Comparatively Barcoded Chromosomes of Brachypodium Perennials Tell the Story of Their Karyotype Structure and Evolution

 , ,
, ,  ,
, 
Abstract
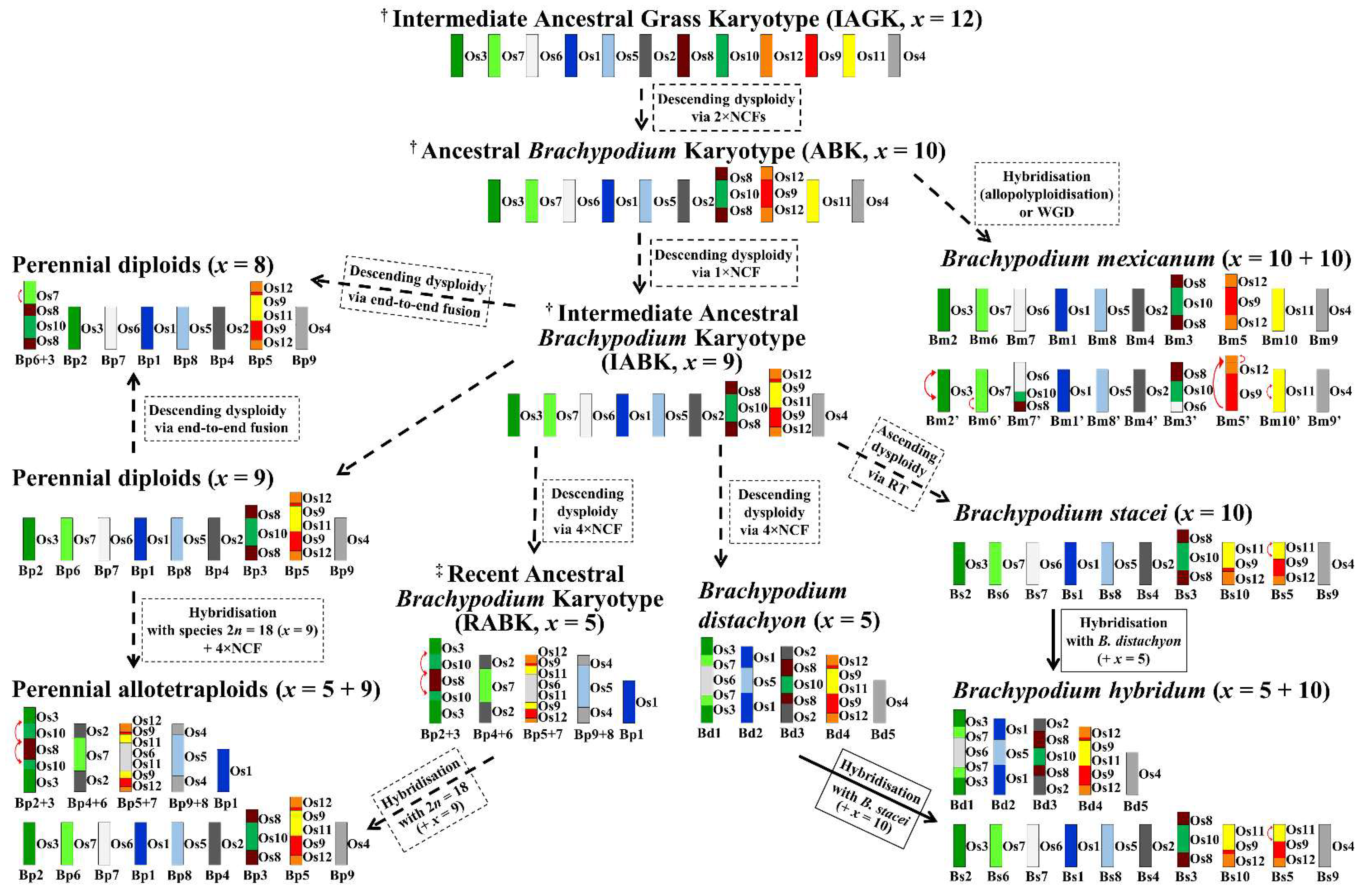
:1. Introduction
2. Results
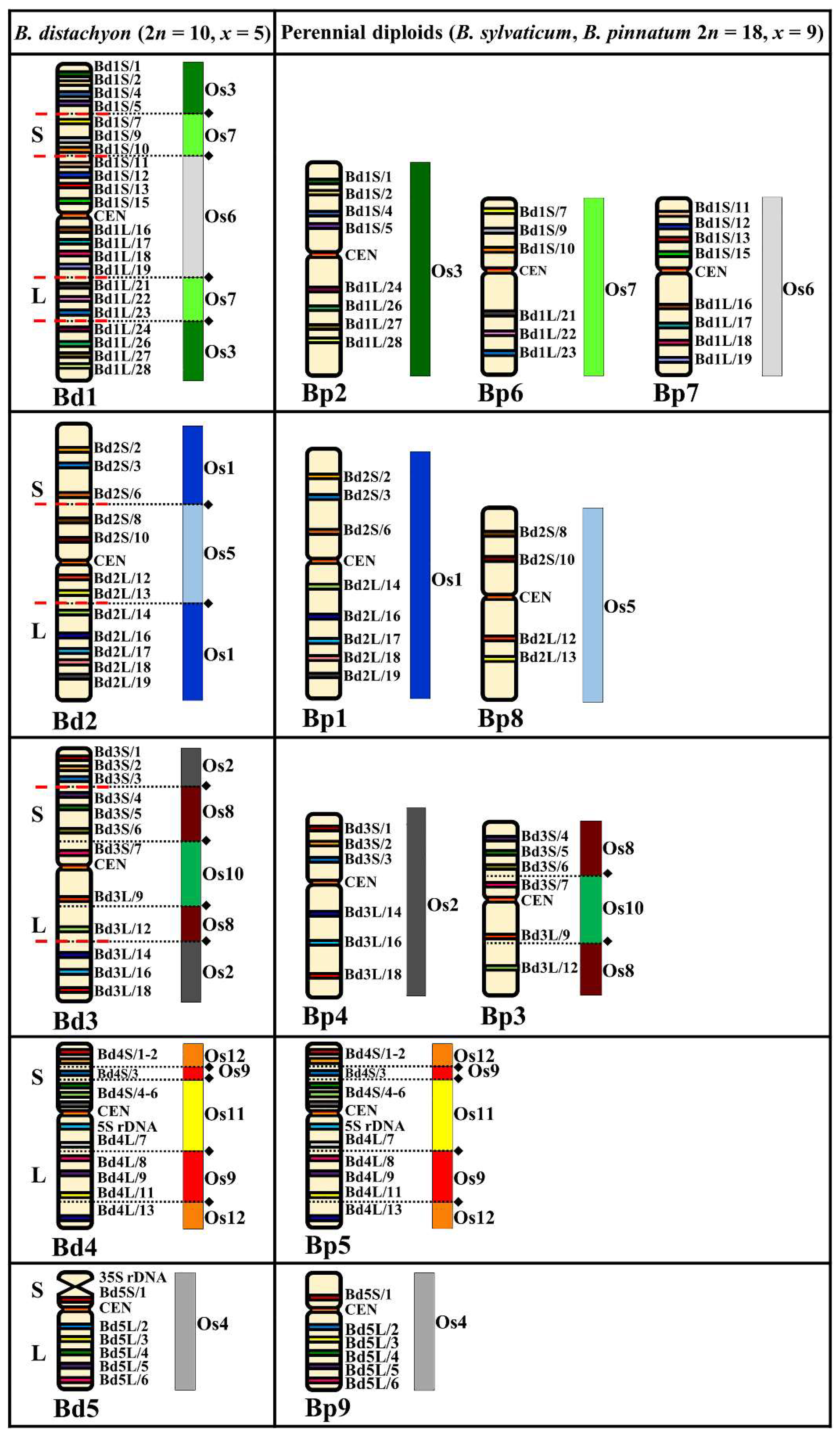
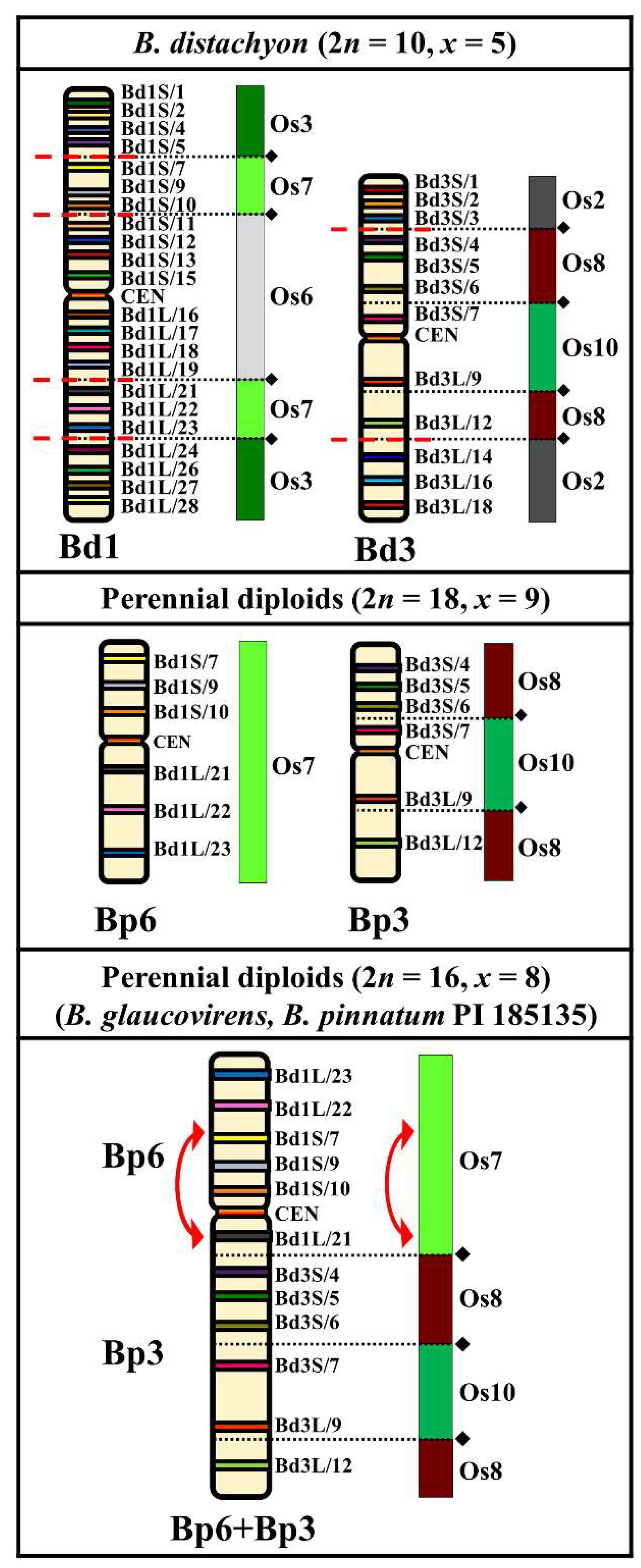
2.1. Comparative Mapping of the Chromosomes in the Perennial Diploids (2n = 18, 16 Chromosomes)
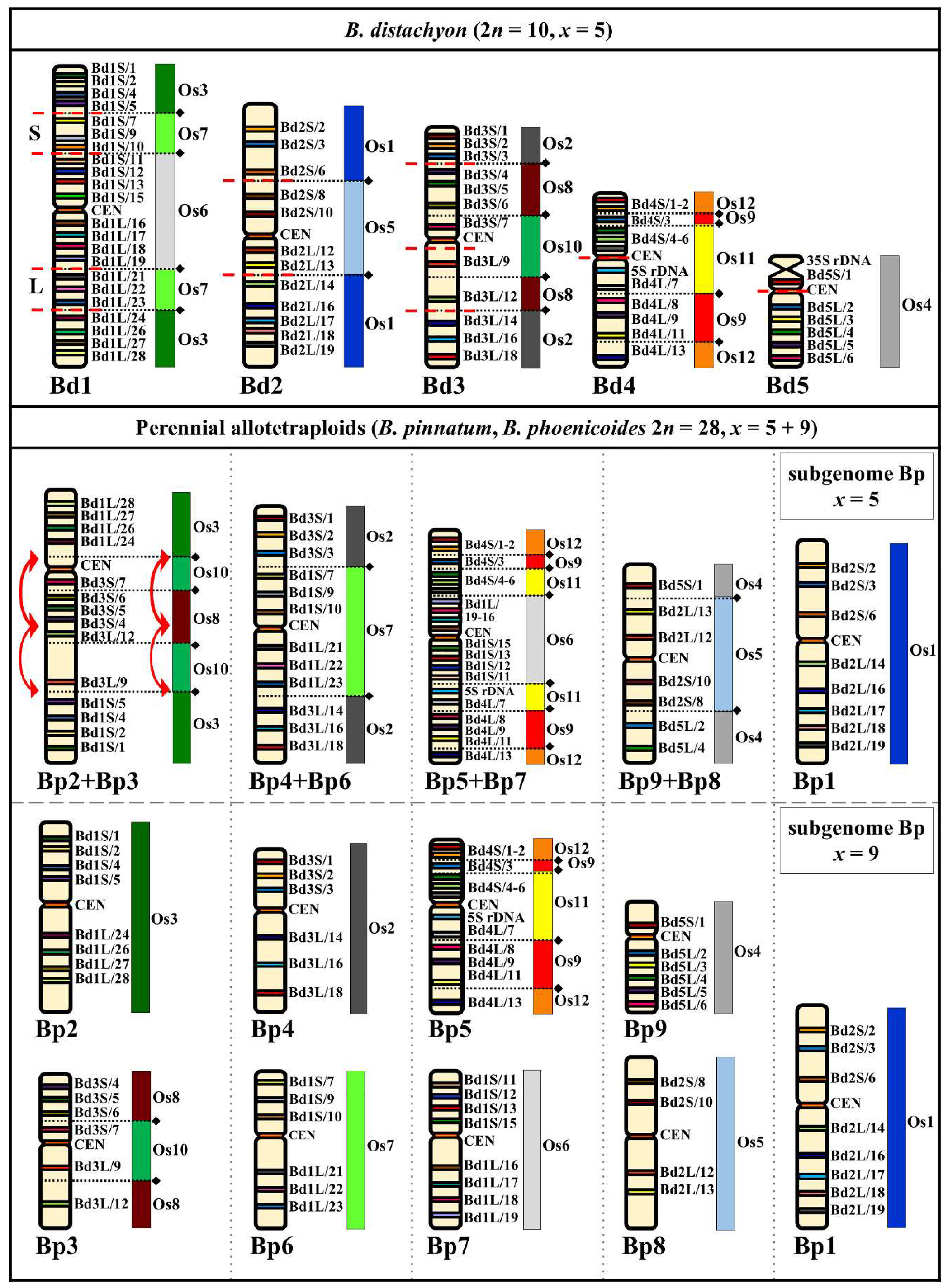
2.2. Comparative Mapping of the Chromosomes in the Perennial Allotetraploids (2n = 28 Chromosomes)
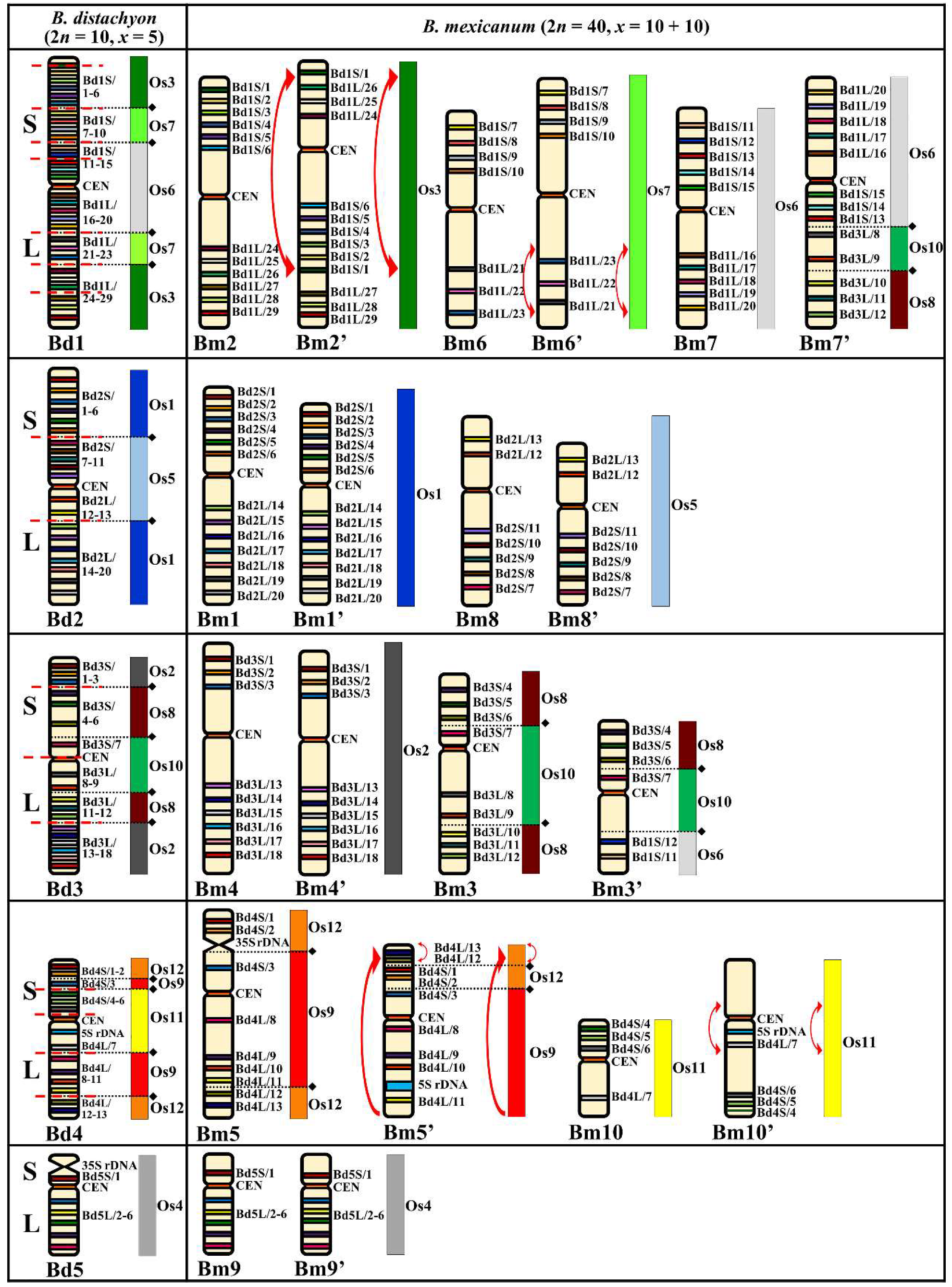
2.3. Comparative Chromosome Barcoding of B. mexicanum (2n = 40 Chromosomes)
3. Discussion
3.1. Karyotype Evolution in the Perennial Diploids
3.2. Karyotype Evolution in the Perennial Polyploids
3.3. Brachypodium Karyotype Evolution
4. Materials and Methods
4.1. Plant Material
4.2. Chromosome Preparation
4.3. Probe Labelling and FISH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABK | Ancestral Brachypodium Karyotype (x = 10) |
| ARCE | Ancestral rice chromosome equivalent |
| Bd | Chromosome complement of B. distachyon (x = 5) |
| Bp | Chromosome complement(s) of Brachypodium perennials (x = 9, 8, and 5) |
| Bm (Bm’) | Chromosome complement(s) of B. mexicanum (x = 10 + 10) |
| Bs | Chromosome complement of B. stacei (x = 10) |
| BAC | Bacterial artificial chromosome |
| CCB | Comparative chromosome barcoding |
| CCP | Comparative chromosome painting |
| CEN | Centromeric BAC BD_CBa0033J12 |
| EEF | End-to-end fusion |
| FISH | Fluorescence in situ hybridization |
| IABK | Intermediate Ancestral Brachypodium Karyotype (x = 9) |
| IAGK | Intermediate Ancestral Grass Karyotype (x = 12) |
| Ma | Million years |
| NCF | Nested chromosome fusion |
| RABK | Recent Ancestral Brachypodium Karyotype (x = 5) |
| WGD | Whole genome duplication |
References
- Mur, L.A.; Allainguillaume, J.; Catalan, P.; Hasterok, R.; Jenkins, G.; Lesniewska, K.; Thomas, I.; Vogel, J. Exploiting the Brachypodium tool box in cereal and grass research. New Phytol. 2011, 191, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P. The rise of Brachypodium as a model system. In Genetics and Genomics of Brachypodium; Vogel, J.P., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–7. [Google Scholar] [CrossRef]
- Scholthof, K.G.; Irigoyen, S.; Catalan, P.; Mandadi, K.K. Brachypodium: A monocot grass model genus for plant biology. Plant Cell 2018, 30, 1673–1694. [Google Scholar] [CrossRef] [PubMed]
- IBI. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Catalan, P.; Chalhoub, B.; Chochois, V.; Garvin, D.F.; Hasterok, R.; Manzaneda, A.J.; Mur, L.A.J.; Pecchioni, N.; Rasmussen, S.K.; Vogel, J.P.; et al. Update on the genomics and basic biology of Brachypodium. Trends Plant Sci. 2014, 19, 414–418. [Google Scholar] [CrossRef]
- Catalan, P.; Muller, J.; Hasterok, R.; Jenkins, G.; Mur, L.A.; Langdon, T.; Betekhtin, A.; Siwinska, D.; Pimentel, M.; Lopez-Alvarez, D. Evolution and taxonomic split of the model grass Brachypodium distachyon. Ann Bot. 2012, 109, 385–405. [Google Scholar] [CrossRef]
- Gordon, S.P.; Liu, L.; Vogel, J.P. The genus Brachypodium as a model for perenniality and polyploidy. In Genetics and Genomics of Brachypodium; Vogel, J.P., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 313–325. [Google Scholar] [CrossRef]
- Catalan, P.; López-Álvarez, D.; Díaz-Pérez, A.; Sancho, R.; López-Herránz, M.L. Phylogeny and evolution of the genus Brachypodium. In Genetics and Genomics of Brachypodium; Vogel, J.P., Ed.; Springer International Group: Cham, Switzerland, 2016; pp. 9–38. [Google Scholar] [CrossRef]
- Betekhtin, A.; Jenkins, G.; Hasterok, R. Reconstructing the evolution of Brachypodium genomes using comparative chromosome painting. PLoS ONE 2014, 9, e115108. [Google Scholar] [CrossRef]
- Khan, M.A.; Stace, C.A. Breeding relationships in the genus Brachypodium (Poaceae). Nord J. Bot. 1999, 19, 257–269. [Google Scholar] [CrossRef]
- Robertson, I.H. Chromosome numbers in Brachypodium Beauv (Gramineae). Genetica 1981, 56, 55–60. [Google Scholar] [CrossRef]
- Wolny, E.; Hasterok, R. Comparative cytogenetic analysis of the genomes of the model grass Brachypodium distachyon and its close relatives. Ann. Bot. 2009, 104, 873–881. [Google Scholar] [CrossRef]
- Díaz-Pérez, A.; López-Álvarez, D.; Sancho, R.; Catalán, P. Reconstructing the origins and the biogeography of species’ genomes in the highly reticulate allopolyploid-rich model grass genus Brachypodium using minimum evolution, coalescence and maximum likelihood approaches. Mol. Phylogenet. Evol. 2018, 127, 256–271. [Google Scholar] [CrossRef]
- Wolny, E.; Lesniewska, K.; Hasterok, R.; Langdon, T. Compact genomes and complex evolution in the genus Brachypodium. Chromosoma 2011, 120, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Sancho, R.; Cantalapiedra, C.P.; Lopez-Alvarez, D.; Gordon, S.P.; Vogel, J.P.; Catalan, P.; Contreras-Moreira, B. Comparative plastome genomics and phylogenomics of Brachypodium: Flowering time signatures, introgression and recombination in recently diverged ecotypes. New Phytol. 2018, 218, 1631–1644. [Google Scholar] [CrossRef] [PubMed]
- Catalan, P.; Shi, Y.; Armstrong, L.; Draper, J.; Stace, C.A. Molecular phylogeny of the grass genus Brachypodium P. Beauv. based on RFLP and RAPD analysis. Bot. J. Linn Soc. 1995, 177, 263–280. [Google Scholar] [CrossRef]
- Gordon, S.P.; Contreras-Moreira, B.; Woods, D.P.; Des Marais, D.L.; Burgess, D.; Shu, S.; Stritt, C.; Roulin, A.C.; Schackwitz, W.; Tyler, L.; et al. Extensive gene content variation in the Brachypodium distachyon pan-genome correlates with population structure. Nat. Commun. 2017, 8, 2184. [Google Scholar] [CrossRef] [PubMed]
- Steinwand, M.A.; Young, H.A.; Bragg, J.N.; Tobias, C.M.; Vogel, J.P. Brachypodium sylvaticum, a model for perennial grasses: Transformation and inbred line development. PLoS ONE 2013, 8, e75180. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.E.; Preece, J.; Kimbrel, J.A.; Marchini, G.L.; Sage, A.; Youens-Clark, K.; Cruzan, M.B.; Jaiswal, P. Sequencing and de novo transcriptome assembly of Brachypodium sylvaticum (Poaceae). Appl. Plant Sci. 2013, 1. [Google Scholar] [CrossRef]
- Griffiths, S.; Sharp, R.; Foote, T.N.; Bertin, I.; Wanous, M.; Reader, S.; Colas, I.; Moore, G. Molecular characterization of Ph1 as a major chromosome pairing locus in polyploid wheat. Nature 2006, 439, 749–752. [Google Scholar] [CrossRef]
- Bossolini, E.; Wicker, T.; Knobel, P.A.; Keller, B. Comparison of orthologous loci from small grass genomes Brachypodium and rice: Implications for wheat genomics and grass genome annotation. Plant J. 2007, 49, 704–717. [Google Scholar] [CrossRef]
- Murat, F.; Armero, A.; Pont, C.; Klopp, C.; Salse, J. Reconstructing the genome of the most recent common ancestor of flowering plants. Nat. Genet. 2017, 49, 490–496. [Google Scholar] [CrossRef]
- Pont, C.; Wagner, S.; Kremer, A.; Orlando, L.; Plomion, C.; Salse, J. Paleogenomics: Reconstruction of plant evolutionary trajectories from modern and ancient DNA. Genome Biol. 2019, 20, 29. [Google Scholar] [CrossRef]
- Salse, J. Deciphering the evolutionary interplay between subgenomes following polyploidy: A paleogenomics approach in grasses. Am. J. Bot. 2016, 103, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salse, J. Ancestors of modern plant crops. Curr. Opin. Plant Biol. 2016, 30, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Alix, K.; Gerard, P.R.; Schwarzacher, T.; Heslop-Harrison, J.S.P. Polyploidy and interspecific hybridization: Partners for adaptation, speciation and evolution in plants. Ann. Bot. 2017, 120, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Sherman-Broyles, S. Double trouble: Taxonomy and definitions of polyploidy. New Phytol. 2017, 213, 487–493. [Google Scholar] [CrossRef]
- Čertner, M.; Fenclová, E.; Kúr, P.; Kolář, F.; Koutecký, P.; Krahulcová, A.; Suda, J. Evolutionary dynamics of mixed-ploidy populations in an annual herb: Dispersal, local persistence and recurrent origins of polyploids. Ann. Bot. 2017, 120, 303–315. [Google Scholar] [CrossRef]
- Paule, J.; Wagner, N.D.; Weising, K.; Zizka, G. Ecological range shift in the polyploid members of the South American genus Fosterella (Bromeliaceae). Ann. Bot. 2017, 120, 233–243. [Google Scholar] [CrossRef]
- Cuadrado, A.; de Bustos, A.; Jouve, N. On the allopolyploid origin and genome structure of the closely related species Hordeum secalinum and Hordeum capense inferred by molecular karyotyping. Ann. Bot. 2017, 120, 245–255. [Google Scholar] [CrossRef]
- Hasterok, R.; Draper, J.; Jenkins, G. Laying the cytotaxonomic foundations of a new model grass, Brachypodium distachyon (L.) Beauv. Chromosome Res. 2004, 12, 397–403. [Google Scholar] [CrossRef]
- Hasterok, R.; Marasek, A.; Donnison, I.S.; Armstead, I.; Thomas, A.; King, I.P.; Wolny, E.; Idziak, D.; Draper, J.; Jenkins, G. Alignment of the genomes of Brachypodium distachyon and temperate cereals and grasses using bacterial artificial chromosome landing with fluorescence in situ hybridization. Genetics 2006, 173, 349–362. [Google Scholar] [CrossRef]
- Lopez-Alvarez, D.; Lopez-Herranz, M.L.; Betekhtin, A.; Catalan, P. A DNA barcoding method to discriminate between the model plant Brachypodium distachyon and its close relatives B. stacei and B. hybridum (Poaceae). PLoS ONE 2012, 7, e51058. [Google Scholar] [CrossRef] [PubMed]
- Idziak, D.; Hazuka, I.; Poliwczak, B.; Wiszynska, A.; Wolny, E.; Hasterok, R. Insight into the karyotype evolution of Brachypodium species using comparative chromosome barcoding. PLoS ONE 2014, 9, e93503. [Google Scholar] [CrossRef] [PubMed]
- Idziak-Helmcke, D.; Betekhtin, A. Methods for cytogenetic chromosome barcoding and chromosome painting in Brachypodium distachyon and its relative species. In Brachypodium Genomics: Methods and Protocols; Sablok, G., Budak, H., Ralph, P.J., Eds.; Springer New York: New York, NY, USA, 2018; pp. 1–19. [Google Scholar] [CrossRef]
- Jenkins, G.; Hasterok, R. BAC ‘landing’ on chromosomes of Brachypodium distachyon for comparative genome alignment. Nat. Protoc. 2007, 2, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Febrer, M.; Goicoechea, J.L.; Wright, J.; McKenzie, N.; Song, X.; Lin, J.; Collura, K.; Wissotski, M.; Yu, Y.; Ammiraju, J.S.; et al. An integrated physical, genetic and cytogenetic map of Brachypodium distachyon, a model system for grass research. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Lysak, M.A.; Berr, A.; Pecinka, A.; Schmidt, R.; McBreen, K.; Schubert, I. Mechanisms of chromosome number reduction in Arabidopsis thaliana and related Brassicaceae species. Proc. Natl. Acad. Sci. USA 2006, 103, 5224–5229. [Google Scholar] [CrossRef] [PubMed]
- Mandakova, T.; Guo, X.; Ozudogru, B.; Mummenhoff, K.; Lysak, M.A. Hybridization-facilitated genome merger and repeated chromosome fusion after 8 million years. Plant J. 2018, 96, 748–760. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Koo, D.-H.; Li, D.; Zhang, T.; Jiang, J.; Luan, F.; Renner, S.S.; Hénaff, E.; Sanseverino, W.; Garcia-Mas, J.; et al. Next-generation sequencing, FISH mapping and synteny-based modeling reveal mechanisms of decreasing dysploidy in Cucumis. Plant J. 2014, 77, 16–30. [Google Scholar] [CrossRef]
- Hou, L.; Xu, M.; Zhang, T.; Xu, Z.; Wang, W.; Zhang, J.; Yu, M.; Ji, W.; Zhu, C.; Gong, Z.; et al. Chromosome painting and its applications in cultivated and wild rice. BMC Plant Biol. 2018, 18, 110. [Google Scholar] [CrossRef]
- Albert, P.S.; Zhang, T.; Semrau, K.; Rouillard, J.M.; Kao, Y.H.; Wang, C.R.; Danilova, T.V.; Jiang, J.; Birchler, J.A. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 1679–1685. [Google Scholar] [CrossRef] [Green Version]
- Lusinska, J.; Majka, J.; Betekhtin, A.; Susek, K.; Wolny, E.; Hasterok, R. Chromosome identification and reconstruction of evolutionary rearrangements in Brachypodium distachyon, B. stacei and B. hybridum. Ann. Bot. 2018, 122, 445–459. [Google Scholar] [CrossRef]
- Wolny, E.; Fidyk, W.; Hasterok, R. Karyotyping of Brachypodium pinnatum (2n = 18) chromosomes using cross-species BAC-FISH. Genome 2013, 56, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Lysak, M.A. Interpretation of karyotype evolution should consider chromosome structural constraints. Trends Genet. 2011, 27, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Mandakova, T.; Lysak, M.A. Chromosomal phylogeny and karyotype evolution in x=7 crucifer species (Brassicaceae). Plant Cell 2008, 20, 2559–2570. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, D.; Wang, Z.; Guo, H.; Zhang, L.; Wang, L.; Li, J.; Paterson, A.H. Telomere-centric genome repatterning determines recurring chromosome number reductions during the evolution of eukaryotes. New Phytol. 2015, 205, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Dinh Thi, V.H.; Coriton, O.; Le Clainche, I.; Arnaud, D.; Gordon, S.P.; Linc, G.; Catalan, P.; Hasterok, R.; Vogel, J.P.; Jahier, J.; et al. Recreating stable Brachypodium hybridum allotetraploids by uniting the divergent genomes of B. distachyon and B. stacei. PLoS ONE 2016, 11, e0167171. [Google Scholar] [CrossRef] [PubMed]
- Mandakova, T.; Lysak, M.A. Post-polyploid diploidization and diversification through dysploid changes. Curr Opin Plant Biol. 2018, 42, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Murat, F.; Xu, J.H.; Tannier, E.; Abrouk, M.; Guilhot, N.; Pont, C.; Messing, J.; Salse, J. Ancestral grass karyotype reconstruction unravels new mechanisms of genome shuffling as a source of plant evolution. Genome Res. 2010, 20, 1545–1557. [Google Scholar] [CrossRef] [Green Version]
- Daverdin, G.; Bahri, B.A.; Wu, X.; Serba, D.D.; Tobias, C.; Saha, M.C.; Devos, K.M. Comparative relationships and chromosome evolution in switchgrass (Panicum virgatum) and its genomic model, foxtail millet (Setaria italica). BioEnergy Res. 2015, 8, 137–151. [Google Scholar] [CrossRef]
- Fonsêca, A.; Ferraz, M.E.; Pedrosa-Harand, A. Speeding up chromosome evolution in Phaseolus: Multiple rearrangements associated with a one-step descending dysploidy. Chromosoma 2015, 125, 413–421. [Google Scholar] [CrossRef]
- Mandakova, T.; Hlouskova, P.; German, D.A.; Lysak, M.A. Monophyletic origin and evolution of the largest crucifer genomes. Plant Physiol. 2017, 174, 2062–2071. [Google Scholar] [CrossRef]
- Idziak, D.; Betekhtin, A.; Wolny, E.; Lesniewska, K.; Wright, J.; Febrer, M.; Bevan, M.W.; Jenkins, G.; Hasterok, R. Painting the chromosomes of Brachypodium: Current status and future prospects. Chromosoma 2011, 120, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, J.; Pan, Y.; Lei, T.; Ge, W.; Wang, L.; Zhang, L.; Li, Y.; Zhao, K.; Liu, T.; et al. Reconstruction of evolutionary trajectories of chromosomes unraveled independent genomic repatterning between Triticeae and Brachypodium. BMC Genom. 2019, 20, 180. [Google Scholar] [CrossRef] [PubMed]
- Hasterok, R.; Dulawa, J.; Jenkins, G.; Leggett, M.; Langdon, T. Multi-substrate chromosome preparations for high throughput comparative FISH. BMC Biotechnol. 2006, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Unfried, I.; Gruendler, P. Nucleotide sequence of the 5.8S and 25S rRNA genes and of the internal transcribed spacers from Arabidopsis thaliana. Nucleic Acids Res. 1990, 18, 4011. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.L.; Dyer, T.A. Sequence organization of the repeating units in the nucleus of wheat which contain 5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4886. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number | 2n | x | Ploidy Level | Origin | Source * |
|---|---|---|---|---|---|---|
| B. distachyon | Bd21 | 10 | 5 | 2× | Iraq | USA |
| B. sylvaticum | PI 297868 | 18 | 9 | 2× | Australia | USA |
| PI 269842 | 18 | 9 | 2× | Tunisia | USA | |
| B. glaucovirens | PI 4202 | 16 | 8 | 2× | Greece, Crete | Germany |
| B. pinnatum | PI 185135 | 16 | 8 | 2× | Iraq | USA |
| PI 230113 | 18 | 9 | 2× | Iran | USA | |
| PI 345982 | 18 | 9 | 2× | Norway | USA | |
| PI 249722 | 28 | 5 + 9 | 4× | Greece | USA | |
| PI 251445 | 28 | 5 + 9 | 4× | Turkey | USA | |
| PI 430277 | 28 | 5 + 9 | 4× | Ireland | USA | |
| B. phoenicoides | PI 253503 | 28 | 5 + 9 | 4× | Spain | USA |
| PI 89817 | 28 | 5 + 9 | 4× | Spain | USA | |
| B. mexicanum | Bmex347 | 40 | 10 + 10 | 4× | Mexico | UK |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lusinska, J.; Betekhtin, A.; Lopez-Alvarez, D.; Catalan, P.; Jenkins, G.; Wolny, E.; Hasterok, R. Comparatively Barcoded Chromosomes of Brachypodium Perennials Tell the Story of Their Karyotype Structure and Evolution. Int. J. Mol. Sci. 2019, 20, 5557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225557
Lusinska J, Betekhtin A, Lopez-Alvarez D, Catalan P, Jenkins G, Wolny E, Hasterok R. Comparatively Barcoded Chromosomes of Brachypodium Perennials Tell the Story of Their Karyotype Structure and Evolution. International Journal of Molecular Sciences. 2019; 20(22):5557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225557
Chicago/Turabian StyleLusinska, Joanna, Alexander Betekhtin, Diana Lopez-Alvarez, Pilar Catalan, Glyn Jenkins, Elzbieta Wolny, and Robert Hasterok. 2019. "Comparatively Barcoded Chromosomes of Brachypodium Perennials Tell the Story of Their Karyotype Structure and Evolution" International Journal of Molecular Sciences 20, no. 22: 5557. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225557