Impressic Acid, a Lupane-Type Triterpenoid from Acanthopanax koreanum, Attenuates TNF-α-Induced Endothelial Dysfunction via Activation of eNOS/NO Pathway

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of IPA in EA.hy926 Cells
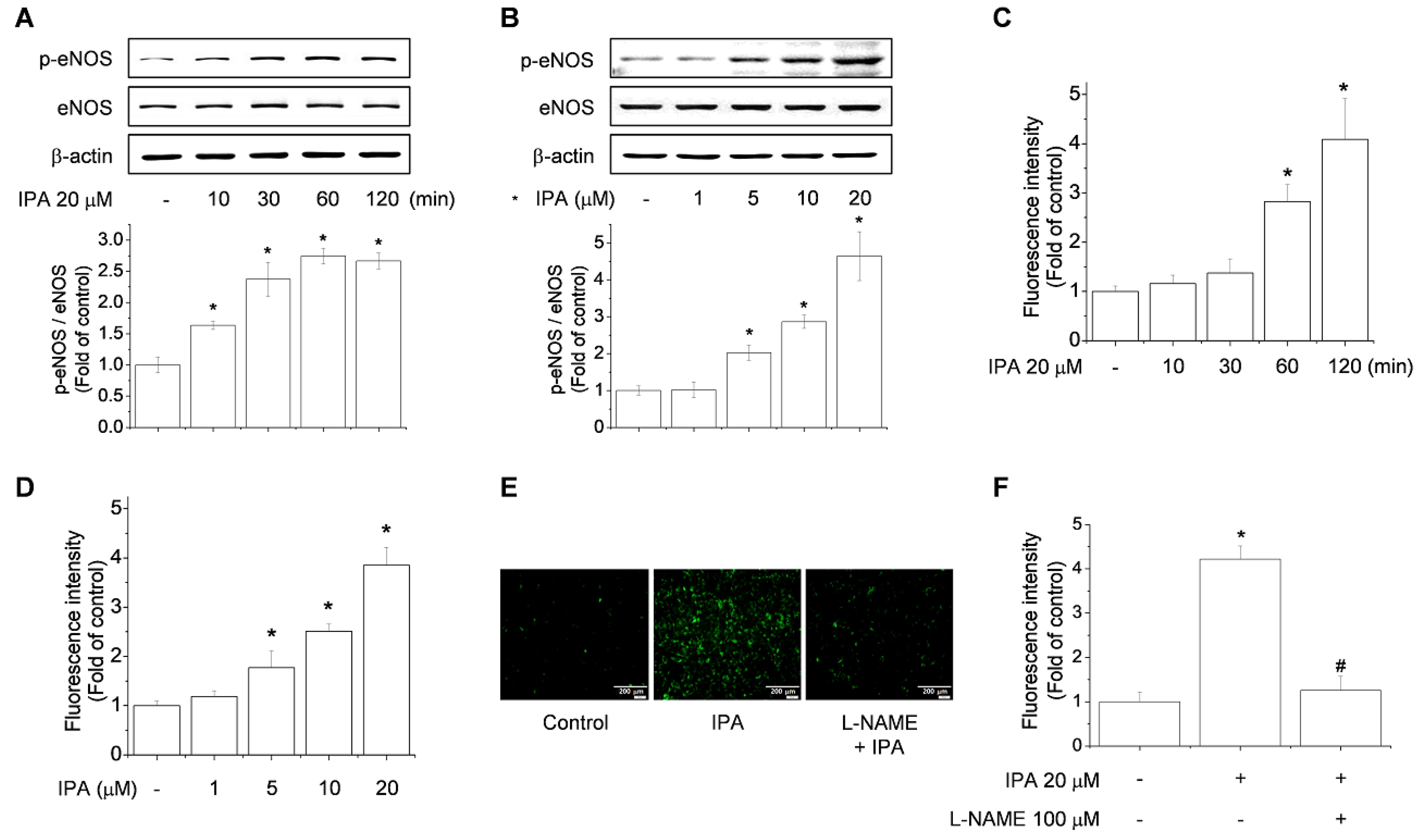
2.2. IPA Increases eNOS Phosphorylation and NO Production in Endothelial Cells
2.3. AMPK and CaMKII Are Required for IPA-Induced eNOS Phosphorylation and NO Production
2.4. Role of Akt and MAPKs in IPA-Induced eNOS Phosphorylation and NO Production
2.5. Effects of IPA on the Inhibition of NF-κB Activation, ICAM-1 Expression, and Monocyte Adhesion in ECs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Treatment
4.3. Measurement of Cell Cytotoxicity
4.4. Protein Extraction and Western Blotting
4.5. Quantification of NO
4.6. Transient Transfection and Luciferase Reporter Assay
4.7. Immunofluorescence Staining
4.8. Cell–Cell Adhesion Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| CaM | Calmodulin |
| CaMKII | Ca2+/calmodulin-dependent kinase II |
| CC | Compound C |
| eNOS | Endothelial nitric oxide synthase |
| IPA | Impressic acid |
| MAPK | Mitogen-activated protein kinase |
| NO | Nitric oxide |
References
- Lee, K.S.; Kim, J.; Kwak, S.N.; Lee, K.S.; Lee, D.K.; Ha, K.S.; Won, M.H.; Jeoung, D.; Lee, H.; Kwon, Y.G.; et al. Functional role of NF-κB in expression of human endothelial nitric oxide synthase. Biochem. Biophys. Res. Commun. 2014, 448, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [PubMed]
- Sessa, W.C. eNOS at a glance. J. Cell. Sci. 2004, 117, 2427–2429. [Google Scholar] [CrossRef] [PubMed]
- Stuehr, D.J. Mammalian nitric oxide synthases. Biochim. Biophys. Acta 1999, 1411, 217–230. [Google Scholar] [CrossRef]
- Fulton, D.; Ruan, L.; Sood, S.G.; Li, C.; Zhang, Q.; Venema, R.C. Agonist-stimulated endothelial nitric oxide synthase activation and vascular relaxation. Role of eNOS phosphorylation at Tyr83. Circ. Res. 2008, 102, 497–504. [Google Scholar] [CrossRef]
- Sriram, K.; Laughlin, J.G.; Rangamani, P.; Tartakovsky, D.M. Shear-Induced Nitric Oxide Production by Endothelial Cells. Biophys. J. 2016, 111, 208–221. [Google Scholar] [CrossRef]
- Morrow, V.A.; Foufelle, F.; Connell, J.M.; Petrie, J.R.; Gould, G.W.; Salt, I.P. Direct activation of AMP-activated protein kinase stimulates nitric-oxide synthesis in human aortic endothelial cells. J. Biol. Chem. 2003, 278, 31629–31639. [Google Scholar] [CrossRef]
- Wu, K.K. Regulation of endothelial nitric oxide synthase activity and gene expression. Ann. N. Y. Acad. Sci. 2002, 962, 122–130. [Google Scholar] [CrossRef]
- Boo, Y.C.; Hwang, J.; Sykes, M.; Michell, B.J.; Kemp, B.E.; Lum, H.; Jo, H. Shear stress stimulates phosphorylation of eNOS at Ser(635) by a protein kinase A-dependent mechanism. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1819–H1828. [Google Scholar] [CrossRef]
- Michell, B.J.; Chen, Z.P.; Tiganis, T.; Stapleton, D.; Katsis, F.; Power, D.A.; Sim, A.T.; Kemp, B.E. Coordinated control of endothelial nitric-oxide synthase phosphorylation by protein kinase C and the cAMP-dependent protein kinase. J. Biol. Chem. 2001, 276, 17625–17628. [Google Scholar] [CrossRef]
- Fulton, D.; Gratton, J.P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 1999, 399, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Ackah, E.; Yu, J.; Zoellner, S.; Iwakiri, Y.; Skurk, C.; Shibata, R.; Ouchi, N.; Easton, R.M.; Galasso, G.; Birnbaum, M.J.; et al. Akt1/protein kinase Balpha is critical for ischemic and VEGF-mediated angiogenesis. J. Clin. Investig. 2005, 115, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.R.; Brunet, A.; Greenberg, M.E. Cellular survival: A play in three Akts. Genes Dev. 1999, 13, 2905–2927. [Google Scholar] [CrossRef] [PubMed]
- Shiojima, I.; Walsh, K. Role of Akt signaling in vascular homeostasis and angiogenesis. Circ. Res. 2002, 90, 1243–1250. [Google Scholar] [CrossRef]
- Yao, R.; Cooper, G.M. Requirement for phosphatidylinositol-3 kinase in the prevention of apoptosis by nerve growth factor. Science 1995, 267, 2003–2006. [Google Scholar] [CrossRef]
- Srivastava, S.K. A New Triterpenic Acid from Schefflera impressa. J. Nat. Prod. 1992, 55, 298–302. [Google Scholar] [CrossRef]
- Park, S.H.; Nhiem, N.X.; Kiem, P.V.; Choi, E.M.; Kim, J.A.; Kim, Y.H. A new norlupane triterpene from the leaves of Acanthopanax koreanum increases the differentiation of osteoblastic MC3T3-e1 cells. Arch. Pharm. Res. 2010, 33, 75–80. [Google Scholar] [CrossRef]
- Kim, J.A.; Yang, S.Y.; Koo, J.E.; Koh, Y.S.; Kim, Y.H. Lupane-type triterpenoids from the steamed leaves of Acanthopanax koreanum and their inhibitory effects on the LPS-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Bioorg. Med. Chem. Lett. 2010, 20, 6703–6707. [Google Scholar] [CrossRef]
- Lim, H.; Min, D.S.; Yun, H.E.; Kim, K.T.; Sun, Y.N.; Dat, L.D.; Kim, Y.H.; Kim, H.P. Impressic acid from Acanthopanax koreanum, possesses matrix metalloproteinase-13 down-regulating capacity and protects cartilage destruction. J. Ethnopharmacol. 2017, 209, 73–81. [Google Scholar] [CrossRef]
- Kim, J.A.; Yang, S.Y.; Song, S.B.; Kim, Y.H. Effects of impressic acid from Acanthopanax koreanum on NF-κB and PPARγ activities. Arch. Pharm. Res. 2011, 34, 1347–1351. [Google Scholar] [CrossRef]
- Fulton, D.; Fontana, J.; Sowa, G.; Gratton, J.P.; Lin, M.; Li, K.X.; Michell, B.; Kemp, B.E.; Rodman, D.; Sessa, W.C. Localization of endothelial nitric-oxide synthase phosphorylated on serine 1179 and nitric oxide in Golgi and plasma membrane defines the existence of two pools of active enzyme. J. Biol. Chem. 2002, 277, 4277–4284. [Google Scholar] [CrossRef] [PubMed]
- Arad, M.; Seidman, C.E.; Seidman, J.G. AMP-activated protein kinase in the heart: Role during health and disease. Circ. Res. 2007, 100, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.P.; Mitchelhill, K.I.; Michell, B.J.; Stapleton, D.; Rodriguez-Crespo, I.; Witters, L.A.; Power, D.A.; Ortiz de Montellano, P.R.; Kemp, B.E. AMP-activated protein kinase phosphorylation of endothelial NO synthase. FEBS Lett. 1999, 443, 285–289. [Google Scholar] [CrossRef]
- Chen, H.; Montagnani, M.; Funahashi, T.; Shimomura, I.; Quon, M.J. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J. Biol. Chem. 2003, 278, 45021–45026. [Google Scholar] [CrossRef]
- Thors, B.; Halldórsson, H.; Thorgeirsson, G. Thrombin and histamine stimulate endothelial nitric-oxide synthase phosphorylation at Ser1177 via an AMPK mediated pathway independent of PI3K-Akt. FEBS Lett. 2004, 573, 175–180. [Google Scholar] [CrossRef]
- Fleming, I.; Fisslthaler, B.; Dimmeler, S.; Kemp, B.E.; Busse, R. Phosphorylation of Thr495 regulates Ca2+/calmodulin-dependent endothelial nitric oxide synthase activity. Circ. Res. 2001, 88, E68–E75. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nemoto, S.; Ishida, K.; Taguchi, K.; Matsumoto, T.; Kamata, K. Involvement of CaM kinase II in the impairment of endothelial function and eNOS activity in aortas of Type 2 diabetic rats. Clin. Sci. 2012, 123, 375–386. [Google Scholar] [CrossRef]
- Dimmeler, S.; Fleming, I.; Fisslthaler, B.; Hermann, C.; Busse, R.; Zeiher, A.M. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999, 399, 601–605. [Google Scholar] [CrossRef]
- Hurt, K.J.; Musicki, B.; Palese, M.A.; Crone, J.K.; Becker, R.E.; Moriarity, J.L.; Snyder, S.H.; Burnett, A.L. Akt-dependent phosphorylation of endothelial nitric-oxide synthase mediates penile erection. Proc. Natl. Acad. Sci. USA 2002, 99, 4061–4066. [Google Scholar] [CrossRef]
- Grossini, E.; Molinari, C.; Mary, D.A.; Uberti, F.; Caimmi, P.P.; Surico, N.; Vacca, G. Intracoronary genistein acutely increases coronary blood flow in anesthetized pigs through beta-adrenergic mediated nitric oxide release and estrogenic receptors. Endocrinology 2008, 149, 2678–2687. [Google Scholar] [CrossRef]
- Merla, R.; Ye, Y.; Lin, Y.; Manickavasagam, S.; Huang, M.H.; Perez-Polo, R.J.; Uretsky, B.F.; Birnbaum, Y. The central role of adenosine in statin-induced ERK1/2, Akt, and eNOS phosphorylation. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1918–H1928. [Google Scholar] [CrossRef] [PubMed]
- Heike, L.P. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogenes 1999, 18, 6853–6866. [Google Scholar]
- Su, J.B. Vascular endothelial dysfunction and pharmacological treatment. World J. Cardiol. 2015, 7, 719–741. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.F.; Lee, I.S.; Shen, G.; Dat, N.T.; Lee, J.J.; Kim, Y.H. Triterpenoids from Acanthopanax koreanum root and their inhibitory activities on NFAT transcription. Arch. Pharm. Res. 2004, 27, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Fleming, I. Molecular mechanisms underlying the activation of eNOS. Pflugers Arch. 2010, 459, 793–806. [Google Scholar] [CrossRef]
- Hien, T.T.; Kim, N.D.; Pokharel, Y.R.; Oh, S.J.; Lee, M.Y.; Kang, K.W. Ginsenoside Rg3 increases nitric oxide production via increases in phosphorylation and expression of endothelial nitric oxide synthase: Essential roles of estrogen receptor-dependent PI3-kinase and AMP-activated protein kinase. Toxicol. Appl. Pharmacol. 2010, 246, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.C.; El Kebir, D.; Chéreau, C.; Lanone, S.; Huang, X.L.; De Buys Roessingh, A.S.; Mercier, J.C.; Dall’Ava-Santucci, J.; Dinh-Xuan, A.T. Involvement of Ca2+/calmodulin-dependent protein kinase II in endothelial NO production and endothelium-dependent relaxation. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2311–H2319. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.W.; Choi, C.Y.; Hwang, Y.P.; Kim, H.G.; Kim, S.J.; Chung, Y.C.; Lee, K.J.; Jeong, T.C.; Jeong, H.G. Betulinic Acid Increases eNOS Phosphorylation and NO Synthesis via the Calcium-Signaling Pathway. J. Agric. Food Chem. 2016, 64, 785–791. [Google Scholar] [CrossRef]
- Zhang, Q.J.; McMillin, S.L.; Tanner, J.M.; Palionyte, M.; Abel, E.D.; Symons, J.D. Endothelial nitric oxide synthase phosphorylation in treadmill-running mice: Role of vascular signalling kinases. J. Physiol. 2009, 587, 3911–3920. [Google Scholar] [CrossRef]
- Peng, X.Q.; Damarla, M.; Skirball, J.; Nonas, S.; Wang, X.Y.; Han, E.J.; Hasan, E.J.; Cao, X.; Boueiz, A.; Damico, R.; et al. Protective role of PI3-kinase/Akt/eNOS signaling in mechanical stress through inhibition of p38 mitogen-activated protein kinase in mouse lung. Acta Pharmacol. Sin. 2010, 31, 175–183. [Google Scholar] [CrossRef]
- Yu, J.; de Muinck, E.D.; Zhuang, Z.; Drinane, M.; Kauser, K.; Rubanyi, G.M.; Qian, H.S.; Murata, T.; Escalante, B.; Sessa, W.C. Endothelial nitric oxide synthase is critical for ischemic remodeling, mural cell recruitment, and blood flow reserve. Proc. Natl. Acad. Sci. USA 2005, 102, 10999–11004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridnour, L.A.; Isenberg, J.S.; Espey, M.G.; Thomas, D.D.; Roberts, D.D.; Wink, D.A. Nitric oxide regulates angiogenesis through a functional switch involving thrombospondin-1. Proc. Natl. Acad. Sci. USA 2005, 102, 13147–13152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salerno, J.C.; Ghosh, D.K.; Razdan, R.; Helms, K.A.; Brown, C.C.; McMurry, J.L.; Rye, E.A.; Chrestensen, C.A. Endothelial nitric oxide synthase is regulated by ERK phosphorylation at Ser602. Biosci. Rep. 2014, 34, e00137. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wu, L.J.; Tashiro, S.I.; Onodera, S.; Ikejima, T. Nitric oxide activated by p38 and NF-kappaB facilitates apoptosis and cell cycle arrest under oxidative stress in evodiamine-treated human melanoma A375-S2 cells. Free Radic. Res. 2008, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Jung, T.W.; Hong, H.C.; Choi, H.Y.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Choi, K.M.; Choi, D.S.; Baik, S.H.; et al. Progranulin protects vascular endothelium against atherosclerotic inflammatory reaction via Akt/eNOS and nuclear factor-κB pathways. PLoS ONE 2013, 8, e76679. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Qiu, X.M.; Hao, Q.; Li, D.J. Anti-inflammatory effects of a Chinese herbal medicine in atherosclerosis via estrogen receptor β mediating nitric oxide production and NF-κB suppression in endothelial cells. Cell Death Dis. 2013, 4, e551. [Google Scholar] [CrossRef]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharmacol. 2009, 78, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Bath, P.M.; Hassall, D.G.; Gladwin, A.M.; Palmer, R.M.; Martin, J.F. Nitric oxide and prostacyclin. Divergence of inhibitory effects on monocyte chemotaxis and adhesion to endothelium in vitro. Arterioscler. Thromb. 1991, 11, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ma, J.; Wang, K.S.; Mi, C.; Wang, Z.; Piao, L.X.; Xu, G.H.; Li, X.; Lee, J.J.; Jin, X. Baicalein inhibits TNF-α-induced NF-κB activation and expression of NF-κB-regulated target gene products. Oncol. Rep. 2016, 36, 2771–2776. [Google Scholar] [CrossRef] [Green Version]
- Kojima, H.; Nakatsubo, N.; Kikuchi, K.; Kawahara, S.; Kirino, Y.; Nagoshi, H.; Hirata, Y.; Nagano, T. Detection and imaging of nitric oxide with novel fluorescent indicators: Diaminofluoresceins. Anal. Chem. 1998, 70, 2446–2453. [Google Scholar] [CrossRef]
- Goetz, R.M.; Thatte, H.S.; Prabhakar, P.; Cho, M.R.; Michel, T.; Golan, D.E. Estradiol induces the calcium-dependent translocation of endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1999, 96, 2788–2793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nethi, S.K.; Veeriah, V.; Barui, A.K.; Rajendran, S.; Mattapally, S.; Misra, S.; Chatterjee, S.; Patra, C.R. Investigation of molecular mechanisms and regulatory pathways of pro-angiogenic nanorods. Nanoscale 2015, 7, 9760–9770. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.W.; Pham, H.T.; Choi, J.H.; Lee, G.H.; Han, E.H.; Cho, Y.H.; Chung, Y.C.; Kim, Y.H.; Jeong, H.G. Impressic Acid, a Lupane-Type Triterpenoid from Acanthopanax koreanum, Attenuates TNF-α-Induced Endothelial Dysfunction via Activation of eNOS/NO Pathway. Int. J. Mol. Sci. 2019, 20, 5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225772
Jin SW, Pham HT, Choi JH, Lee GH, Han EH, Cho YH, Chung YC, Kim YH, Jeong HG. Impressic Acid, a Lupane-Type Triterpenoid from Acanthopanax koreanum, Attenuates TNF-α-Induced Endothelial Dysfunction via Activation of eNOS/NO Pathway. International Journal of Molecular Sciences. 2019; 20(22):5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225772
Chicago/Turabian StyleJin, Sun Woo, Hoa Thi Pham, Jae Ho Choi, Gi Ho Lee, Eun Hee Han, Young Ho Cho, Young Chul Chung, Young Ho Kim, and Hye Gwang Jeong. 2019. "Impressic Acid, a Lupane-Type Triterpenoid from Acanthopanax koreanum, Attenuates TNF-α-Induced Endothelial Dysfunction via Activation of eNOS/NO Pathway" International Journal of Molecular Sciences 20, no. 22: 5772. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225772