Host Genotype and Precipitation Influence of Fungal Endophyte Symbiosis and Mycotoxin Abundance in a Locoweed

 ,
,  and
and
Abstract
:1. Introduction
2. Results
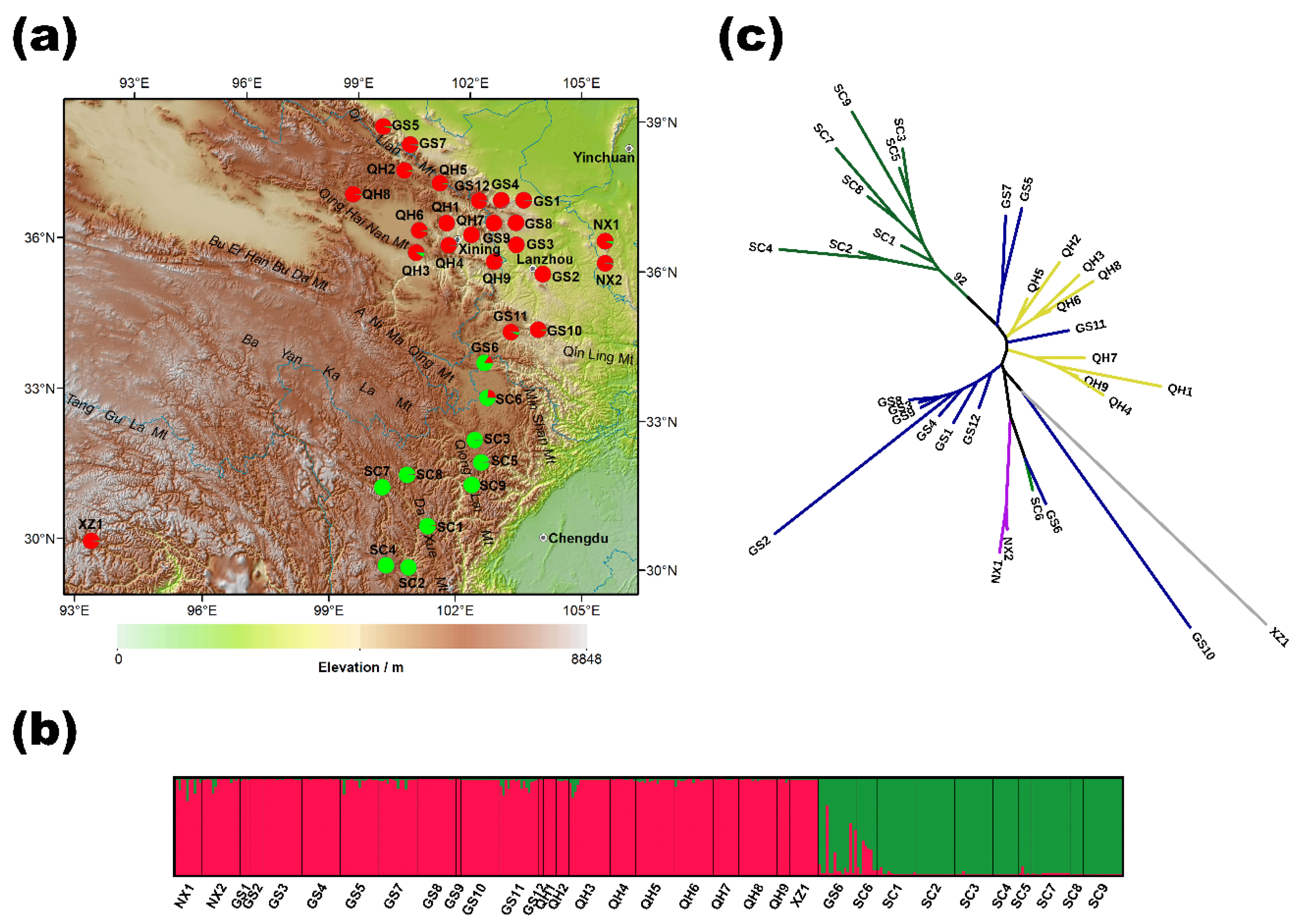
2.1. Genetic Variation in O. ochrocephala
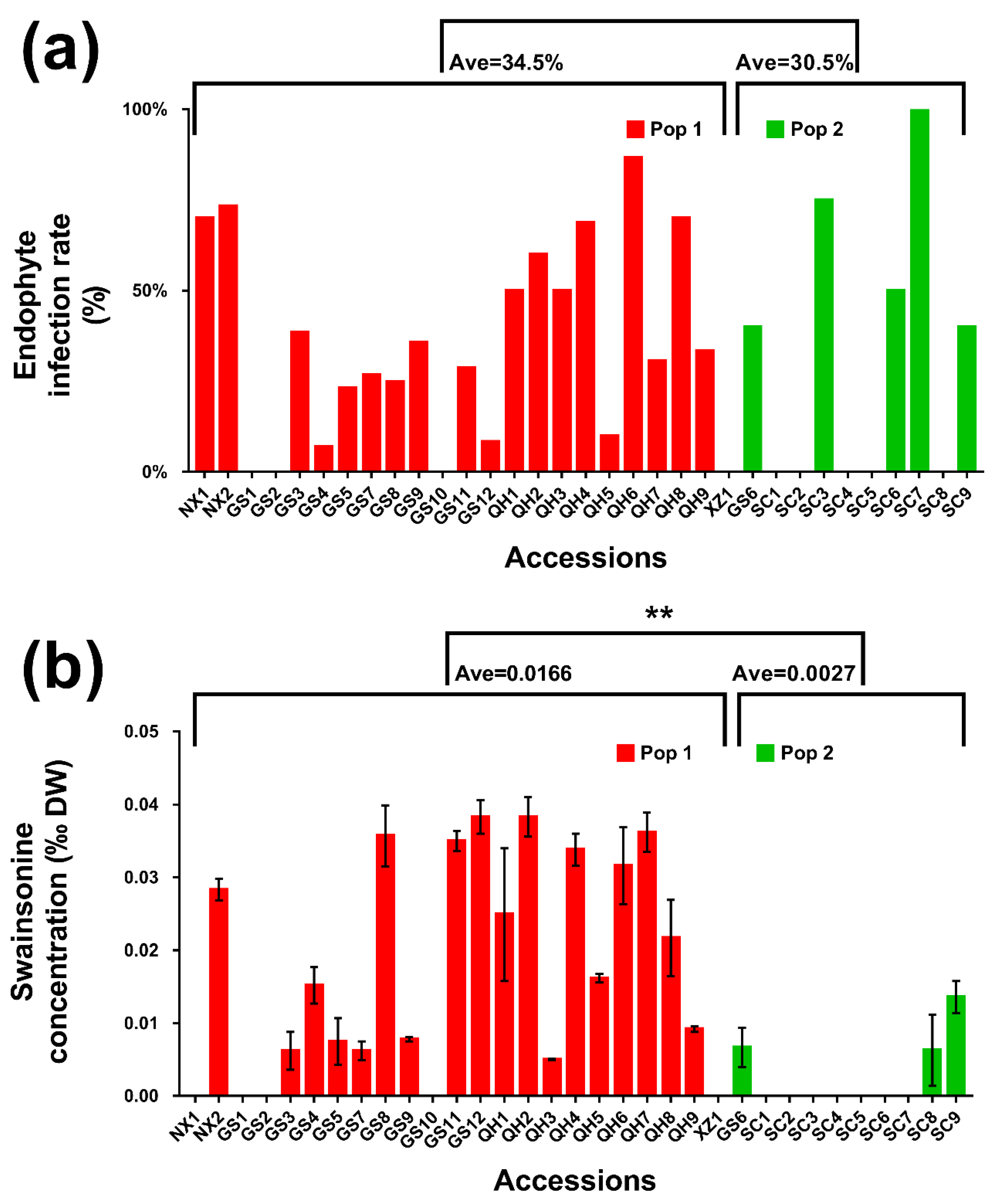
2.2. Endophyte Infection Rate and Swainsonine Concentrations
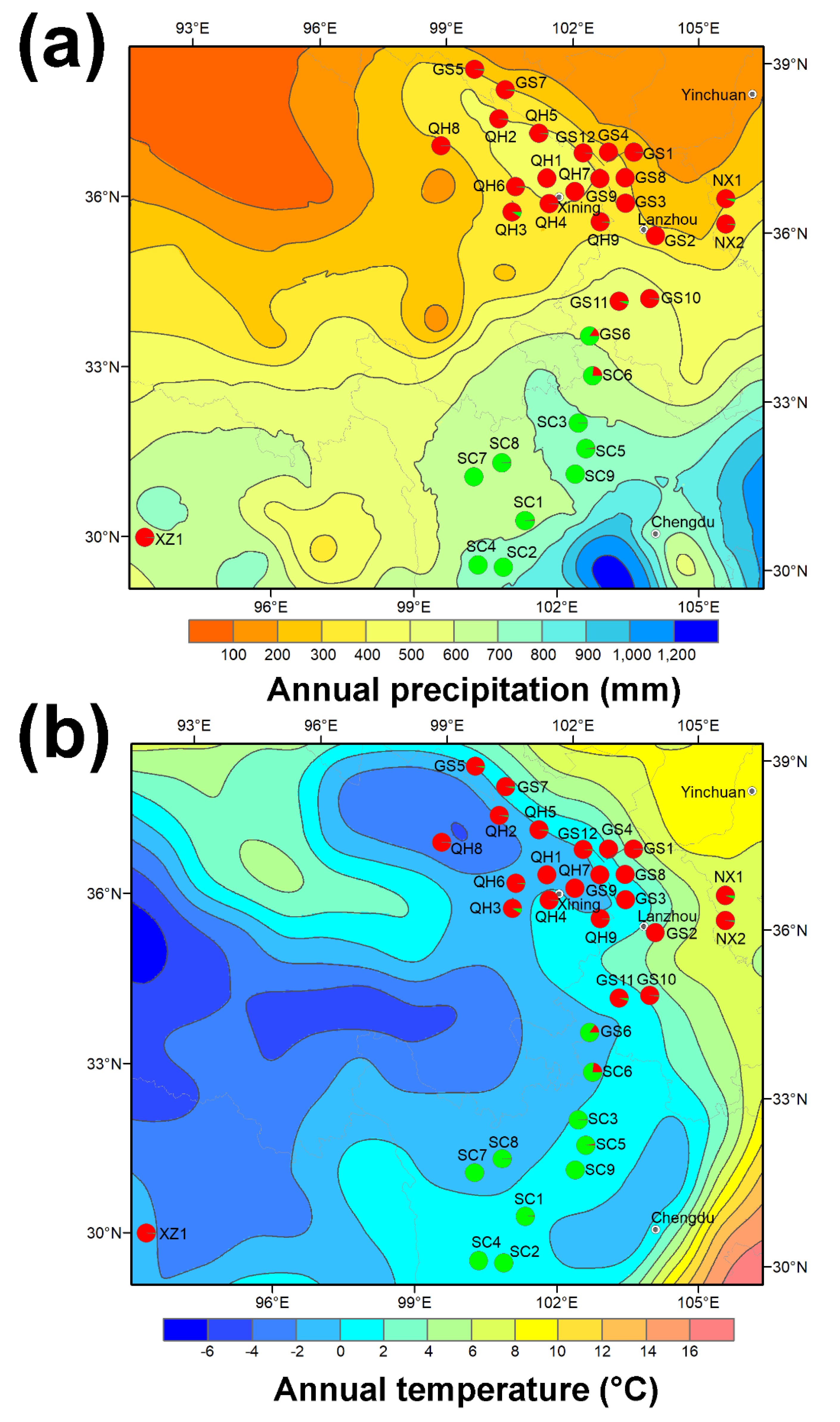
2.3. Interaction between Genetics and Environment and Their Integrated Effect on Endophyte Symbiosis and Swainsonine Production
3. Discussion
4. Materials and Methods
4.1. Sample Collection and DNA Extraction
4.2. Microsatellite Genotyping
4.3. Measuring Swainsonine Content
4.4. Determination of Endophyte Infection Rate
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Young, C.A.; Helander, M.; Schardl, C.L. Endophytic Epichloe species and their grass hosts: From evolution to applications. Plant Mol. Biol. 2016, 90, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.M.; Uroz, S.; Barker, D.G. Ancestral alliances: Plant mutualistic symbioses with fungi and bacteria. Science 2017, 356, eaad4501. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Gundel, P.E.; Helander, M. Chemical ecology mediated by fungal endophytes in grasses. J. Chem. Ecol. 2013, 39, 962–968. [Google Scholar] [CrossRef]
- Faeth, S.; Saikkonen, K. Variability is the nature of the endophyte-grass interaction. In Proceedings of the 6th International Symposium on Fungal Endophytes of Grasses, Christchurch, New Zealand, 25–28 March 2006. [Google Scholar]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J.; Biere, A. Herbivore-specific induction of defence metabolites in a grass-endophyte association. Funct. Ecol. 2017, 31, 318–324. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Bortfeld-Miller, M.; Ackermann, M.; Vorholt, J.A. A synthetic community approach reveals plant genotypes affecting the phyllosphere microbiota. PLoS Genet 2014, 10, e1004283. [Google Scholar] [CrossRef]
- Sapkota, R.; Knorr, K.; Jorgensen, L.N.; O’Hanlon, K.A.; Nicolaisen, M. Host genotype is an important determinant of the cereal phyllosphere mycobiome. New Phytol. 2015, 207, 1134–1144. [Google Scholar] [CrossRef]
- Compant, S.; Saikkonen, K.; Mitter, B.; Campisano, A.; Mercado-Blanco, J. Editorial special issue: Soil, plants and endophytes. Plant Soil 2016, 405, 1–11. [Google Scholar] [CrossRef]
- Zhao, M.; Gao, X.; Wang, J.; He, X.; Han, B. A review of the most economically important poisonous plants to the livestock industry on temperate grasslands of China. J. Appl. Toxicol. 2013, 33, 9–17. [Google Scholar] [CrossRef]
- Wang, Q.; Nagao, H.; Li, Y.; Wang, H.; Kakishima, M. Embellisia oxytropis, a new species isolated from Oxytropis kansuensis in China. Mycotaxon 2006, 95, 255–260. [Google Scholar]
- Pryor, B.; Creamer, R.; Shoemaker, R.; McLain-Romero, J.; Hambleton, S. Undifilum, a new genus for endophytic Embellisia oxytropis and parasitic Helminthosporium bornmuelleri on legumes. Botany 2009, 87, 178–194. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molyneux, R.J.; James, L.F. Loco intoxication: Indolizidine alkaloids of spotted locoweed (Astragalus lentiginosus). Science 1982, 216, 190–191. [Google Scholar] [CrossRef]
- Creamer, R.; Baucom, D. Fungal endophytes of locoweeds: A commensal relationship? J. Plant Physiol. Pathol. 2013, 1. [Google Scholar] [CrossRef]
- Ralphs, M.; Cook, D.; Gardner, D.; Grum, D. Transmission of the locoweed endophyte to the next generation of plants. Fungal Ecol. 2011, 4, 251–255. [Google Scholar] [CrossRef]
- Cook, D.; Grum, D.S.; Gardner, D.R.; Welch, K.D.; Pfister, J.A. Influence of endophyte genotype on swainsonine concentrations in Oxytropis sericea. Toxicon Off. J. Int. Soc. Toxinology 2013, 61, 105–111. [Google Scholar] [CrossRef]
- Cook, D.; Gardner, D.R.; Pfister, J.A. Swainsonine-containing plants and their relationship to endophytic fungi. J Agric Food Chem 2014, 62, 7326–7334. [Google Scholar] [CrossRef]
- Chung, M.; Gelembiuk, G.; Givnish, T.J. Population genetics and phylogeography of endangered Oxytropis campestris var. chartacea and relatives: Arctic-alpine disjuncts in eastern North America. Mol. Ecol. 2004, 13, 3657–3673. [Google Scholar] [CrossRef]
- Morjan, C.L.; Rieseberg, L.H. How species evolve collectively: Implications of gene flow and selection for the spread of advantageous alleles. Mol. Ecol. 2004, 13, 1341–1356. [Google Scholar] [CrossRef]
- Kozurahova, E.; Richards, A.J. Breeding sysmtems of rare and endemic Astragalus, Oxytropis and Onobrychis species (Fabaceae) tested with alternative methods. Comptes Rendus De L’académie Bulg. Des. Sci. 2016, 69. [Google Scholar]
- Archambault, A.; Strömvik, M.V. Evolutionary relationships in Oxytropis species, as estimated from the nuclear ribosomal internal transcribed spacer (ITS) sequences point to multiple expansions into the Arctic. Botany 2012, 90, 770–779. [Google Scholar] [CrossRef]
- Harrison, J.G.; Parchman, T.L.; Cook, D.; Gardner, D.R.; Forister, M.L. A heritable symbiont and host-associated factors shape fungal endophyte communities across spatial scales. J. Ecol. 2018, 106, 2274–2286. [Google Scholar] [CrossRef]
- Schirrmann, M.K.; Zoller, S.; Croll, D.; Stukenbrock, E.H.; Leuchtmann, A.; Fior, S. Genomewide signatures of selection in Epichloë reveal candidate genes for host specialization. Mol. Ecol. 2018, 27, 3070–3086. [Google Scholar] [CrossRef] [PubMed]
- Gladieux, P. What makes a specialized endophyte special? Mol. Ecol. 2018, 27, 3037–3039. [Google Scholar] [CrossRef] [Green Version]
- Saikkonen, K.; Wali, P.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef]
- Li, Y.; Nan, Z. A new species, Embellisia astragali sp. nov., causing standing milk-vetch disease in China. Mycologia 2007, 99, 406–411. [Google Scholar] [CrossRef]
- Cook, D.; Donzelli, B.G.; Creamer, R.; Baucom, D.L.; Gardner, D.R.; Pan, J.; Moore, N.; Krasnoff, S.B.; Jaromczyk, J.W.; Schardl, C.L. Swainsonine biosynthesis genes in diverse symbiotic and pathogenic fungi. G3: Genes Genomes Genet. 2017, 7, 1791–1797. [Google Scholar] [CrossRef]
- Oldrup, E.; McLain-Romero, J.; Padilla, A.; Moya, A.; Gardner, D.; Creamer, R. Localization of endophytic Undifilum fungi in locoweed seed and influence of environmental parameters on a locoweed in vitro culture system. Botany 2010, 88, 512–521. [Google Scholar] [CrossRef]
- He, W.; Zhuang, H.; Fu, Y.; Guo, L.; Guo, B.; Guo, L.; Zhang, X.; Wei, Y. De novo transcriptome assembly of a Chinese locoweed (Oxytropis ochrocephala) species provides insights into genes associated with drought, salinity, and cold tolerance. Front Plant Sci. 2015, 6, 1086. [Google Scholar] [CrossRef]
- Wu, C.; Zhao, B.; Lu, H.; Ma, F.; Cao, D.; Liu, X.; Liu, Z.; Wang, Z. Dynamic change of swainsonine in major locoweed species in China. Acta Agric. Boreali-Occident. Sin. 2014, 23, 51–58. [Google Scholar]
- Duan, Q.; Huang, X.; Li, C.; Ni, J.; Di, D. Pretreatment of sample based on macroporous adsorptive resin and content determination of swainsonine in Oxytropis kansuensis Bunge by HPLC. Anal. Test. Technol. Instrum. 2010, 16, 6–10. [Google Scholar]
- Cook, D.; Gardner, D.R.; Welch, K.D.; Roper, J.M.; Ralphs, M.H.; Green, B.T. Quantitative PCR method to measure the fungal endophyte in locoweeds. J. Agric. Food Chem. 2009, 57, 6050–6054. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Quan, H.; Zhou, Q.; Ren, Z.; Xue, R.; Zhao, B.; Creamer, R. Endogenous fungi isolated from three locoweed species from rangeland in western China. Afr. J. Microbiol. Res. 2017, 11, 155–170. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 157. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 117693430500100003. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Resour. 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Cornuet, J.-M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.-M.; Estoup, A. DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar]
- Jensen, J.L.; Bohonak, A.J.; Kelley, S.T. Isolation by distance, web service. BMC Genet. 2005, 6, 13. [Google Scholar] [CrossRef]
- Miller, M.P. Available online: http://www.marksgeneticsoftware.net/tfpga.htm (accessed on 14 October 2018).
- Smouse, P.E.; Long, J.C.; Sokal, R.R. Multiple regression and correlation extensions of the Mantel test of matrix correspondence. Syst. Zool. 1986, 35, 627–632. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation | Fixation Indices |
|---|---|---|---|---|---|
| Amongst populations | 1 | 173.928 | 0.505 Va | 9.69 | Fct = 0.0969 *** |
| Amongst accessions within populations | 31 | 757.888 | 0.937 Vb | 17.95 | Fsc = 0.199 *** |
| Within accessions | 703 | 2653.730 | 3.775 Vc | 72.36 | Fst = 0.276 *** |
| Total | 735 | 3585.546 | 5.217 | 100 |
| Environment | Elevation | Genetics | |||
|---|---|---|---|---|---|
| Genetic Distance | Population Partition | Cluster | He | ||
| Geographic distance | --- | r = 0.523 *** | --- | --- | --- |
| Elevation | --- | r = 0.424 *** | r = 0.621 *** | r = 0.662 *** | r = 0.010 |
| Annual precipitation | r = 0.730 *** | r = 0.484 *** | r = 0.584 *** | r = 0.762 *** | r = −0.256 |
| Annual temperature | r = −0.489 ** | r = 0.258 * | r = −0.067 | r = 0.049 | r = −0.371 * |
| Factors | Endophyte Infection Rate | Swainsonine Content | |
|---|---|---|---|
| G | Genetic distance | r = 0.157 * | r = 0.218 *** |
| Population partition | r = 0.014 | r = 0.492 ** | |
| Cluster | r = 0.136 | r = 0.570 ** | |
| He | r = 0.020 | r = 0.377 * | |
| E | Geographic distance | r = 0.093 | r = 0.340 *** |
| Elevation | r = 0.001 | r = −0.197 | |
| Annual precipitation | r = −0.261 | r = −0.444 * | |
| Annul temperature | r = 0.008 | r = −0.096 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.; Guo, L.; Wang, L.; Zhao, Q.; Guo, L.; Cao, W.; Mur, L.A.J.; Wei, Y. Host Genotype and Precipitation Influence of Fungal Endophyte Symbiosis and Mycotoxin Abundance in a Locoweed. Int. J. Mol. Sci. 2019, 20, 5285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215285
He W, Guo L, Wang L, Zhao Q, Guo L, Cao W, Mur LAJ, Wei Y. Host Genotype and Precipitation Influence of Fungal Endophyte Symbiosis and Mycotoxin Abundance in a Locoweed. International Journal of Molecular Sciences. 2019; 20(21):5285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215285
Chicago/Turabian StyleHe, Wei, Linwei Guo, Lei Wang, Qianqian Zhao, Lizhu Guo, Wei Cao, Luis A. J. Mur, and Yahui Wei. 2019. "Host Genotype and Precipitation Influence of Fungal Endophyte Symbiosis and Mycotoxin Abundance in a Locoweed" International Journal of Molecular Sciences 20, no. 21: 5285. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215285