Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway

 , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
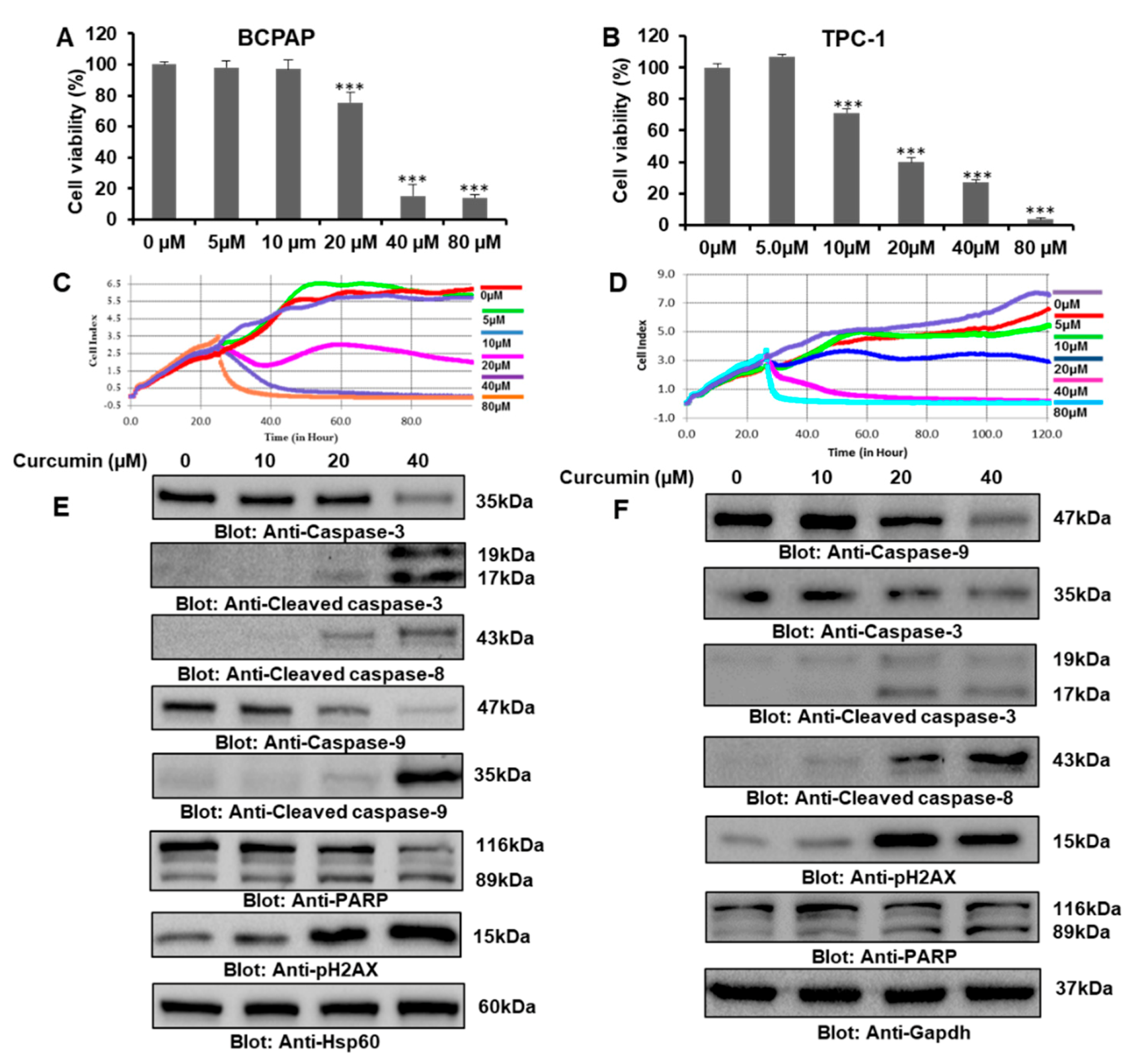
2.1. Curcumin-Mediated Inhibition of Cell Proliferation and Apoptosis in PTC Cells
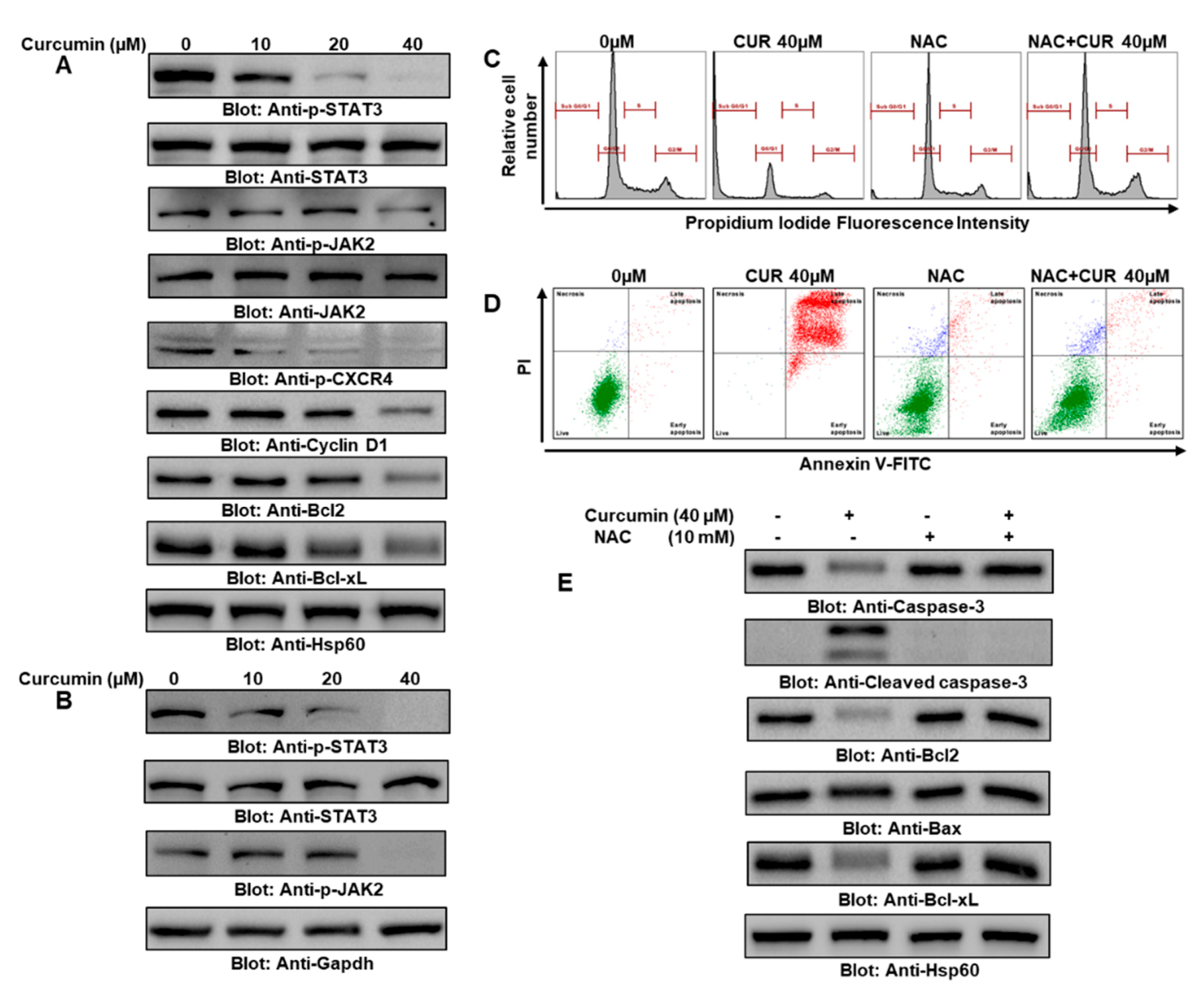
2.2. Curcumin Suppressed the Constitutive JAK/STAT3 Signaling Pathway and Upregulated the ROS Level to Induce Apoptosis
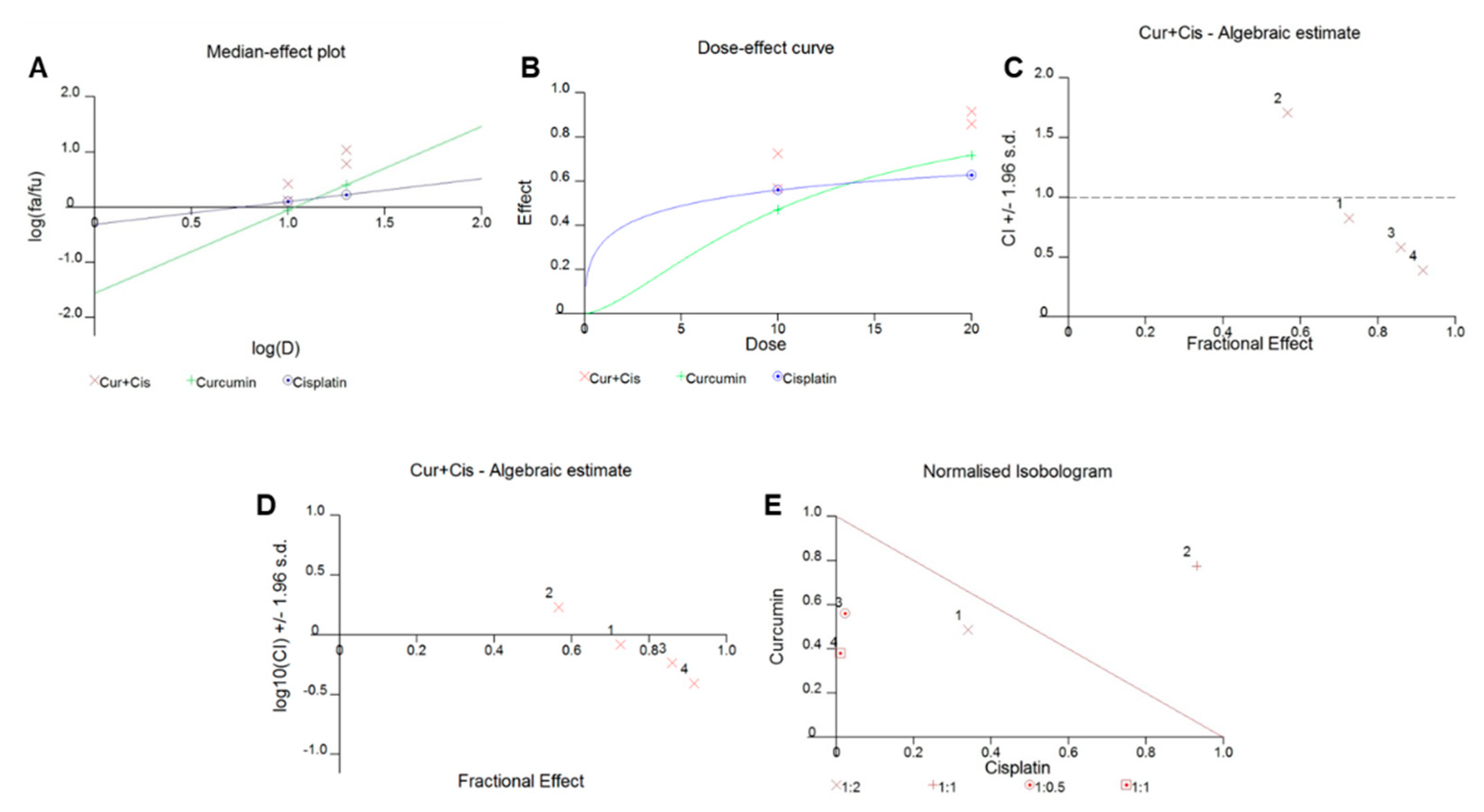
2.3. Synergistic Activity of Curcumin and Cisplatin in PTC Cell Lines
2.4. Cotreatment with Curcumin and Cisplatin Augmented the Inhibition of Cell Viability
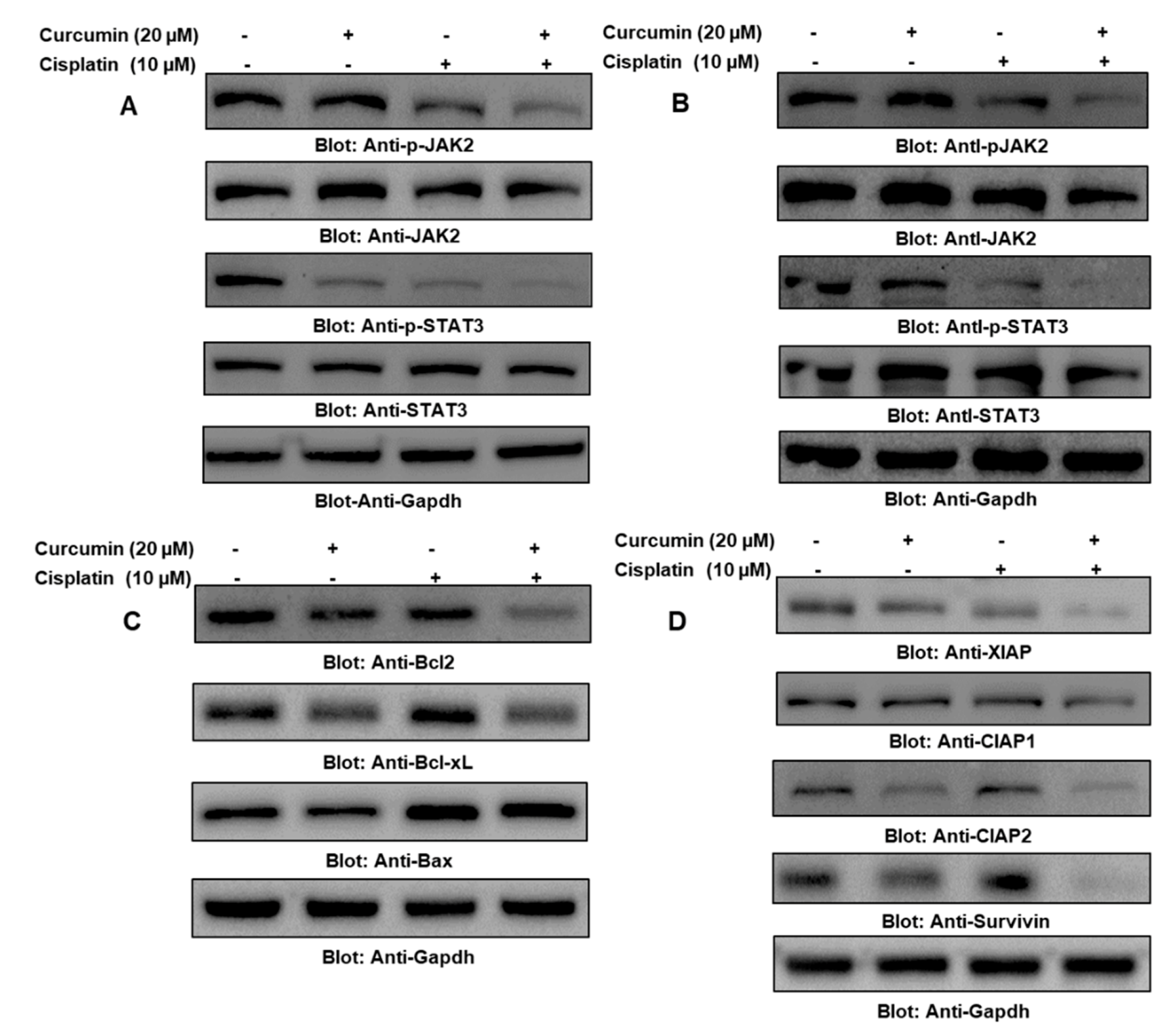
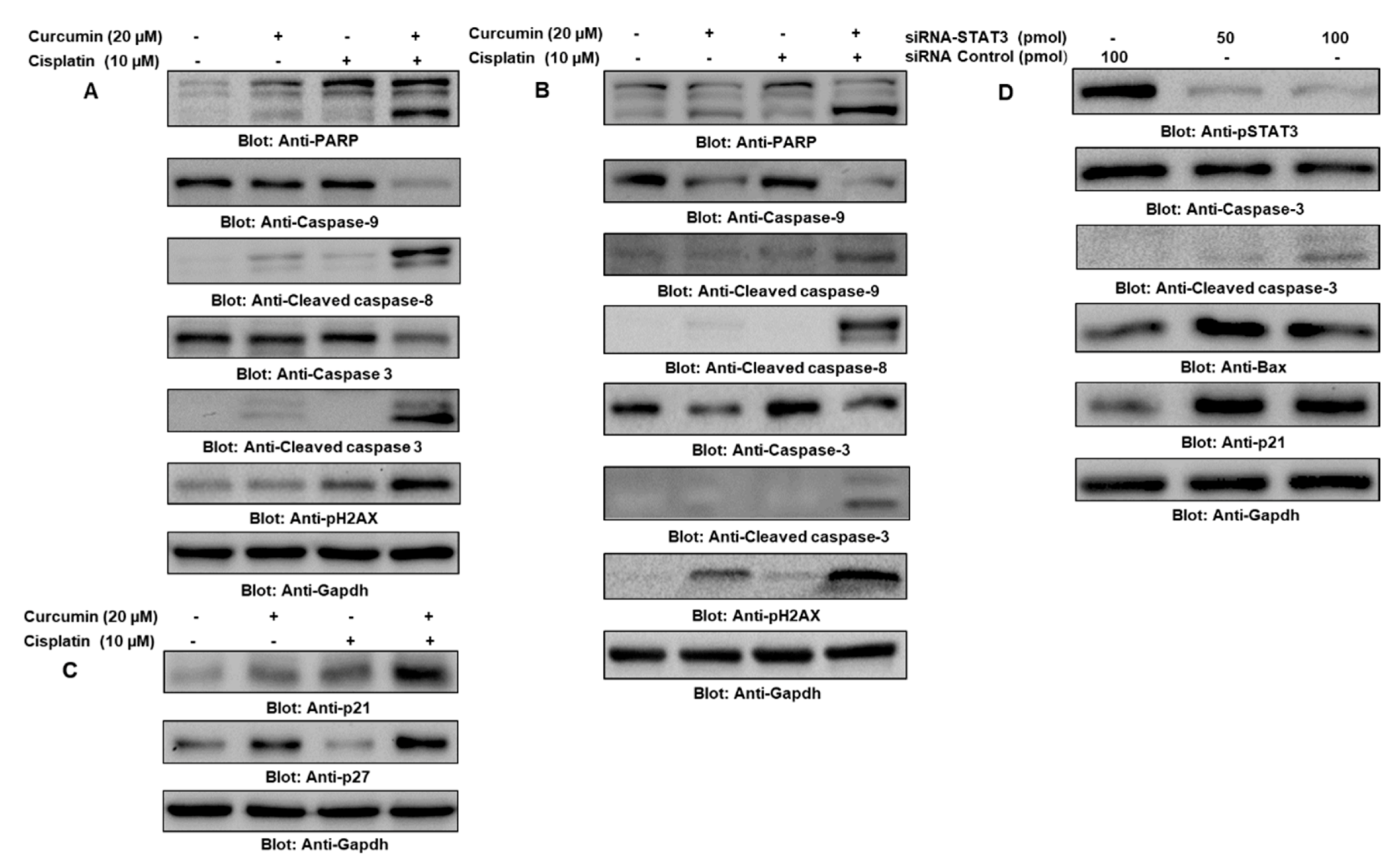
2.5. Cotreatment of Curcumin and Cisplatin Enhanced Apoptosis via the Downregulation of the JAK/STAT Survival Pathway in PTC Cells
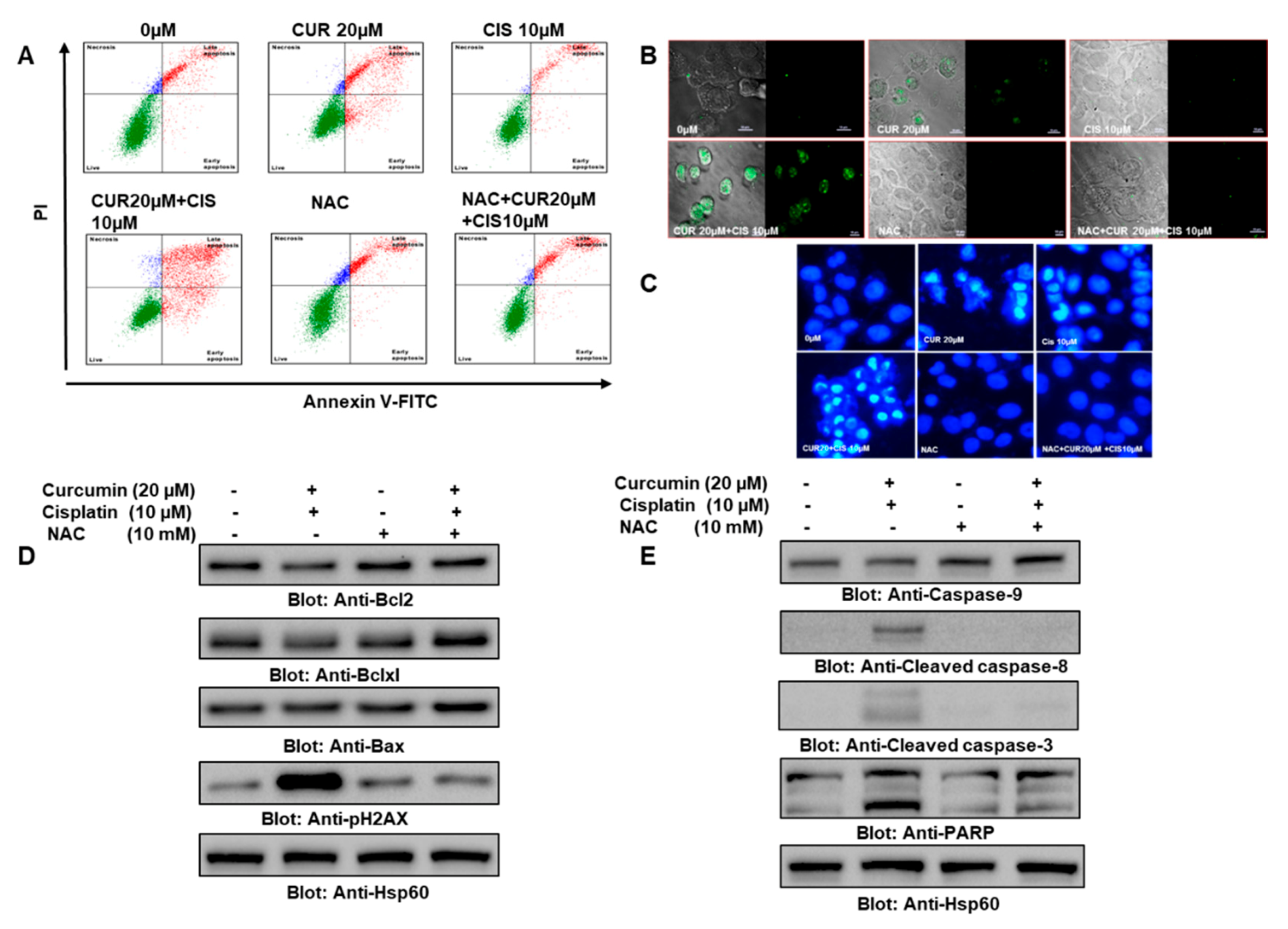
2.6. ROS Were Central in Curcumin- and Cisplatin Cotreatment-Induced Apoptosis in PTC Cells
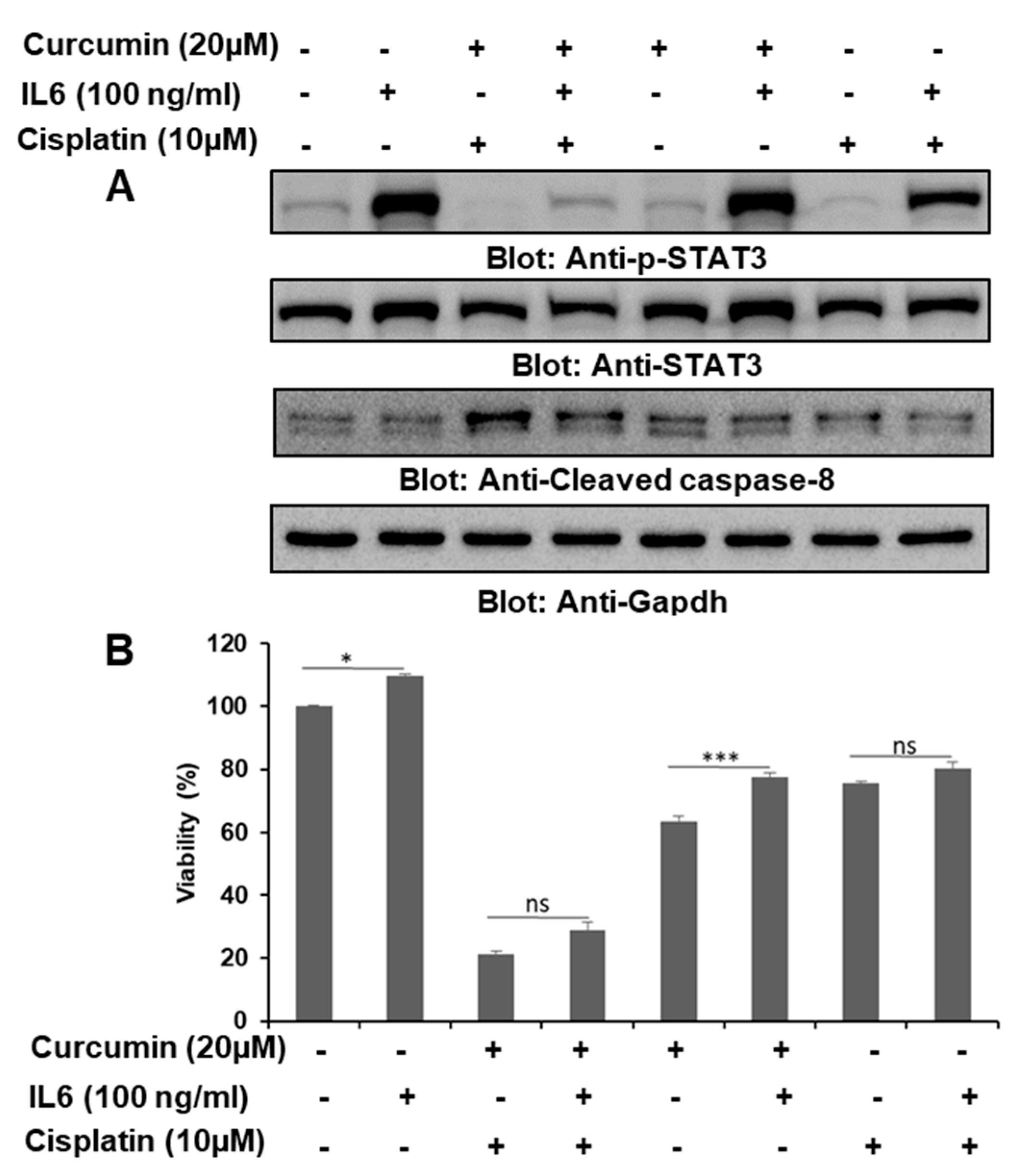
2.7. IL6 Was Important in the Curcumin- and Cisplatin-Induced Downregulation of p-STAT3
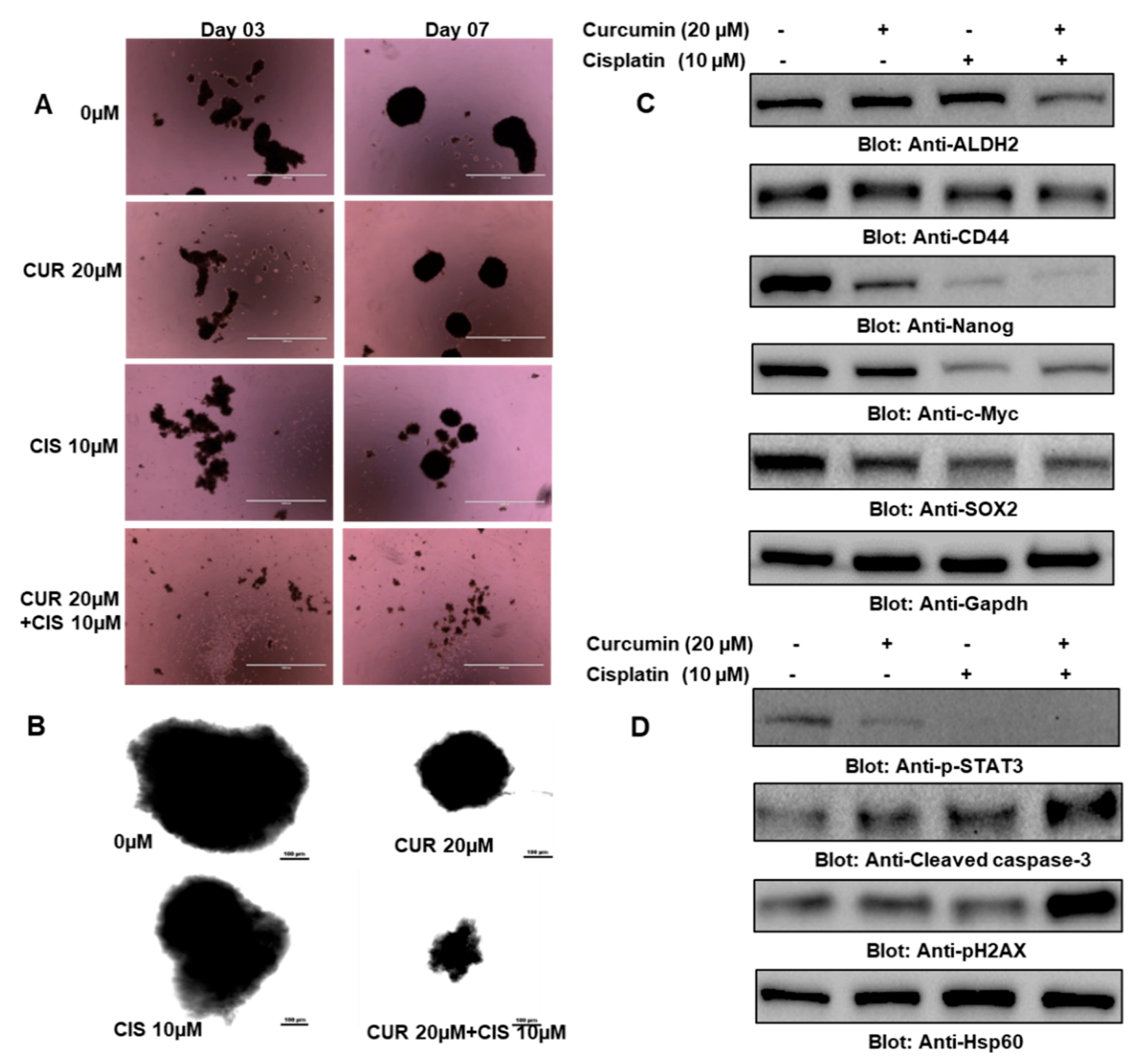
2.8. Cotreatment of Curcumin and Cisplatin Alleviated the Stemness of Thyroid Cancer Stem Cells (CSCs)
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Lines and Cell Culture
4.3. Measurement of Real-Time Cell Proliferation (Cell Index) of PTC Cells Treated with Curcumin, Cisplatin, and Curcumin + Cisplatin Using an RTCA xCELLigencecell Analyzer
4.4. Cell-Counting Kit-8 (CCK8) Assay
4.5. Cell Migration and Colonogenic Assay
4.6. Annexin V Staining
4.7. Cell Lysis and Immunoblotting
4.8. Measurement of Mitochondrial Membrane Potential
4.9. Gene Silencing Using siRNA
4.10. Thyrosphere-Forming Assay and Stemness Profile
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sosonkina, N.; Starenki, D.; Park, J.I. The role of stat3 in thyroid cancer. Cancers 2014, 6, 526–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprague, B.L.; Warren Andersen, S.; Trentham-Dietz, A. Thyroid cancer incidence and socioeconomic indicators of health care access. Cancer Causes Control 2008, 19, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Xing, M. Molecular pathogenesis and mechanisms of thyroid cancer. Nat. Rev. Cancer 2013, 13, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Zaballos, M.A.; Santisteban, P. Key signaling pathways in thyroid cancer. J. Endocrinol. 2017, 235, R43–R61. [Google Scholar] [CrossRef]
- Constantinescu, S.N.; Girardot, M.; Pecquet, C. Mining for jak-stat mutations in cancer. Trends Biochem. Sci 2008, 33, 122–131. [Google Scholar] [CrossRef]
- Fathi, N.; Rashidi, G.; Khodadadi, A.; Shahi, S.; Sharifi, S. Stat3 and apoptosis challenges in cancer. Int. J. Biol. Macromol. 2018, 117, 993–1001. [Google Scholar] [CrossRef]
- Li, M.; Yue, G.G.; Song, L.H.; Huang, M.B.; Lee, J.K.; Tsui, S.K.; Fung, K.P.; Tan, N.H.; Lau, C.B. Natural small molecule bigelovin suppresses orthotopic colorectal tumor growth and inhibits colorectal cancer metastasis via il6/stat3 pathway. Biochem. Pharmacol. 2018, 150, 191–201. [Google Scholar] [CrossRef]
- Shi, W.; Yan, D.; Zhao, C.; Xiao, M.; Wang, Y.; Ma, H.; Liu, T.; Qin, H.; Zhang, C.; Li, C.; et al. Inhibition of il-6/stat3 signaling in human cancer cells using evista. Biochem. Biophys. Res. Commun. 2017, 491, 159–165. [Google Scholar] [CrossRef]
- Yan, L.I.; Li, L.I.; Li, Q.; Di, W.; Shen, W.; Zhang, L.; Guo, H. Expression of signal transducer and activator of transcription 3 and its phosphorylated form is significantly upregulated in patients with papillary thyroid cancer. Exp. Ther. Med. 2015, 9, 2195–2201. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gill, A.; Atmore, B.; Johns, A.; Delbridge, L.; Lai, R.; McMullen, T. Upregulation of the signal transducers and activators of transcription 3 (stat3) pathway in lymphatic metastases of papillary thyroid cancer. Int. J. Clin. Exp. Pathol. 2011, 4, 356–362. [Google Scholar]
- Giuffrida, R.; Adamo, L.; Iannolo, G.; Vicari, L.; Giuffrida, D.; Eramo, A.; Gulisano, M.; Memeo, L.; Conticello, C. Resistance of papillary thyroid cancer stem cells to chemotherapy. Oncol. Lett. 2016, 12, 687–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.; Minsky, N.; Morshed, S.A.; Davies, T.F. Stemness in human thyroid cancers and derived cell lines: The role of asymmetrically dividing cancer stem cells resistant to chemotherapy. J. Clin. Endocrinol. Metab. 2014, 99, E400–E409. [Google Scholar] [CrossRef] [PubMed]
- Hardin, H.; Zhang, R.; Helein, H.; Buehler, D.; Guo, Z.; Lloyd, R.V. The evolving concept of cancer stem-like cells in thyroid cancer and other solid tumors. Lab. Investig. 2017, 97, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Hardin, H.; Lloyd, R.V. Cancer stem-like cells and thyroid cancer. Endocr. Relat. Cancer 2014, 21, T285–T300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golonko, A.; Lewandowska, H.; Swislocka, R.; Jasinska, U.T.; Priebe, W.; Lewandowski, W. Curcumin as tyrosine kinase inhibitor in cancer treatment. Eur. J. Med. Chem. 2019, 181, 111512. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Tastan, H.; Ozercan, I.H.; Guler, O.; Kahraman, N.; Kucuk, O.; Ozpolat, B. Chemopreventive and antitumor efficacy of curcumin in a spontaneously developing hen ovarian cancer model. Cancer Prev. Res. (Phila.) 2018, 11, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, F.; Shafiee, M.; Banikazemi, Z.; Pourhanifeh, M.H.; Khanbabaei, H.; Shamshirian, A.; Amiri Moghadam, S.; ArefNezhad, R.; Sahebkar, A.; Avan, A.; et al. Curcumin inhibits nf-kb and wnt/beta-catenin pathways in cervical cancer cells. Pathol. Res. Pract. 2019, 215, 152556. [Google Scholar] [CrossRef]
- Kuttikrishnan, S.; Siveen, K.S.; Prabhu, K.S.; Khan, A.Q.; Ahmed, E.I.; Akhtar, S.; Ali, T.A.; Merhi, M.; Dermime, S.; Steinhoff, M.; et al. Curcumin induces apoptotic cell death via inhibition of pi3-kinase/akt pathway in b-precursor acute lymphoblastic leukemia. Front. Oncol. 2019, 9, 484. [Google Scholar] [CrossRef]
- Seo, S.U.; Woo, S.M.; Lee, H.S.; Kim, S.H.; Min, K.J.; Kwon, T.K. Mtorc1/2 inhibitor and curcumin induce apoptosis through lysosomal membrane permeabilization-mediated autophagy. Oncogene 2018, 37, 5205–5220. [Google Scholar] [CrossRef] [Green Version]
- Wan Mohd Tajuddin, W.N.B.; Lajis, N.H.; Abas, F.; Othman, I.; Naidu, R. Mechanistic understanding of curcumin’s therapeutic effects in lung cancer. Nutrients 2019, 11, 2989. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.; Zeng, S.; Zhang, X.; Zhao, J.; Zhang, X.; Chen, X.; Yang, W.; Yang, Y.; Dong, Z.; et al. The natural polyphenol curcumin induces apoptosis by suppressing stat3 signaling in esophageal squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 303. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhang, C.; Weng, Q.; Ye, B. Curcumin protects against acute renal injury by suppressing jak2/stat3 pathway in severe acute pancreatitis in rats. Exp. Ther. Med. 2017, 14, 1669–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.M.; Xu, R.; Huang, X.Y.; Cheng, S.M.; Huang, M.F.; Yue, H.Y.; Wang, X.; Zou, Y.; Lu, A.P.; Liu, D.Y. Curcumin suppressed activation of dendritic cells via jak/stat/socs signal in mice with experimental colitis. Front. Pharmacol. 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wu, J.; Ye, B.; Wang, Q.; Xie, X.; Shen, H. Protective effect of curcumin on tnbs-induced intestinal inflammation is mediated through the jak/stat pathway. BMC Complement. Altern. Med. 2016, 16, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.L.; Norhaizan, M.E. Curcumin combination chemotherapy: The implication and efficacy in cancer. Molecules 2019, 24, 2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Tian, D.; Qiao, X.; Li, J.; Zhang, L. Modulation of myb-induced nf-kb -stat3 signaling and resulting cisplatin resistance in ovarian cancer by dietary factors. J. Cell Physiol. 2019, 234, 21126–21134. [Google Scholar] [CrossRef]
- Baharuddin, P.; Satar, N.; Fakiruddin, K.S.; Zakaria, N.; Lim, M.N.; Yusoff, N.M.; Zakaria, Z.; Yahaya, B.H. Curcumin improves the efficacy of cisplatin by targeting cancer stem-like cells through p21 and cyclin d1-mediated tumour cell inhibition in non-small cell lung cancer cell lines. Oncol. Rep. 2016, 35, 13–25. [Google Scholar] [CrossRef]
- HemaIswarya, S.; Doble, M. Potential synergism of natural products in the treatment of cancer. Phytother. Res. 2006, 20, 239–249. [Google Scholar] [CrossRef]
- Lumachi, F.; Basso, S.M.; Orlando, R. Cytokines, thyroid diseases and thyroid cancer. Cytokine 2010, 50, 229–233. [Google Scholar] [CrossRef]
- Notarangelo, T.; Sisinni, L.; Trino, S.; Calice, G.; Simeon, V.; Landriscina, M. Il6/stat3 axis mediates resistance to braf inhibitors in thyroid carcinoma cells. Cancer Lett. 2018, 433, 147–155. [Google Scholar] [CrossRef]
- Kobawala, T.P.; Trivedi, T.I.; Gajjar, K.K.; Patel, D.H.; Patel, G.H.; Ghosh, N.R. Significance of interleukin-6 in papillary thyroid carcinoma. J. Thyroid Res. 2016, 2016, 6178921. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C. Drug combination studies and their synergy quantification using the chou-talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siironen, P.; Louhimo, J.; Nordling, S.; Ristimaki, A.; Maenpaa, H.; Haapiainen, R.; Haglund, C. Prognostic factors in papillary thyroid cancer: An evaluation of 601 consecutive patients. Tumour Biol. J Int. Soc. Oncodevelopmental Biol. Med. 2005, 26, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Hay, I.D. Papillary thyroid carcinoma. Endocrinol. Metab. Clin. N. Am. 1990, 19, 545–576. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, X.; Xu, S.; Bao, J.; Yu, H. Curcumin induces endoplasmic reticulum stress-associated apoptosis in human papillary thyroid carcinoma bcpap cells via disruption of intracellular calcium homeostasis. Medicine 2018, 97, e11095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cheng, X.; Gao, Y.; Zhang, C.; Bao, J.; Guan, H.; Yu, H.; Lu, R.; Xu, Q.; Sun, Y. Curcumin inhibits metastasis in human papillary thyroid carcinoma bcpap cells via down-regulation of the tgf-beta/smad2/3 signaling pathway. Exp. Cell Res. 2016, 341, 157–165. [Google Scholar] [CrossRef]
- Carpenter, R.L.; Lo, H.W. Stat3 target genes relevant to human cancers. Cancers 2014, 6, 897–925. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Ma, Q.; Li, L.; Wang, H. The cxcl12-cxcr4 axis promotes migration, invasiveness, and emt in human papillary thyroid carcinoma b-cpap cells via nf-kappab signaling. Biochem. Cell Biol. 2018, 96, 619–626. [Google Scholar] [CrossRef]
- Zhu, X.; Bai, Q.; Lu, Y.; Lu, Y.; Zhu, L.; Zhou, X.; Wu, L. Expression and function of cxcl12/cxcr4/cxcr7 in thyroid cancer. Int. J. Oncol. 2016, 48, 2321–2329. [Google Scholar] [CrossRef]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Borkhuu, O.; Bao, W.; Yang, Y.T. Signaling pathways in thyroid cancer and their therapeutic implications. J. Clin. Med. Res. 2016, 8, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunigal, S.; Lakka, S.S.; Sodadasu, P.K.; Estes, N.; Rao, J.S. Stat3-sirna induces fas-mediated apoptosis in vitro and in vivo in breast cancer. Int. J. Oncol. 2009, 34, 1209–1220. [Google Scholar] [PubMed]
- Aoki, Y.; Feldman, G.M.; Tosato, G. Inhibition of stat3 signaling induces apoptosis and decreases survivin expression in primary effusion lymphoma. Blood 2003, 101, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, J.; Prasad, S.; Aggarwal, B.B. Curcumin and cancer cells: How many ways can curry kill tumor cells selectively? AAPS J. 2009, 11, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ding, Y.; Ye, N.; Wild, C.; Chen, H.; Zhou, J. Direct activation of bax protein for cancer therapy. Med. Res. Rev. 2016, 36, 313–341. [Google Scholar] [CrossRef]
- Naseri, M.H.; Mahdavi, M.; Davoodi, J.; Tackallou, S.H.; Goudarzvand, M.; Neishabouri, S.H. Up regulation of bax and down regulation of bcl2 during 3-nc mediated apoptosis in human cancer cells. Cancer Cell Int. 2015, 15, 55. [Google Scholar] [CrossRef] [Green Version]
- Perlman, H.; Zhang, X.; Chen, M.W.; Walsh, K.; Buttyan, R. An elevated bax/bcl-2 ratio corresponds with the onset of prostate epithelial cell apoptosis. Cell Death Differ. 1999, 6, 48–54. [Google Scholar] [CrossRef]
- Al Zaid Siddiquee, K.; Turkson, J. Stat3 as a target for inducing apoptosis in solid and hematological tumors. Cell Res. 2008, 18, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Buettner, R.; Mora, L.B.; Jove, R. Activated stat signaling in human tumors provides novel molecular targets for therapeutic intervention. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2002, 8, 945–954. [Google Scholar]
- Zaanan, A.; Okamoto, K.; Kawakami, H.; Khazaie, K.; Huang, S.; Sinicrope, F.A. The mutant kras gene up-regulates bcl-xl protein via stat3 to confer apoptosis resistance that is reversed by bim protein induction and bcl-xl antagonism. J. Biol. Chem. 2015, 290, 23838–23849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Ray, R.M.; Johnson, L.R. Stat3-mediated transcription of bcl-2, mcl-1 and c-iap2 prevents apoptosis in polyamine-depleted cells. Biochem. J. 2005, 392, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynh, J.; Etemadi, N.; Hollande, F.; Ernst, M.; Buchert, M. The jak/stat3 axis: A comprehensive drug target for solid malignancies. Semin. Cancer Biol. 2017, 45, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.H.; Choi, J.H.; Chung, Y.Y.; Lim, G.L.; Park, Y.N.; Park, S.W. Predominant activation of jak/stat3 pathway by interleukin-6 is implicated in hepatocarcinogenesis. Neoplasia 2015, 17, 586–597. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, S.I.A.; Jantan, I.; Haque, M.A. Naturally occurring immunomodulators with antitumor activity: An insight on their mechanisms of action. Int. Immunopharmacol 2017, 50, 291–304. [Google Scholar] [CrossRef]
- Shafabakhsh, R.; Pourhanifeh, M.H.; Mirzaei, H.R.; Sahebkar, A.; Asemi, Z.; Mirzaei, H. Targeting regulatory t cells by curcumin: A potential for cancer immunotherapy. Pharmacol. Res. 2019, 147, 104353. [Google Scholar] [CrossRef]
- Xu, B.; Yu, L.; Zhao, L.Z. Curcumin up regulates t helper 1 cells in patients with colon cancer. Am. J. Transl. Res. 2017, 9, 1866–1875. [Google Scholar]
- Wang, Z.; Zhang, K.; Zhu, Y.; Wang, D.; Shao, Y.; Zhang, J. Curcumin inhibits hypoxia-induced proliferation and invasion of mg-63 osteosarcoma cells via downregulating notch1. Mol. Med. Rep. 2017, 15, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Long, Q.; Zhang, L.; Shi, Y.; Liu, X.; Li, X.; Guan, B.; Tian, Y.; Wang, X.; Li, L.; et al. Curcumin inhibits cancer-associated fibroblast-driven prostate cancer invasion through maoa/mtor/hif-1alpha signaling. Int. J. Oncol. 2015, 47, 2064–2072. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Chang, Y.; Li, R.; Xu, Q.; Lei, J.; Yin, C.; Li, T.; Wu, Y.; Ma, Q.; Li, X. Curcumin inhibits hypoxia inducible factor1alphainduced epithelialmesenchymal transition in hepg2 hepatocellular carcinoma cells. Mol. Med. Rep. 2014, 10, 2505–2510. [Google Scholar] [CrossRef]
- Ye, M.X.; Zhao, Y.L.; Li, Y.; Miao, Q.; Li, Z.K.; Ren, X.L.; Song, L.Q.; Yin, H.; Zhang, J. Curcumin reverses cis-platin resistance and promotes human lung adenocarcinoma a549/ddp cell apoptosis through hif-1alpha and caspase-3 mechanisms. Phytomedicine 2012, 19, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The xcelligence system for real-time and label-free monitoring of cell viability. Methods Mol. Biol. 2011, 740, 33–43. [Google Scholar] [PubMed]
- Khan, A.Q.; Siveen, K.S.; Prabhu, K.S.; Kuttikrishnan, S.; Akhtar, S.; Shaar, A.; Raza, A.; Mraiche, F.; Dermime, S.; Uddin, S. Curcumin-Mediated Degradation of S-Phase Kinase Protein 2 Induces Cytotoxic Effects in Human Papillomavirus-Positive and Negative Squamous Carcinoma Cells. Front. Oncol. 2018, 8, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Asmari, A.K.; Albalawi, S.M.; Athar, M.T.; Khan, A.Q.; Al-Shahrani, H.; Islam, M. Moringa oleifera as an anti-cancer agent against breast and colorectal cancer cell lines. PLoS ONE 2015, 10, e0135814. [Google Scholar] [CrossRef]
- Iskandarani, A.; Bhat, A.A.; Siveen, K.S.; Prabhu, K.S.; Kuttikrishnan, S.; Khan, M.A.; Krishnankutty, R.; Kulinski, M.; Nasr, R.R.; Mohammad, R.M.; et al. Bortezomib-mediated downregulation of s-phase kinase protein-2 (skp2) causes apoptotic cell death in chronic myelogenous leukemia cells. J. Transl. Med. 2016, 14, 69. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, K.S.; Siveen, K.S.; Kuttikrishnan, S.; Iskandarani, A.; Tsakou, M.; Achkar, I.W.; Therachiyil, L.; Krishnankutty, R.; Parray, A.; Kulinski, M.; et al. Targeting of x-linked inhibitor of apoptosis protein and pi3-kinase/akt signaling by embelin suppresses growth of leukemic cells. PLoS ONE 2017, 12, e0180895. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.R.; Al-Rasheed, M.; Manogaran, P.S.; Al-Hussein, K.A.; Platanias, L.C.; Al Kuraya, K.; Uddin, S. Curcumin induces apoptosis via inhibition of pi3’-kinase/akt pathway in acute t cell leukemias. Apoptosis 2006, 11, 245–254. [Google Scholar] [CrossRef]
- Caria, P.; Pillai, R.; Dettori, T.; Frau, D.V.; Zavattari, P.; Riva, G.; Romano, G.; Pani, F.; Bentivegna, A.; Giovannoni, R.; et al. Thyrospheres from b-cpap cell line with braf and tert promoter mutations have different functional and molecular features than parental cells. J. Cancer 2017, 8, 1629–1639. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.Q.; Ahmed, E.I.; Elareer, N.; Fathima, H.; Prabhu, K.S.; Siveen, K.S.; Kulinski, M.; Azizi, F.; Dermime, S.; Ahmad, A.; et al. Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020438
Khan AQ, Ahmed EI, Elareer N, Fathima H, Prabhu KS, Siveen KS, Kulinski M, Azizi F, Dermime S, Ahmad A, et al. Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway. International Journal of Molecular Sciences. 2020; 21(2):438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020438
Chicago/Turabian StyleKhan, Abdul Q., Eiman I. Ahmed, Noor Elareer, Hamna Fathima, Kirti S. Prabhu, Kodappully S. Siveen, Michal Kulinski, Fouad Azizi, Said Dermime, Aamir Ahmad, and et al. 2020. "Curcumin-Mediated Apoptotic Cell Death in Papillary Thyroid Cancer and Cancer Stem-Like Cells through Targeting of the JAK/STAT3 Signaling Pathway" International Journal of Molecular Sciences 21, no. 2: 438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020438