Effect of Tff3 Deficiency and ER Stress in the Liver

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
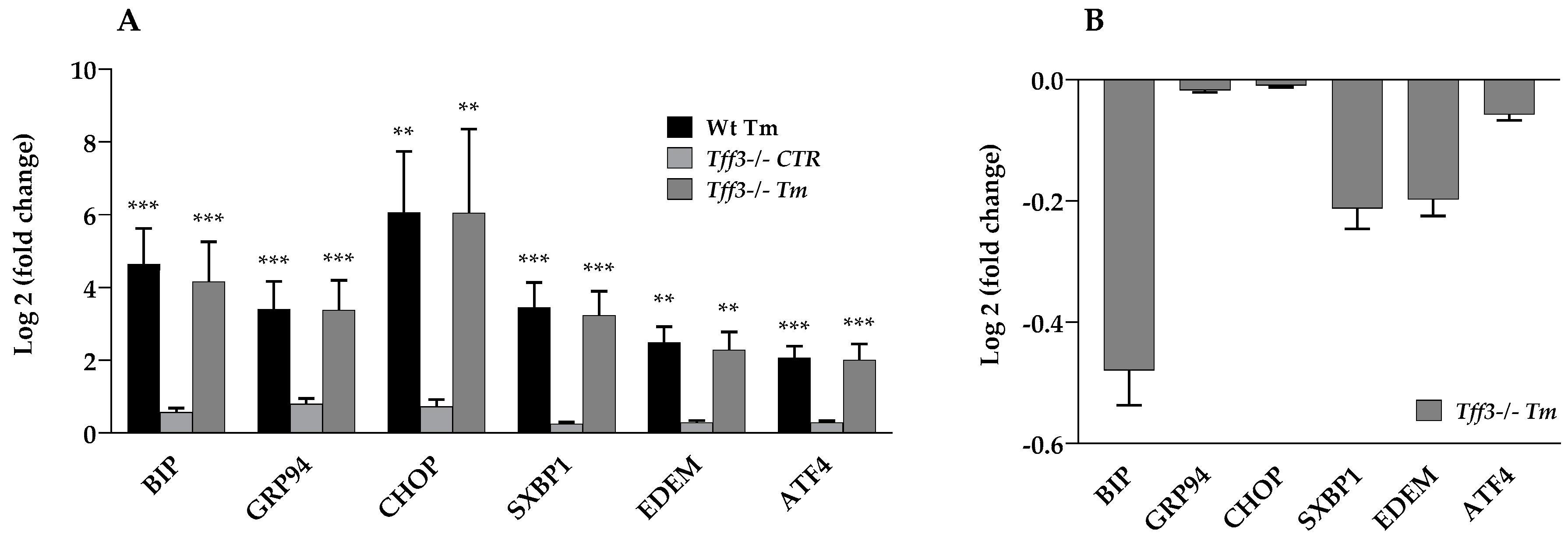
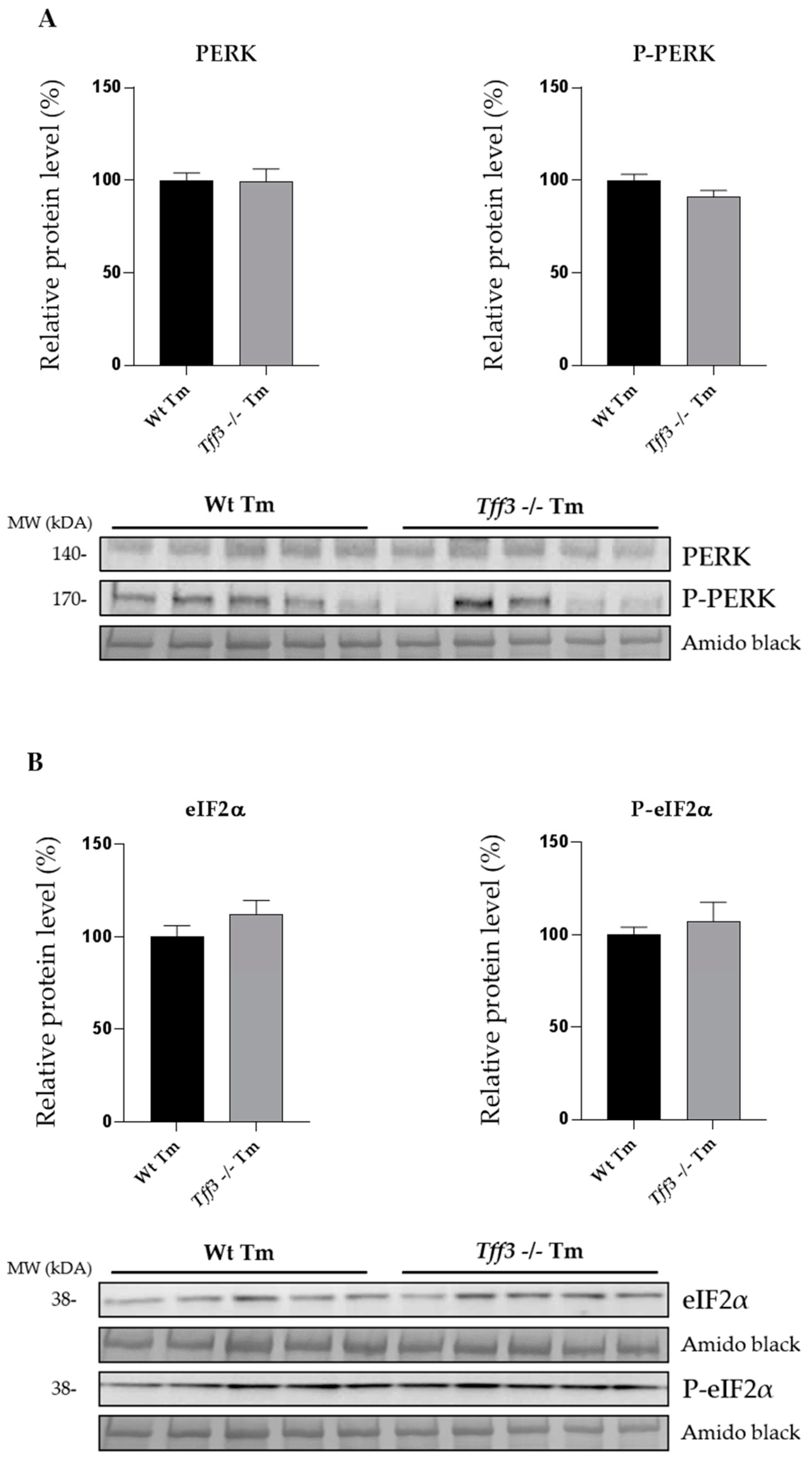
2.1. Expression of ER Stress Markers
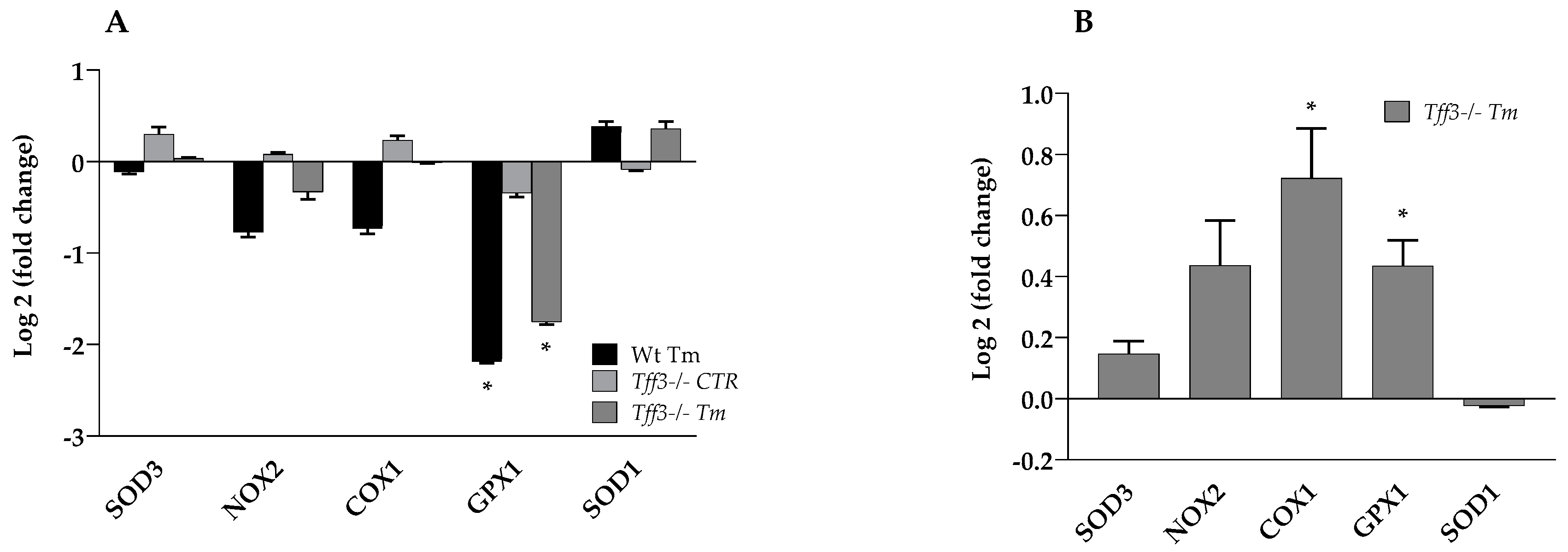
2.2. Expression of Oxidative Stress Marker Genes
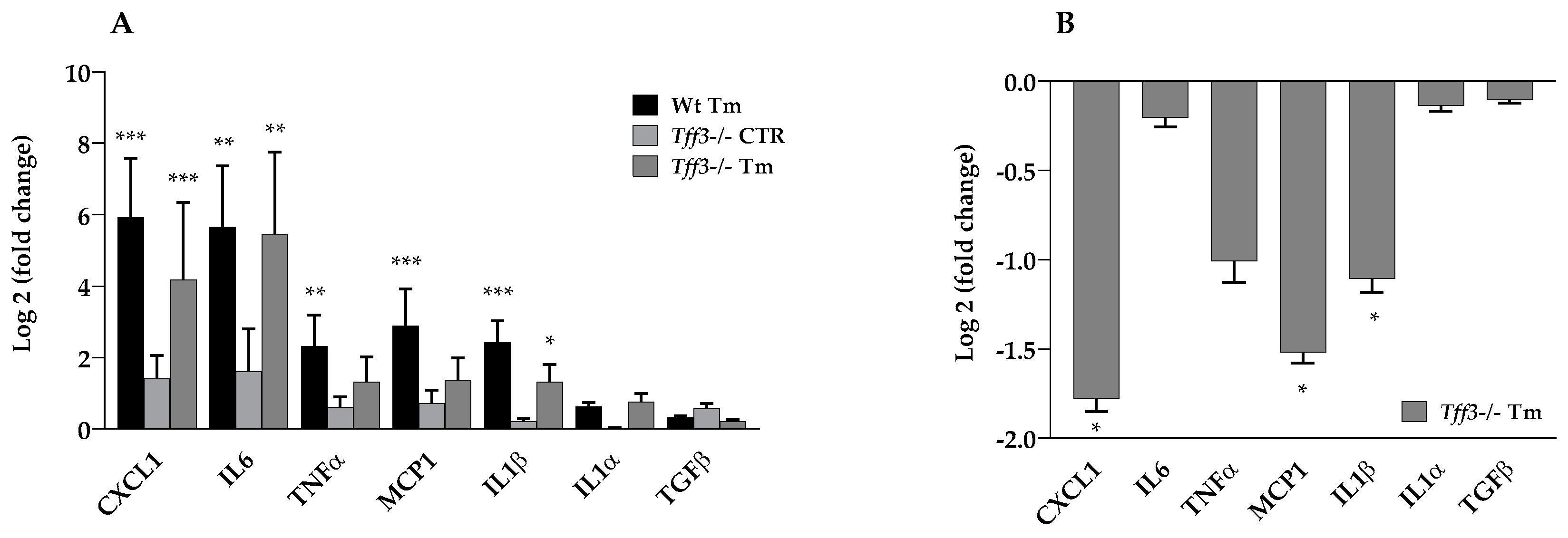
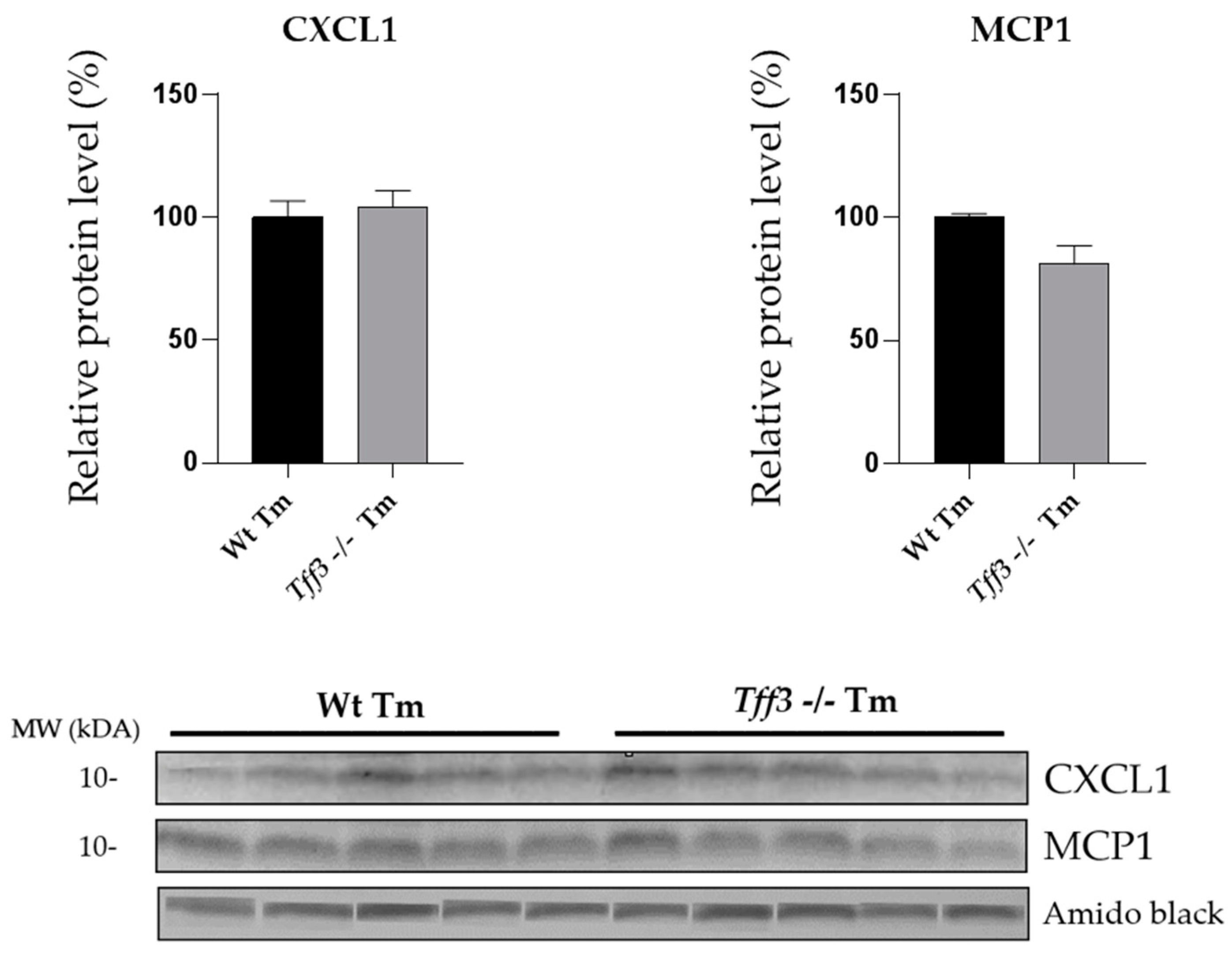
2.3. Tff3 Deficiency Reduces Cytokine Expression in Acute ERS
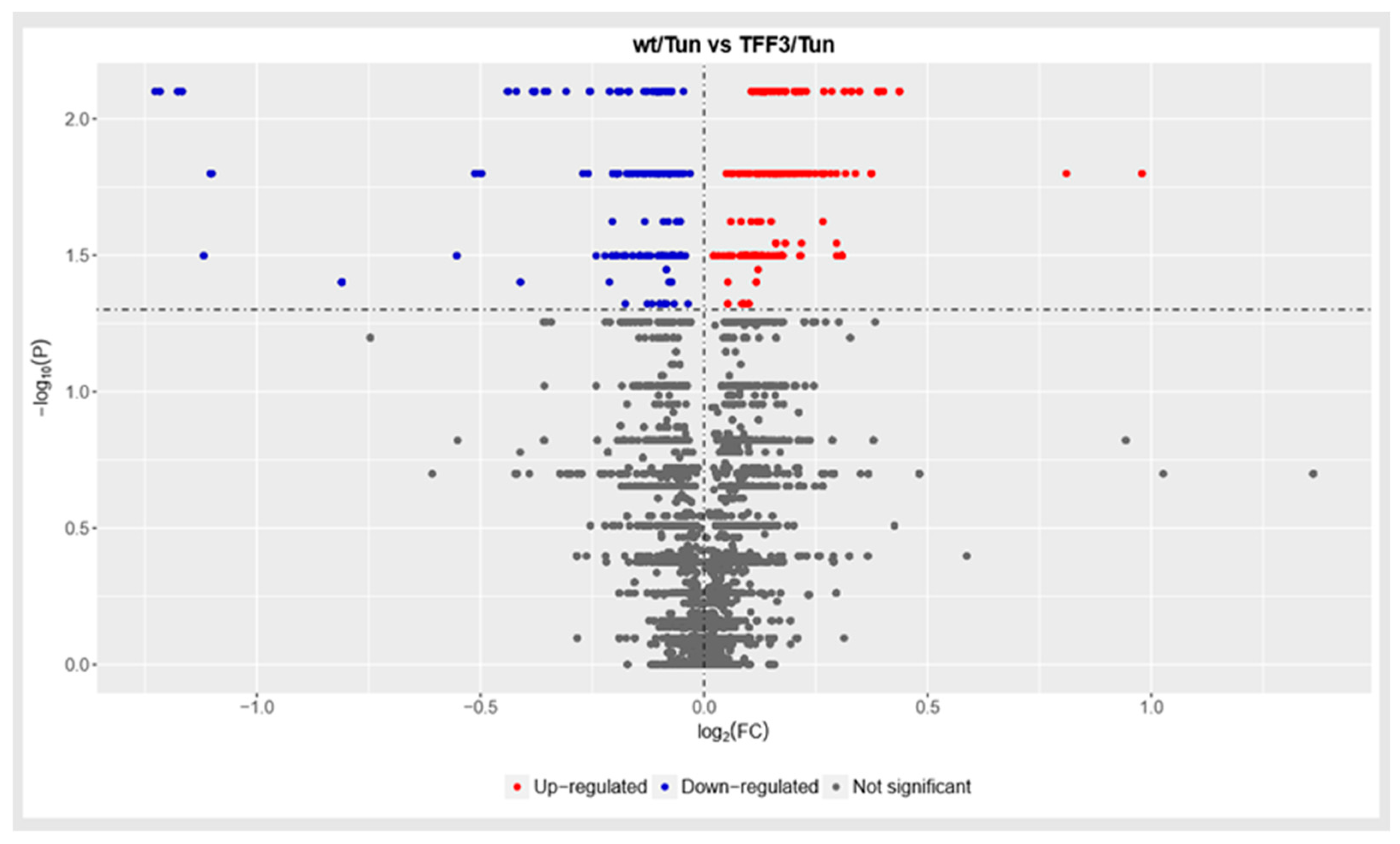
2.4. Shotgun Proteomic Analysis of Liver Proteome ERS Provoked WT and Tff3 Deficient Mice
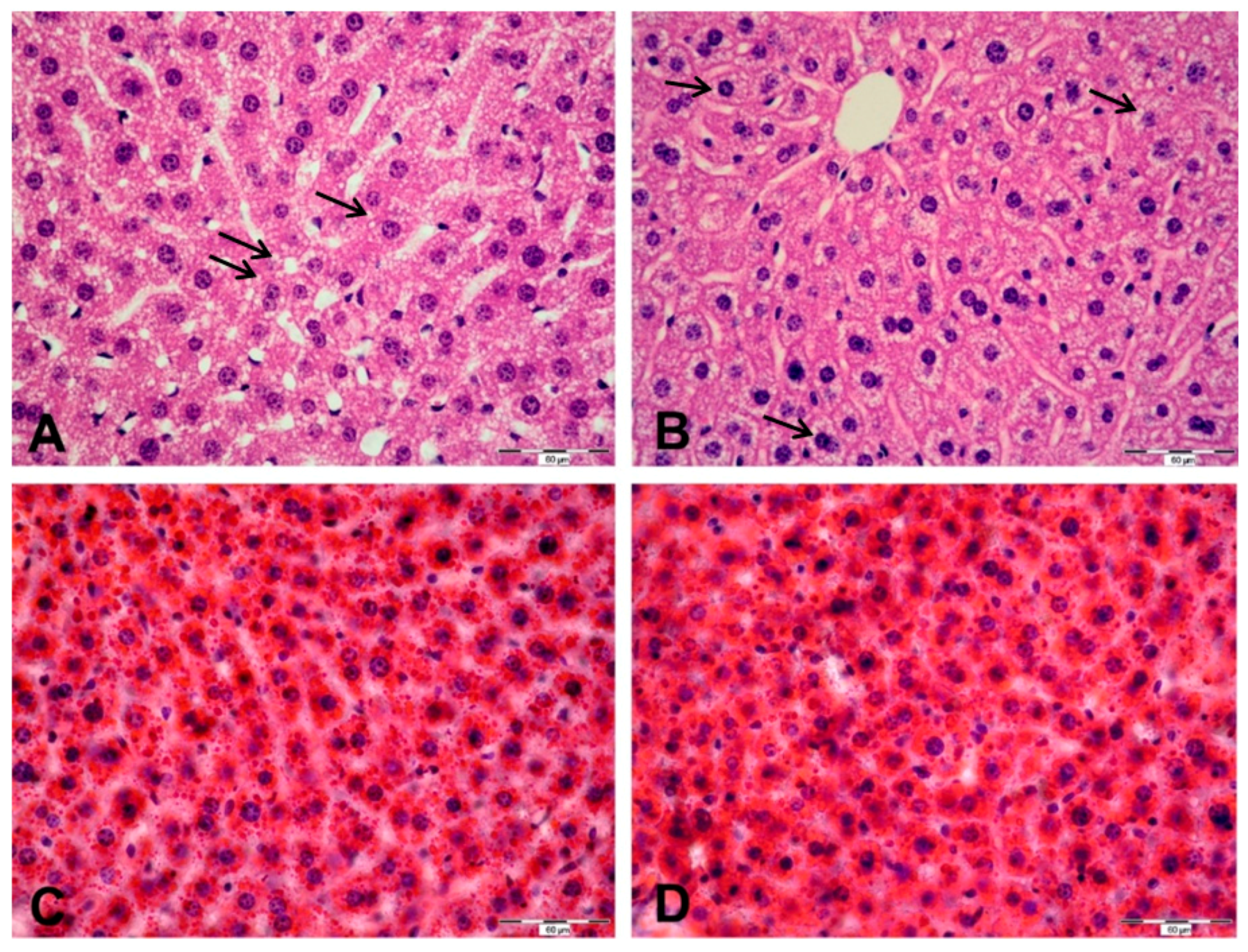
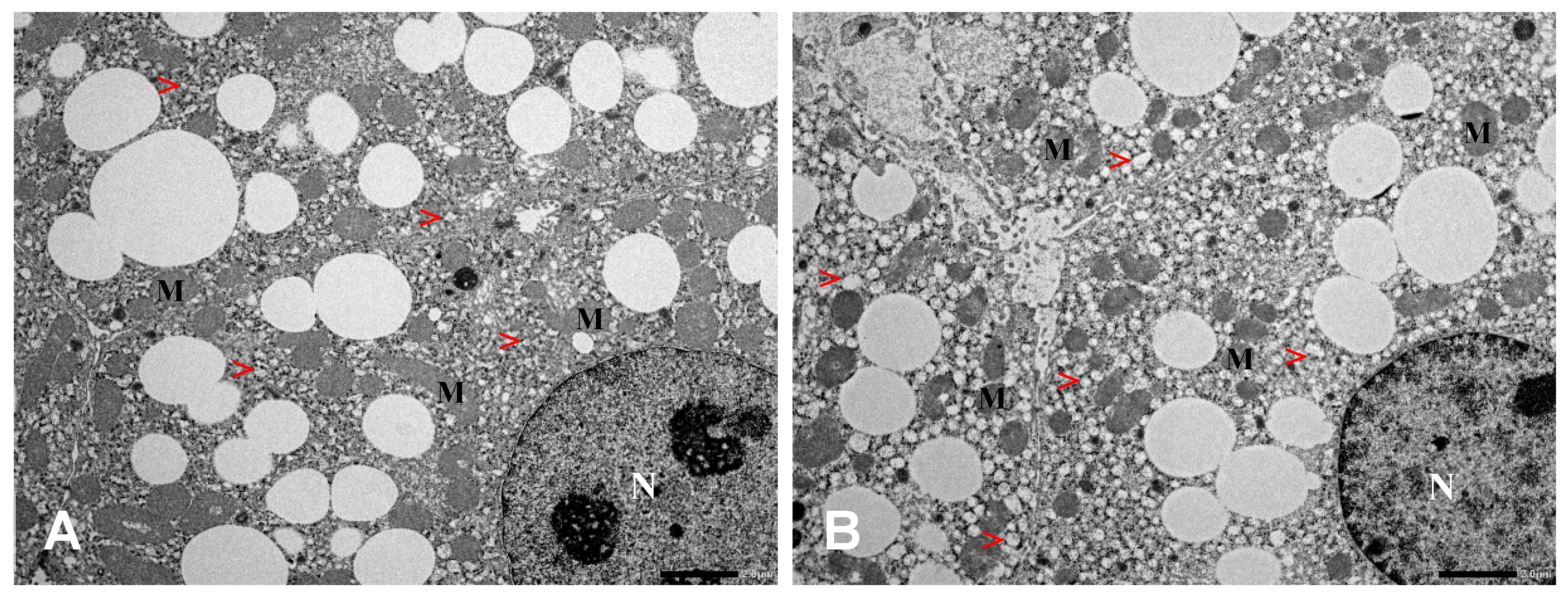
2.5. Histological and Ultrastructural Changes
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Inducing ER Stress
4.3. Quantitative PCR
4.4. Western Blot
4.5. Proteomic Analysis
4.5.1. Protein Extraction and Sample Preparation
4.5.2. LC-MS/MS Analysis
4.5.3. Data Analysis
4.6. Histology
4.7. Ultrastructure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACTB | actin beta |
| ATF4 | activating transcription factor 4 |
| BIP | endoplasmic reticulum chaperone bip |
| B2M | beta-2-microglobulin |
| CHOP | DNA damage-inducible transcript 3 |
| COX1 | cyclooxygenase I |
| CXCL1 | C-X-C motif chemokine ligand 1 |
| eIF2α | eukaryotic translation initiation factor 2α |
| EDEM1 | ER degradation enhancing alpha-mannosidase like protein 1 |
| ER | endoplasmic reticulum |
| ERS | endoplasmic reticulum stress |
| GSTP1 | glutathione S-transferase pi 1 |
| GSTP2 | glutathione S-transferase pi 2 |
| GPX1 | glutathione peroxidase 1 |
| GRP94 | heat shock protein 90 beta family member 1 |
| IL1α | interleukin 1 alpha |
| IL1β | interleukin 1 beta |
| IL6 | interleukin 6 |
| MCP1 | monocyte chemoattractant protein-1 |
| MT1 | metallothionein-1 |
| MT2 | metallothionein-1 |
| NOX2 | NADPH oxidase 2 |
| PERK | protein kinase R (PKR)-like endoplasmic reticulum kinase |
| SOD1 | superoxide dismutase 1 |
| SOD3 | superoxide dismutase 3 |
| sXBP1 | spliced X-box binding protein 1 |
| Tffs | trefoil factor family proteins |
| TNFα | tumor necrosis factor alpha |
| TGFα | tumor growth factor alpha |
| Tm | tunicamycin |
| UPR | unfolded protein response |
| Wt | wild type |
References
- Wang, S.; Kaufman, R.J. The impact of the unfolded protein response on human disease. J. Cell Biol. 2012, 197, 857–867. [Google Scholar] [CrossRef] [Green Version]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Luís, A.; McCarthy, N.; Montibeller, L.; More, S.; et al. Endoplasmic reticulum stress signalling—from basic mechanisms to clinical applications. FEBS J. 2019, 286, 241–278. [Google Scholar] [CrossRef]
- Fu, S.; Watkins, S.M.; Hotamisligil, G.S. The role of endoplasmic reticulum in hepatic lipid homeostasis and stress signaling. Cell Metab. 2012, 15, 623–634. [Google Scholar] [CrossRef]
- Torres, L.F.; Karam, S.M.; Wendling, C.; Chenard, M.P.; Kershenobich, D.; Tomasetto, C.; Rio, M.C. Trefoil Factor 1 (TFF1/pS2) Deficiency Activates the Unfolded Protein Response. Mol. Med. 2002, 8, 273–282. [Google Scholar] [CrossRef]
- Baus-Loncar, M.; Schmid, J.; Lalani, E.N.; Rosewell, I.; Goodlad, R.A.; Stamp, G.W.H.; Blin, N.; Kayademir, T. Trefoil Factor 2 (Tff2) Deficiency in Murine Digestive Tract Influences the Immune System. Cell. Physiol. Biochem. 2005, 16, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Lubka, M.; Shah, A.A.; Blin, N.; Baus-Lončar, M. The intestinal trefoil factor (Tff3), also expressed in the inner ear, interacts with peptides contributing to apoptosis. J. Appl. Genet. 2009, 50, 167–171. [Google Scholar] [CrossRef]
- Desmots, F.; Russell, H.R.; Michel, D.; McKinnon, P.J. Scythe regulates apoptosis-inducing factor stability during endoplasmic reticulum stress-induced apoptosis. J. Biol. Chem. 2008, 283, 3264–3271. [Google Scholar] [CrossRef]
- Ren, C.; Dokter-Fokkens, J.; Figueroa Lozano, S.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Lactic Acid Bacteria May Impact Intestinal Barrier Function by Modulating Goblet Cells. Mol. Nutr. Food Res. 2018, 62, 1–14. [Google Scholar] [CrossRef]
- Asada, R.; Saito, A.; Kawasaki, N.; Kanemoto, S.; Iwamoto, H.; Oki, M.; Miyagi, H.; Izumi, S.; Imaizumi, K. The endoplasmic reticulum stress transducer OASIS is involved in the terminal differentiation of goblet cells in the large intestine. J. Biol. Chem. 2012, 287, 8144–8153. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef]
- Loncar, M.B.; Al-azzeh, E.; Sommer, P.S.M.; Marinovic, M.; Schmehl, K.; Kruschewski, M.; Blin, N.; Stohwasser, R.; Gött, P.; Kayademir, T. Tumour necrosis factor α and nuclear factor κB inhibit transcription of human TFF3 encoding a gastrointestinal healing peptide. Gut 2003, 52, 1297–1303. [Google Scholar] [CrossRef]
- Bernstein, H.G.; Dobrowolny, H.; Trübner, K.; Steiner, J.; Bogerts, B.; Hoffmann, W. Differential regional and cellular distribution of TFF3 peptide in the human brain. Amino Acids 2015, 47, 1053–1063. [Google Scholar] [CrossRef]
- Liu, S.Q.; Tefft, B.J.; Roberts, D.T.; Zhang, L.Q.; Ren, Y.; Li, Y.C.; Huang, Y.; Zhang, D.; Phillips, H.R.; Wu, Y.H. Cardioprotective proteins upregulated in the liver in response to experimental myocardial ischemia. Am. J. Physiol. Circ. Physiol. 2012, 303, H1446–H1458. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.Q.; Roberts, D.; Zhang, B.; Ren, Y.; Zhang, L.Q.; Wu, Y.H. Trefoil Factor 3 as an Endocrine Neuroprotective Factor from the Liver in Experimental Cerebral Ischemia/Reperfusion Injury. PLoS ONE 2013, 8, 1–15. [Google Scholar] [CrossRef]
- Xue, Y.; Shen, L.; Cui, Y.; Zhang, H.; Chen, Q.; Cui, A.; Fang, F.; Chang, Y. Tff3, as a Novel Peptide, Regulates Hepatic Glucose Metabolism. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef]
- Ge, H.; Gardner, J.; Wu, X.; Rulifson, I.; Wang, J.; Xiong, Y.; Ye, J.; Belouski, E.; Cao, P.; Tang, J.; et al. Trefoil factor 3 (TFF3) is regulated by food intake, improves glucose tolerance and induces mucinous metaplasia. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Brown, A.C.; Olver, W.I.; Donnelly, C.J.; May, M.E.; Naggert, J.K.; Shaffer, D.J.; Roopenian, D.C. Searching QTL by gene expression: Analysis of diabesity. BMC Genet. 2005, 6, 1–9. [Google Scholar] [CrossRef]
- Guillén, N.; Navarro, M.A.; Arnal, C.; Noone, E.; Arbonés-Mainar, J.M.; Acín, S.; Surra, J.C.; Muniesa, P.; Roche, H.M.; Osada, J. Microarray analysis of hepatic gene expression identifies new genes involved in steatotic liver. Physiol. Genom. 2009, 37, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Mashimo, H.; Wu, D.C.; Podolsky, D.K.; Fishman, M.C. Impaired Defense of Intestinal Mucosa in Mice Lacking Intestinal Trefoil Factor. Science 1996, 274, 262–265. [Google Scholar] [CrossRef]
- Sigmund, C.D. Viewpoint: Are Studies in Genetically Altered Mice Out of Control? Curt D. Sigmund Arterioscler. Arterioscler. Thromb. 2000, 1425–1429. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.H.; Ryan, T.E.; Smith, C.D.; Gilliam, L.A.A.; Lin, C.T.; Reese, L.R.; Torres, M.J.; Neufer, P.D. A direct comparison of metabolic responses to high-fat diet in c57bl/6j and c57bl/6nj mice. Diabetes 2016, 65, 3249–3261. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Cardozo, A.K.; Cnop, M. The Role for Endoplasmic Reticulum Stress in Diabetes Mellitus. Endocr. Rev. 2008, 29, 42–61. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Xu, C.F.; Yu, C.H.; Chen, W.X.; Li, Y.M. Role of endoplasmic reticulum stress in the pathogenesis of nonalcoholic fatty liver disease. World J. Gastroenterol. 2014. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’H, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. (Berl.) 2008. [Google Scholar] [CrossRef]
- Abdullahi, A.; Stanojcic, M.; Parousis, A.; Patsouris, D.; Jeschke, M.G. Modeling Acute ER Stress in Vivo and in Vitro. Shock 2017, 47, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Oslowski, C.M.; Urano, F. Measuring ER Stress and the Unfolded Protein Response Using Mammalian Tissue Culture System. Methods Enzymol. 2011, 490, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Luhr, M.; Torgersen, M.L.; Szalai, P.; Hashim, A.; Brech, A.; Staerk, J.; Engedal, N. The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress. J. Biol. Chem. 2019. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Y.; Tang, J.; Jiang, J.; Chen, Z. New insights into the roles of CHOP-induced apoptosis in ER stress. Acta Biochim. Biophys Sin 2014, 46, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Cormier, J.H.; Tamura, T.; Sunryd, J.C.; Hebert, D.N. EDEM1 recognition and delivery of misfolded proteins to the SEL1L-containing ERAD complex. Mol. Cell 2009, 34, 627–633. [Google Scholar] [CrossRef]
- Eletto, D.; Dersh, D.; Argon, Y. GRP94 in ER quality control and stress responses. Semin. Cell Dev. Biol. 2010, 21, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Balcer-Kubiczek, E.K.; Harrison, G.H.; Xu, J.F.; Gutierrez, P.L. Coordinate late expression of trefoil peptide genes (pS2/TFF1 and ITF/TFF3) in human breast, colon and gastric tumor cells exposed to X-rays. Mol. Cancer Ther. 2002, 1, 405–415. [Google Scholar]
- Cohen, G.; Hochstein, P. Glutathione Peroxidase: The primary agent for the elimination of hydrogen peroxide in erythrocytes. Biochemistry 1963, 6, 1420–1428. [Google Scholar] [CrossRef]
- Bujak, M.; Bujak, I.T.; Sobočanec, S.; Mihalj, M.; Novak, S.; Cosić, A.; Levak, M.T.; Kopačin, V.; Mihaljević, B.; Balog, T.; et al. Trefoil factor 3 deficiency affects liver lipid metabolism. Cell. Physiol. Biochem. 2018, 47, 827–841. [Google Scholar] [CrossRef]
- Perlemuter, G.; Davit-Spraul, A.; Cosson, C.; Conti, M.; Bigorgne, A.; Paradis, V.; Corre, M.P.; Prat, L.; Kuoch, V.; Basdevant, A.; et al. Increase in liver antioxidant enzyme activities in non-alcoholic fatty liver disease. Liver Int. 2005, 25, 946–953. [Google Scholar] [CrossRef]
- Ucar, F.; Sezer, S.; Erdogan, S.; Akyol, S.; Armutcu, F.; Akyol, O. The relationship between oxidative stress and nonalcoholic fatty liver disease: Its effects on the development of nonalcoholic steatohepatitis. Redox Rep. 2013, 18, 127–133. [Google Scholar] [CrossRef]
- Marnett, L.J.; Rowlinson, S.W.; Goodwin, D.C.; Kalgutkar, A.S.; Lanzo, C.A. Arachidonic acid oxygenation by COX-1 and COX-2. Mechanisms of catalysis and inhibition. J. Biol. Chem. 1999, 274, 22903–22906. [Google Scholar] [CrossRef]
- Gores, G.J.; Flarsheim, C.E.; Dawson, T.L.; Nieminen, A.L.; Herman, B.; Lemasters, J.J. Swelling, reductive stress, and cell death during chemical hypoxia in hepatocytes. Am. J. Physiol. Physiol. 1989, 257, 347–354. [Google Scholar] [CrossRef]
- Xiao, J.; Liong, E.C.; Huang, H.; On Tse, W.; Lau, K.S.; Pan, J.; Nanji, A.A.; Fung, M.L.; Xing, F.; Tipoe, G.L. Cyclooxygenase-1 serves a vital hepato-protective function in chemically induced acute liver injury. Toxicol. Sci. 2014, 143, 430–440. [Google Scholar] [CrossRef]
- Jackson, L.M.; Wu, K.C.; Mahida, Y.R.; Jenkins, D.; Hawkey, C.J. Cyclooxygenase (COX) 1 and 2 in normal, inflamed, and ulcerated human gastric mucosa. Gut 2000, 47, 762–770. [Google Scholar] [CrossRef]
- Rodrigues, S.; Nguyen, Q.D.; Faivre, S.; Bruyneel, E.; Thim, L.; Westley, B.; May, F.; Flatau, G.; Mareel, M.; Gespach, C.; et al. Activation of cellular invasion by trefoil peptides and src is mediated by cyclooxygenase- and thromboxane A2 receptor-dependent signaling pathways. FASEB J. 2002, 15, 1517–1528. [Google Scholar] [CrossRef]
- Xue, L.; Aihara, E.; Podolsky, D.K.; Wang, T.C.; Montrose, M.H. In vivo action of trefoil factor 2 (TFF2) to speed gastric repair is independent of cyclooxygenase. Gut 2010, 59, 1184–1191. [Google Scholar] [CrossRef]
- Cook, G.A.; Familari, M.; Thim, L.; Giraud, A.S. The trefoil peptides TFF2 and TFF3 are expressed in rat lymphoid tissues and participate in the immune response. FEBS Lett. 1999, 456, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Fu, T.; Znalesniak, E.B.; Kalinski, T.; Möhle, L.; Biswas, A.; Salm, F.; Dunay, I.R.; Hoffmann, W. TFF Peptides Play a Role in the Immune Response Following Oral Infection of Mice with Toxoplasma Gondii. Eur. J. Microbiol. Immunol. (Bp). 2015, 5, 221–231. [Google Scholar] [CrossRef]
- Kolattukudy, P.E.; Niu, J. Inflammation, endoplasmic reticulum stress, autophagy, and the monocyte chemoattractant protein-1/CCR2 pathway. Circ. Res. 2012, 110, 174–189. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, J.; Fan, H.N.; Zhu, J.S. Function and therapeutic advances of chemokine and its receptor in nonalcoholic fatty liver disease. Therap. Adv. Gastroenterol. 2018, 11, 1–13. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Invest. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Mandrekar, P.; Ambade, A.; Lim, A.; Szabo, G.; Catalano, D. An essential role for monocyte chemoattractant protein-1 in alcoholic liver injury: Regulation of proinflammatory cytokines and hepatic steatosis in mice. Hepatology 2011, 54, 2185–2197. [Google Scholar] [CrossRef] [Green Version]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Gao, B.; Tsukamoto, H. Inflammation in Alcoholic and Nonalcoholic Fatty Liver Disease: Friend or Foe? Gastroenterology 2016, 150, 1704–1709. [Google Scholar] [CrossRef] [Green Version]
- Isoda, K.; Sawada, S.; Ayaori, M.; Matsuki, T.; Horai, R.; Kagata, Y.; Miyazaki, K.; Kusuhara, M.; Okazaki, M.; Matsubara, O.; et al. Deficiency of interleukin-1 receptor antagonist deteriorates fatty liver and cholesterol metabolism in hypercholesterolemic mice. J. Biol. Chem. 2005, 280, 7002–7009. [Google Scholar] [CrossRef]
- Miura, K.; Kodama, Y.; Inokuchi, S.; Schnabl, B.; Aoyama, T.; Ohnishi, H.; Olefsky, J.M.; Brenner, D.A.; Seki, E. Toll-like receptor 9 promotes steatohepatitis by induction of interleukin-1beta in mice. Gastroenterology 2010, 139, 323–334. [Google Scholar] [CrossRef]
- Kamari, Y.; Shaish, A.; Vax, E.; Shemesh, S.; Kandel-Kfir, M.; Arbel, Y.; Olteanu, S.; Barshack, I.; Dotan, S.; Voronov, E.; et al. Lack of interleukin-1α or interleukin-1β inhibits transformation of steatosis to steatohepatitis and liver fibrosis in hypercholesterolemic mice. J. Hepatol. 2011, 55, 1086–1094. [Google Scholar] [CrossRef]
- Cheng, Z.; Teo, G.; Krueger, S.; Rock, T.M.; Koh, H.W.; Choi, H.; Vogel, C. Differential dynamics of the mammalian mRNA and protein expression response to misfolding stress. Mol. Syst. Biol. 2016, 12, 855. [Google Scholar] [CrossRef]
- Nejdl, L.; Gumulec, J.; Ruttkay-Nedecky, B.; Stiborova, M.; Zitka, O.; Adam, V.; Kizek, R.; Masarik, M.; Eckschlager, T. The role of metallothionein in oxidative Stress. Int. J. Mol. Sci. 2013, 14, 6044–6066. [Google Scholar] [CrossRef]
- Thirumoorthy, N.; Shyam Sunder, A.; Manisenthil Kumar, K.T.; Senthil kumar, M.; Ganesh, G.N.K.; Chatterjee, M. A review of metallothionein isoforms and their role in pathophysiology. World J. Surg. Oncol. 2011, 9, 1–7. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakajima, K. Significance of metallothionein expression in liver disease. Curr. Pharm. Biotechnol. 2013, 14, 420–426. [Google Scholar] [CrossRef]
- Waller-Evans, H.; Hue, C.; Fearnside, J.; Rothwell, A.R.; Lockstone, H.E.; Caldérari, S.; Wilder, S.P.; Cazier, J.B.; Scott, J.; Gauguier, D. Nutrigenomics of high fat diet induced obesity in mice suggests relationships between susceptibility to fatty liver disease and the proteasome. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef]
- Kondoh, M.; Tsukada, M.; Kuronaga, M.; Higashimoto, M.; Takiguchi, M.; Himeno, S.; Watanabe, Y.; Sato, M. Induction of hepatic metallothionein synthesis by endoplasmic reticulum stress in mice. Toxicol. Lett. 2004, 148, 133–139. [Google Scholar] [CrossRef]
- Arriaga, J.M.; Bravo, A.I.; Mordoh, J.; Bianchini, M. Metallothionein 1G promotes the differentiation of HT-29 human colorectal cancer cells. Oncol. Rep. 2017, 37, 2633–2651. [Google Scholar] [CrossRef] [Green Version]
- Baltruskeviciene, E.; Kazbariene, B.; Badaras, R.; Bagdonaitė, L.; Krikštaponienė, A.; Zdanavičius, L.; Aleknavicius, E.; Didziapetrienė, J. Glutathione and glutathione S-transferase levels in patients with liver metastases of colorectal cancer and other hepatic disorders. Turk. J. Gastroenterol. 2016, 27, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.W.; Zhang, J.; Ancrum, T.; Manevich, Y.; Townsend, D.M.; Tew, K.D. Glutathione S-transferase P-mediated protein S-glutathionylation of resident endoplasmic reticulum proteins influences sensitivity to drug-induced unfolded protein response. Antioxid. Redox Signal. 2017, 26, 247–261. [Google Scholar] [CrossRef]
- Jo, H.; Choe, S.S.; Shin, K.C.; Jang, H.; Lee, J.H.; Seong, J.K.; Back, S.H.; Kim, J.B. Endoplasmic reticulum stress induces hepatic steatosis via increased expression of the hepatic very low-density lipoprotein receptor. Hepatology 2013, 57, 1366–1377. [Google Scholar] [CrossRef]
- McKeegan, D.; Kaab, H.; Bain, M.; Kuleš, J.; O’Reilly, E.; Horvatić, A.; Eckersall, P.D.; Guillemin, N. Quantitative proteomics using tandem mass tags in relation to the acute phase protein response in chicken challenged with Escherichia coli lipopolysaccharide endotoxin. J. Proteom. 2019, 192, 64–77. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013; Available online: https://www.R-project.org/ (accessed on 5 June 2019).
- Komsta, L. Outliers: Tests for Outliers (R Package Version 0.14). 2011. Available online: https://rdrr.io/cran/outliers/ (accessed on 5 June 2019).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 1st ed.; Springer: Cham, Switzerland, 2009. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession No. | Primer Sequence Forward (5′-3′) Reverse (5′-3′) | Optimized qPCR Conditions (Annealing Temp/MgCl) |
|---|---|---|---|
| ERS markers | |||
| ATF4 | NM_009716.3 | CCACTCCAGAGCATTCCTTTAG CTCCTTTACACATGGAGGGATTAG | 59 °C; 3.5 mM |
| BIP | NM_001163434.1 | GAGACTGCTGAGGCGTATTT CAGCATCTTTGGTTGCTTGTC | 58 °C; 3.5 mM |
| CHOP | NM_007837.4 | TTGAGCCTAACACGTCGATTAT CACTTCCTTCTGGAACACTCTC | 58 °C; 3 mM |
| EDEM | NM_138677.2 | TGAAAGCATGTGAGGGTAGTG GAGAGAAGGGAAGACAGGATAGA | 61 °C; 3.5 mM |
| GRP94 | NM_011631.1 | AAGAATGAAGGAAAAACAGGACAAAA CAAATGGAGAAGATTCCGCC | 58 °C; 3 mM |
| sXBP1 | NM_008934.4 | GAGTCCGCAGCAGGTG GTGTCAGAGTCCATGGGA | 56 °C; 3 mM |
| Cytokines | |||
| CXCL1 | NM_008176.3 | GTGTCAACCACTGTGCTAGT CACACATGTCCTCACCCTAATAC | 61 °C; 3.5 mM |
| IL1α | NM_010554.4 | CCTTACACCTACCAGAGTGATTT CCTTACACCTACCAGAGTGATTT | 65 °C; 3 mM |
| IL1β | NM_008361.4 | ATGGGCAACCACTTACCTATTT GTTCTAGAGAGTGCTGCCTAATG | 64 °C; 3 mM |
| IL6 | NM_031168.2 | GATAAGCTGGAGTCACAGAAGG TTGCCGAGTAGATCTCAAAGTG | 59 °C; 3.5 mM |
| MCP1 | NM_011333.3 | CCTGGATCGGAACCAAATGA CGGGTCAACTTCACATTCAAAG | 62 °C; 3 mM |
| TGFα | NM_031199.4 | CTTTAGGAAGGACCTGGGTTG GTGTGTCCAGGCTCCAAATA | 66 °C; 3 mM |
| TNFα | NM_013693.3 | GTCTCAGAATGAGGCTGGATAAG CATTGCACCTCAGGGAAGAA | 63 °C; 2.5 mM |
| Oxidative stress markers | |||
| COX1 | NM_008969.4 | GTGCCAGAACCAGGGTGTCT GTAGCCCGTGCGAGTACAATC | 58 °C 3 mM |
| GPX1 | NM_008160.6 | GGTTCGAGCCCAATTTTACA CATTCCGCAGGAAGGTAAAG | 58 °C 2.5 mM |
| NOX2 | NM_007807.5 | ACTCCTTGGGTCAGCACTGG GTTCCTGTCCAGTTGTCTTCG | 62 °C 3 mM |
| SOD1 | NM_011434.2 | GCCTTCTGCTCGAAGTGGAT GGAAGCATGGCGATGAAAGC | 59 °C 3.5 mM |
| SOD3 | NM_011435.3 | TGGCTGATGGTTGTACCCTG TGAGAAGATAGGCGACACGC | 60 °C 2.5 mM |
| Housekeeping genes | |||
| ACTB | NM_007393.5 | GCAAGCAGGAGTACGATGAG CCATGCCAATGTTGTCTCTT | 61 °C; 3.5 mM |
| B2M | NM_009735.3 | CCTGCAGAGTTAAGCATGACAGT TCATGATGCTTGATCACATGTCT | 60 °C; 3 mM |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šešelja, K.; Bazina, I.; Welss, J.; Schicht, M.; Paulsen, F.; Bijelić, N.; Rođak, E.; Horvatić, A.; Gelemanović, A.; Mihalj, M.; et al. Effect of Tff3 Deficiency and ER Stress in the Liver. Int. J. Mol. Sci. 2019, 20, 4389. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184389
Šešelja K, Bazina I, Welss J, Schicht M, Paulsen F, Bijelić N, Rođak E, Horvatić A, Gelemanović A, Mihalj M, et al. Effect of Tff3 Deficiency and ER Stress in the Liver. International Journal of Molecular Sciences. 2019; 20(18):4389. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184389
Chicago/Turabian StyleŠešelja, Kate, Iva Bazina, Jessica Welss, Martin Schicht, Friedrich Paulsen, Nikola Bijelić, Edi Rođak, Anita Horvatić, Andrea Gelemanović, Martina Mihalj, and et al. 2019. "Effect of Tff3 Deficiency and ER Stress in the Liver" International Journal of Molecular Sciences 20, no. 18: 4389. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20184389