TBX1 and Basal Cell Carcinoma: Expression and Interactions with Gli2 and Dvl2 Signaling

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Recruitment of BCC Cases
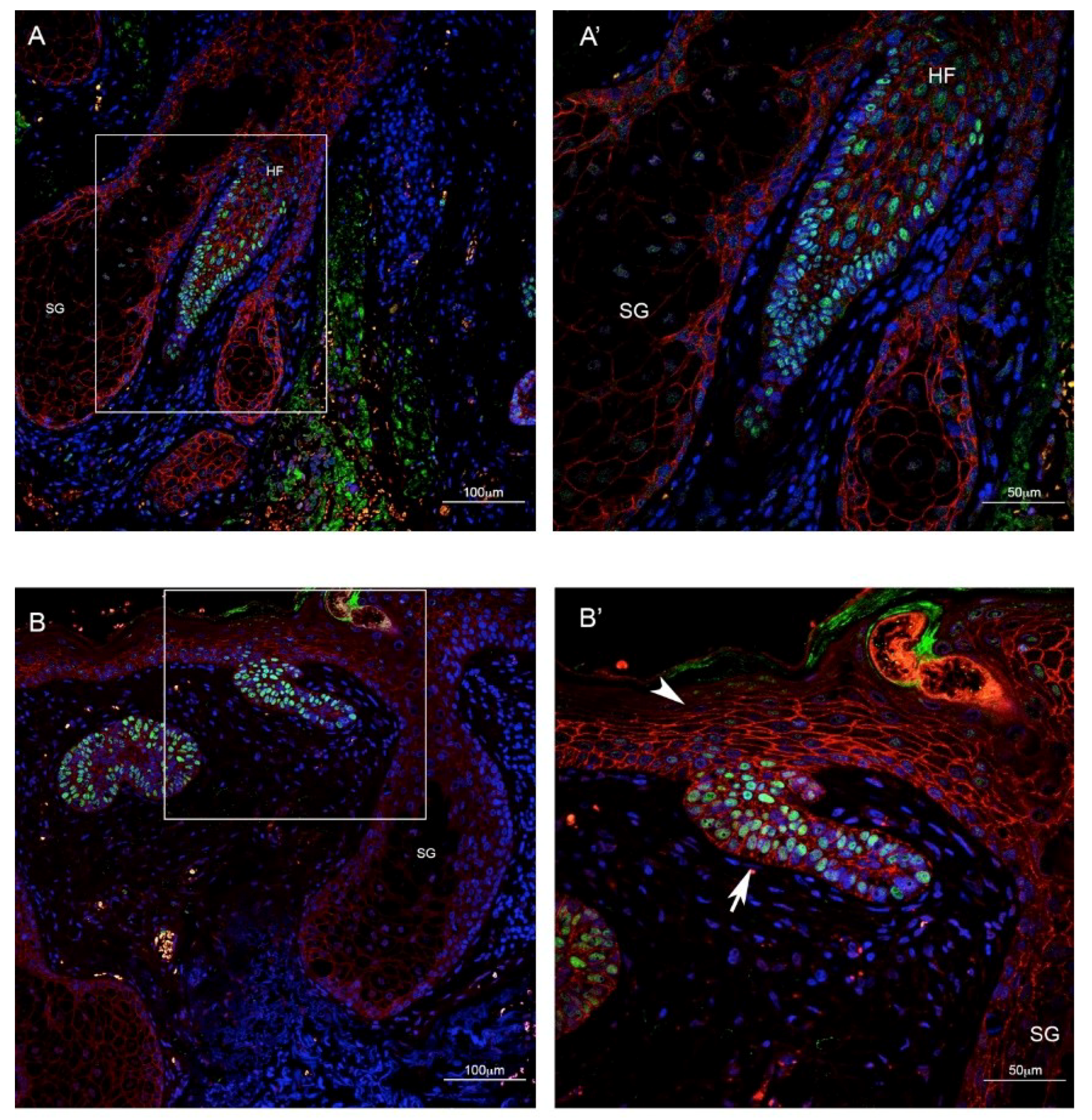
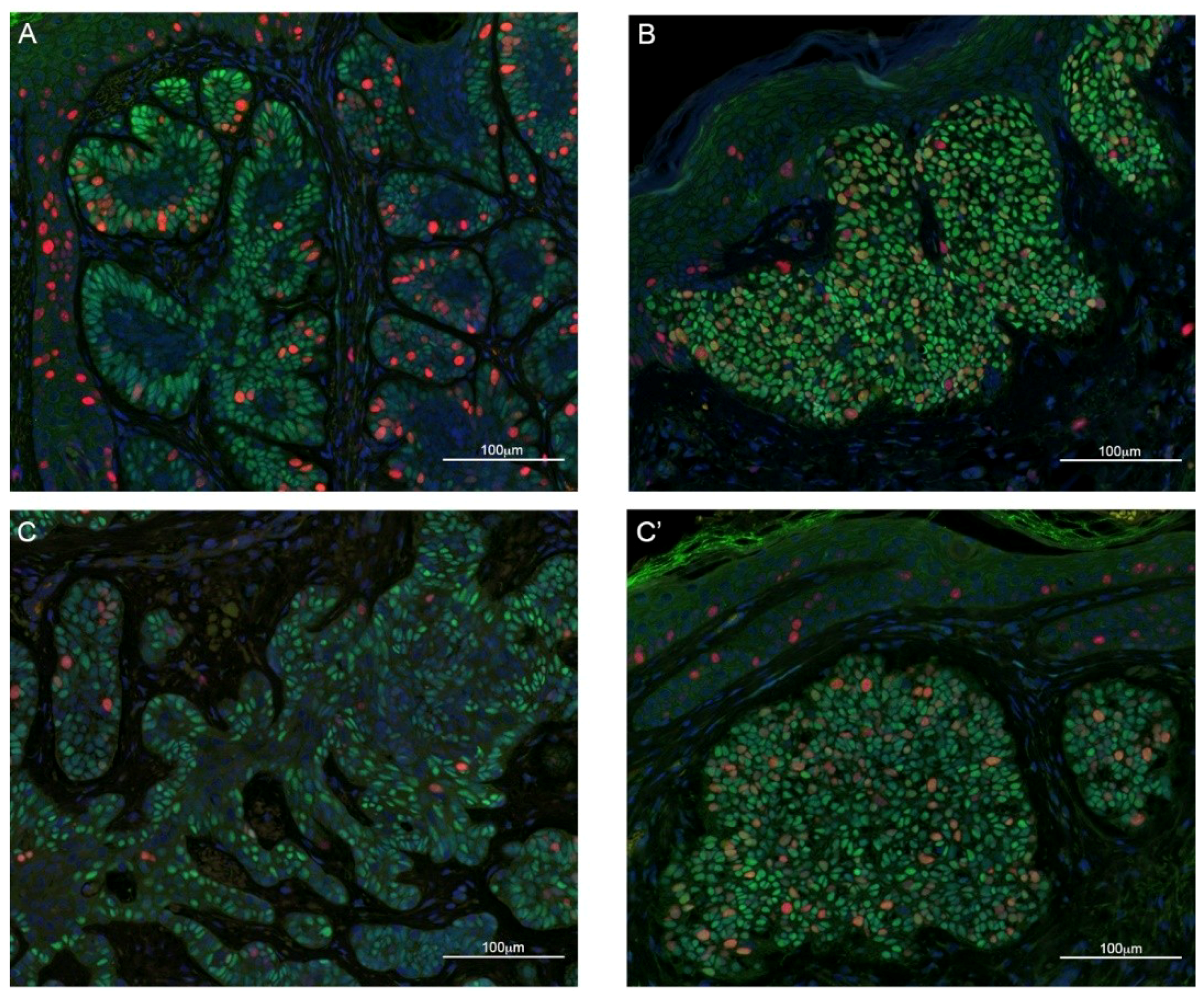
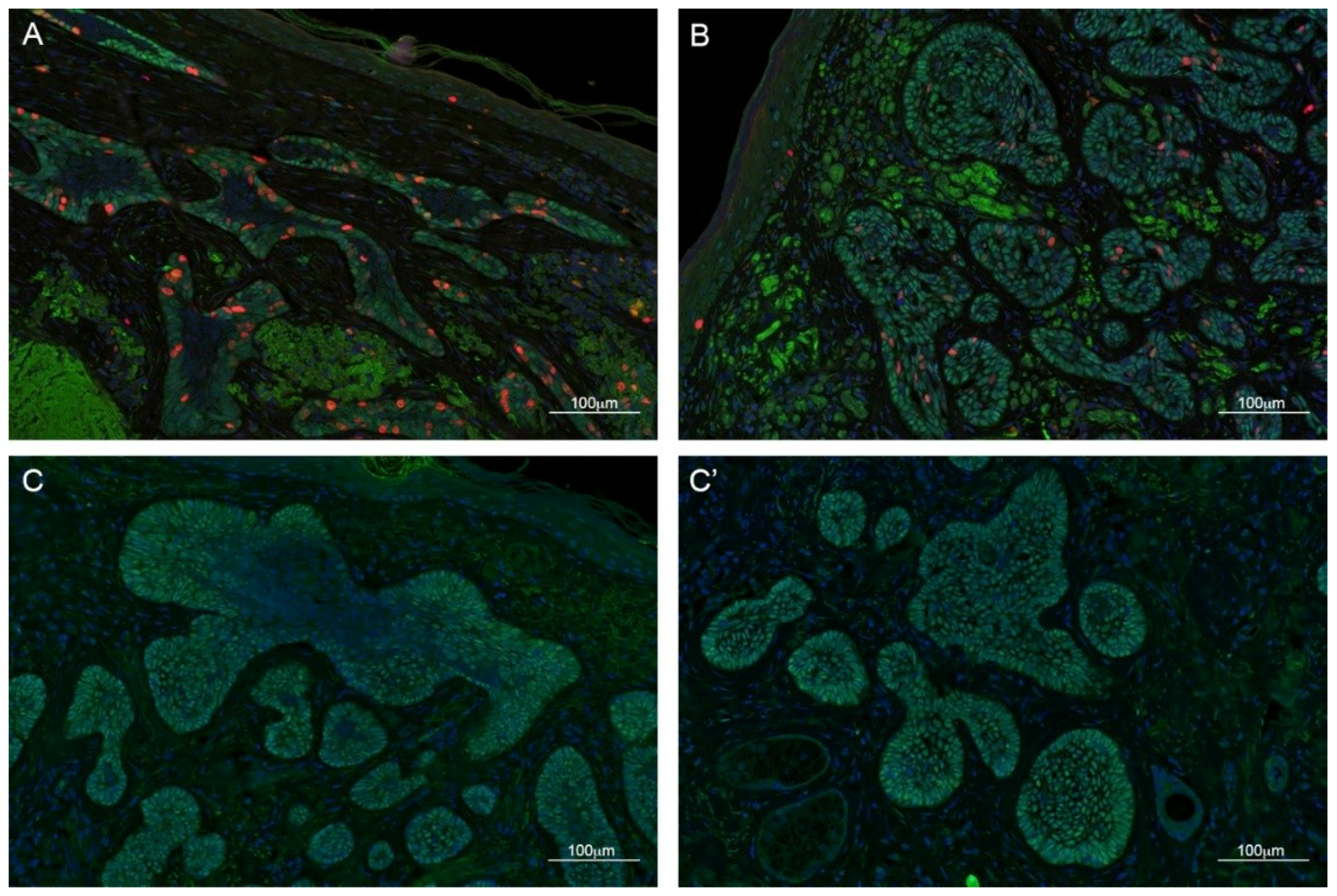
2.2. TBX1 is Expressed in the Hair Follicle and in BCC
2.3. TBX1 Expression and BCC Histopathology
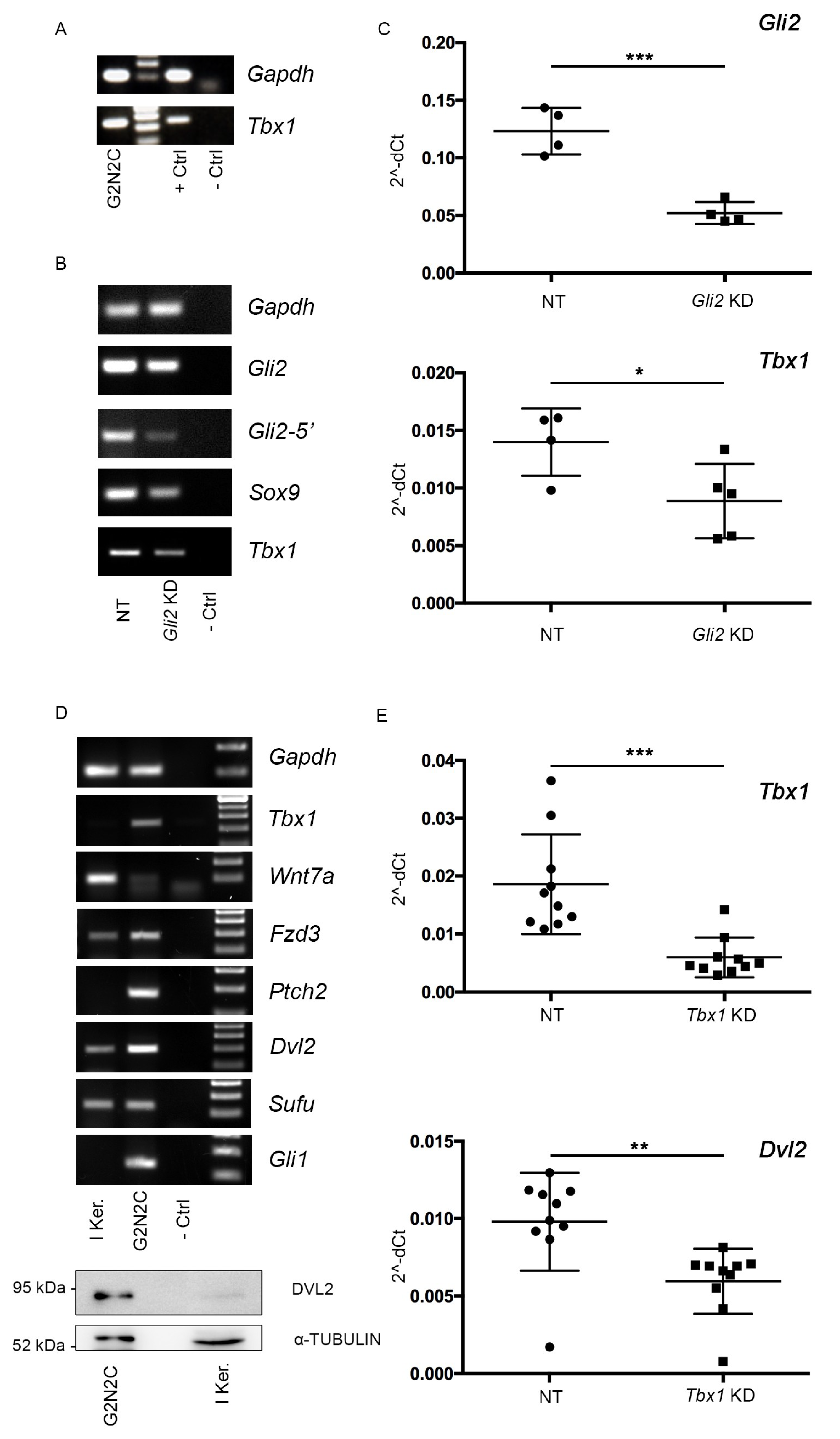
2.4. A Murine Cellular Model of BCC Suggests a Tbx1-Dependent Pathway
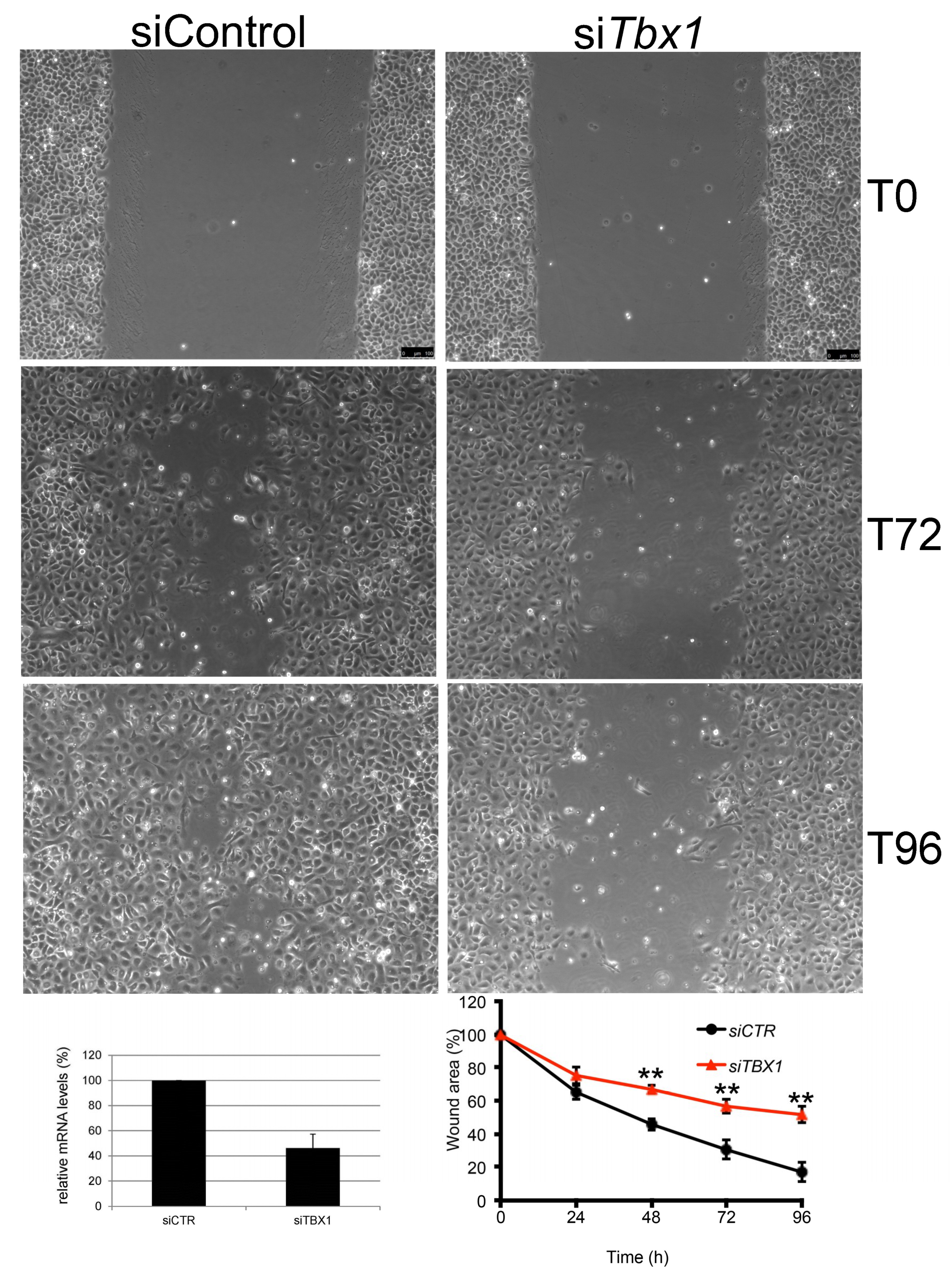
2.5. TBX1 is Important for Migration of G2N2C Cells
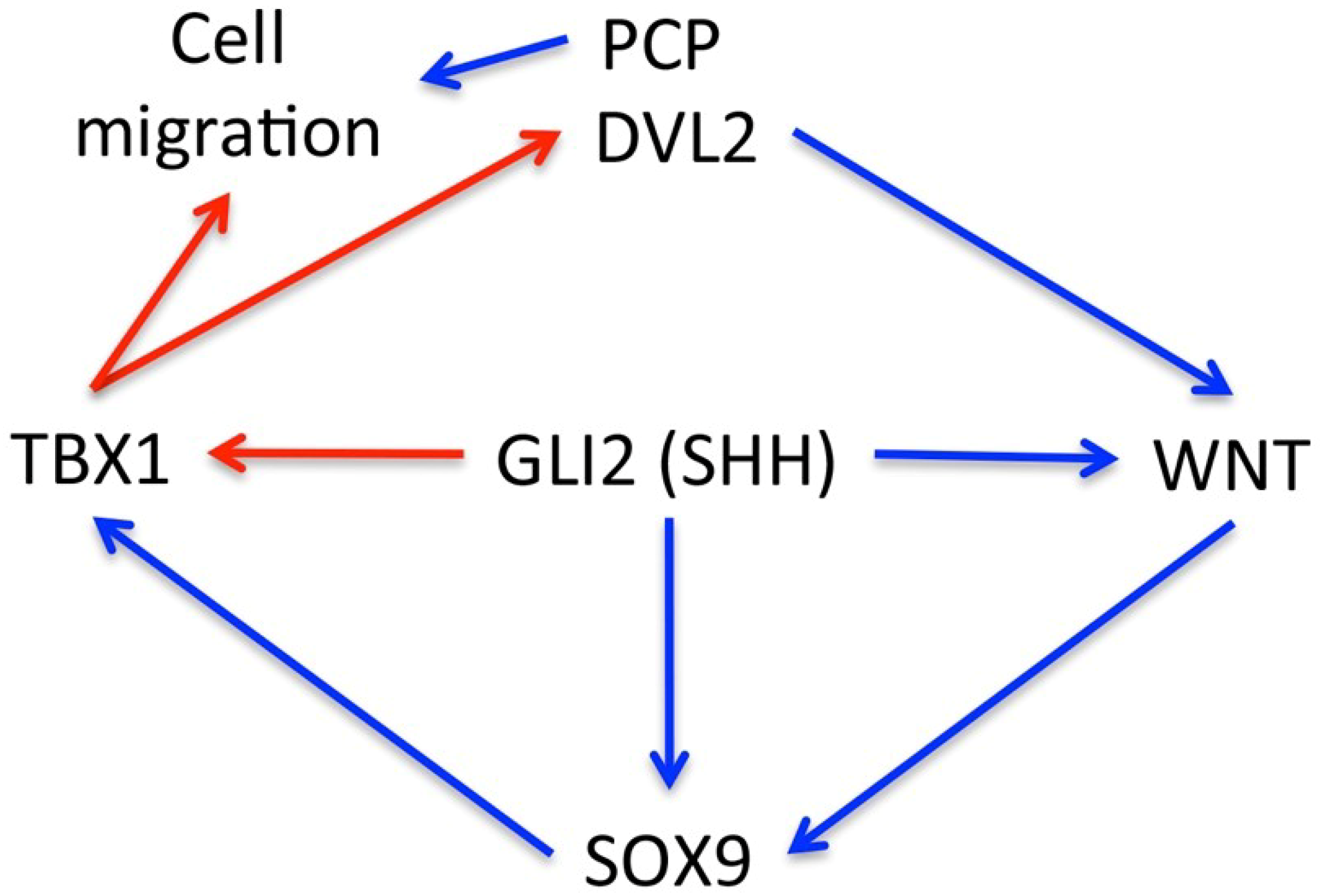
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Tissue Microarray Construction
4.3. Immunofluorescence
4.4. Cell Culture Experiments
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pellegrini, C.; Maturo, M.G.; Di Nardo, L.; Ciciarelli, V.; García-Rodrigo, C.G.; Fargnoli, M.C. Understanding the Molecular Genetics of Basal Cell Carcinoma. Int. J. Mol. Sci. 2017, 18, 2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, K.K.; Van Keymeulen, A.; Lapouge, G.; Beck, B.; Michaux, C.; Achouri, Y.; Sotiropoulou, P.A.; Blanpain, C. Identification of the cell lineage at the origin of basal cell carcinoma. Nat. Cell Biol. 2010, 12, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Youssef, K.K.; Lapouge, G.; Bouvrée, K.; Rorive, S.; Brohée, S.; Appelstein, O.; Larsimont, J.-C.; Sukumaran, V.; Van de Sande, B.; Pucci, D.; et al. Adult interfollicular tumour-initiating cells are reprogrammed into an embryonic hair follicle progenitor-like fate during basal cell carcinoma initiation. Nat. Cell Biol. 2012, 14, 1282–1294. [Google Scholar] [CrossRef] [PubMed]
- Grachtchouk, M.; Pero, J.; Yang, S.H.; Ermilov, A.N.; Michael, L.E.; Wang, A.; Wilbert, D.; Patel, R.M.; Ferris, J.; Diener, J.; et al. Basal cell carcinomas in mice arise from hair follicle stem cells and multiple epithelial progenitor populations. J. Clin. Investig. 2011, 121, 1768–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, S.C.; Eberl, M.; Vagnozzi, A.N.; Belkadi, A.; Veniaminova, N.A.; Verhaegen, M.E.; Bichakjian, C.K.; Ward, N.L.; Dlugosz, A.A.; Wong, S.Y. Basal cell carcinoma preferentially arises from stem cells within hair follicle and mechanosensory niches. Cell Stem Cell 2015, 16, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Y.; Wang, J.; Mancianti, M.-L.; Epstein, E.H. Basal cell carcinomas arise from hair follicle stem cells in Ptch1(+/-) mice. Cancer Cell 2011, 19, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Bichakjian, C.K.; Olencki, T.; Aasi, S.Z.; Alam, M.; Andersen, J.S.; Berg, D.; Bowen, G.M.; Cheney, R.T.; Daniels, G.A.; Glass, L.F.; et al. Basal Cell Skin Cancer, Version 1.2016, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2016, 14, 574–597. [Google Scholar] [CrossRef]
- Fulcoli, F.G.; Franzese, M.; Liu, X.; Zhang, Z.; Angelini, C.; Baldini, A. Rebalancing gene haploinsufficiency in vivo by targeting chromatin. Nat. Commun. 2016, 7, 11688. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Fulcoli, F.G.; Ferrentino, R.; Martucciello, S.; Illingworth, E.A.; Baldini, A. Transcriptional control in cardiac progenitors: Tbx1 interacts with the BAF chromatin remodeling complex and regulates Wnt5a. Plos Genet. 2012, 8, e1002571. [Google Scholar] [CrossRef] [Green Version]
- Racedo, S.E.; Hasten, E.; Lin, M.; Devakanmalai, G.S.; Guo, T.; Ozbudak, E.M.; Cai, C.-L.; Zheng, D.; Morrow, B.E. Reduced dosage of β-catenin provides significant rescue of cardiac outflow tract anomalies in a Tbx1 conditional null mouse model of 22q11.2 deletion syndrome. Plos Genet. 2017, 13, e1006687. [Google Scholar] [CrossRef]
- Yamagishi, H.; Maeda, J.; Hu, T.; McAnally, J.; Conway, S.J.; Kume, T.; Meyers, E.N.; Yamagishi, C.; Srivastava, D. Tbx1 is regulated by tissue-specific forkhead proteins through a common Sonic hedgehog-responsive enhancer. Genes Dev. 2003, 17, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsimont, J.-C.; Youssef, K.K.; Sánchez-Danés, A.; Sukumaran, V.; Defrance, M.; Delatte, B.; Liagre, M.; Baatsen, P.; Marine, J.-C.; Lippens, S.; et al. Sox9 Controls Self-Renewal of Oncogene Targeted Cells and Links Tumor Initiation and Invasion. Cell Stem Cell 2015, 17, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Heller, E.; Beronja, S.; Oshimori, N.; Stokes, N.; Fuchs, E. An RNA interference screen uncovers a new molecule in stem cell self-renewal and long-term regeneration. Nature 2012, 485, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dentice, M.; Luongo, C.; Huang, S.; Ambrosio, R.; Elefante, A.; Mirebeau-Prunier, D.; Zavacki, A.M.; Fenzi, G.; Grachtchouk, M.; Hutchin, M.; et al. Sonic hedgehog-induced type 3 deiodinase blocks thyroid hormone action enhancing proliferation of normal and malignant keratinocytes. Proc. Natl. Acad. Sci. USA. 2007, 104, 14466–14471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, H.; Goich, S.; Wang, A.; Grachtchouk, M.; Lowe, L.; Mo, R.; Lin, K.; de Sauvage, F.J.; Sasaki, H.; Hui, C.; et al. Dissecting the oncogenic potential of Gli2: Deletion of an NH(2)-terminal fragment alters skin tumor phenotype. Cancer Res. 2002, 62, 5308–5316. [Google Scholar] [PubMed]
- Sharma, M.; Castro-Piedras, I.; Simmons, G.E.; Pruitt, K. Dishevelled: A masterful conductor of complex Wnt signals. Cell. Signal. 2018, 47, 52–64. [Google Scholar] [CrossRef]
- Alfano, D.; Altomonte, A.; Cortes, C.; Bilio, M.; Kelly, R.G.; Baldini, A. Tbx1 regulates extracellular matrix-cell interactions in the second heart field. Hum. Mol. Genet. 2019, 28, 2295–2308. [Google Scholar] [CrossRef]
- Atwood, S.X.; Whitson, R.J.; Oro, A.E. Advanced treatment for basal cell carcinomas. Cold Spring Harb. Perspect. Med. 2014, 4, a013581. [Google Scholar] [CrossRef]
- Sekulic, A.; Migden, M.R.; Oro, A.E.; Dirix, L.; Lewis, K.D.; Hainsworth, J.D.; Solomon, J.A.; Yoo, S.; Arron, S.T.; Friedlander, P.A.; et al. Efficacy and safety of vismodegib in advanced basal-cell carcinoma. N. Engl. J. Med. 2012, 366, 2171–2179. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Danés, A.; Larsimont, J.-C.; Liagre, M.; Muñoz-Couselo, E.; Lapouge, G.; Brisebarre, A.; Dubois, C.; Suppa, M.; Sukumaran, V.; Del Marmol, V.; et al. A slow-cycling LGR5 tumour population mediates basal cell carcinoma relapse after therapy. Nature 2018, 562, 434–438. [Google Scholar] [CrossRef]
- Bien-Willner, G.A.; Stankiewicz, P.; Lupski, J.R. SOX9cre1, a cis-acting regulatory element located 1.1 Mb upstream of SOX9, mediates its enhancement through the SHH pathway. Hum. Mol. Genet. 2007, 16, 1143–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, V.E.; Gordon, J.; O’Neil, J.D.; Ramos, I.; Richie, E.R.; Manley, N.R. Tissue-specific roles for sonic hedgehog signaling in establishing thymus and parathyroid organ fate. Dev. Camb. Engl. 2016, 143, 4027–4037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, V.; Yamagishi, C.; Hu, T.; Kathiriya, I.S.; Yamagishi, H.; Srivastava, D. Tbx1, a DiGeorge syndrome candidate gene, is regulated by sonic hedgehog during pharyngeal arch development. Dev. Biol. 2001, 235, 62–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trempus, C.S.; Wei, S.J.; Humble, M.M.; Dang, H.; Bortner, C.D.; Sifre, M.I.; Kissling, G.E.; Sunman, J.A.; Akiyama, S.K.; Roberts, J.D.; et al. A novel role for the T-box transcription factor Tbx1 as a negative regulator of tumor cell growth in mice. Mol. Carcinog. 2011, 50, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Kump, E.; Wernli, M.; Erb, P. Gene silencing of transcription factor Gli2 inhibits basal cell carcinomalike tumor growth in vivo. Int. J. Cancer 2008, 122, 50–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histological Subtype | n |
|---|---|
| BCC with aggressive growth | 27 |
| Basosquamous Carcinoma | 2 |
| Infiltrating BCC | 17 |
| Micronodular BCC | 2 |
| Sclerosing/morphoeic BCC | 6 |
| BCC with indolent growth | 24 |
| BCC with adnexal differentiation | 5 |
| Nodular BCC | 15 |
| Superficial BCC | 4 |
| Total | 51 |
| Age | Mean ± SD | 66.5 ± 13.2 years | |
| Range (Min–Max) | 38–87 years | ||
| Sex | Male | n = 36 | 71% |
| Female | n = 15 | 29% | |
| Tumor Site | Area H | n = 18 | 35% |
| Area M | n = 6 | 12% | |
| Area L | n = 24 | 47% | |
| ND | n = 3 | 6% | |
| Histologic Subtype | BCC with indolent growth | n = 24 | 47% |
| BCC with aggressive growth | n = 27 | 53% | |
| Tumor Size | >2 cm | n = 7 | 14% |
| <2 cm | n =36 | 71% | |
| ND | n = 8 | 16% |
| Sample Code | Histological Type | Differentiation Sstate | TBX1+ Cells | TBX1 Signal |
|---|---|---|---|---|
| 15 | Nodular BCC | differentiated | D | 2 |
| BAS59 | Nodular BCC | differentiated | D + P | 3 |
| BAS60 | BCC with adnexal differentiation | differentiated | D + P | 3 |
| BAS5 | Nodular BCC | differentiated | D | 3 |
| BAS11 | Nodular BCC | differentiated | P | 2 |
| BAS14 | Superficial BCC | differentiated | D | 3 |
| BAS24 | Nodular BCC | differentiated | D + P | 3 |
| BAS29 | Superficial BCC | differentiated | D | 2 |
| BAS38 | Nodular BCC | differentiated | P | 1 |
| BAS40 | Nodular BCC | differentiated | P | 1 |
| BAS41 | Nodular BCC | differentiated | P + D | 2 |
| BAS42 | Nodular BCC | differentiated | D | 3 |
| A | BCC with adnexal differentiation | differentiated | D | 1 |
| 8 | Nodular BCC | differentiated | D | 2 |
| 2 | Nodular BCC | differentiated | D + P | 3 |
| B | Superficial BCC | differentiated | P | 2 |
| E | Superficial BCC | differentiated | D | 3 |
| G | BCC with adnexal differentiation | differentiated | P | 2 |
| I | Nodular BCC | differentiated | P | 2 |
| L | BCC with adnexal differentiation | differentiated | P | 2 |
| M | Nodular BCC | differentiated | D + P | 3 |
| n | Nodular BCC | differentiated | P + D | 2 |
| R | Nodular BCC | differentiated | D | 3 |
| S | BCC with adnexal differentiation | differentiated | D + P | 3 |
| BAS7 | Micronodular BCC | undifferentiated | P | 2 |
| BAS8 | Infiltrating BCC | undifferentiated | D | 1 |
| BAS9 | Basosquamous Carcinoma | undifferentiated | P | 1 |
| BAS12 | Infiltrating BCC | undifferentiated | P + D | 2 |
| BAS13 | Infiltrating BCC | undifferentiated | D | 2 |
| BAS19 | Infiltrating BCC | undifferentiated | D | 3 |
| BAS67 | Sclerosing/morphoeic BCC | undifferentiated | D | 1 |
| BAS68 | Micronodular BCC | undifferentiated | D | 3 |
| BAS69 | Infiltrating BCC | undifferentiated | P + D | 1 |
| BAS28 | Infiltrating BCC | undifferentiated | D | 1 |
| BAS44 | Basosquamous Carcinoma | undifferentiated | D | 2 |
| BAS47 | Sclerosing/morphoeic BCC | undifferentiated | P + D | 2 |
| BAS48 | Infiltrating BCC | undifferentiated | P + D | 2 |
| BAS51 | Infiltrating BCC | undifferentiated | D + P | 3 |
| 12 | Infiltrating BCC | undifferentiated | D | 2 |
| 3 | Infiltrating BCC | undifferentiated | D | 2 |
| 1 | Infiltrating BCC | undifferentiated | P + D | 2 |
| C | Sclerosing/morphoeic BCC | undifferentiated | D + P | 1 |
| BAS81 | Infiltrating BCC | undifferentiated | D + P | 1 |
| D | Sclerosing/morphoeic BCC | undifferentiated | D | 1 |
| F | Sclerosing/morphoeic BCC | undifferentiated | D | 1 |
| H | Sclerosing/morphoeic BCC | undifferentiated | D | 1 |
| O | Infiltrating BCC | undifferentiated | P + D | 3 |
| P | Infiltrating BCC | undifferentiated | P + D | 3 |
| Q | Infiltrating BCC | undifferentiated | P | 2 |
| T | Infiltrating BCC | undifferentiated | P | 3 |
| BAS89 | Infiltrating BCC | undifferentiated | P | 1 |
| Primer Name | Sequence 5′-3′ |
|---|---|
| Gapdh-F: | TGCACCACCAACTGCTTAGC |
| Gapdh-R: | TCTTCTGGGTGGCAGTGATG |
| Tbx1_qRT-PCR-F: | CTGACCAATAACCTGCTGGATGA |
| Tbx1_qRT-PCR-R: | GGCTGATATCTGTGCATGGAGTT |
| Tbx1_RT-PCR-F: | TTTGTGCCCGTAGATGACAA |
| Tbx1_RT-PCR-R: | AATCGGGGCTGATATCTGTG |
| Gli2-F: | ATCAAGACAGAGAGCTCCGG |
| Gli2-R: | ATGCCACTGTCATTGTTGGG |
| Gli2-5′-F: | GCCGATTGACATGAGACACC |
| Gli2-5′-R: | CTGAAGGGTGACTCTCCAGG |
| Sox9-F: | GTACCCGCATCTGCACAAC |
| Sox9-R: | CTCCTCCACGAAGGGTCTCT |
| Wnt7a-F: | CGCTCATGAACTTACACAATAACG |
| Wnt7a-R: | ACAGGAGCCTGACACACCAT |
| Fzd3-F: | AGGTGGGCACAGTTTGTTTT |
| Fzd3-R: | GAAATGGCCGAAAATCCCGA |
| Ptch2-F: | GTCCACCTAGTGCTCCCAAC |
| Ptch2-R: | GTGCCCCCTAGTAGCAGTTC |
| Dvl2-F: | AGCAGTGCCTCCCGCCTCCTCA |
| Dvl2-R: | CCCATCACCACGCTCGTTACTTTG |
| Sufu-F: | TACTACGGACAGTGCCCATT |
| Sufu-R: | GTCTGTCTCAATGCCTTTGTCA |
| Gli1-F: | GAATTCGTGTGCCATTG |
| Gli1-R: | GGACTTCCGACAGCCTTCAA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caprio, C.; Varricchio, S.; Bilio, M.; Feo, F.; Ferrentino, R.; Russo, D.; Staibano, S.; Alfano, D.; Missero, C.; Ilardi, G.; et al. TBX1 and Basal Cell Carcinoma: Expression and Interactions with Gli2 and Dvl2 Signaling. Int. J. Mol. Sci. 2020, 21, 607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020607
Caprio C, Varricchio S, Bilio M, Feo F, Ferrentino R, Russo D, Staibano S, Alfano D, Missero C, Ilardi G, et al. TBX1 and Basal Cell Carcinoma: Expression and Interactions with Gli2 and Dvl2 Signaling. International Journal of Molecular Sciences. 2020; 21(2):607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020607
Chicago/Turabian StyleCaprio, Cinzia, Silvia Varricchio, Marchesa Bilio, Federica Feo, Rosa Ferrentino, Daniela Russo, Stefania Staibano, Daniela Alfano, Caterina Missero, Gennaro Ilardi, and et al. 2020. "TBX1 and Basal Cell Carcinoma: Expression and Interactions with Gli2 and Dvl2 Signaling" International Journal of Molecular Sciences 21, no. 2: 607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020607