Molecular Characterization and Disease Control of Stem Canker on Royal Poinciana (Delonix regia) Caused by Neoscytalidium dimidiatum in the United Arab Emirates

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Symptoms of Stem Canker Disease on Royal Poinciana
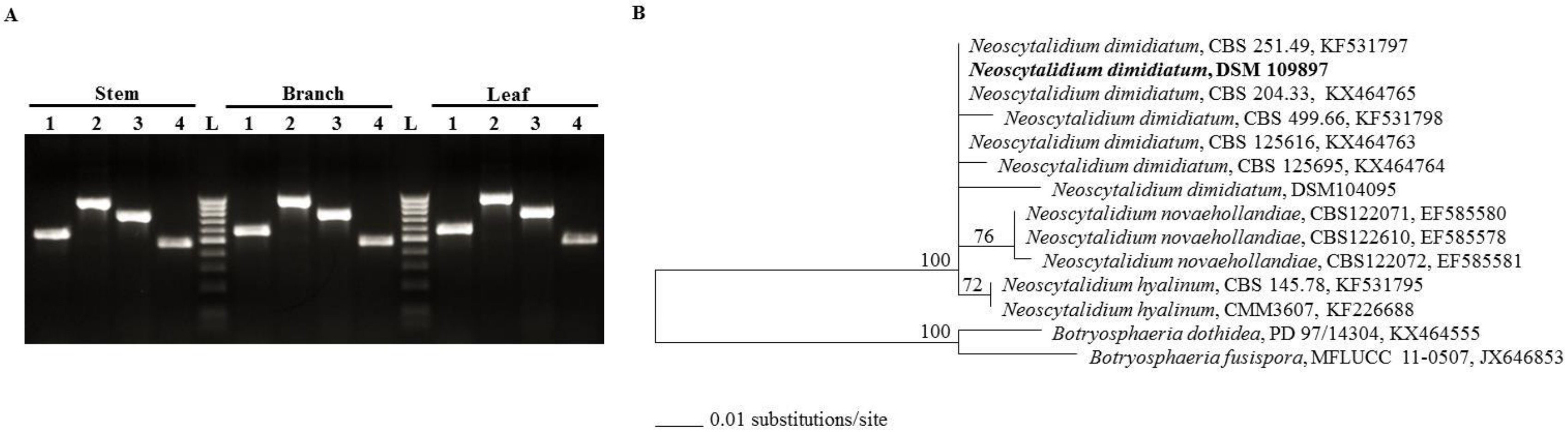
2.2. Identification and Molecular Characterization of Neoscytalidium Dimidiatum
2.3. Pathogenicity Tests of Neoscytalidium Dimidiatum on Royal Poinciana Seedlings and Apple Fruits
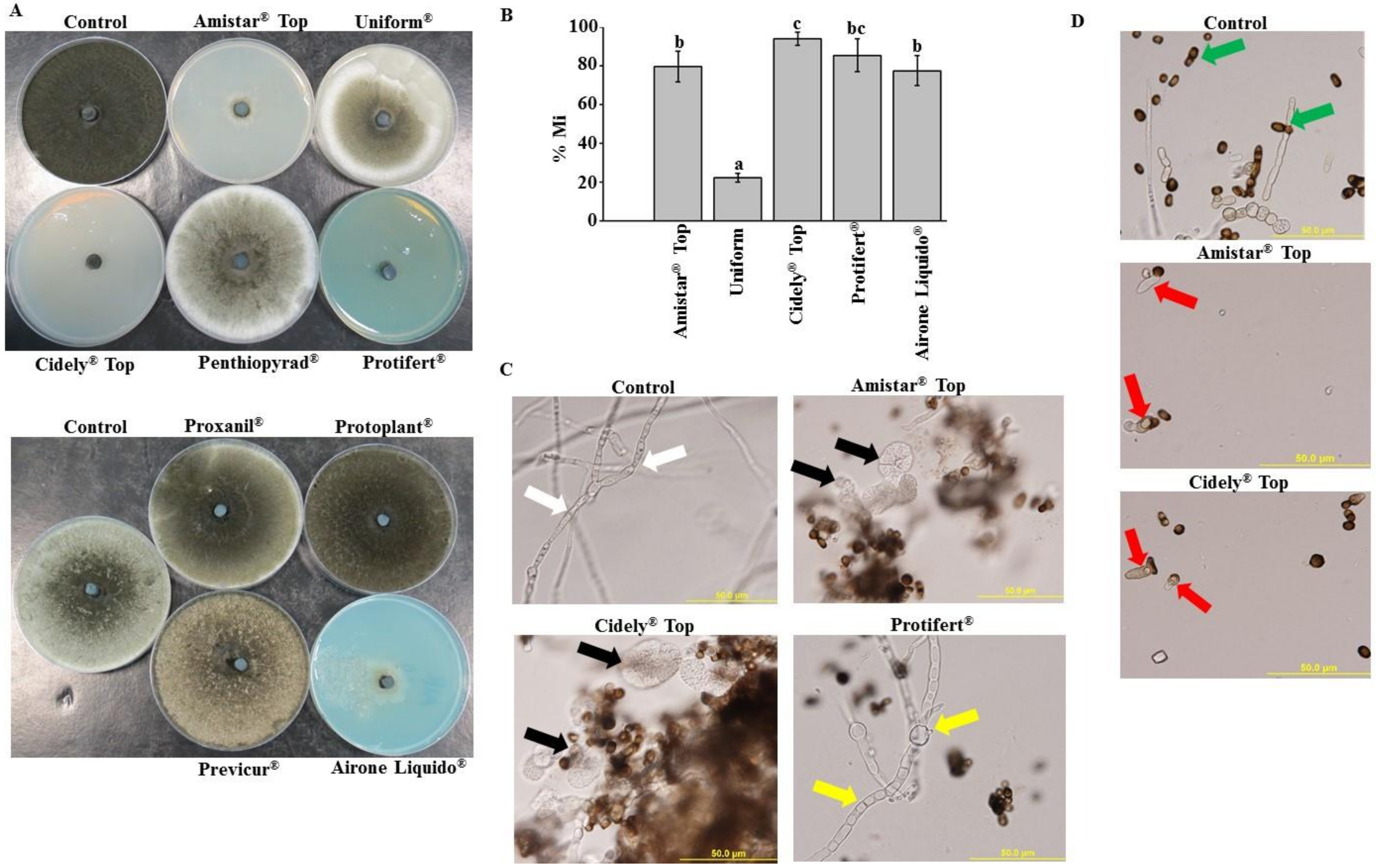
2.4. In Vitro Evaluation of Chemical Fungicides to Neoscytalidium Dimidiatum
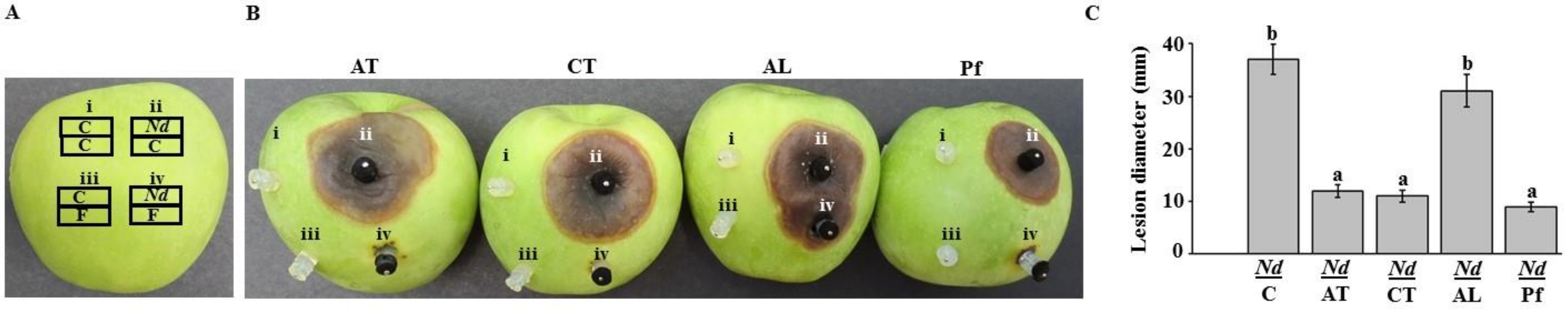
2.5. Assessment of Chemical Fungicides on Neoscytalidium Dimidiatum Using Apple Bioassay
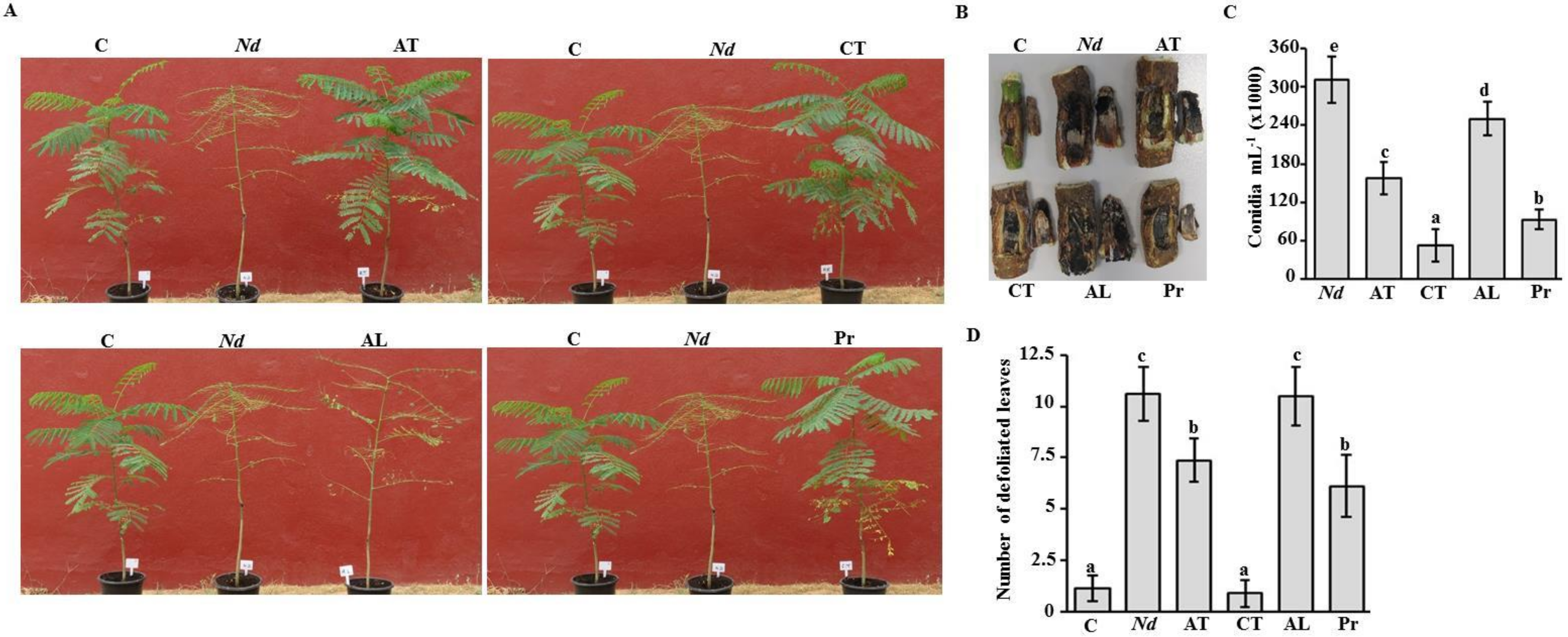
2.6. Fungicide Effects on Royal Poinciana Infected with Neoscytalidium Dimidiatum
2.7. Effect of Cidely® Top on Royal Poinciana Trees Naturally Infected with Neoscytalidium Dimidiatum
3. Discussion
4. Materials and Methods
4.1. Fungal Culture and Isolation
4.2. Molecular Identification of the Pathogen
4.3. In Vivo Pathogenicity Tests and Koch’s Postulates
4.4. In Vitro Evaluation of Fungicides Against N. Dimidiatum
4.5. In Vivo Evaluation of Selected Fungicides
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gledhill, D. The Names of Plants, 4th ed.; Cambridge University Press: Cambridge, UK, 2008; p. 137. [Google Scholar]
- Kirtikar, K.R.; Basu, B.D. Indian medicinal plants, 2nd ed.; International Book Distributors: Dehradun, India, 1999; Volume 2, p. 852. [Google Scholar]
- Gilman, E.F.; Watson, D.G.; Klein, R.W.; Koeser, A.K.; Hilbert, D.R.; McLean, D.C. Delonix regia: Royal Poinciana; The Institute of Food and Agricultural Sciences (IFAS), University of Florida: Gainesville, FL, USA, 2019; Available online: edis.ifas.ufl.edu/pdffiles/ST/ST22800.pdf (accessed on 3 January 2020).
- Cayley, D.M. Fungi associated with “die back” in stone fruit trees. Ann. Appl. Biol. 1923, 10, 253–275. [Google Scholar] [CrossRef]
- Anagnostakis, S.L.; Ferrandino, F.J. Isolation of Nectria galligena from cankers on sweet birch. Plant Dis. 1998, 82, 440–441. [Google Scholar] [CrossRef]
- Rouxel, T.; Balesdent, M.H. The stem canker (blackleg) fungus, Leptosphaeria maculans, enters the genomic era. Mol. Plant Pathol. 2005, 6, 225–241. [Google Scholar] [CrossRef]
- Chungu, D.; Muimba-Kankolongo, A.; Wingfield, M.J.; Roux, J. Identification of fungal pathogens occurring in eucalypt and pine plantations in Zambia by comparing DNA sequences. Forestry 2010, 83, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Darge, W.A. First report of Lasioplodia theobromae causing needle blight and stem canker diseases on Araucaria heterophylla in Ethiopia. J. Hortic. Res. 2017, 25, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Reckhaus, P. Hendersonula dieback of mango in Niger. Plant Dis. 1987, 71, 1045. [Google Scholar] [CrossRef]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plants and Plant Products in the United States; The American Phytopathological Society (APS) Press: St. Paul, MN, USA, 1989; Volume 8, p. 1252. [Google Scholar]
- Ray, J.D.; Burgess, T.; Lanoiselet, V.M. First record of Neoscytalidium dimidiatum and N. novaehollandiae on Mangifera indica and N. dimidiatum on Ficus carica in Australia. Australas. Plant Dis. Notes 2010, 5, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.H.; Lin, Q.L.; Mo, J.J.; Wu, F.F.; Chen, J. Fruit internal brown rot caused by Neoscytalidium dimidiatum on pitahaya in Guangdong province, China. Australas. Plant Dis. Notes 2015, 10, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Al-Bedak, O.A.; Mohamed, R.A.; Seddek, N.H. First detection of Neoscytalidium dimidiatum associated with canker disease in Egyptian Ficus trees. Forest Pathol. 2017, 1–7. [Google Scholar] [CrossRef]
- Hajlaoui, M.R.; Nouri, M.T.; Hamrouni, N.; Trouillas, F.P.; Ben Yahmed, N.; Eddouzi, J.; Mnari-Hattab, M. First record ofdieback and decline of plum caused by Neoscytalidium dimidiatum in Tunisia. New Dis. Rep. 2018, 38, 20. [Google Scholar] [CrossRef] [Green Version]
- Nouri, M.T.; Lawrence, D.P.; Yaghmour, M.A.; Michailides, T.J.; Trouillas, F.P. Neoscytalidium dimidiatum causing canker, shoot blight and fruit rot of almond in California. Plant Dis. 2018, 102, 1638–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshafie, A.E.; Ba-Omar, T. First report of Albizia lebbeck caused by Scytalidium dimidiatum in Oman. Mycopathologia 2002, 154, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Saeed, E.E.; Sham, A.; El-Tarabily, K.A.; Abu Elsamen, F.; Iratni, R.; AbuQamar, S.F. Chemical control of dieback disease on date palm caused by the fungal pathogen, Thielaviopsis punctulata, in United Arab Emirates. Plant Dis. 2016, 100, 2370–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, E.E.; Sham, A.; AbuZarqa, A.; Al Shurafa, K.; Al Naqbi, T.S.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Detection and management of mango dieback disease in the United Arab Emirates. Int. J. Mol. Sci. 2017, 18, 2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alwahshi, K.J.; Saeed, E.E.; Sham, A.; Alblooshi, A.A.; Alblooshi, M.M.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular identification and disease management of date palm sudden decline syndrome in the United Arab Emirates. Int. J. Mol. Sci. 2019, 20, 923. [Google Scholar] [CrossRef] [Green Version]
- Chuang, M.F.; Ni, H.F.; Yang, H.R.; Shu, S.L.; Lai, S.Y.; Jiang, Y.L. First report of stem canker disease of pitaya (Hylocereus undatus and H. polyrhizus) caused by Neoscytalidium dimidiatum in Taiwan. Plant Dis. 2012, 96, 906–907. [Google Scholar] [CrossRef]
- Mohd, M.H.; Salleh, B.; Zakaria, L. Identification and molecular characterizations of Neoscytalidium dimidiatum causing stem canker of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 2013, 161, 841–849. [Google Scholar] [CrossRef]
- Kamil, F.H.; Saeed, E.E.; El-Tarabily, K.A.; AbuQamar, S.F. Biological control of mango dieback disease caused by Lasiodiplodia theobromae using streptomycete and non-streptomycete actinobacteria in the United Arab Emirates. Front Microbiol. 2018, 9, 829. [Google Scholar] [CrossRef]
- Al-Tememe, Z.A.M.; Lahuf, A.; Abdalmoohsin, R.G.; Al-Amirry, A.T. Occurrence, identification, pathogenicity and control of Neoscytalidium dimidiatum fungus, the causal agent of sooty canker on Eucalyptus camaldulensis in Kerbala Province of Iraq. 2019. Plant Arch. 2019, 19, 31–38. [Google Scholar]
- Murad, N.Y.; Al-Dabagh, M.N. Evaluation some of pesticides in control of Neoscytalidium dimidiatum (Penz) Crous and Slippers causing wilt and canker on cypress trees in Iraq. Iraqi J. Agric. Sci. 2014, 19, 25–38. [Google Scholar]
- Farr, D.F.; Elliott, M.; Rossman, A.Y.; Edmonds, R.L. Fusicoccum arbuti sp. nov. causing cankers on pacific madrone in western North America with notes on Fusicoccum dimidiatum, the correct name for Scytalidium dimidiatum and Nattrassia mangiferae. Mycologia 2005, 97, 730–741. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 91, 315–322. [Google Scholar]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.D.; Ngoc, D.T.B.; Thang, N.D.; Tuan, L.N.A.; Thach, B.D.; Hien, N.Q. Synthesis and in vitro antifungal efficiency of alginate-stabilized Cu2O-Cu nanoparticles against Neoscytalidium dimidiatum causing brown spot disease on dragon fruit plants (Hylocereus undatus). Vietnam J. Chem. 2019, 57, 318–323. [Google Scholar]
- Hassan, W.A.; Pasha, A.A.; Mohammad, M.B. Sooty canker on some thin bark trees caused by Nattrassia mangiferae. Egypt. J. Agric. Res. 2009, 87, 443–456. [Google Scholar]
- Hassan, W.A.; Haleem, R.A.; Hassan, P.H. Effect of heat-stress predisposition on the development of sooty canker caused by Neoscytalidium dimidiatum (Penz.) Crous & Slippers. Acta Agrobot. 2011, 64, 207–212. [Google Scholar]
- Alhammadi, M.S.; Al-Shariqi, R.; Maharachchikumbura, S.; Al-Sadi, A.M. Molecular identification of fungal pathogens associated with date palm root diseases in the United Arab Emirates. J. Plant Pathol. 2018, 99, 1–7. [Google Scholar]
- Pavlic, D.; Wingfield, M.J.; Barber, P.; Slippers, B.; Hardy, G.E.S.; Burgess, T.I. Seven new species of the Botryosphaeriaceae from baobab and other native trees in Western Australia. Mycologia 2008, 100, 851–866. [Google Scholar] [CrossRef] [Green Version]
- Polizzi, G.; Aiello, D.; Vitale, A.; Giuffrida, F.; Groenewald, Z.; Crous, P.W. (2009) First report of shoot blight, canker, and gumumosis caused by Neoscytalidium dimidiatum on citrus in Italy. Plant Dis. 2009, 93, 1215. [Google Scholar] [CrossRef]
- Madrid, H.; Ruı´z-Cendoya, M.; Cano, J.; Stchigel, A.; Orofino, R.; Guarro, J. Genotyping and in vitro antifungal susceptibility of Neoscytalidium dimidiatum isolates from different origins. Int. J. Antimicrob. Agents 2009, 34, 351–354. [Google Scholar] [CrossRef]
- Alwan, S.L.; Hussein, H.N. Efficacy of ecofriendly biocontrol Azotobacter chroococcum and Lactobacillus rhamnosus for enhancing plant growth and reducing infection by Neoscytalidium spp. in fig (Ficus carica L.) saplings. J. Kerbala Agric. Sci. 2019, 6, 16–25. [Google Scholar]
- Hortova, B.; Novotny, D.; Erban, T. Physiological characteristics and pathogenicity of eight Neofabraea isolates from apples in Czechia. Europ. J. Hort. Sci. 2014, 79, 327–334. [Google Scholar]
- AbuQamar, S.; Moustafa, K.; Tran, L.-S. Mechanisms and strategies of plant defense against Botrytis cinerea. Crit. Rev. Biotechnol. 2017, 37, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Mengiste, T.; Laluk, K.; AbuQamar, S. Mechanisms of induced resistance against B. cinerea. In Post-harvest Pathology; Chapter 2; Prusky, D., Gullino, M.L., Eds.; Springer Science + Business Media: Berlin, Germany, 2010; Volume 2, pp. 13–30. [Google Scholar]
- Rusmarini, W.; Shah, U.K.D.; Abdullah, M.P.; Mamat, S.; Hun, T.G. Identification of Trichoderma harzianum T3.13 and its interaction with Neoscytalidium dimidiatum U1, a pathogenic fungus islated from dragon fruit (Hylocereus polyrhizus) in Malaysia. Int. J. Environ. Agric. Res. 2017, 3, 3205–3228. [Google Scholar]
- Budzinski, H.; Couderchet, M. Environmental and human health issues related to pesticides: From usage and environmental fate to impact. Environ. Sci. Pollut. Res. 2018, 25, 14277. [Google Scholar] [CrossRef] [Green Version]
- Kuai, X.; Barraco, C.; Després, C. Combining fungicides and prospective NPR1-based “just-in-time” immunomodulating chemistries for crop protection. Front. Plant Sci. 2017, 8, 1715. [Google Scholar] [CrossRef] [Green Version]
- Saeed, E.E.; Sham, A.; Salmin, Z.; Abdelmowla, Y.; Iratni, R.; El-Tarabily, K.A.; AbuQamar, S.F. Streptomyces globosus UAE1, a potential effective biocontrol agent for black scorch disease in date palm plantations. Front. Microbiol. 2017, 8, 1455. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Hardy, G.E.St.J.; Sivasithamparam, K.; Hussein, A.M.; Kurtböke, D.I. The potential for the biological control of cavity spot disease of carrots caused by Pythium coloratum by streptomycete and non-streptomycete actinomycetes in Western Australia. New Phytol. 1997, 137, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Pervez, M.A.; Ahmad, S.; Iftikhar, Y.; Yasin, M.; Nawaz, A.; Ghazanfar, M.U.; Dasti, A.A.; Saleem, A. Determination of minimum inhibitory concentrations of fungicides against fungus Fusarium mangiferae. Pak. J. Bot. 2010, 42, 3525–3532. [Google Scholar]
- Khan, S.H.; Idrees, M.; Muhammad, F.; Mahmood, A.; Zaidi, S.H. Incidence of shisham (Dalbergia sissoo Roxb.) decline and in vitro response of isolated fungus spp. to various fungicides. Int. J. Agric. Biol. 2004, 6, 611–614. [Google Scholar]
- Yanase, Y.; Katsuta, H.; Tomiya, K.; Enomoto, M.; Sakamoto, O. Development of a novel fungicide, penthiopyrad. J. Pestic. Sci. 2013, 38, 167–168. [Google Scholar] [CrossRef] [Green Version]
- AbuQamar, S.F.; Moustafa, K.; Tran, L.S. ‘Omics’ and plant responses to Botrytis cinerea. Front. Plant Sci. 2016, 7, 1658. [Google Scholar] [CrossRef] [Green Version]
- Kirsop, B.E.; Doyle, A. Maintenance of microorganisms and cultured cells, a manual of laboratory methods, 2nd ed.; Academic Press: London, UK, 1991; p. 308. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplifed ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–555. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Raish, S.M.; Saeed, E.E.; Sham, A.; Alblooshi, K.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular Characterization and Disease Control of Stem Canker on Royal Poinciana (Delonix regia) Caused by Neoscytalidium dimidiatum in the United Arab Emirates. Int. J. Mol. Sci. 2020, 21, 1033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031033
Al Raish SM, Saeed EE, Sham A, Alblooshi K, El-Tarabily KA, AbuQamar SF. Molecular Characterization and Disease Control of Stem Canker on Royal Poinciana (Delonix regia) Caused by Neoscytalidium dimidiatum in the United Arab Emirates. International Journal of Molecular Sciences. 2020; 21(3):1033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031033
Chicago/Turabian StyleAl Raish, Seham M., Esam Eldin Saeed, Arjun Sham, Khulood Alblooshi, Khaled A. El-Tarabily, and Synan F. AbuQamar. 2020. "Molecular Characterization and Disease Control of Stem Canker on Royal Poinciana (Delonix regia) Caused by Neoscytalidium dimidiatum in the United Arab Emirates" International Journal of Molecular Sciences 21, no. 3: 1033. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031033