Genome-Wide Identification and Characterization of the Mitochondrial Transcription Termination Factors (mTERFs) in Capsicum annuum L.

Abstract
:1. Introduction
2. Results
2.1. Basic Characterization of CamTERFs and Comparison between Available Capsicum Genome Databases
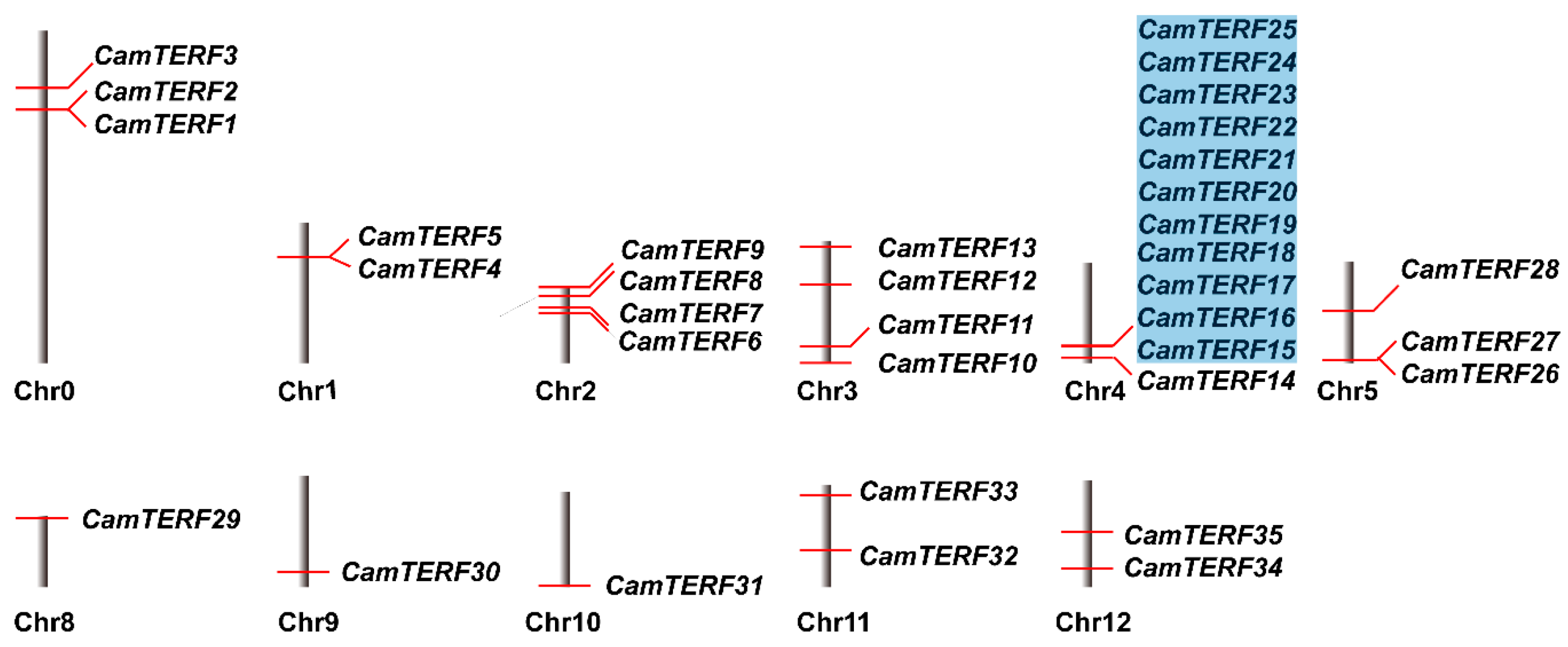
2.2. Analysis of CamTERFs Chromosomal Location and Subcellular Localization
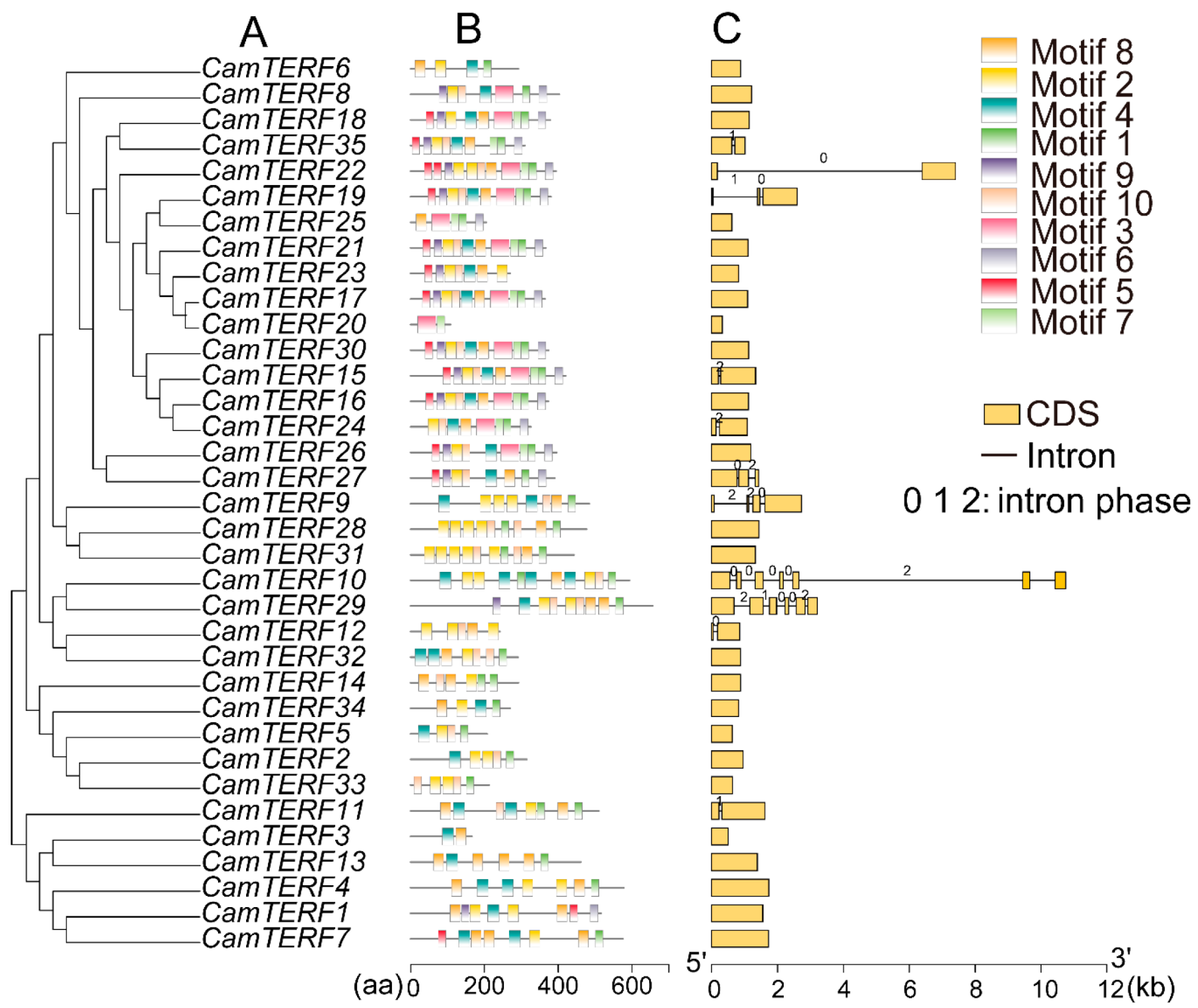
2.3. Phylogenetic Relationships, Gene Structures, and Motifs of CamTERFs
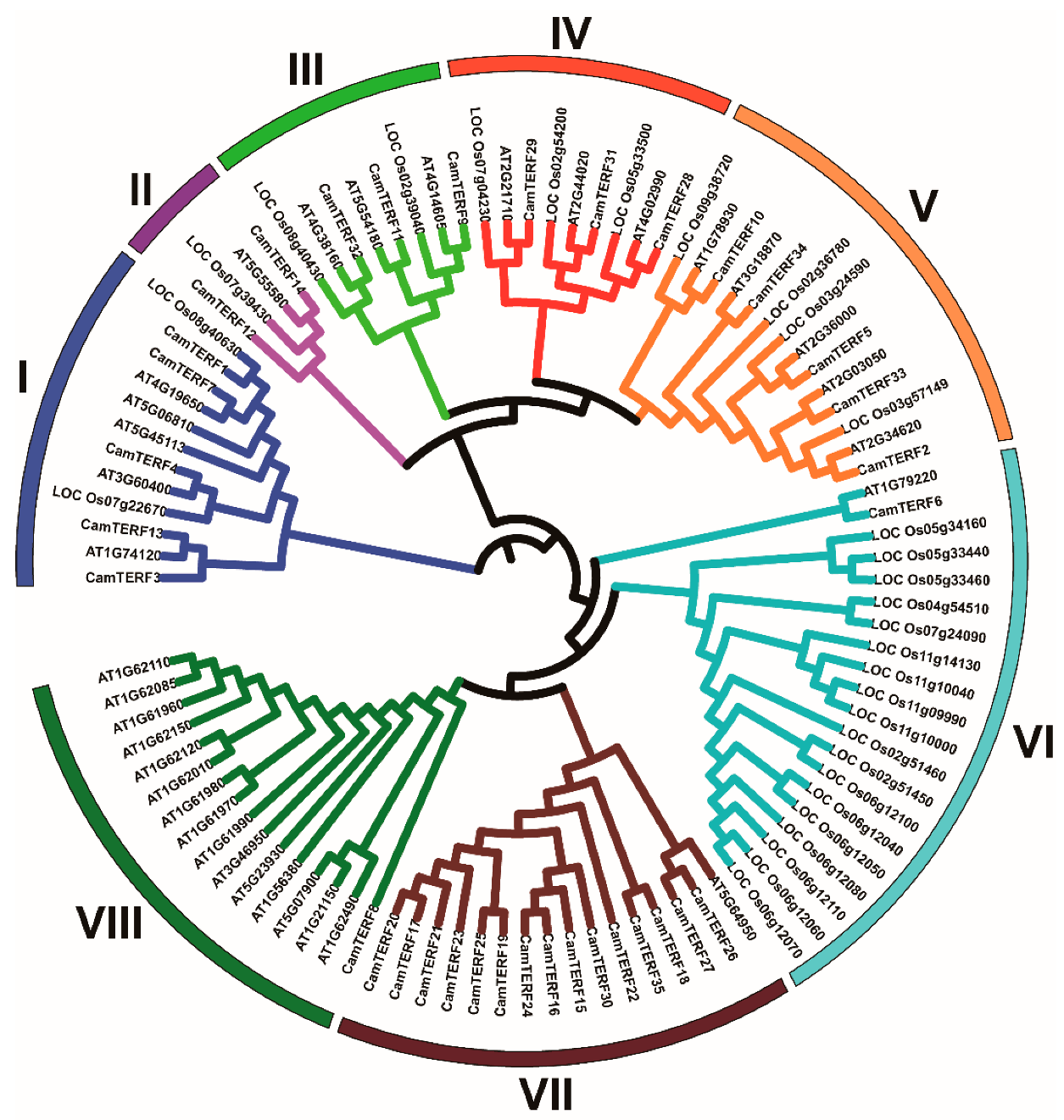
2.4. Homologous Relationships among mTERFs of Capsicum, Rice, and Arabidopsis
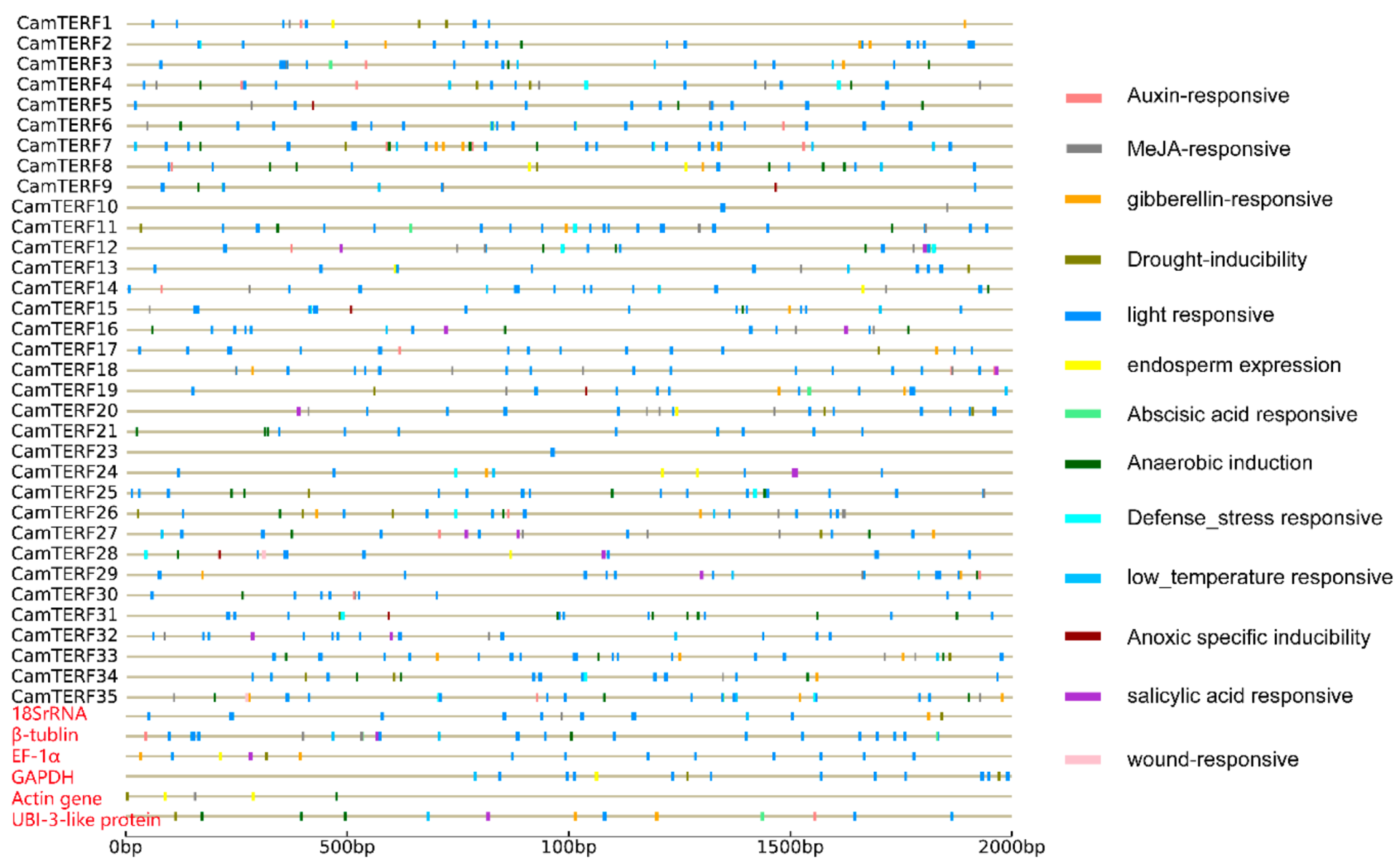
2.5. Analysis of CamTERF Promoter cis-Elements
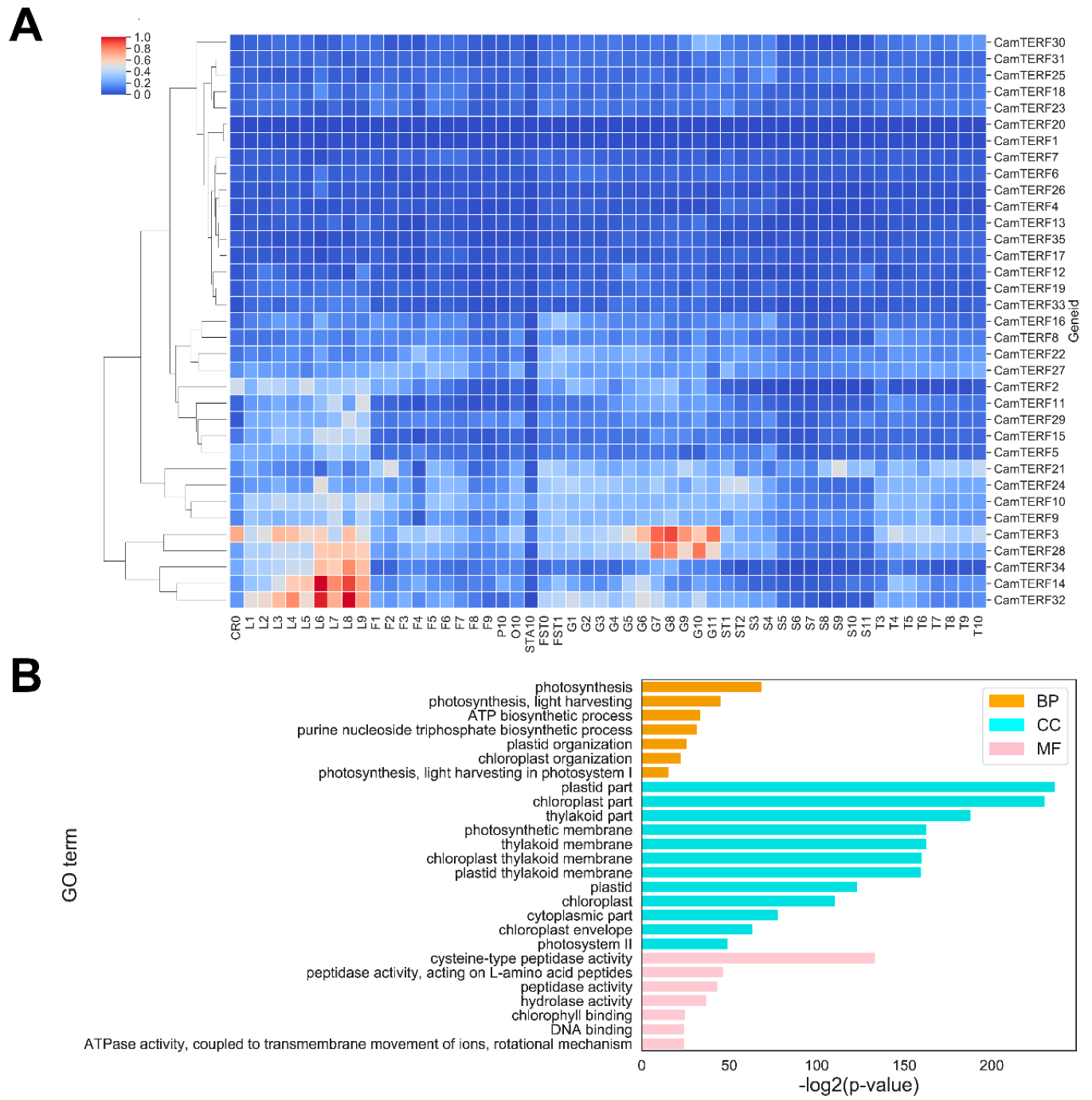
2.6. Analysis of CamTERFs Expression Patterns and GO Enrichment Analysis of CamTERFs Co-Expressing Genes
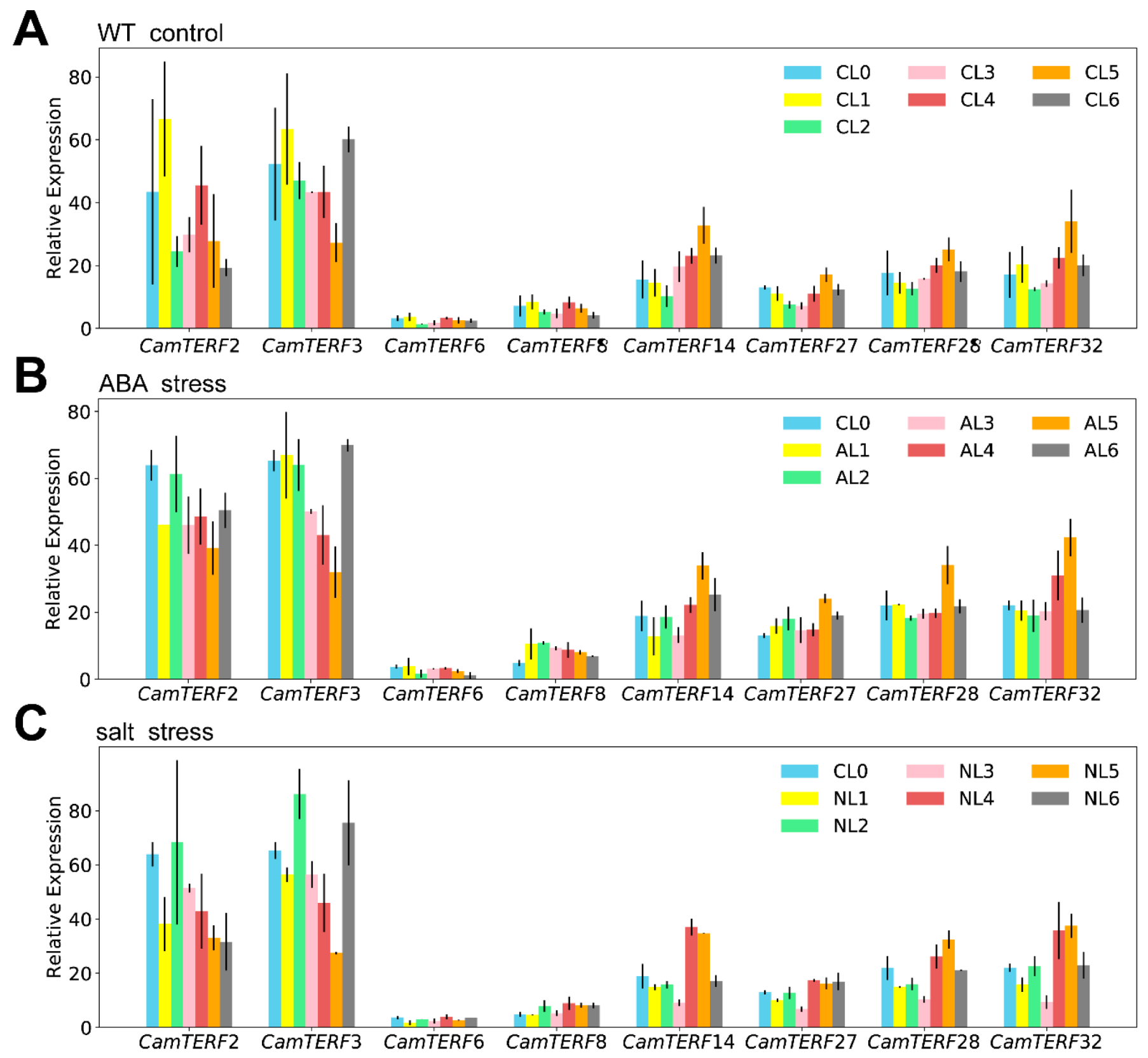
2.7. Analysis of the Expression of CamTERFs under Salt Stress and ABA Treatment
3. Discussion
4. Materials and Methods
4.1. Capsicum mTERF Family Identification
4.2. Basic Characterization and Homology of CamTERFs
4.3. Analysis of Chromosomal Location and Subcellular Localization
4.4. Analysis of Gene Structure, Phylogeny, and Conserved Motifs
4.5. Analysis of Homologous Evolutionary Relationships between mTERFs of Rice, Arabidopsis, and Capsicum
4.6. Analysis of Cis-Regulatory Elements in CamTERFs
4.7. Analysis of CamTERFs Expression and GO Enrichment for CamTERF Co-Expressing Members
4.8. ABA and Salt-Stress Treatment
4.9. cDNA Generation and the Quantitative/Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| mTERFs | Mitochondrial transcription ter |
| CamTERFs | Capsicum annuum Mitochondr |
| FPKM | Fragments Per Kilobase Million |
| GO | Gene Ontology |
| MeJA | Methyl Jasmonate |
| BP | Biological Process |
| MF | Molecular Function |
| CC | Cellular Component |
| RNA-Seq | RNA sequencing; |
| mtDNA | Mitochondrial DNA |
| ABA | Abscisic acid |
| RT-qPCR | Reverse transcription quantitative PCR |
References
- Gray, M.W.; Burger, G.; Lang, B.F. Mitochondrial evolution. Science 1999, 283, 1476–1481. [Google Scholar] [CrossRef] [Green Version]
- Lang, B.F.; Gray, M.W.; Burger, G. Mitochondrial genome evolution and the origin of eukaryotes. Annu. Rev. Genet. 1999, 33, 351–397. [Google Scholar] [CrossRef] [PubMed]
- Liere, K.; Weihe, A.; Börner, T. The transcription machineries of plant mitochondria and chloroplasts: Composition, function, and regulation. J. Plant Physiol. 2011, 168, 1345–1360. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Silva, P.; Martinez-Azorin, F.; Micol, V.; Attardi, G. The human mitochondrial transcription termination factor (mTERF) is a multizipper protein but binds to DNA as a monomer, with evidence pointing to intramolecular leucine zipper interactions. EMBO J. 1997, 16, 1066–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberti, M.; Polosa, P.L.; Bruni, F.; Manzari, C.; Deceglie, S.; Gadaleta, M.N.; Cantatore, P. The MTERF family proteins: Mitochondrial transcription regulators and beyond. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2009, 1787, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Linder, T.; Park, C.B.; Asin-Cayuela, J.; Pellegrini, M.; Larsson, N.G.; Falkenberg, M.; Gustafsson, C.M. A family of putative transcription termination factors shared amongst metazoans and plants. Curr. Genet. 2005, 48, 265–269. [Google Scholar] [CrossRef]
- Kruse, B.; Narasimhan, N.; Attardi, G. Termination of transcription in human mitochondria: Identification and purification of a DNA binding protein factor that promotes termination. Cell 1989, 58, 391–397. [Google Scholar] [CrossRef]
- Asin-Cayuela, J.; Schwend, T.; Farge, G.; Gustafsson, C.M. The human mitochondrial transcription termination factor (mTERF) is fully active in vitro in the non-phosphorylated form. J. Biol. Chem. 2005, 280, 25499–25505. [Google Scholar] [CrossRef] [Green Version]
- Park, C.B.; Asin-Cayuela, J.; Cámara, Y.; Shi, Y.; Pellegrini, M.; Gaspari, M.; Falkenberg, M. MTERF3 is a negative regulator of mammalian mtDNA transcription. Cell 2007, 130, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Cámara, Y.; Asin-Cayuela, J.; Park, C.B.; Metodiev, M.D.; Shi, Y.; Ruzzenente, B.; Franz, T. MTERF4 regulates translation by targeting the methyltransferase NSUN4 to the mammalian mitochondrial ribosome. Cell Metab. 2011, 13, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Wenz, T.; Luca, C.; Torraco, A.; Moraes, C.T. RETRACTED: mTERF2 Regulates Oxidative Phosphorylation by Modulating mtDNA Transcription; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Meskauskiene, R.; Würsch, M.; Laloi, C.; Vidi, P.A.; Coll, N.S.; Kessler, F.; Apel, K. A mutation in the Arabidopsis mTERF-related plastid protein SOLDAT10 activates retrograde signaling and suppresses 1O2-induced cell death. Plant J. 2009, 60, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.V.; Toepel, J.; Burgess, S.; Uhmeyer, A.; Blifernez, O.; Doebbe, A.; Kruse, O. Time-course global expression profiles of Chlamydomonas reinhardtii during photo-biological H2 production. PLoS ONE 2011, 6, e29364. [Google Scholar] [CrossRef] [Green Version]
- Wobbe, L.; Nixon, P.J. The mTERF protein MOC1 terminates mitochondrial DNA transcription in the unicellular green alga Chlamydomonas reinhardtii. Nucleic Acids Res. 2013, 41, 6553–6567. [Google Scholar] [CrossRef]
- Babiychuk, E.; Vandepoele, K.; Wissing, J.; Garcia-Diaz, M.; De Rycke, R.; Akbari, H.; Van Montagu, M.C. Plastid gene expression and plant development require a plastidic protein of the mitochondrial transcription termination factor family. Proc. Natl. Acad. Sci. USA 2011, 108, 6674–6679. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, P.; Riano-Pachon, D.M.; Corrêa, L.G.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: Updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2009, 38 (Suppl. 1), D822–D827. [Google Scholar]
- Kleine, T. Arabidopsis thaliana mTERF proteins: Evolution and functional classification. Front. Plant Sci. 2012, 3, 233. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Cai, M.; Zhang, X.; Li, Y.; Zhang, J.; Zhao, H.; Qiu, F. Genome-wide identification, evolution and expression analysis of mTERF gene family in maize. PLoS ONE 2014, 9, e94126. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Leister, D.; Kleine, T. Arabidopsis thaliana mTERF10 and mTERF11, but not mTERF12, are involved in the response to salt stress. Front. Plant Sci. 2017, 8, 1213. [Google Scholar] [CrossRef] [Green Version]
- Robles, P.; Micol, J.L.; Quesada, V. Arabidopsis MDA1, a nuclear-encoded protein, functions in chloroplast development and abiotic stress responses. PLoS ONE 2012, 7, e42924. [Google Scholar] [CrossRef]
- Huang, S.Y.; Qiu, S.J.; Chen, J.Y.; Chen, J.P.; Li, X.Y.; Li, L. Advances of mTERF Protein Family. Life Sci. Res. 2016, 20, 455–459. [Google Scholar]
- Romani, I.; Manavski, N.; Morosetti, A.; Tadini, L.; Maier, S.; Kühn, K.; Kleine, T. A member of the arabidopsis mitochondrial transcription termination factor family is required for maturation of chloroplast transfer RNAIle (GAU). Plant Physiol. 2015, 169, 627–646. [Google Scholar] [CrossRef] [Green Version]
- Robles, P.; Núñez-Delegido, E.; Ferrández-Ayela, A.; Sarmiento-Mañús, R.; Micol, J.L.; Quesada, V. Arabidopsis mTERF6 is required for leaf patterning. Plant Sci. 2018, 266, 117–129. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.L.; Zhang, X.L.; Yu, Q.B.; Wang, X.; Yuan, X.B.; Yang, Z.N. A nuclear-encoded protein, mTERF6, mediates transcription termination of rpoA polycistron for plastid-encoded RNA polymerase-dependent chloroplast gene expression and chloroplast development. Sci. Rep. 2018, 8, 11929. [Google Scholar] [CrossRef]
- Pfalz, J.; Liere, K.; Kandlbinder, A.; Dietz, K.J.; Oelmüller, R. pTAC2,-6, and-12 are components of the transcriptionally active plastid chromosome that are required for plastid gene expression. Plant Cell 2006, 18, 176–197. [Google Scholar] [CrossRef] [Green Version]
- Xiong, H.B.; Wang, J.; Huang, C.; Rochaix, J.D.; Lin, F.M.; Zhang, J.X.; Yang, Z.N. mTERF8, a Member of the Mitochondrial Transcription Termination Factor Family, is Involved in the Transcription Termination of Chloroplast Gene psbJ1. Plant Physiol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Quesada, V.; Sarmiento-Mañús, R.; González-Bayón, R.; Hricová, A.; Pérez-Marcos, R.; Graciá-Martínez, E.; Micol, J.L. Arabidopsis RUGOSA2 encodes an mTERF family member required for mitochondrion, chloroplast and leaf development. Plant J. 2011, 68, 738–753. [Google Scholar] [CrossRef]
- Hammani, K.; Barkan, A. An mTERF domain protein functions in group II intron splicing in maize chloroplasts. Nucleic Acids Res. 2014, 42, 5033–5042. [Google Scholar] [CrossRef] [Green Version]
- Robles, P.; Micol, J.L.; Quesada, V. Mutations in the plant-conserved MTERF9 alter chloroplast gene expression, development and tolerance to abiotic stress in Arabidopsis thaliana. Physiol. Plant. 2015, 154, 297–313. [Google Scholar] [CrossRef]
- Hsu, Y.W.; Wang, H.J.; Hsieh, M.H.; Hsieh, H.L.; Jauh, G.Y. Arabidopsis mTERF15 is required for mitochondrial nad2 intron 3 splicing and functional complex I activity. PLoS ONE 2014, 9, e112360. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Lee, U.; Small, I.; des Francs-Small, C.C.; Vierling, E. Mutations in an Arabidopsis mitochondrial transcription termination factor–related protein enhance thermotolerance in the absence of the major molecular chaperone HSP101. Plant Cell 2012, 24, 3349–3365. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Jung, K. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Yang, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Pang, X.; Wan, H.; Ahammed, G.J.; Yu, J.; Yao, Z.; Yang, Y. Identification of optimal reference genes for normalization of qPCR analysis during pepper fruit development. Front. Plant Sci. 2017, 8, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Zhang, Y.; Hu, Z.; Huang, X.; Zhang, B.; Lu, Q.; Lu, C. mTERF5 Acts as a Transcriptional Pausing Factor to Positively Regulate Transcription of Chloroplast psbEFLJ. Mol. Plant 2019. [Google Scholar] [CrossRef]
- Xu, Q.; Dunbrack, R.L., Jr. Assignment of protein sequences to existing domain and family classification systems: Pfam and the PDB. Bioinformatics 2012, 28, 2763–2772. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Sonnhammer, E.L. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39 (Suppl. 2), W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Zheng, Y.; Jiao, C.; Sun, H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.; Zhao, P.X. iTAK: A program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2013, 42, D1182–D1187. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Sinharoy, S.; Udvardi, M.; Zhao, P.X. PlantTFcat: An online plant transcription factor and transcriptional regulator categorization and analysis tool. BMC Bioinform. 2013, 14, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Grosdidier, A. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; Von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various biological data handling tools with a user-friendly interface. bioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, H.; Deng, Y.; Zheng, J.; Liu, M.; Ou, L.; He, S. PepperHub, an informatics hub for the chili pepper research community. Mol. Plant 2017, 10, 1129–1132. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experimentswith HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Klopfenstein, D.V.; Zhang, L.; Pedersen, B.S.; Ramírez, F.; Vesztrocy, A.W.; Naldi, A.; Dampier, W. GOATOOLS: A Python library for Gene Ontology analyses. Sci. Rep. 2018, 8, 10872. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Y.; Bachmann, D.; La, H.; Ma, Z.; Venkatesh, P.N.; Ramamoorthy, R.; Ramachandran, S. Ds insertion mutagenesis as an efficient tool to produce diverse variations for rice breeding. Plant Mol. Biol. 2007, 65, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
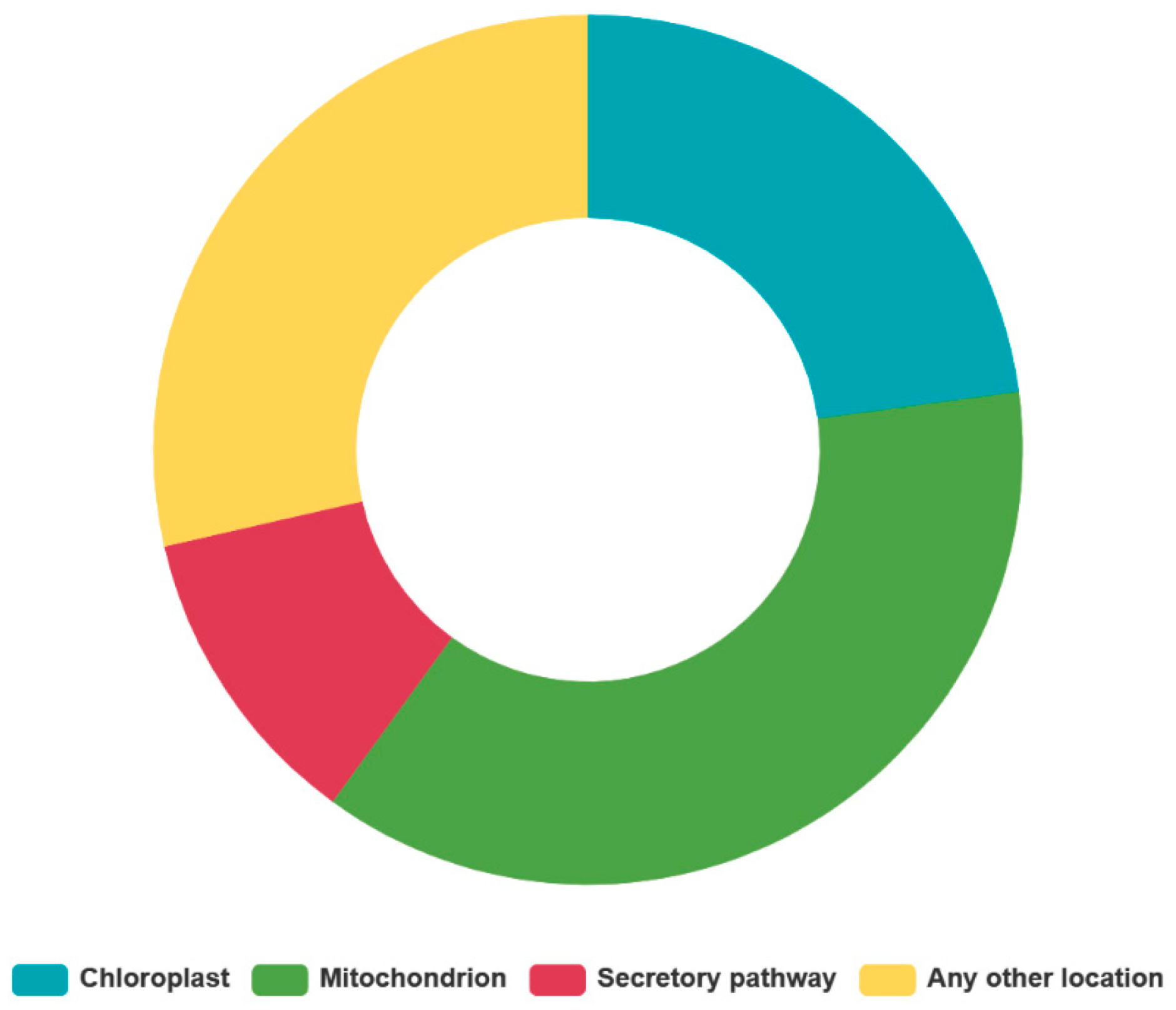{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Accession No. | CM334 Homelog | E-Value | Chr | Start | End | ORF bp | Size aa | Protein Molecular Weight/KD | PI |
|---|---|---|---|---|---|---|---|---|---|---|
| CamTERF1 | Capana00g003386 | CA00g83930 | 0 | Chr00 | 544847272 | 544848822 | 1551 | 516 | 59.52 | 9.24 |
| CamTERF2 | Capana00g003424 | CA01g01340 | 0 | Chr00 | 545339784 | 545340731 | 948 | 315 | 35.86 | 9.3 |
| CamTERF3 | Capana00g003922 | CA04g12790 | 1.25 × 10−12 | Chr00 | 591637478 | 591637978 | 501 | 166 | 19.33 | 7.6 |
| CamTERF4 | Capana01g003479 | CA01g27850 | 0 | Chr01 | 227806286 | 227808022 | 1737 | 578 | 66.29 | 8.02 |
| CamTERF5 | Capana01g003663 | CA00g07130 | 6.03 × 10−41 | Chr01 | 232164334 | 232164957 | 624 | 207 | 24.25 | 9.13 |
| CamTERF6 | Capana02g001023 | CA02g09080 | 0 | Chr02 | 107229127 | 107230005 | 879 | 292 | 33.69 | 9.73 |
| CamTERF7 | Capana02g001304 | CA02g00270 | 0 | Chr02 | 119529666 | 119531393 | 1728 | 575 | 65.45 | 8.98 |
| CamTERF8 | Capana02g002368 | CA02g19520 | 0 | Chr02 | 143793081 | 143794292 | 1212 | 403 | 45.81 | 9.87 |
| CamTERF9 | Capana02g003623 | CA02g30630 | 1.22 × 10−17 | Chr02 | 163004467 | 163007189 | 1455 | 484 | 54.96 | 9.2 |
| CamTERF10 | Capana03g000004 | CA03g36450 | 4.29 × 10−69 | Chr03 | 95975 | 106725 | 1782 | 593 | 67.82 | 9.18 |
| CamTERF11 | Capana03g001822 | CA03g21280 | 0 | Chr03 | 36374556 | 36376173 | 1533 | 510 | 57.4 | 8.83 |
| CamTERF12 | Capana03g003090 | CA03g11980 | 1.51 × 10−17 | Chr03 | 168574645 | 168575497 | 729 | 242 | 28.26 | 9.22 |
| CamTERF13 | Capana03g004299 | CA03g04110 | 0 | Chr03 | 250309314 | 250310699 | 1386 | 461 | 53.08 | 9.25 |
| CamTERF14 | Capana04g000706 | CA04g17520 | 0 | Chr04 | 12169693 | 12170571 | 879 | 292 | 33.97 | 9.86 |
| CamTERF15 | Capana04g001184 | CA00g83430 | 0 | Chr04 | 36527349 | 36528682 | 1263 | 420 | 47.68 | 9.46 |
| CamTERF16 | Capana04g001185 | CA00g83420 | 0 | Chr04 | 36553095 | 36554216 | 1122 | 373 | 42.93 | 9.65 |
| CamTERF17 | Capana04g001188 | CA00g83410 | 0 | Chr04 | 36586260 | 36587354 | 1095 | 364 | 41.92 | 9.28 |
| CamTERF18 | Capana04g001189 | CA00g83400 | 6.61 × 10−18 | Chr04 | 36664744 | 36665883 | 1140 | 379 | 43.25 | 9.49 |
| CamTERF19 | Capana04g001190 | CA01g17910 | 0 | Chr04 | 36679829 | 36682422 | 1143 | 380 | 43.5 | 9.47 |
| CamTERF20 | Capana04g001191 | Chr04 | 36685395 | 36685721 | 327 | 108 | 12.4 | 9.52 | ||
| CamTERF21 | Capana04g001192 | CA00g85840 | 0 | Chr04 | 36719507 | 36720607 | 1101 | 366 | 42.01 | 9.51 |
| CamTERF22 | Capana04g001196 | CA04g14910 | 0 | Chr04 | 36830156 | 36837562 | 1185 | 394 | 45.17 | 10.02 |
| CamTERF23 | Capana04g001201 | CA10g08900 | 0 | Chr04 | 36952799 | 36953611 | 813 | 270 | 30.41 | 9.42 |
| CamTERF24 | Capana04g001203 | CA00g85820 | 0 | Chr04 | 37032362 | 37033440 | 981 | 326 | 38 | 9.64 |
| CamTERF25 | Capana04g001204 | CA00g85830 | 1.02 × 10−15 | Chr04 | 37046910 | 37047527 | 618 | 205 | 23.63 | 9.52 |
| CamTERF26 | Capana05g000328 | CA05g03010 | 7.52 × 10−78 | Chr05 | 6905983 | 6907173 | 1191 | 396 | 45.16 | 9.67 |
| CamTERF27 | Capana05g000329 | CA05g03020 | 0 | Chr05 | 6908441 | 6909864 | 1173 | 390 | 44.77 | 10.02 |
| CamTERF28 | Capana05g001366 | CA05g09510 | 0 | Chr05 | 112851518 | 112852951 | 1434 | 477 | 53.95 | 6.1 |
| CamTERF29 | Capana08g002535 | CA00g64290 | 0 | Chr08 | 148385074 | 148388277 | 1971 | 656 | 75.9 | 9.16 |
| CamTERF30 | Capana09g000680 | CA09g07900 | 0 | Chr09 | 32649482 | 32650606 | 1125 | 374 | 43.07 | 9.65 |
| CamTERF31 | Capana10g000180 | CA10g01160 | 0 | Chr10 | 3405765 | 3407096 | 1332 | 443 | 50.49 | 6.12 |
| CamTERF32 | Capana11g000966 | CA11g10880 | 0 | Chr11 | 79029802 | 79030677 | 876 | 291 | 33.23 | 9.57 |
| CamTERF33 | Capana11g001839 | CA00g74110 | 4.08 × 10−15 | Chr11 | 197890867 | 197891505 | 639 | 212 | 24.48 | 8.93 |
| CamTERF34 | Capana12g001016 | CA08g18550 | 0 | Chr12 | 40961437 | 40962249 | 813 | 270 | 31.42 | 9.1 |
| CamTERF35 | Capana12g001548 | CA12g14250 | 6.69 × 10−15 | Chr12 | 119671476 | 119672487 | 930 | 309 | 35.48 | 9.55 |
| Protein ID | Chloroplast Transit Peptide | Mitochondrial Targeting Peptide | Signal Peptide | Other Location | Loc | RC |
|---|---|---|---|---|---|---|
| CamTERF1 | 0.064 | 0.339 | 0.114 | 0.612 | _ | 4 |
| CamTERF2 | 0.888 | 0.047 | 0.016 | 0.07 | C | 1 |
| CamTERF3 | 0.079 | 0.536 | 0.06 | 0.568 | _ | 5 |
| CamTERF4 | 0.062 | 0.883 | 0.008 | 0.21 | M | 2 |
| CamTERF5 | 0.24 | 0.049 | 0.029 | 0.749 | _ | 3 |
| CamTERF6 | 0.098 | 0.127 | 0.165 | 0.532 | _ | 4 |
| CamTERF7 | 0.852 | 0.021 | 0.082 | 0.165 | C | 2 |
| CamTERF8 | 0.049 | 0.681 | 0.007 | 0.028 | M | 2 |
| CamTERF9 | 0.014 | 0.823 | 0.026 | 0.349 | M | 3 |
| CamTERF10 | 0.862 | 0.067 | 0.01 | 0.138 | C | 2 |
| CamTERF11 | 0.733 | 0.043 | 0.021 | 0.102 | C | 2 |
| CamTERF12 | 0.018 | 0.567 | 0.125 | 0.16 | M | 3 |
| CamTERF13 | 0.178 | 0.302 | 0.108 | 0.04 | M | 5 |
| CamTERF14 | 0.106 | 0.27 | 0.019 | 0.732 | _ | 3 |
| CamTERF15 | 0.177 | 0.12 | 0.054 | 0.147 | C | 5 |
| CamTERF16 | 0.082 | 0.857 | 0.006 | 0.031 | M | 2 |
| CamTERF17 | 0.009 | 0.176 | 0.763 | 0.07 | S | 3 |
| CamTERF18 | 0.014 | 0.797 | 0.163 | 0.042 | M | 2 |
| CamTERF19 | 0.261 | 0.635 | 0.021 | 0.098 | M | 4 |
| CamTERF20 | 0.06 | 0.164 | 0.142 | 0.883 | _ | 2 |
| CamTERF21 | 0.007 | 0.194 | 0.85 | 0.058 | S | 2 |
| CamTERF22 | 0.086 | 0.773 | 0.008 | 0.082 | M | 2 |
| CamTERF23 | 0.005 | 0.419 | 0.44 | 0.085 | S | 5 |
| CamTERF24 | 0.274 | 0.078 | 0.024 | 0.39 | _ | 5 |
| CamTERF25 | 0.685 | 0.039 | 0.176 | 0.248 | C | 3 |
| CamTERF26 | 0.069 | 0.284 | 0.085 | 0.06 | M | 5 |
| CamTERF27 | 0.131 | 0.598 | 0.034 | 0.07 | M | 3 |
| CamTERF28 | 0.212 | 0.659 | 0.026 | 0.029 | M | 3 |
| CamTERF29 | 0.665 | 0.268 | 0.01 | 0.103 | C | 4 |
| CamTERF30 | 0.009 | 0.538 | 0.611 | 0.025 | S | 5 |
| CamTERF31 | 0.029 | 0.588 | 0.064 | 0.492 | M | 5 |
| CamTERF32 | 0.071 | 0.339 | 0.012 | 0.707 | _ | 4 |
| CamTERF33 | 0.034 | 0.174 | 0.109 | 0.785 | _ | 2 |
| CamTERF34 | 0.815 | 0.029 | 0.203 | 0.017 | C | 2 |
| CamTERF35 | 0.139 | 0.095 | 0.053 | 0.679 | _ | 3 |
| Motif | Width | Sequence |
|---|---|---|
| 1 | 21 | YLVSHPALLMYSLEKRIKPRY |
| 2 | 29 | PKLLFYDVEKTLKPKLZFLKELGLSGSDL |
| 3 | 50 | VPPDSPMFLHGVQVLSSLKKSKLDRKJGIFKSFGWSDDDILTMFRKLPYC |
| 4 | 31 | YLRSLGGSDEBVVKLJKRCPWLLSYSLEKTL |
| 5 | 21 | YLINSLGFSKQEAJSASAKVT |
| 6 | 22 | SESKFLEKYVLPYKDELPDLYE |
| 7 | 21 | SEARIQTALTFFMKELGYKSA |
| 8 | 28 | LLRNFGFSNDKIRKMVLRCPQLLTQNPE |
| 9 | 21 | LVVNFFKQTGFBBTQIKKLVS |
| 10 | 21 | VKVIARDPKLLTRSLDTHJKP |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, B.; Xie, L.; Yi, T.; Lv, J.; Yang, H.; Cheng, X.; Liu, F.; Zou, X. Genome-Wide Identification and Characterization of the Mitochondrial Transcription Termination Factors (mTERFs) in Capsicum annuum L. Int. J. Mol. Sci. 2020, 21, 269. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010269
Tang B, Xie L, Yi T, Lv J, Yang H, Cheng X, Liu F, Zou X. Genome-Wide Identification and Characterization of the Mitochondrial Transcription Termination Factors (mTERFs) in Capsicum annuum L. International Journal of Molecular Sciences. 2020; 21(1):269. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010269
Chicago/Turabian StyleTang, Bingqian, Lingling Xie, Ting Yi, Junheng Lv, Huiping Yang, Xiang Cheng, Feng Liu, and Xuexiao Zou. 2020. "Genome-Wide Identification and Characterization of the Mitochondrial Transcription Termination Factors (mTERFs) in Capsicum annuum L." International Journal of Molecular Sciences 21, no. 1: 269. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010269