Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes

 ,
, 
Abstract
:1. Introduction
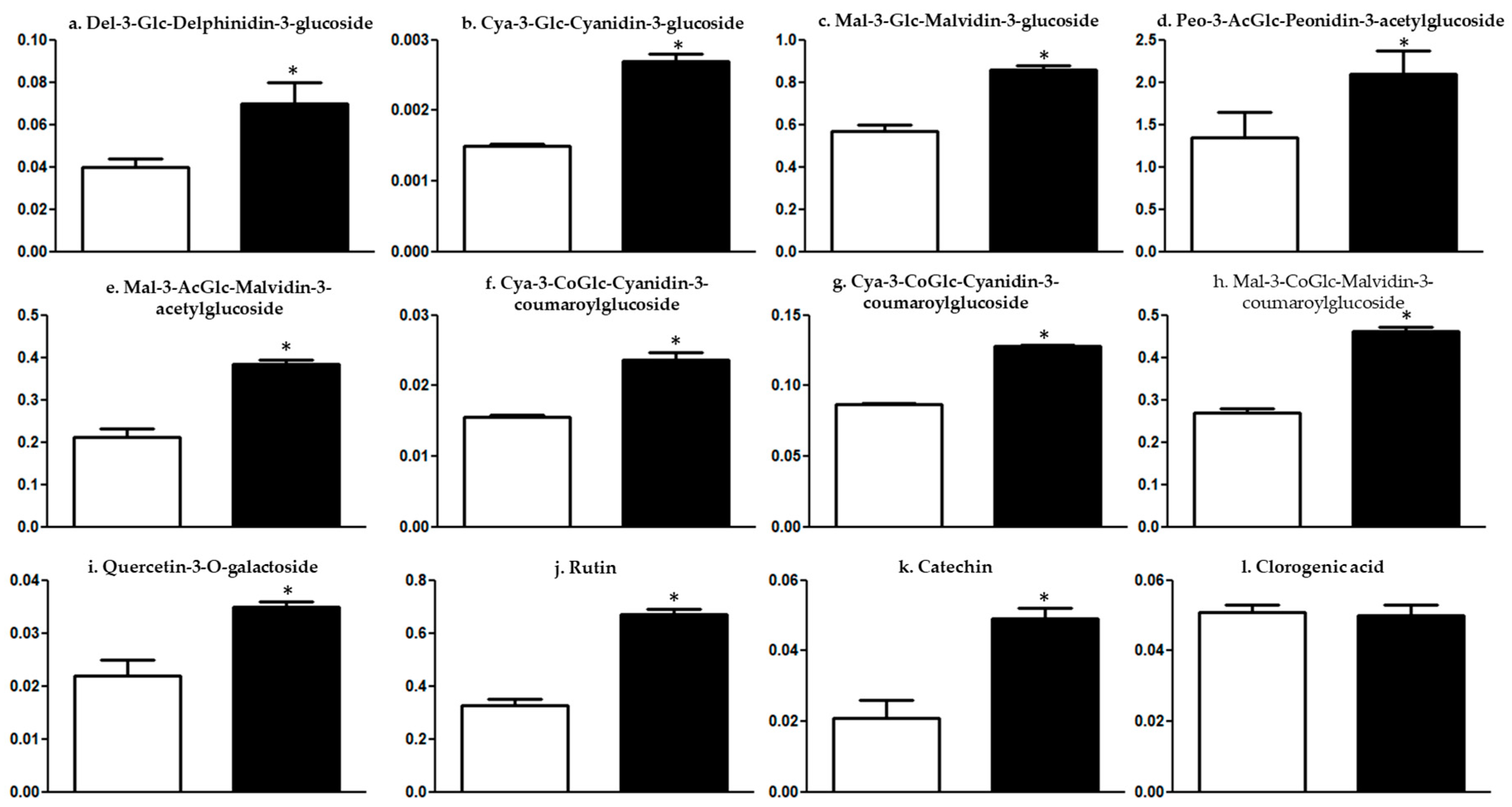
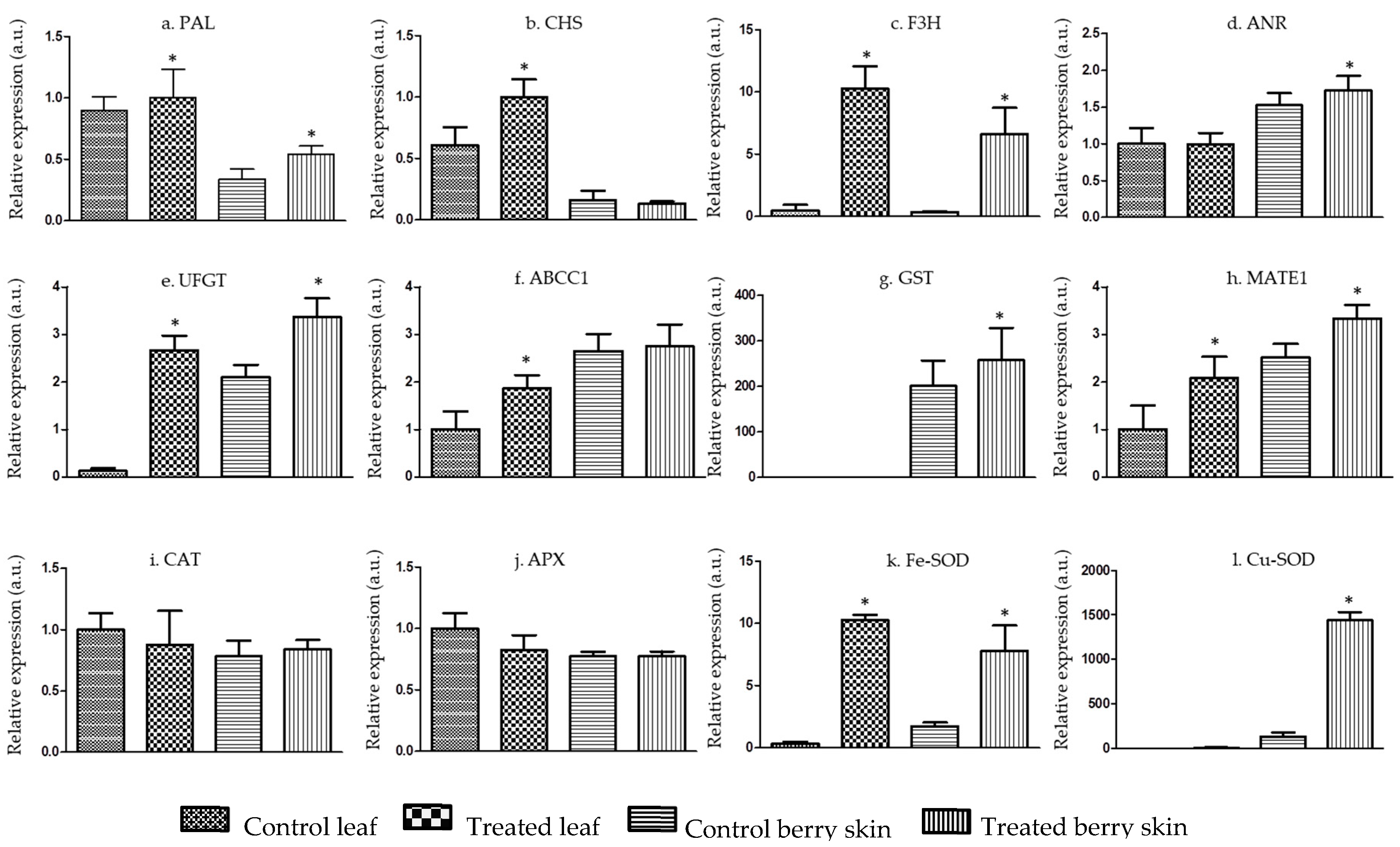
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material, Treatment and Sample Collection
3.2. Extract Preparation
3.3. HPLC-DAD-UV/VIS Analysis
3.4. Total RNA Extraction, cDNA Synthesis and Quantitative Real-Time PCR
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rinaudo, M. Chitin and chitosan: Properties and application. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Reactive oxygen and nitrogen species indefense/stress responses activated by chitosan in sycamore cultured cells. Int. J. Mol. Sci. 2015, 16, 3019–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, M.; Mujtaba, M.; Bulut, E.; Akyuz, B.; Zelencova, L.; Sofi, K. Fluctuation in physicochemical properties of chitins extracted from different body parts of honeybee. Carbohydr. Polym. 2015, 132, 9–16. [Google Scholar] [CrossRef]
- Allan, C.R.; Hadwiger, L.A. The fungicidal effect of chitosan on fungi of varying cell wall composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Sharif, R.; Mujtaba, M.; Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A Review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2018, 25, 313–326. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Tommaso, G.; Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, F.; Ippolito, A.; Venere, D.; Salerno, M. Effects of pre-and postharvest chitosan treatments to control storage grey table grapes. J. Food Sci. 2002, 67, 1862–1867. [Google Scholar] [CrossRef]
- Reglinski, T.; Elmer, P.A.G.; Taylor, J.T.; Wood, P.N.; Hoyte, S.M. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Li, B.; Liu, J.; Tian, S. Physiological responses and quality attributes of table grape fruit to chitosan preharvest spray and postharvest coating during storage. Food Chem. 2008, 106, 501–508. [Google Scholar] [CrossRef]
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Ines, A.; Olivera, A.A.; Falco, V. Chitosan upregulates the genes of the ROS pathway, and enhances the antioxidant potential of grape (Vitis vinifera L. ‘Touriga Francax and ’Tinto Cão’) tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downey, M.O.; Harvey, J.S.; Simon, R. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine Res. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Ollé, D.; Guiraud, J.L.; Souquet, J.M.; Terrier, N.; Ageorges, A.; Cheynier, V.; Verries, C. Effect of pre- and post-veraison water deficit on proanthocyanidin and anthocyanin accumulation during Shiraz berry development. Aust. J. Grape Wine Res. 2011, 17, 90–100. [Google Scholar] [CrossRef]
- Rudolf, J.R.; Resurreccion, A.V.A. Elicitation of Resveratrol in Peanut Kernels by Application of Abiotic Stresses. J. Agric. Food Chem. 2005, 53, 10186–10192. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Tassoni, A.; Franceschetti, M.; Righetti, L.; Naldrett, M.J.; Bagni, N. Chitosan treatment induces changes of protein expression profile and stilbene distribution in Vitis vinifera cell suspensions. Proteomics 2009, 9, 610–624. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Baroja, E.; Santamaría, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 15, 213–221. [Google Scholar] [CrossRef]
- Arakawa, O. Photoregulation of anthocyanin synthesis in apple fruit under UV-B and red light. Plant Cell Physiol. 1988, 29, 1385–1389. [Google Scholar]
- Zhao, Q.; He, F.; Reeves, M.J.; Pan, Q.H.; Duan, C.Q.; Wang, J. Expression of structural genes related to anthocyanin biosynthesis of Vitis amurensis. J. For. Res. 2016, 27, 647–657. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C.; Robinson, S.P. Analysis of the expression of anthocyanin pathway genes in developing Vitis vinifera L. cv Shiraz grape berries and the implications for pathway regulation. Plant Physiol. 1996, 111, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar]
- Chen, J.Y.; Wen, P.F.; Kong, W.F.; Pan, Q.H.; Wan, S.B.; Huang, W.D. Changes and subcellular localizations of the enzymes involved in phenylpropanoid metabolism during grape berry development. Plant Physiol. 2006, 163, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.L.; Holton, T.A.; Ablett, E.M.; Lee, L.S.; Henry, R.J. cDNA microarray analysis of development grape (Vitis vinifera cv Shiraz) berry skin. Funct. Integr. Genom. 2005, 5, 40–58. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, S.D.; Gaspero, G.; Marconi, R.; Nonis, A.; Peterlunger, E.; Paillard, S.; Adam-Blondon, A.F.; Testolin, R. Colour variation in red grapevines (Vitis vinifera L.): Genomic organisation, expression of flavonoid 3′-hydroxylase, flavonoid 3′, 5′-hydroxylase genes and related metabolite profiling of red cyanidin-/blue delphinidin-based anthocyanins in berry skin. BMC Genom. 2006, 7, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Holton, T.A.; Tanaka, Y. Blue roses-a pigment of our imagination? Trends Biotechnol. 1994, 12, 40–42. [Google Scholar] [CrossRef]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M.; et al. ABCC1, an ATP binding cassette protein from 695 grape berry, transports anthocyanidin 3-O-Glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar] [CrossRef] [Green Version]
- Goodman, C.D.; Casati, P.; Walbot, V. A Multidrug Resistance–Associated Protein Involved in 708 Anthocyanin Transport in Zea mays. Plant Cell 2004, 16, 1812–1826. [Google Scholar] [CrossRef] [Green Version]
- Conde, A.; Pimentel, D.; Neves, A.; Dinis, L.T.; Bernardo, S.; Correia, C.M.; Geros, H.; Pereira, J.M. Kaolin Foliar Application Has a Stimulatory Effect on Phenylpropanoid and Flavonoid Pathways in Grape Berries. Front. Plant Sci. 2016, 7, 1150. [Google Scholar] [CrossRef] [Green Version]
- Demurtas, O.C.; Francisco, R.B.; Diretto, G.; Ferrante, P.; Frusciante, S.; Pietrella, M.; Aprea, G.; Borghi, L.; Feeney, M.; Frigerio, L.; et al. ABCC Transporters Mediate the Vacuolar Accumulation of Crocins in Saffron 3 Stigmas. Plant Cell 2019. [Google Scholar] [CrossRef] [Green Version]
- Conn, S.; Curtin, C.; Bezier, A.; Franco, C.; Zhang, W. Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J. Exp. Bot. 2008, 59, 3621–3634. [Google Scholar] [CrossRef] [Green Version]
- Tepperman, J.M.; Dunsmuir, P. Transformed plants with elevated levels of chloroplastic SOD are not more resistant to superoxide toxicity. Plant Mol. Biol. 1990, 14, 501–511. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Lai, Z.; Lin, Y.; Lai, G.; Lian, C. Genome-wide identification and characterization of the superoxide dismutase gene family in Musa acuminata cv. Tianbaojiao (AAA group). BMC Genom. 2015, 16, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Hao, C.; Cheng, Z.M.; Zhong, Y. Genome-Wide Identification, Characterization, and Expression Analysis of the Grapevine Superoxide Dismutase (SOD) Family. Int. J. Genom. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Lai, Z.X. Superoxide dismutase multigene family in longan somatic embryos: A comparison of CuZn-SOD, Fe-SOD, and Mn-SOD gene structure, splicing, phylogeny, and expression. Mol. Breed. 2013, 32, 595–615. [Google Scholar] [CrossRef]
- Baek, K.H.; Skinner, D.Z.; Ling, P.; Chen, X. Molecular structure and organization of the wheat genomic manganese superoxide dismutase gene. Genome 2006, 49, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Kliebenstein, D.J.; Monde, R.A.; Last, R.L. Superoxide dismutase in Arabidopsis: An eclectic enzyme family with disparate regulation and protein localization. Plant Physiol. 1998, 2, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Filiz, E.; Tombuloğlu, H. Genome-wide distribution of superoxide dismutase (SOD) gene families in Sorghum bicolor. Turk. J. Biol. 2015, 39, 49–59. [Google Scholar] [CrossRef]
- Molina-Rueda, J.J.; Tsai, C.J.; Kirby, E.G. The Populus superoxide dismutase gene family and its responses to drought stress in transgenic poplar overexpressing a pine cytosolic glutamine synthetase (GS1a). PLoS ONE 2013, 8, e56421. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Xia, M.; Chen, J.; Deng, F.; Yuan, R.; Zhang, X.; Shen, F. Genome-wide analysis of superoxide dismutase gene family in Gossypium raimondii and G. arboretum. Plant Gene 2016, 6, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Valletta, A.; De Angelis, G.; Badiali, C.; Brasili, E.; Miccheli, A.; Di Cocco, M.E.; Pasqua, G. Acetic acid acts as an elicitor exerting a chitosan-like effect on xanthone biosynthesis in Hypericum perforatum L. root cultures. Plant Cell Rep. 2016, 35, 1009–1020. [Google Scholar] [CrossRef]
- Aires, A.; Carvalho, R.; Matos, M.; Carnide, V.; Silva, A.P.; Gonçalves, B. Variation of chemical constituents, antioxidant activity, and endogenous plant hormones throughout different ripening stages of highbush blueberry (Vaccinium corymbosum L.) cultivars produced in centre of Portugal. J. Food Biochem. 2017, 41, e12414. [Google Scholar] [CrossRef]
- Gasic, K.; Hernandez, A.; Korban, S.S. RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction. Plant Mol. Biol. Rep. 2004, 22, 437–438. [Google Scholar] [CrossRef]
- Arvidsson, S.; Kwasniewski, M.; Riaño-Pachón, D.M.; Mueller-Roeber, B. QuantPrime—A flexible tool for reliable high-throughput primer design for quantitative PCR. BMC Bioinform. 2008, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Gene | Primer Sequence | Accession | Annealing Temperature | |
|---|---|---|---|---|
| Fe-SOD | Superoxide dismutase | F 5′ CCTTTGTGAACCTAGGCGAACC 3′ R 5′ TGGCCGGGTTAGCTTGAACTC 3′ | XM_019222444 | 55 °C |
| Cu/Zn-SOD | Superoxide dismutase | F 5′ AGATTGGCATGTGGTGTTGTTG 3′ R 5′ ACTCCCACATTACCCAACAACA 3′ | NM_001281138.1 | 55 °C |
| CAT | Catalase | F 5′ GGTGTTCACACCTTCACTCT 3′ R 5′ GAGATCCTGAGTAGCATGACTG 3′ | KP271927 | 55 °C |
| APX | Ascorbate peroxidase | F 5′ ATCTGGTGGTCATACTCTGG 3′ R 5′ TCTAGGAGAGCCTTGTCTGA 3′ | XM_010655137 | 55 °C |
| PAL | Phenylalanine ammonia-lyase | F 5′ CCTACTGTTCAGAGCTCCAG 3′ R 5′ GCCACTAGGTATGTGGTAGACA 3′ | XM_003633937 | 55 °C |
| CHS | Chalcone synthase | F 5′ CACTCTTCGAACTCGTCTCT 3′ R 5′ CCACCAAGCTCTTCTCTATG 3′ | KT589834 | 55 °C |
| F3H | Flavanone3-hydroxylase | F5′CAGTGCAAGACTGGCGCGAGATCGTA3′ R 5′ TAGCCTCAGACAACACCTCCAGCAACT 3′ | KY006128 | 52 °C |
| ANR | Anthocyanidin reductase | F 5′ CTGTCAGGTTCAGTCTCCAT 3′ R 5′ GTTGGGACTTTGTACTGAGG 3′ | NM_001280956 | 55 °C |
| UFGT | UDP glucose: flavonoid 3-o-glucosyltransferase | F 5′ TGCAGGGCCTAACTCACTCT 3′ R 5′ GCAGTCGCCTTAGGTAGCAC 3′ | GSVIVT0102441901 | 55 °C |
| ABCC1 | Anthocyanin transporter | F 5′ CTCCACTGGTCCTCTGCTTC 3′ R 5′ AGCCTGCTTCGAAAGTACCA 3′ | GSVIVT01028722001 | 55 °C |
| MATE1 | Tonoplast transporter | F 5′ TGCTTTTGTGATTTTGTTAGAGG 3′ R 5′ CCCTTCCCCGATTGAGAGTA 3′ | GSVIVT01028885001 | 55 °C |
| GST | glutathioneS-transferase | F 5′ AAGGATCCATGGTGATGAAGGTGTATGGC 3′ R 5′ AACTGCAGAAGCCAACCAACCAACAAAC 3′ | GSVIVG01035256001 | 55 °C |
| ACT | Actin | F 5′ GTGCCTGCCATGTATGTTGCC 3′ R 5′ GCAAGGTCAAGACGAAGGATA 3′ | GSVIVT010265800 01 | 55 °C |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010306
Singh RK, Martins V, Soares B, Castro I, Falco V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. International Journal of Molecular Sciences. 2020; 21(1):306. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010306
Chicago/Turabian StyleSingh, Rupesh Kumar, Viviana Martins, Bruno Soares, Isaura Castro, and Virgílio Falco. 2020. "Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes" International Journal of Molecular Sciences 21, no. 1: 306. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010306