Lo, Y.M. Stability of endogenous and added RNA in blood specimens, serum, and plasma. Clin. Chem. 2002, 48, 1647–1653. [Google Scholar] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular miRNAs: The mystery of their origin and function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Théry, C.; Regnault, A.; Garin, J.; Wolfers, J.; Zitvogel, L.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Molecular characterization of dendritic cell-derived exosomes. Selective accumulation of the heat shock protein hsc73. J. Cell Biol. 1999, 147, 599–610. [Google Scholar] [CrossRef]
- Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, N.; Lankar, D.; Faure, F.; Regnault, A.; Dumont, C.; Raposo, G.; Hivroz, C. TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/ζ complex. J. Immunol. 2002, 168, 3235–3241. [Google Scholar] [CrossRef]
- Raposo, G.; Tenza, D.; Mecheri, S.; Peronet, R.; Bonnerot, C.; Desaymard, C. Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol. Biol. Cell. 1997, 8, 2631–2645. [Google Scholar] [CrossRef]
- van Niel, G.; Raposo, G.; Candalh, C.; Boussac, M.; Hershberg, R.; Cerf-Bensussan, N.; Heyman, M. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology 2001, 121, 337–349. [Google Scholar] [CrossRef]
- Mears, R.; Craven, R.A.; Hanrahan, S.; Totty, N.; Upton, C.; Young, S.L.; Patel, P.; Selby, P.J.; Banks, R.E. Proteomic analysis of melanoma-derived exosomes by two-dimensional polyacrylamide gel electrophoresis and mass spectrometry. Proteomics 2004, 4, 4019–4031. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, C.; Karpinski, K.; Haug, A.T.; Vester, H.; Schmitt, A.; Bauer, J.S.; van Griensven, M. Five freely circulating miRNAs and bone tissue miRNAs are associated with osteoporotic fractures. J. Bone Miner. Res. 2014, 29, 1718–1728. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, G.; Wang, Z.; Yao, Y.; Chen, R.; Pu, X.Y.; Wang, J. Circulating MicroRNA-21 is a Potential Diagnostic Biomarker in Gastric Cancer. Dis. Markers 2015, 2015, 8. [Google Scholar] [CrossRef]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.Y. Secreted microRNAs: A new form of intercellular communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef]
- Tiberio, P.; Callari, M.; Angeloni, V.; Daidone, M.G.; Appierto, V. Challenges in using circulating miRNAs as cancer biomarkers. Bio. Med. Res. Int. 2015, 2015, 731479. [Google Scholar] [CrossRef]
- Pimentel, F.; Bonilla, P.; Ravishankar, Y.G.; Contag, A.; Gopal, N.; LaCour, S.; Lee, T.; Niemz, A. Technology in microrna profiling: Circulating micrornas as noninvasive cancer biomarkers in breast cancer. J. Lab. Autom. 2015, 20, 574–588. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034.1. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify 40. genes suited for normalization, applied to bladder andcolon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar]
- Permuth-Wey, J.; Chen, D.T.; Fulp, W.J.; Yoder, S.J.; Zhang, Y.; Georgeades, C.; Husain, K.; Centeno, B.A.; Magliocco, A.M.; Coppola, D.; et al. Plasma MicroRNAs as Novel Biomarkers for Patients with Intraductal Papillary Mucinous Neoplasms of the Pancreas. Cancer Prev. Res. (Phila) 2015, 8, 826–834. [Google Scholar] [CrossRef] [PubMed]
- DeCicco, D.; Zhu, H.; Brureau, A.; Schwaber, J.S.; Vadigepalli, R. MicroRNA network changes in the brain stem underlie the development of hypertension. Physiol. Genomics 2015, 47, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Pontrelli, P.; Accetturo, M.; Gesualdo, L. miRNome analysis using real-time PCR. Methods Mol. Biol. 2014, 1186, 210–232. [Google Scholar]
- Nassirpour, R.; Mathur, S.; Gosink, M.M.; Li, Y.; Shoieb, A.M.; Wood, J.; O’Neil, S.P.; Homer, B.L.; Whiteley, L.O. Identification of tubular injury microRNA biomarkers in urine: Comparison of next-generation sequencing and qPCR-based profiling platforms. BMC Genomics 2014, 15, 485. [Google Scholar] [CrossRef]
- Moldovan, L.; Batte, K.E.; Trgovcich, J.; Wisler, J.; Marsh, C.B.; Piper, M. Methodological challenges in utilizing miRNAs as circulating biomarkers. J. Cell Mol. Med. 2014, 18, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, N.; Meryet-Figuière, M.; Guttin, A.; Issartel, J.P.; Lambert, B.; Briand, M.; Louis, M.H.; Vernon, M.; Lebailly, P.; Lecluse, Y.; et al. Towards a new standardized method for circulating miRNAs profiling in clinical studies: Interest of the exogenous normalization to improve miRNA signature accuracy. Mol. Oncol. 2016, 10, 981–992. [Google Scholar] [CrossRef]
- Benes, V.; Castoldi, M. Expression profiling of microrna using real-time quantitative pcr, how to use it and what is available. Methods 2010, 50, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Brase, J.C.; Wuttig, D.; Kuner, R.; Sültmann, H. Serum microRNAs as non-invasive biomarkers for cancer. Mol. Cancer 2010, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shen, J.; Medico, L.; Wang, D.; Ambrosone, C.B.; Liu, S. A pilot study of circulating miRNAs as potential biomarkers of early stage breast cancer. PLoS ONE 2010, 5, e13735. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Miller, N.; Lowery, A.J.; Sweeney, K.J.; Newell, J.; Kerin, M. Circulating micrornas as novel minimally invasive biomarkers for breast cancer. Ann. Surg. 2010, 251, 499–505. [Google Scholar] [CrossRef]
- Roth, C.; Rack, B.; Müller, V.; Janni, W.; Pantel, K.; Schwarzenbach, H. Circulating microRNAs as blood-based markers for patients with primary and metastatic breast cancer. Breast Cancer Res. 2010, 12, R90. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, S.; Khateri, S.; Soroush, M.R.; Shamsara, M.; Naeli, P.; Najafi, A.; Korsching, E.; Mowla, S.J. MicroRNA expression in serum samples of sulfur mustard veterans as a diagnostic gateway to improve care. PLoS ONE 2018, 13, e0194530. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Y.; Du, L.; Li, J.; Jiang, X.; Zheng, G.; Qu, A.; Wang, H.; Wang, L.; Zhang, X.; et al. Identification and validation of reference genes for the detection of serum microRNAs by reverse transcription-quantitative polymerase chain reaction in patients with bladder cancer. Mol. Med. Rep. 2015, 12, 615–622. [Google Scholar] [PubMed]
- Wang, Z.; Lu, Y.; Zhang, X.; Ren, X.; Wang, Y.; Li, Z.; Xu, C.; Han, J. Serum microRNA is a promising biomarker for osteogenesis imperfecta. Intractable Rare Dis. Res. 2012, 1, 81–85. [Google Scholar] [Green Version]
- Song, J.; Bai, Z.; Han, W.; Zhang, J.; Meng, H.; Bi, J.; Ma, X.; Han, S.; Zhang, Z. Identification of suitable reference genes for qPCR analysis of serum microRNA in gastric cancer patients. Dig. Dis. Sci. 2012, 57, 897–904. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Yuan, J.; Wu, J.; Deng, X.; Peng, J.; Wang, S.; Yang, C.; Ge, J.; Zou, Y. Evaluation of the performance of serum miRNAs as normalizers in microRNA studies focused on cardiovascular disease. J. Thorac. Dis. 2018, 10, 2599–2607. [Google Scholar] [CrossRef]
- Zalewski, K.; Misiek, M.; Kowalik, A.; Bakuła-Zalewska, E.; Kopczyński, J.; Zielińska, A.; Bidziński, M.; Radziszewski, J.; Góźdź, S.; Kowalewska, M. Normalizers for MicroRNA Quantification in Plasma of Patients with Vulvar Intraepithelial Neoplasia Lesions and Vulvar Carcinoma. Tumour Biol. 2017, 39, 1010428317717140. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, L.; Cheng, K.; Wang, X.; Ren, G.; Xie, P. Identification of suitable plasma-based reference genes for miRNAome analysis of major depressive disorder. J. Affect Disord. 2014, 163, 133–139. [Google Scholar] [CrossRef]
- Barry, S.E.; Chan, B.; Ellis, M.; Yang, Y.; Plit, M.L.; Guan, G.; Wang, X.; Britton, W.J.; Saunders, B.M. Identification of miR-93 as a suitable miR for normalizing miRNA in plasma of tuberculosis patients. J. Cell Mol. Med. 2015, 19, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wu, Y.; Huang, J.; Li, Q.; Kang, K.; Qu, J.; Li, F.; Gou, D. Identification of reference genes for circulating microRNA analysis in colorectal cancer. Sci. Rep. 2016, 6, 35611. [Google Scholar] [CrossRef]
- Li, Y.; Xiang, G.M.; Liu, L.L.; Liu, C.; Liu, F.; Jiang, D.N.; Pu, X.Y. Assessment of endogenous reference gene suitability for serum exosomal microRNA expression analysis in liver carcinoma resection studies. Mol. Med. Rep. 2015, 12, 4683–4691. [Google Scholar] [CrossRef]
- Zhu, H.T.; Dong, Q.Z.; Wang, G.; Zhou, H.J.; Ren, N.; Jia, H.L.; Ye, Q.H.; Qin, L.X. Identification of suitable reference genes for qRT-PCR analysis of circulating microRNAs in hepatitis B virus-infected patients. Mol. Biotechnol. 2012, 50, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, L.; Liu, F.; Xiang, G.; Jiang, D.; Pu, X. Identification of endogenous controls for analyzing serum exosomal miRNA in patients with hepatitis B or hepatocellular carcinoma. Dis. Markers 2015, 2015, 12. [Google Scholar] [CrossRef]
- Zheng, G.; Wang, H.; Zhang, X.; Yang, Y.; Wang, L.; Du, L.; Li, W.; Li, J.; Qu, A.; Liu, Y.; et al. Identification and validation of reference genes for qPCR detection of serum microRNAs in colorectal adenocarcinoma patients. PLoS ONE 2013, 8, e83025. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Dong, J.; Wang, L.E.; Ma, H.; Liu, J.; Zhao, Y.; Tang, J.; Chen, X.; Dai, J.; Wei, Q.; et al. Serum microRNA profiling and breast cancer risk: The use of miR-484/191 as endogenous controls. Carcinogenesis 2012, 33, 828–834. [Google Scholar] [CrossRef]
- Allen-Rhoades, W.; Kurenbekova, L.; Satterfield, L.; Parikh, N.; Fuja, D.; Shuck, R.L.; Rainusso, N.; Trucco, M.; Barkauskas, D.A.; Jo, E.; et al. Cross-species identification of a plasma microRNA signature for detection, therapeutic monitoring, and prognosis in osteosarcoma. Cancer Med. 2015, 4, 977–988. [Google Scholar] [CrossRef]
- Schlosser, K.; McIntyre, L.A.; White, R.J.; Stewart, D.J. Customized Internal Reference Controls for Improved Assessment of Circulating MicroRNAs in Disease. PLoS ONE 2015, 10, e0127443. [Google Scholar] [CrossRef]
- Chen, J.; Li, K.; Pang, Q.; Yang, C.; Zhang, H.; Wu, F.; Cao, H.; Liu, H.; Wan, Y.; Xia, W.; et al. Identification of suitable reference gene and biomarkers of serum miRNAs for osteoporosis. Sci. Rep. 2016, 6, 36347. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Z.; Liao, B.Y.; Yu, L.; Gao, X.; Lu, S.; Wang, S.; Dai, Z.; Zhang, X.; Chen, Q.; et al. Human miR-1228 as a stable endogenous control for the quantification of circulating microRNAs in cancer patients. Int. J. Cancer 2014, 135, 1187–1194. [Google Scholar] [PubMed]
- Zhang, Y.; Tang, W.; Peng, L.; Tang, J.; Yuan, Z. Identification and validation of microRNAs as endogenous controls for quantitative polymerase chain reaction in plasma for stable coronary artery disease. Cardiol. J. 2016, 23, 694–703. [Google Scholar] [CrossRef]
- Tang, G.; Shen, X.; Lv, K.; Wu, Y.; Bi, J.; Shen, Q. Different normalization strategies might cause inconsistent variation in circulating microRNAs in patients with hepatocellular carcinoma. Med. Sci. Monit. 2015, 21, 617–624. [Google Scholar]
- Tay, J.W.; James, I.; Hughes, Q.W.; Tiao, J.Y.; Baker, R.I. Identification of reference miRNAs in plasma useful for the study of oestrogen-responsive miRNAs associated with acquired Protein S deficiency in pregnancy. BMC Res. Notes 2017, 10, 312. [Google Scholar]
- Chen, Z.H.; Zhang, G.L.; Li, H.R.; Luo, J.D.; Li, Z.X.; Chen, G.M.; Yang, J. A panel of five circulating microRNAs as potential biomarkers for prostate cancer. Prostate 2012, 72, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liang, H.; Guan, D.; Wang, C.; Hu, X.; Cui, L.; Chen, S.; Zhang, C.; Zhang, J.; Zen, K.; et al. A combination of Let-7d, Let-7g and Let-7i serves as a stable reference for normalization of serum microRNAs. PLoS ONE 2013, 8, e79652. [Google Scholar]
- Sanders, I.; Holdenrieder, S.; Walgenbach-Brünagel, G.; von Ruecker, A.; Kristiansen, G.; Müller, S.C.; Ellinger, J. Evaluation of reference genes for the analysis of serum miRNA in patients with prostate cancer, bladder cancer and renal cell carcinoma. Int. J. Urol. 2012, 19, 1017–1025. [Google Scholar]
- Ghai, V.; Wang, K. Recent progress toward the use of circulating microRNAs as clinical biomarkers. Arch. Toxicol. 2016, 90, 2959–2978. [Google Scholar] [CrossRef] [PubMed]
- Armand-Labit, V.; Pradines, A. Circulating cell-free microRNAs as clinical cancer biomarkers. Biomol. Concepts 2017, 8, 61–81. [Google Scholar] [CrossRef] [PubMed]
- Shajari, E.; Mollasalehi, H. Ribonucleic-acid-biomarker candidates for early-phase group detection of common cancers. Genomics 2018. [Google Scholar] [CrossRef]
- Masè, M.; Grasso, M.; Avogaro, L.; D’Amato, E.; Tessarolo, F.; Graffigna, A.; Denti, M.A.; Ravelli, F. Selection of reference genes is critical for miRNA expression analysis in human cardiac tissue. A focus on atrial fibrillation. Sci. Rep. 2017, 7, 41127. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; da Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Pathologies | Biological Fluids | Population* | Platforms for miRNA Detection | RG Stability Analysis Methods | RGs | References |
|---|---|---|---|---|---|---|
| Breast Cancer | Plasma | Women with early stage breast (20) | microarray, qPCR | miR-16 | [61] | |
| Breast Cancer | Whole blood | Breast cancer (148) | Real-Time qPCR | geNorm | miR-16 | [62] |
| Breast Cancer | Serum | Consecutive breast cancer (83) | qPCR | miR-16 | [63] | |
| SMV | Serum | SMV (28) | qPCR | geNorm, NormFinder, Bestkeeper, comparative delta Ct | miR-423-3p, miR-103, miR-16 | [64] |
| Bladder Cancer | Serum | NMIBC (40), MIBC (40), and Bladder cancer (46) | MiSeq sequencing, qPCR | geNorm, NormFinder | miR-193a-5p, miR-16-5p | [65] |
| Osteogenesis Imperfecta | Serum | osteogenesis imperfecta (22) | qPCR | geNorm, NormFinder, Bestkeeper, comparative delta Ct | snRNAU6/miR-92a, miR-16, let-7a | [66] |
| Gastric Cancer | Serum | Gastric cancer (20) | qPCR | geNorm, NormFinder, Bestkeeper, comparative delta Ct | miR-16, miR-93 | [67] |
| Cardiovascular Disease | Serum | Cardiovascular disease (35) | qPCR | NormFinder, Bestkeeper, comparative delta Ct | let-7i, miR-16 | [68] |
| Vulvar Carcinoma | Plasma | Vulvar intraepithelial neoplasia lesions (17), and vulvar squamous cell carcinoma (27) | qPCR | geNorm, NormFinder, BestKeeper, comparative delta Ct | miR-93-5p, miR-425-5p | [69] |
| MDD | Plasma | MDD (32) | Microarray, qPCR | geNorm, NormFinder, BestKeeper, comparative delta Ct | miR-93-5p, miR-101-3p | [70] |
| Tuberculosis | Plasma | Patients with tuberculosis (24), and patients LTBI (24) | qPCR | geNorm, NormFinder | miR-93 | [71] |
| Colorectal Cancer | Serum | Colorectal cancer (96) | Microarray, qPCR | geNorm, NormFinder, BestKeeper | miR-93-5p, miR-25-3p, miR-106b-5p | [72] |
| Liver Carcinoma | Serum Exosomal | Pre-operative and post-operative patients with HCC (53) | qPCR | geNorm, NormFinder, BestKeeper, comparative delta Ct | miR-221, miR-26a, let-7a | [73] |
| HBV | Serum | HBV (52) | TaqMan low density arrays, qPCR | geNorm, NormFinder | miR-26a, miR-221, miR-22* | [74] |
| HBV and HCC | Serum Exosomal | HCC (50), and HBV (50) | Real-Time qPCR | geNorm, NormFinder | miR-221, miR-191, let-7a, miR-181a, miR-26a | [75] |
| Colorectal Adenocarcinoma | Serum | Colorectal adenocarcinoma (203) | Miseq sequencing, qPCR | geNorm, NormFinder | miR-191-5p, U6 | [76] |
| Breast Cancer | Serum | Breast cancer (225), lung cancer (65), cervical cancer (30), gastric cancer (45), HCC (35), esophageal cancer (5), colon cancer (5), rectal cancer (5), pancreatic cancer (5), and oral cancer (5) | Solexa sequencing, TaqMan low density array, qPCR | NormFinder | miR-191, miR-484 | [77] |
| Osteosarcoma | Plasma | Engineered mouse models of osteosarcoma, osteosarcoma human patients (60) | qPCR | geNorm | miR-320a, miR-15a | [78] |
| PAH or Septic Shock | Plasma | PAH (18), and patients with septic shock (4) | qPCR | NormFinder | miR-142-3p, miR-320a | [79] |
| Osteoporosis | Serum | Osteoporosis-induced animal model; normal BMD (19), osteopenic (7), and osteoporosis woman (10) | Microarray, qPCR | geNorm, NormFinder | miR-25-3p | [80] |
| Different Cancer Types | Plasma | HCC (88), colorectal cancer (62), lung cancer (103), esophageal cancer (23), gastric cancer (21), renal cancer (24), CaP (20), and breast cancer (21) | Microarray, qPCR | geNorm, NormFinder | miR-1228 | [81] |
| CAD | Plasma | CAD (119) | Microarray, qPCR | NormFinder, BestKeeper | miR-6090, miR-4516 | [82] |
| HCC | Plasma | HCC (30), gastric carcinoma (30), hepatic cirrhosis (20), and HBV (20) | qPCR | geNorm, NormFinder, BestKeeper, comparative delta Ct | miR-106a, miR-21 | [83] |
| Protein S Deficiency in Pregnancy | Plasma | Non-pregnant females not taking oral contraceptives (14), non-pregnant females currently taking oral contraceptives (14), and pregnant females (14) | nanoString nCounter, qPCR, Digital Droplet PCR | NormFinder, Bestkeeper | miR-188-5p, miR-222-3p | [84] |
| CaP | Plasma | CaP (105), and benign prostatic hyperplasia (61) | qPCR | comparative delta Ct | U6 | [85] |
| Different Pathology Types | Serum | Non-Small Cell Lung Cancer (87), pancreatic cancer (88), gastric cancer (87), esophageal cancer (46), colorectal cancer (30), patients with HCC (30), breast cancer (58), ovarian cancer (26), cervical cancer (40), nephritis (101), colitis (73), pancreatitis (18), pneumonia (8), and type 2 diabetes (320) | Illumina technology, qPCR | geNorm, NormFinder | let-7d/g/i | [86] |
| CaP, Bladder Cancer and RCC | Serum | CaP (24), NMIBC (12), MIBC (12), and RCC (24) | qPCR | geNorm, NormFinder, comparative delta Ct | SNORD43, RNU1-4 | [87] |
| Pathology | RGs | References |
|---|---|---|
| Breast Cancer | miR-16 | [61] |
| miR-16 | [62] | |
| miR-16 | [63] | |
| miR-191, miR-484 | [77] | |
| miR-1228 | [81] | |
| let-7d/g/i | [86] | |
| Urogenital Malignances (Bladder, Prostate, Renal) | miR-16-5p, miR-193a-5p | [65] |
| miR-1228 | [81] | |
| U6 | [85] | |
| SNORD43, RNU1-4 | [87] | |
| Bone Diseases (Osteogenesis imperfecta, Osteosarcoma, Osteoporosis) | miR-16, U6, miR-92a, let-7a | [66] |
| miR-320a, miR-15a | [78] | |
| miR-25-3p | [80] | |
| Gastrointestinal Disorders (Esophagus Cancer, Colorectal Cancer, Gastric, Cancer, Pancreatic Disease) | miR-93, miR-16 | [67] |
| miR-93-5p, miR-25-3p, miR-106b-5p | [72] | |
| miR-191-5p, U6 | [76] | |
| miR-1228 | [81] | |
| let-7d/g/i | [86] | |
| Cardiovascular Pathologies(Hypertension, Heart Failure, CAD) | miR-16, let-7i | [68] |
| miR-6090, miR-4516 | [82] | |
| Female Reproductive System Cancers(Vulvar, Ovarian, Cervical) | miR-93-5p, miR-425-5p | [69] |
| let-7d/g/i | [86] | |
| Lung Diseases(Tuberculosis, Pulmonary cancers) | miR-93 | [71] |
| miR-320a, miR-142-3p | [79] | |
| miR-1228 | [81] | |
| Liver Disorders | miR-221, let-7a, miR-26a | [73] |
| miR-221, miR-22*, miR-26a | [74] | |
| miR-221, let-7a, miR-191, miR-26a, miR-181a | [75] | |
| miR-1228 | [81] | |
| miR-106a, miR-21 | [83] | |
| let-7d/g/i | [86] | |
| MDD | miR-93-5p, miR-101-3p | [70] |
| SMV | miR-423-3p, miR-103, miR-16 | [64] |
| Protein S deficiency in pregnancy | miR-188-5p and miR-222-3p | [84] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, S.; Ciuffi, S.; Brandi, M.L. Human Circulating miRNAs Real-time qRT-PCR-based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization. Int. J. Mol. Sci. 2019, 20, 4353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184353
Donati S, Ciuffi S, Brandi ML. Human Circulating miRNAs Real-time qRT-PCR-based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization. International Journal of Molecular Sciences. 2019; 20(18):4353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184353
Chicago/Turabian StyleDonati, Simone, Simone Ciuffi, and Maria L. Brandi. 2019. "Human Circulating miRNAs Real-time qRT-PCR-based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization" International Journal of Molecular Sciences 20, no. 18: 4353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184353