Metabolic Adaptation to Sulfur of Hyperthermophilic Palaeococcus pacificus DY20341T from Deep-Sea Hydrothermal Sediments

Abstract
:1. Introduction
2. Results and Discussions
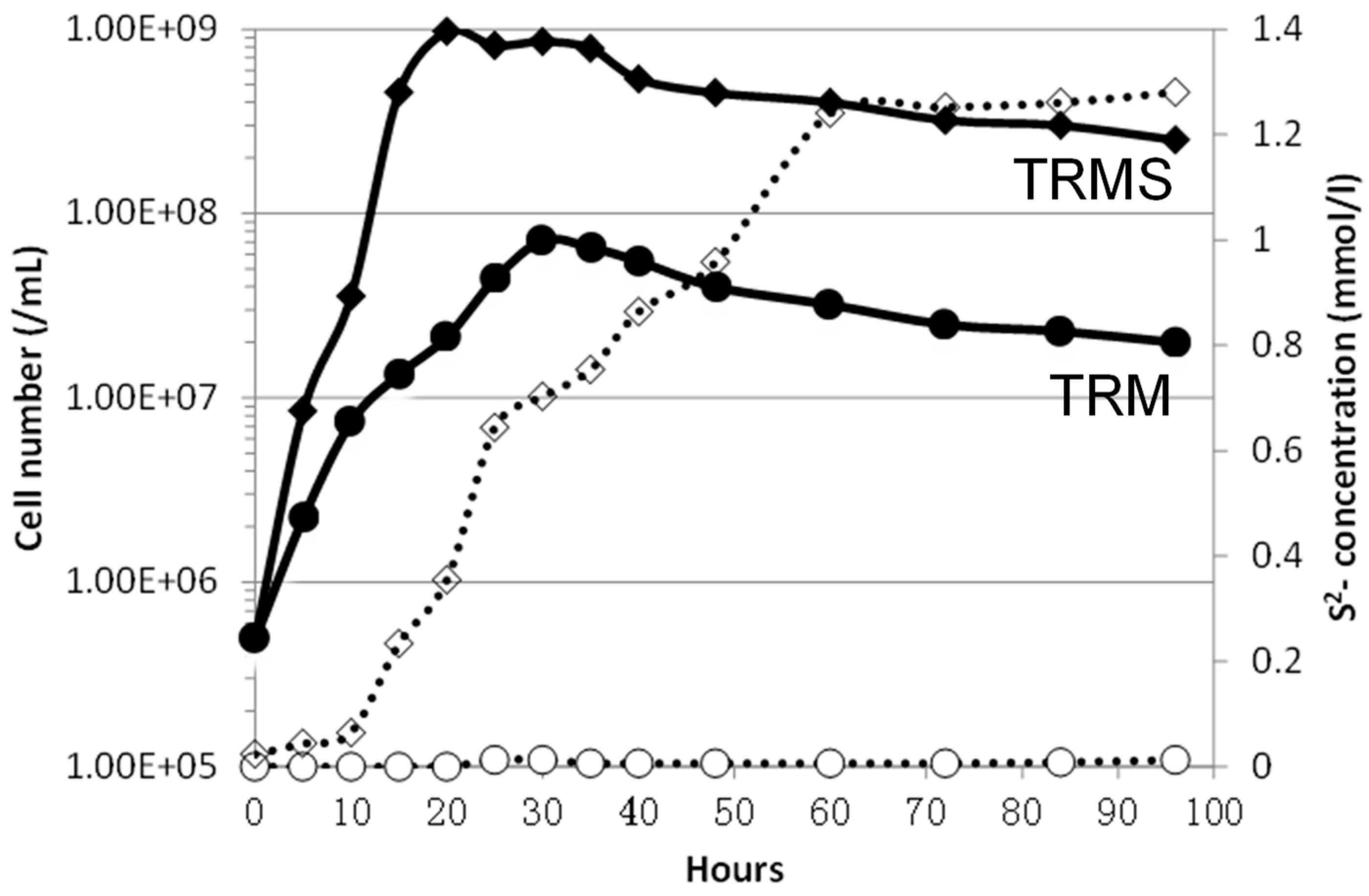
2.1. The Growth Curve of Pa. pacificus with or without Elemental Sulfur
2.2. Genomic and Transcriptomic Features of Pa. pacificus DY20341T
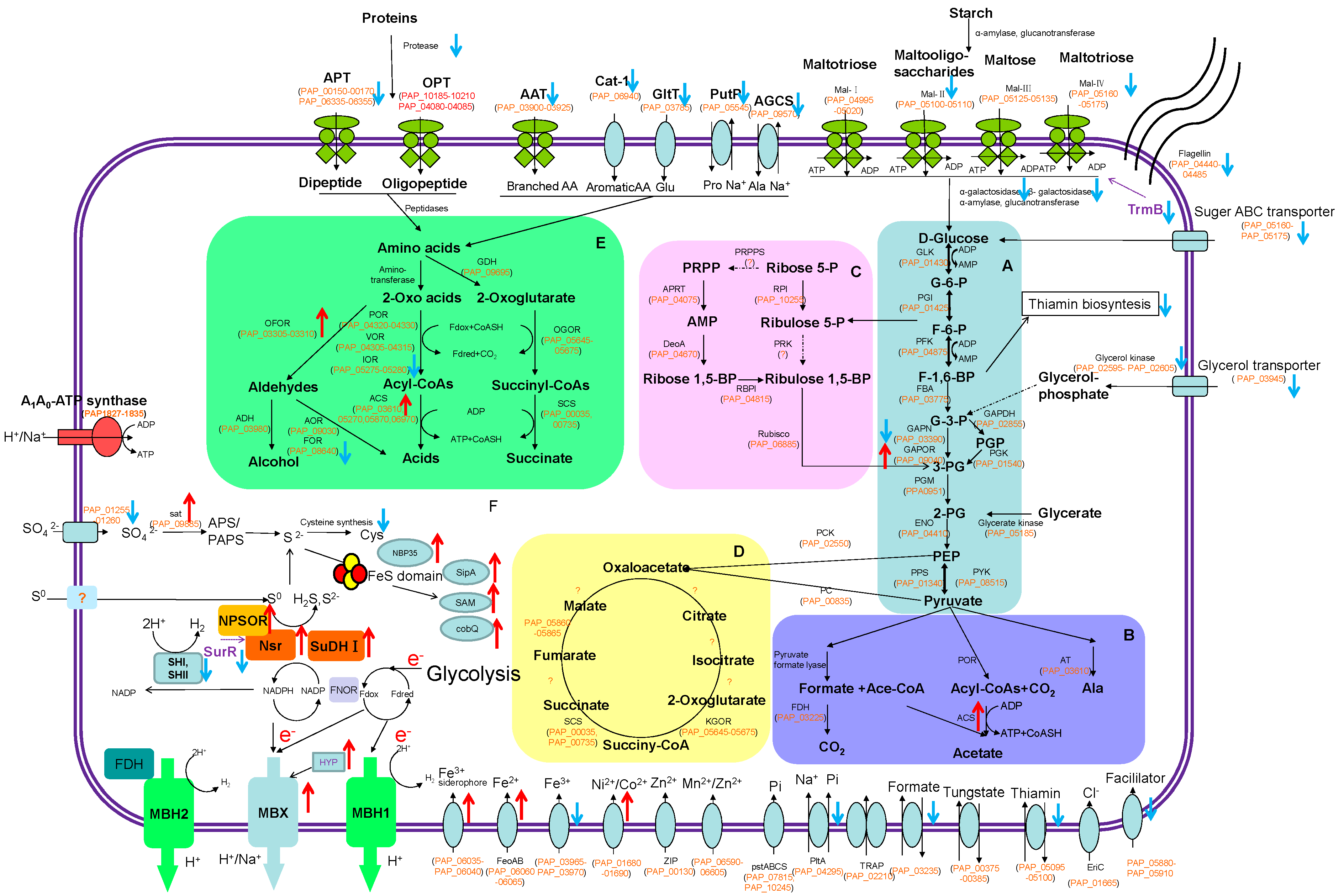
2.3. Metabolic Response to Growth on Sulfur
2.3.1. Central Metabolism
2.3.2. Utilization of Proteins
2.3.3. Utilization of Carbohydrates
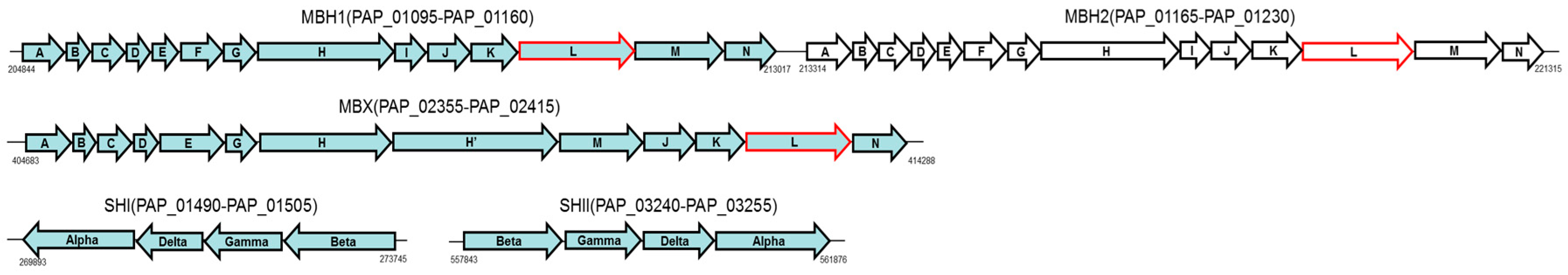
2.3.4. Energy Metabolism
2.3.5. Sulfur Assimilation
2.3.6. Transcription Regulation under Sulfur as an Electron Acceptor
2.3.7. Cell Mobility
2.3.8. Mobile Elements (Transporters, Viruses, and CRISPR Elements)
2.3.9. Adaptation against High Temperature, High Pressure, and Oxygen
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
3.2. RNA Purification and Transcriptome Sequence
3.3. Data Processing and Analysis
3.4. Determination of Protease Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gartman, A.; Yucel, M.; Madison, A.S.; Chu, D.W.; Ma, S.; Janzen, C.P.; Becker, E.L.; Beinart, R.A.; Girguis, P.R.; Luther, G.W. Sulfide oxidation across diffuse flow zones of hydrothermal vents. Aquat. Geochem. 2011, 17, 583–601. [Google Scholar] [CrossRef]
- Bonchosmolovskaya, E.A. Bacterial sulfur reduction in hot vents. FEMS Microbiol. Rev. 1994, 15, 65–77. [Google Scholar] [CrossRef]
- Offre, P.; Spang, A.; Schleper, C. Archaea in biogeochemical cycles. Annu. Rev. Microbiol. 2013, 67, 437–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, S.D.; Baross, J.A.; Delaney, J.R. Volcanoes, fluids, and life at mid-ocean ridge spreading centers. Annu. Rev. Earth Planet. Sci. 2002, 30, 385–491. [Google Scholar] [CrossRef] [Green Version]
- Roussel, E.; Bonavita, M.C.; Querellou, J.; Cragg, B.A.; Webster, G.; Prieur, D.; Parkes, R.J. Extending the sub-sea-floor biosphere. Science 2008, 320, 1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teske, A.; Edgcomb, V.P.; Rivers, A.R.; Thompson, J.R.; Gomez, A.D.; Molyneaux, S.J.; Wirsen, C.O.A. A molecular and physiological survey of a diverse collection of hydrothermal vent Thermococcus and Pyrococcus isolates. Extremophiles 2009, 13, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Robb, F.T.; Place, A.R.; Sowers, K.R.; Schreier, H.J.; DasSarma, S.; Fleischmann, E.M. Archaea: A Laboratory Manual: Thermophiles; Cold Spring Harbor Laboratory Press: Suffolk County, NY, USA, 1995. [Google Scholar]
- Schut, G.J.; Bridger, S.L.; Adams, M.W.W. Insights into the metabolism of elemental sulfur by the hyperthermophilic archaeon Pyrococcus furiosus: Characterization of a coenzyme A- dependent NAD(P)H sulfur oxidoreductase. J. Bacteriol. 2007, 189, 4431–4441. [Google Scholar] [CrossRef] [Green Version]
- Bridger, S.L.; Clarkson, S.M.; Stirrett, K.; Debarry, M.; Lipscomb, G.L.; Schut, G.J.; Westpheling, J.; Scott, R.A.; Adams, M.W.W. Deletion strains reveal metabolic roles for key elemental sulfur-rResponsive proteins in Pyrococcus furiosus. J. Bacteriol. 2011, 193, 6498–6504. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.; Kwon, J.S.; Yun, S.H.; Lim, H.L.; Kim, J.; Kim, S.J.; Kang, S.G.; Lee, J.; Kim, S.; Chung, Y.H. Proteomic insights into sulfur metabolism in the hydrogen-producing hyperthermophilic archaeon Thermococcus onnurineus NA1. Int. J. Mol. Sci. 2015, 16, 9167–9195. [Google Scholar] [CrossRef]
- Ladenstein, R.; Ren, B. Protein disulfides and protein disulfide oxidoreductases in hyperthermophiles. FEBS J. 2006, 273, 4170–4185. [Google Scholar] [CrossRef]
- Takai, K.; Sugai, A.; Itoh, T.; Horikoshi, K. Palaeococcus ferrophilus gen. nov., s nov., a barophilic, hyperthermophilic arcaeon from a deep sea hydrothermal vent chimney. Int. J. Syst. Evol. Microbiol. 2000, 50, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Amend, J.P.; Meyer-Dombard, D.R.; Sheth, S.N.; Zolotova, N.; Amend, A.C. Palaeococcus helgesonii sp. nov., a facultatively anaerobic, hyperthermophilic archaeon from a geothermal well on Vulcano Island, Italy. Arch. Microbiol. 2003, 179, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, X.; Jiang, L.; Alain, K.; Jebbar, M.; Shao, Z. Palaeococcus pacificus sp. nov., an archaeon from deep-sea hydrothermal sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 2155–2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Jebbar, M.; Shao, Z. Complete genome sequence of hyperthermophilic piezophilic archaeon Palaeococcus pacificus DY20341T, isolated from deep-sea hydrothermal sediments. Genome Announc. 2015, 3, e01080-15. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Atomi, H.; Imanaka, T. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Sato, T.; Atomi, H.; Imanaka, T. Archaeal type III RuBisCOs function in a pathway for AMP metabolism. Science 2007, 315, 1003–1006. [Google Scholar] [CrossRef]
- Bowden, S.D.; Bauer, M.W.; Shockley, K.R.; Pysz, M.A.; Kelly, R.M. An incomplete TCA cycle increases survival of Salmonella Typhimurium during infection of resting and activated murine macrophages. PLoS ONE 2010, 5, e13871. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.E.; Fukui, T.; Fujiwara, S. Proteolysis in hyperthermophilic microorganisms. Archaea 2002, 1, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Mardanov, A.V.; Bagramyan, K.; Adams, M.W.W. Metabolic versatility and indigenous origin of the archaeon Thermococcus sibiricus, isolated from a Siberian oil reservoir, as revealed by genome analysis. Appl. Environ. Microbiol. 2009, 75, 4580–4588. [Google Scholar] [CrossRef] [Green Version]
- Shikata, K.; Atomi, H.; Kanai, T.; Matsumi, R.; Fujiwara, S.; Imanaka, T. A novel ADP-forming succinyl-CoA synthetase in Thermococcus kodakaraensis structurally related to the archaeal nucleoside diphosphate-forming acetyl-CoA synthetases. J. Biol. Chem. 2007, 282, 26963–26970. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Brehm, S.D.; Datta, S.; Adams, M.W. Growth of hyperthermophilic archaeon Pyrococcus furiosus on chitin involves two family 18 chitinases. Appl. Environ. Microbiol. 2003, 69, 3119–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadimitriou, K.; Matsuoka, R.; Beppu, H.; Nakajima, A.; Okada, Y.; Atomi, H.; Imanaka, T. Analysis of the complete genome sequence of the archaeon Pyrococcus chitonophagus DSM 10152 (formerly Thermococcus chitonophagus). Extremophiles 2016, 20, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Imanaka, T.; Fukui, T.; Fujiwara, S. Chitinase from Thermococcus kodakaraensis KOD1. Methods Enzymol. 2001, 330, 319–329. [Google Scholar] [PubMed]
- Sapra, R.; Bagramyan, K.; Adams, M.W.W. A simple energy-conserving system: Proton reduction coupled to proton translocation. Proc. Natl. Acad. Sci. USA 2003, 100, 7545–7550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, T.; Cooper, A.J.L. Complete genome sequence of the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1 and comparison with Pyrococcus genomes. Genome Res. 2005, 15, 352–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhu, W.; Major, T.A.; Roy, H.; Palioura, S.; Jahn, D.; Whitman, W.B.; Yates, J.R.; Ibba, M.; Soll, D. Complete genome sequence of Thermococcus sp. strain 4557, a hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent area. J. Bacteriol. 2011, 193, 5544–5545. [Google Scholar] [CrossRef] [Green Version]
- Vannier, P.; Sieprawskalupa, M.; Whitman, W.B.; White, R.H. Complete genome sequence of the hyperthermophilic, piezophilic, heterotrophic, and carboxydotrophic archaeon Thermococcus barophilus MP. J. Bacteriol. 2011, 193, 1481. [Google Scholar] [CrossRef] [Green Version]
- Schut, G.J.; Newcomer, E.C.; Young, E.G.; Adams, M.W. Whole-genome dna microarray analysis of a hyperthermophile and an archaeon: Pyrococcus furiosus grown on carbohydrates or peptides. J. Bacteriol. 2003, 185, 3935–3947. [Google Scholar] [CrossRef] [Green Version]
- Kanai, T.; Drevland, R.M.; Downs, D.M.; Graham, D.E. Distinct physiological roles of the three [NiFe]-hydrogenase orthologs in the hyperthermophilic archaeon Thermococcus kodakarensis. J. Bacteriol. 2011, 193, 3109–3116. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.J.; Kwon, J.; Yun, S.H.; Lim, H.L.; Kim, M.S.; Kang, S.G.; Lee, J.H.; Choi, J.S.; Kim, S.I.; Chung, Y.H. Proteome analyses of hydrogen-producing hyperthermophilic archaeon Thermococcus onnurineus NA1 in different one-carbon substrate culture conditions. Mol. Cell. Proteom. 2012, 11, M111.015420. [Google Scholar] [CrossRef] [Green Version]
- Henstra, A.M.; Dijkema, C.; Stams, A.J.M. Archaeoglobus fulgidus couples CO oxidation to sulfate reduction and acetogenesis with transient formate accumulation. Environ. Microbiol. 2007, 9, 1836–1841. [Google Scholar] [CrossRef] [PubMed]
- Toohey, J.I.; Cooper, A.J.L. Thiosulfoxide (sulfane) sulfur: New chemistry and new regulatory roles in biology. Molecules 2014, 19, 12789–12813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauerwald, A.; Ghosh, S.; Diruggiero, J. RNA-dependent cysteine biosynthesis in archaea. Science 2005, 307, 1969–1972. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Escobarparamo, P.; Mongodin, E.F.; Nelson, K.E.; Diruggiero, J. Cysteine is not the sulfur source for iron-sulfur cluster and methionine biosynthesis in the methanogenic archaeon Methanococcus maripaludis. J. Biol. Chem. 2010, 285, 31923–31929. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, S.M.; Dunn, D.M.; Maeder, D.L.; Holleyshanks, R.; Chatard, J.; Horlacher, R.; Robb, F.T.; Boos, W.; Weiss, R.B. The elemental sulfur-responsive protein (sipa) from the hyperthermophilic archaeon Pyrococcus furiosus is regulated by sulfide in an iron-dependent manner. J. Bacteriol. 2010, 192, 5841–5843. [Google Scholar] [CrossRef] [Green Version]
- Boyd, J.M.; Clark, D.S. Archaeal ApbC/Nbp35 homologs function as iron-sulfur cluster carrier proteins. J. Bacteriol. 2009, 191, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Lipscomb, G.L.; Singer, G.A.C. SurR: A transcriptional activator and repressor controlling hydrogen and elemental sulphur metabolism in Pyrococcus furiosus. Mol. Microbiol. 2009, 71, 332–349. [Google Scholar] [CrossRef] [Green Version]
- Bairoch, A.M. A possible mechanism for metal-ion induced DNA-protein dissociation in a family of prokaryotic transcriptional regulators. Nucleic Acids Res. 1993, 21, 2515. [Google Scholar] [CrossRef]
- Lee, S.; Engelmann, A.; Horlacher, R.; Qu, Q.; Vierke, G.; Hebbeln, C.; Thomm, M.; Boos, W. TrmB, a Sugar-specific Transcriptional Regulator of the Trehalose/Maltose ABC Transporter from the Hyperthermophilic Archaeon Thermococcus litoralis. J. Biol. Chem. 2003, 278, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Surma, M.; Hausner, W.; Thomm, M.; Boos, W. The role of TrmB and TrmB-like transcriptional regulators for sugar transport and metabolism in the hyperthermophilic archaeon Pyrococcus furiosus. Arch. Microbiol. 2008, 190, 247–256. [Google Scholar] [CrossRef]
- Escobarparamo, P.; Ghosh, S.; Diruggiero, J. Evidence for Genetic Drift in the diversification of a geographically isolated population of the hyperthermophilic archaeon Pyrococcus. Mol. Biol. Evol. 2005, 22, 2297–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.; Escobarparamo, P.; Mongodin, E.F.; Nelson, K.E.; Diruggiero, J. Extensive genome rearrangements and multiple horizontal gene transfers in a population of Pyrococcus isolates from Vulcano Island, Italy. Appl. Environ. Microbiol. 2008, 74, 6447–6451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diruggiero, J.; Dunn, D.M.; Maeder, D.L.; Holleyshanks, R.; Chatard, J.; Horlacher, R.; Robb, F.T.; Boos, W.; Weiss, R.B. Evidence of recent lateral gene transfer among hyperthermophilic Archaea. Mol. Microbiol. 2000, 38, 684–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, F.T.; Clark, D.S. Adaptation of proteins from hyperthermophiles to high pressure and high temperature. J. Mol. Microbiol. Biotechnol. 1999, 1, 101. [Google Scholar]
- Hickey, D.A.; Singer, G.A.C. Genomic and proteomic adaptations to growth at high temperature. Genome Biol. 2004, 5, 117. [Google Scholar] [CrossRef] [Green Version]
- Giulio, M.D. A comparison of proteins from Pyrococcus furiosus and Pyrococcus abyssi: Barophily in the physicochemical properties of amino acids and in the genetic code. Gene 2005, 346, 1–6. [Google Scholar] [CrossRef]
- Jun, X.; Liu, L.; Xu, M.; Oger, P.; Wang, F.; Jebbar, M.; Xiao, X. Complete genome sequence of the obligate piezophilic hyperthermophilic archaeon Pyrococcus yayanosii CH1. J. Bacteriol. 2011, 193, 4297–4298. [Google Scholar] [CrossRef] [Green Version]
- Nath, A.; Subbiah, K. Insights into the molecular basis of piezophilic adaptation: Extraction of piezophilic signatures. J. Theor. Biol. 2016, 390, 117–126. [Google Scholar] [CrossRef]
- Jenney, F.E.; Verhagen, M.F.; Cui, X.; Adams, M.W.W. Anaerobic microbes: Oxygen detoxification without superoxide dismutase. Science 1999, 286, 306–309. [Google Scholar] [CrossRef]
- Cline, J.D. Spectrophotometric determination of hydrogen sulfide in natural waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Fang, X.; Yang, H.; Wang, J.; Kristiansen, K.; Wang, J. SNP detection for massively parallel whole-genome resequencing. Genome Res. 2009, 19, 1124–1132. [Google Scholar] [CrossRef] [Green Version]
- Audic, S.; Claverie, J. The Significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]
- Zdobnov, E.M.; Tegenfeldt, F.; Kuznetsov, D.; Waterhouse, R.M.; Simao, F.A.; Ioannidis, P.; Seppey, M.; Loetscher, A.; Kriventseva, E.V. OrthoDB v9.1: Cataloging evolutionary and functional annotations for animal, fungal, plant, archaeal, bacterial and viral orthologs. Nucleic Acids Res. 2017, 45, D744–D749. [Google Scholar] [CrossRef]
- Sondergaard, D.; Pedersen, C.N.S.; Greening, C. HydDB: A web tool for hydrogenase classification and analysis. Sci. Rep. 2016, 6, 34212. [Google Scholar] [CrossRef]
- Cuppenyard, C. Sigma’s non-specific protease activity assay—Casein as a substrate. J. Vis. Exp. 2008, 19, 899. [Google Scholar]
- Felbeck, H.; Somero, G.N. Primary production in deep-sea hydrothermal vent organisms: Roles of sulfide-oxidizing bacteria. Trends Biochem. Sci. 1982, 7, 201–204. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Gene ID | TRMS- RPKM | TRM- RPKM | log2 (TRMS/TRM) | p-Value | Predicted Protein (Top Hit Species) |
|---|---|---|---|---|---|---|
| Up | PAP_06520 | 15,920 | 212 | 6.23 | 0 |
Fe-Mo cluster-binding protein, SipA (Thermococcus gammatolerans EJ3) |
| PAP_06515 | 1980 | 101 | 4.29 | 0 |
Fe–Mo cofactor-binding protein (Thermococcus barophilus MP) | |
| PAP_06425 | 3373 | 253 | 3.74 | 0 |
Cobalamin biosynthesis protein, CobQ: iron–sulfur cluster binding
domain (Thermococcus barophilus MP) | |
| PAP_01595 | 32,502 | 3010 | 3.43 | 0 |
Ni–Fe hydrogenase metallocenter assembly protein, HypF (Thermococcus barophilus MP) | |
| PAP_06430 | 2931 | 276 | 3.41 | 1.27 × 10−244 |
Cobalamin biosynthesis protein, CobQ: iron–sulfur cluster binding
domain (Thermococcus barophilus MP) | |
| PAP_02400 | 1490 | 170 | 3.13 | 6.34 × 10−191 |
NADH dehydrogenase subunit B (Thermococcus onnurineus NA1) | |
| PAP_04045 | 1170 | 136 | 3.10 | 0 |
Hypothetical protein (Thermococcus litoralis DSM 5473) | |
| PAP_03165 | 8830 | 1083 | 3.03 | 1.83 × 10−200 |
Hypothetical protein (Thermococcus kodakarensis KOD) | |
| PAP_08665 | 1288 | 165 | 2.97 | 0 |
Hypothetical protein (Pyrococcus yayanosii) | |
| PAP_06035 | 3912 | 515 | 2.93 | 0 | ABC-type iron (III)-siderophore transporter permease, ABC.FEV.P (Thermococcus barophilus MP) | |
| Down | PAP_03140 | 7 | 97 | −3.79 | 2.26 × 10−22 |
50S ribosomal protein L35Ae (Thermococcus barophilus MP) |
| PAP_04135 | 21 | 223 | −3.41 | 7.98 × 10−46 |
Hypothetical protein (Thermococcus barophilus MP) | |
| PAP_06725 | 12 | 103 | −3.10 | 1.51 × 10−22 |
Cobalamin adenosyltransferase (Thermococcus litoralis DSM 5473) | |
| PAP_04130 | 36 | 293 | −3.02 | 3.40 × 10−54 |
Diacetylchitobiose deacetylase (Thermococcus barophilus MP) | |
| PAP_05255 | 14 | 106 | −2.92 | 4.46 × 10−20 |
Nucleotide-binding protein (Thermococcus litoralis DSM 5473) | |
| PAP_07560 | 30 | 222 | −2.89 | 7.70 × 10−40 |
Hypothetical protein (Ktedonobacter racemifer DSM 44963) | |
| PAP_08745 | 87 | 631 | −2.86 | 7.29 × 10−9 |
Cystathionine beta-synthase (Pyrococcus sp. NA2) | |
| PAP_04125 | 32 | 225 | −2.81 | 2.03 × 10−39 |
Hydrolase (Thermococcus barophilus MP) | |
| PAP_04120 | 52 | 364 | −2.80 | 1.53 × 10−62 |
Hypothetical protein (Thermococcus barophilus MP) | |
| PAP_06600 | 10 | 70 | −2.80 | 2.49 × 10−13 |
Cro regulatory protein (Thermococcus litoralis DSM 5473) |
| Protein Function | Protein Name (Abbreviation) | Pa. pacificus | P. furious | T. onnurineus |
|---|---|---|---|---|
| Hydrogenase | MBH1 | ↑ | ↓ | ↓ |
| MBH2 | - | ↓ | Does not exist | |
| MBX | ↑ | ↑ | ↑ | |
| SHI | ↓ | ↓ | ↓ | |
| SHII | - | ↓ | ↓ | |
| Sulfur-reducing metabolism | SuDHI | ↑ | ↑ | ↑ |
| SuDHII | Does not exist | Does not exist | ↑ | |
| NPSOR | ↑ | Does not exist | ↑ | |
| Nsr | ↑ | ↑ | ↑ | |
| Iron-sulfur metabolism | SipA | ↑ | ↑ | ↑ |
| SipB | Does not exist | Does not exist | ↑ | |
| FeoB | ↑ | ↑ | ↑ | |
| SufB | ↓ | ↑ | ↑ | |
| Mrp/NBP35 | ↑ | N.D. | ↑ | |
| Regulator | SurR | ↓ | ↓ | ↓ |
| Superoxide-reducing system | NROR | ↑ | N.D. | ↓ |
| SOR | - | N.D. | ↓ | |
| Rr | - | N.D. | ↓ | |
| ATP synthesis | ATP synthase | - | ↑ | ↑ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Zhang, X.; Shao, Z. Metabolic Adaptation to Sulfur of Hyperthermophilic Palaeococcus pacificus DY20341T from Deep-Sea Hydrothermal Sediments. Int. J. Mol. Sci. 2020, 21, 368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010368
Zeng X, Zhang X, Shao Z. Metabolic Adaptation to Sulfur of Hyperthermophilic Palaeococcus pacificus DY20341T from Deep-Sea Hydrothermal Sediments. International Journal of Molecular Sciences. 2020; 21(1):368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010368
Chicago/Turabian StyleZeng, Xiang, Xiaobo Zhang, and Zongze Shao. 2020. "Metabolic Adaptation to Sulfur of Hyperthermophilic Palaeococcus pacificus DY20341T from Deep-Sea Hydrothermal Sediments" International Journal of Molecular Sciences 21, no. 1: 368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010368