Multifaceted Analyses of Epidermal Serine Protease Activity in Patients with Atopic Dermatitis

 and
and 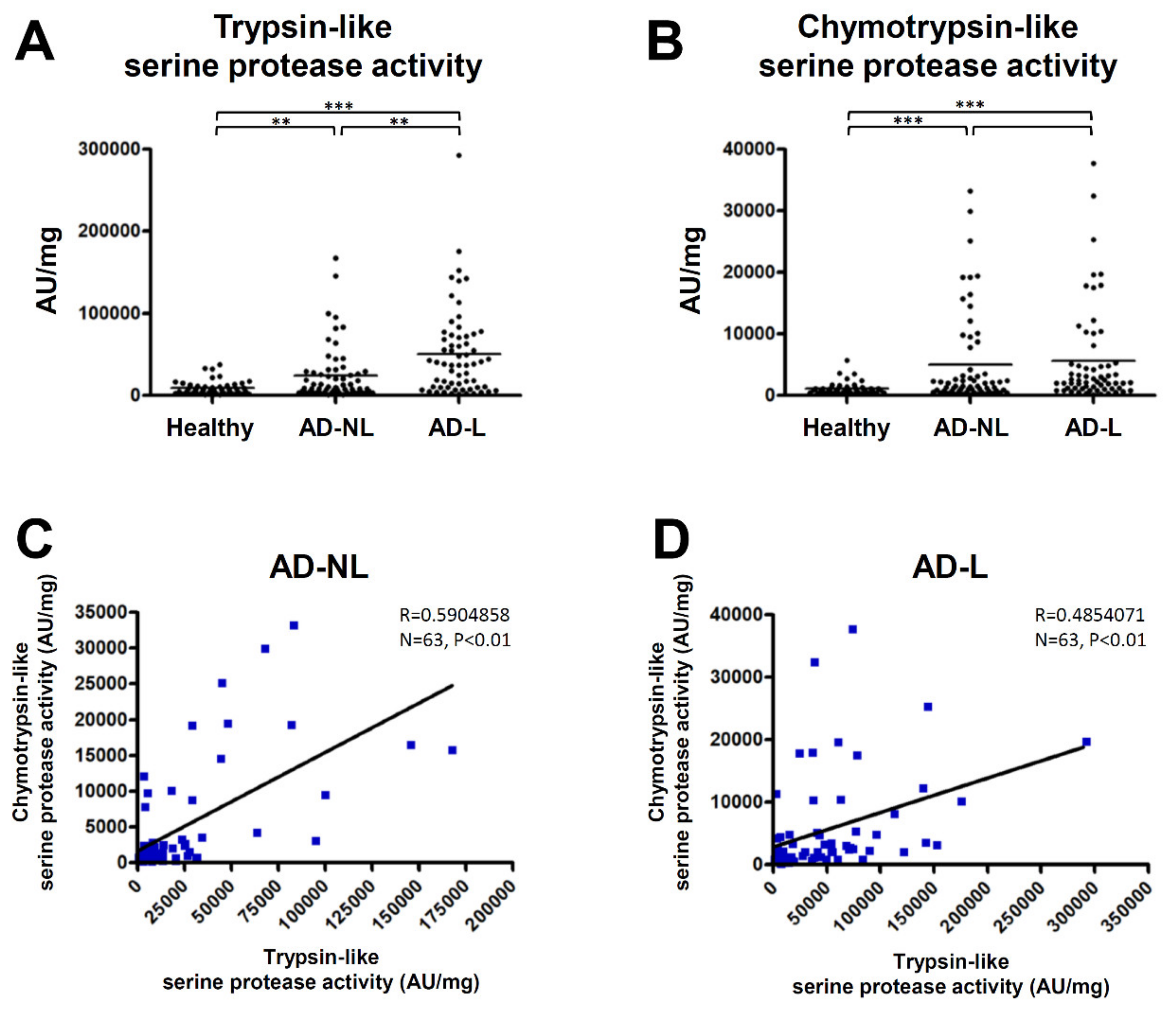{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Serine Protease Activity in the SC of AD Is Increased in Both Non-Lesions and Lesions
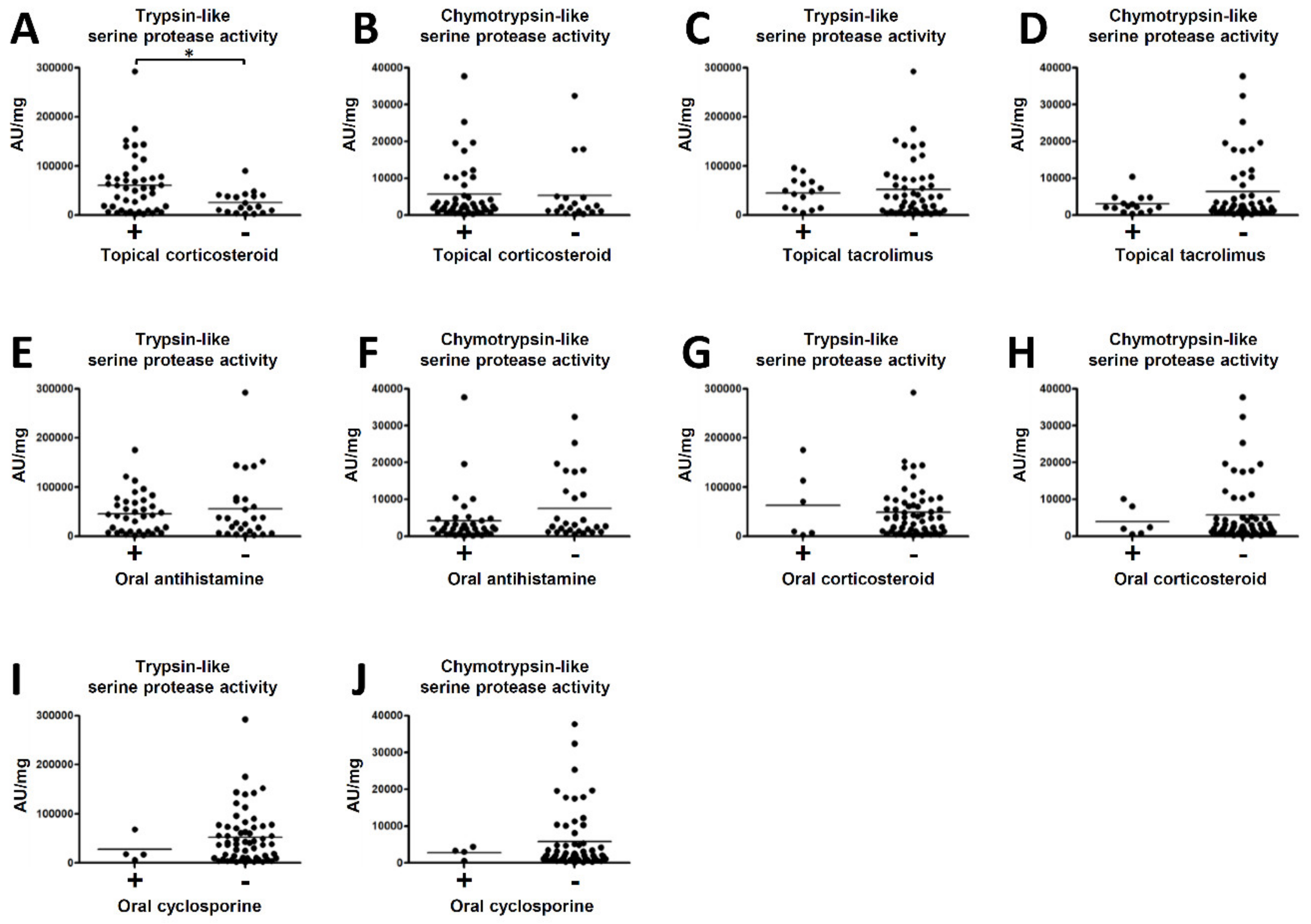
2.2. The Association between Serine Protease Activity in the SC of AD and Medical Treatments
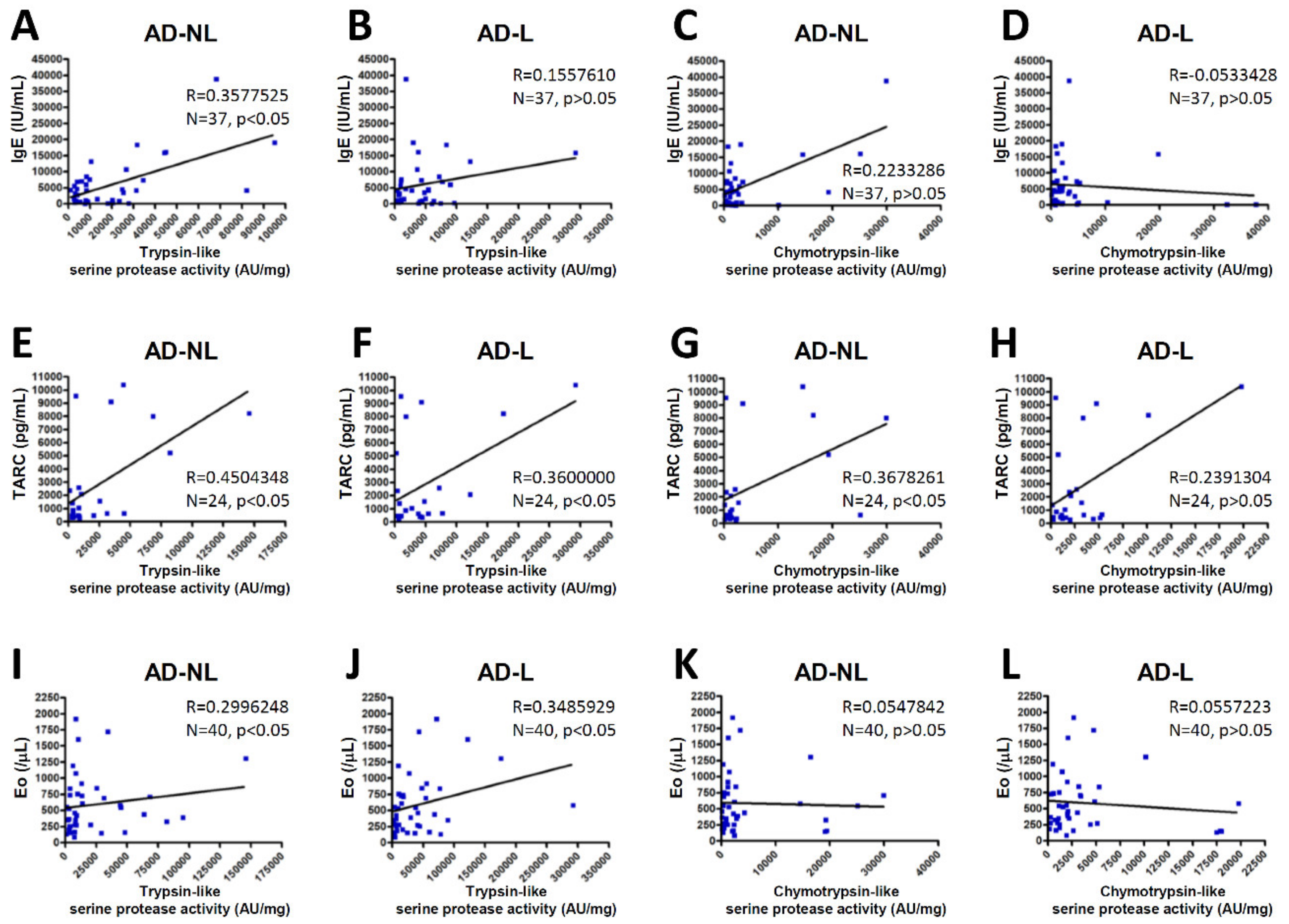
2.3. Serine Protease Activity in the SC of AD Correlate with Biomarkers of AD
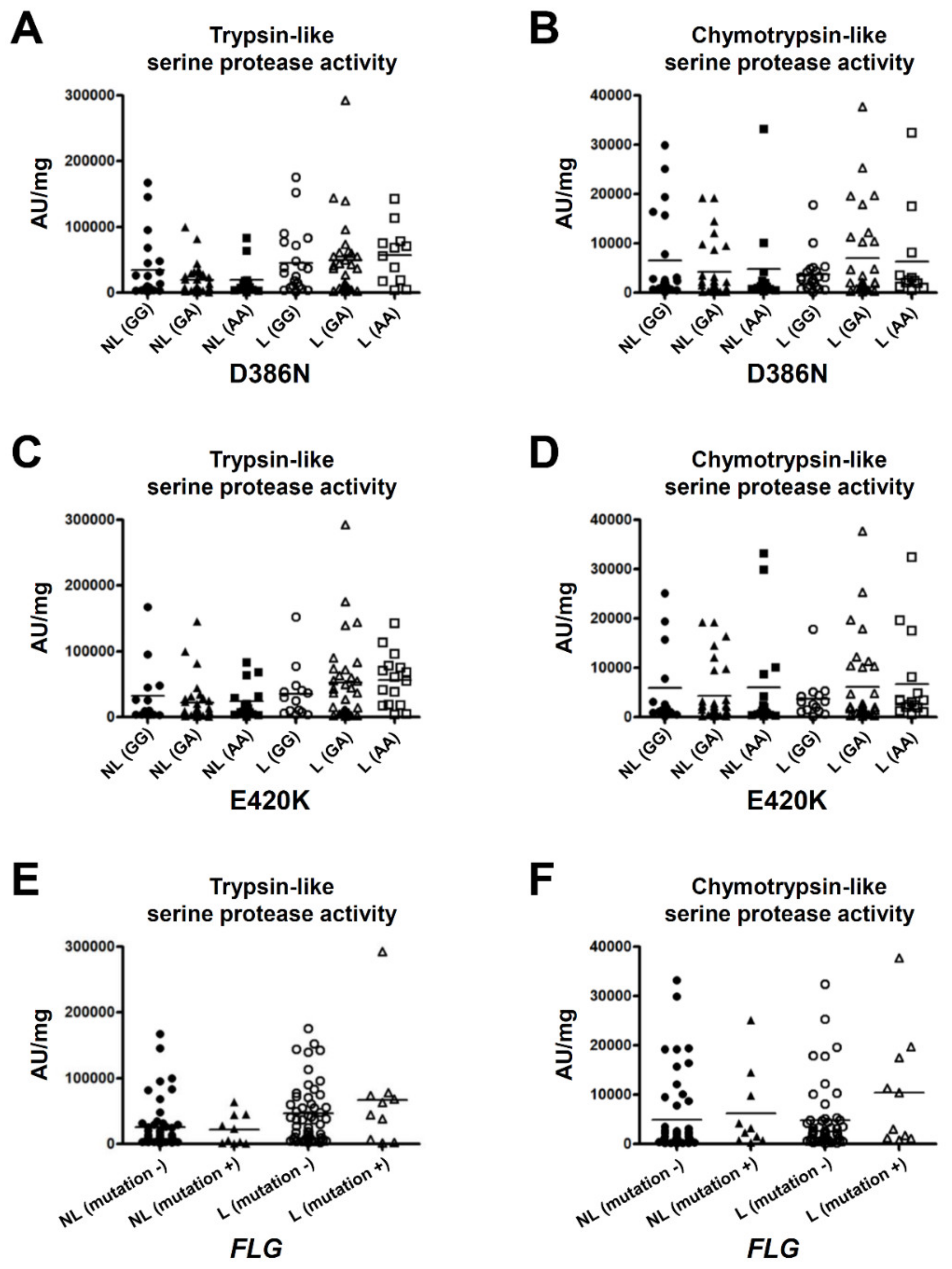
2.4. p.D386N and p.E420K of SPINK5 and Loss-Of-Function Mutations in FLG Do Not Affect the Serine Protease Activity in the SC
3. Discussion
4. Subjects and Methods
4.1. Human Samples
4.2. Collection of the Stratum Corneum
4.3. Protease Assays
4.4. Blood Examinations and Medical Treatment Histories
4.5. Analysis of the SPINK5 Gene and the FLG Gene
4.6. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| D | Domain |
| FLG | Filaggrin |
| IgE | Immunoglobulin E |
| IL | Interleukin |
| KLK | Kallikrein-related peptidase |
| LEKTI | Lympho-epithelial Kazal-type-related inhibitor |
| SC | Stratum corneum |
| SLPI | Secretory leukocyte protease inhibitor |
| SPINK5 | Serine protease inhibitor Kazal type 5 |
| TARC | Thymus and activation-regulated chemokine |
| Th | T-helper lymphocytes |
References
- Eichenfield, L.F.; Tom, W.L.; Chamlin, S.L.; Feldman, S.R.; Hanifin, J.M.; Simpson, E.L.; Berger, T.G.; Bergman, J.N.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 1. Diagnosis and assessment of atopic dermatitis. J. Am. Acad. Dermatol. 2014, 70, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Chavanas, S.; Bodemer, C.; Rochat, A.; Hamel-Teillac, D.; Ali, M.; Irvine, A.D.; Bonafe, J.L.; Wilkinson, J.; Taieb, A.; Barrandon, Y.; et al. Mutations in SPINK5, encoding a serine protease inhibitor, cause Netherton syndrome. Nat. Genet. 2000, 25, 141–142. [Google Scholar] [CrossRef]
- Descargues, P.; Deraison, C.; Bonnart, C.; Kreft, M.; Kishibe, M.; Ishida-Yamamoto, A.; Elias, P.; Barrandon, Y.; Zambruno, G.; Sonnenberg, A.; et al. Spink5-Deficient mice mimic Netherton syndrome through degradation of desmoglein 1 by epidermal protease hyperactivity. Nat. Genet. 2005, 37, 56–65. [Google Scholar] [CrossRef]
- Komatsu, N.; Saijoh, K.; Jayakumar, A.; Clayman, G.L.; Tohyama, M.; Suga, Y.; Mizuno, Y.; Tsukamoto, K.; Taniuchi, K.; Takehara, K.; et al. Correlation between SPINK5 gene mutations and clinical manifestations in Netherton syndrome patients. J. Investig. Dermatol. 2008, 128, 1148–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, N.; Saijoh, K.; Kuk, C.; Liu, A.C.; Khan, S.; Shirasaki, F.; Takehara, K.; Diamandis, E.P. Human tissue kallikrein expression in the stratum corneum and serum of atopic dermatitis patients. Exp. Dermatol. 2007, 16, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, N.; Saijoh, K.; Kuk, C.; Shirasaki, F.; Takehara, K.; Diamandis, E.P. Aberrant human tissue kallikrein levels in the stratum corneum and serum of patients with psoriasis: Dependence on phenotype, severity and therapy. Br. J. Dermatol. 2007, 156, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2007, 13, 975–980. [Google Scholar] [CrossRef]
- Voegeli, R.; Rawlings, A.V.; Breternitz, M.; Doppler, S.; Schreier, T.; Fluhr, J.W. Increased stratum corneum serine protease activity in acute eczematous atopic skin. Br. J. Dermatol. 2009, 161, 70–77. [Google Scholar] [CrossRef]
- Sotiropoulou, G.; Pampalakis, G.; Diamandis, E.P. Functional roles of human kallikrein-related peptidases. J. Biol. Chem. 2009, 284, 32989–32994. [Google Scholar] [CrossRef] [Green Version]
- Caubet, C.; Jonca, N.; Brattsand, M.; Guerrin, M.; Bernard, D.; Schmidt, R.; Egelrud, T.; Simon, M.; Serre, G. Degradation of corneodesmosome proteins by two serine proteases of the kallikrein family, SCTE/KLK5/hK5 and SCCE/KLK7/hK7. J. Investig. Dermatol. 2004, 122, 1235–1244. [Google Scholar] [CrossRef] [Green Version]
- Morizane, S. The role of kallikrein-related peptidases in atopic dermatitis. Acta Med. Okayama 2019, 73, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Briot, A.; Deraison, C.; Lacroix, M.; Bonnart, C.; Robin, A.; Besson, C.; Dubus, P.; Hovnanian, A. Kallikrein 5 induces atopic dermatitis-like lesions through PAR2-mediated thymic stromal lymphopoietin expression in Netherton syndrome. J. Exp. Med. 2009, 206, 1135–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furio, L.; de Veer, S.; Jaillet, M.; Briot, A.; Robin, A.; Deraison, C.; Hovnanian, A. Transgenic kallikrein 5 mice reproduce major cutaneous and systemic hallmarks of Netherton syndrome. J. Exp. Med. 2014, 211, 499–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, L.; Backman, A.; Ny, A.; Edlund, M.; Ekholm, E.; Ekstrand-Hammarstrom, B.; Tornell, J.; Wallbrandt, P.; Wennbo, H.; Egelrud, T. Epidermal overexpression of stratum corneum chymotryptic enzyme in mice: A model for chronic itchy dermatitis. J. Investig. Dermatol. 2002, 118, 444–449. [Google Scholar] [CrossRef] [Green Version]
- Morizane, S.; Yamasaki, K.; Kajita, A.; Ikeda, K.; Zhan, M.; Aoyama, Y.; Gallo, R.L.; Iwatsuki, K. TH2 cytokines increase kallikrein 7 expression and function in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 259–261. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Hoffert, U. Reddish, scaly, and itchy: How proteases and their inhibitors contribute to inflammatory skin diseases. Arch. Immunol. Ther. Exp. 2009, 57, 345–354. [Google Scholar] [CrossRef]
- Deraison, C.; Bonnart, C.; Lopez, F.; Besson, C.; Robinson, R.; Jayakumar, A.; Wagberg, F.; Brattsand, M.; Hachem, J.P.; Leonardsson, G.; et al. LEKTI fragments specifically inhibit KLK5, KLK7, and KLK14 and control desquamation through a pH-dependent interaction. Mol. Biol. Cell 2007, 18, 3607–3619. [Google Scholar] [CrossRef] [Green Version]
- Magert, H.J.; Standker, L.; Kreutzmann, P.; Zucht, H.D.; Reinecke, M.; Sommerhoff, C.P.; Fritz, H.; Forssmann, W.G. LEKTI, a novel 15-domain type of human serine proteinase inhibitor. J. Biol. Chem. 1999, 274, 21499–21502. [Google Scholar] [CrossRef] [Green Version]
- Hovnanian, A. Netherton syndrome: Skin inflammation and allergy by loss of protease inhibition. Cell Tissue Res. 2013, 351, 289–300. [Google Scholar] [CrossRef]
- Fortugno, P.; Furio, L.; Teson, M.; Berretti, M.; El Hachem, M.; Zambruno, G.; Hovnanian, A.; D’Alessio, M. The 420K LEKTI variant alters LEKTI proteolytic activation and results in protease deregulation: Implications for atopic dermatitis. Hum. Mol. Genet. 2012, 21, 4187–4200. [Google Scholar] [CrossRef] [Green Version]
- Kato, A.; Fukai, K.; Oiso, N.; Hosomi, N.; Murakami, T.; Ishii, M. Association of SPINK5 gene polymorphisms with atopic dermatitis in the Japanese population. Br. J. Dermatol. 2003, 148, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Kusunoki, T.; Okafuji, I.; Yoshioka, T.; Saito, M.; Nishikomori, R.; Heike, T.; Sugai, M.; Shimizu, A.; Nakahata, T. SPINK5 polymorphism is associated with disease severity and food allergy in children with atopic dermatitis. J. Allergy Clin. Immunol. 2005, 115, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Nishio, Y.; Noguchi, E.; Shibasaki, M.; Kamioka, M.; Ichikawa, E.; Ichikawa, K.; Umebayashi, Y.; Otsuka, F.; Arinami, T. Association between polymorphisms in the SPINK5 gene and atopic dermatitis in the Japanese. Genes Immun. 2003, 4, 515–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walley, A.J.; Chavanas, S.; Moffatt, M.F.; Esnouf, R.M.; Ubhi, B.; Lawrence, R.; Wong, K.; Abecasis, G.R.; Jones, E.Y.; Harper, J.I.; et al. Gene polymorphism in Netherton and common atopic disease. Nat. Genet. 2001, 29, 175–178. [Google Scholar] [CrossRef]
- Ramesh, K.; Matta, S.A.; Chew, F.T.; Mok, Y.K. Exonic mutations associated with atopic dermatitis disrupt lympho-epithelial Kazal-type related inhibitor action and enhance its degradation. Allergy 2019. [Google Scholar] [CrossRef]
- Brown, S.J.; McLean, W.H. One remarkable molecule: Filaggrin. J. Investig. Dermatol. 2012, 132, 751–762. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.J.; Irvine, A.D.; Terron-Kwiatkowski, A.; Sandilands, A.; Campbell, L.E.; Zhao, Y.; Liao, H.; Evans, A.T.; Goudie, D.R.; Lewis-Jones, S.; et al. Loss-of-function mutations in the gene encoding filaggrin cause ichthyosis vulgaris. Nat. Genet. 2006, 38, 337–342. [Google Scholar] [CrossRef]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Nemoto-Hasebe, I.; Akiyama, M.; Nomura, T.; Sandilands, A.; McLean, W.H.; Shimizu, H. FLG mutation p.Lys4021X in the C-terminal imperfect filaggrin repeat in Japanese patients with atopic eczema. Br. J. Dermatol. 2009, 161, 1387–1390. [Google Scholar] [CrossRef]
- Kono, M.; Nomura, T.; Ohguchi, Y.; Mizuno, O.; Suzuki, S.; Tsujiuchi, H.; Hamajima, N.; McLean, W.H.; Shimizu, H.; Akiyama, M. Comprehensive screening for a complete set of Japanese-population-specific filaggrin gene mutations. Allergy 2014, 69, 537–540. [Google Scholar] [CrossRef]
- Bieber, T.; D’Erme, A.M.; Akdis, C.A.; Traidl-Hoffmann, C.; Lauener, R.; Schappi, G.; Schmid-Grendelmeier, P. Clinical phenotypes and endophenotypes of atopic dermatitis: Where are we, and where should we go? J. Allergy Clin. Immunol. 2017, 139, S58–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuperstock, J.E.; Brook, C.D.; Ryan, M.W.; Platt, M.P. Correlation between the number of allergen sensitizations and immunoglobulin E: Monosensitization vs. polysensitization. Int. Forum Allergy Rhinol. 2017, 7, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y. Thymus and activation-regulated chemokine as a clinical biomarker in atopic dermatitis. J. Dermatol. 2014, 41, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Braathen, L.R.; Simon, H.U. Eosinophils and atopic dermatitis. Allergy 2004, 59, 561–570. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nomura, H.; Suganuma, M.; Takeichi, T.; Kono, M.; Isokane, Y.; Sunagawa, K.; Kobashi, M.; Sugihara, S.; Kajita, A.; Miyake, T.; et al. Multifaceted Analyses of Epidermal Serine Protease Activity in Patients with Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030913
Nomura H, Suganuma M, Takeichi T, Kono M, Isokane Y, Sunagawa K, Kobashi M, Sugihara S, Kajita A, Miyake T, et al. Multifaceted Analyses of Epidermal Serine Protease Activity in Patients with Atopic Dermatitis. International Journal of Molecular Sciences. 2020; 21(3):913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030913
Chicago/Turabian StyleNomura, Hayato, Mutsumi Suganuma, Takuya Takeichi, Michihiro Kono, Yuki Isokane, Ko Sunagawa, Mina Kobashi, Satoru Sugihara, Ai Kajita, Tomoko Miyake, and et al. 2020. "Multifaceted Analyses of Epidermal Serine Protease Activity in Patients with Atopic Dermatitis" International Journal of Molecular Sciences 21, no. 3: 913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030913