Interleukin-27 Gene Delivery Targeting IL-6Rα-Expressing Cells as a Stress Response Therapy

 ,
, 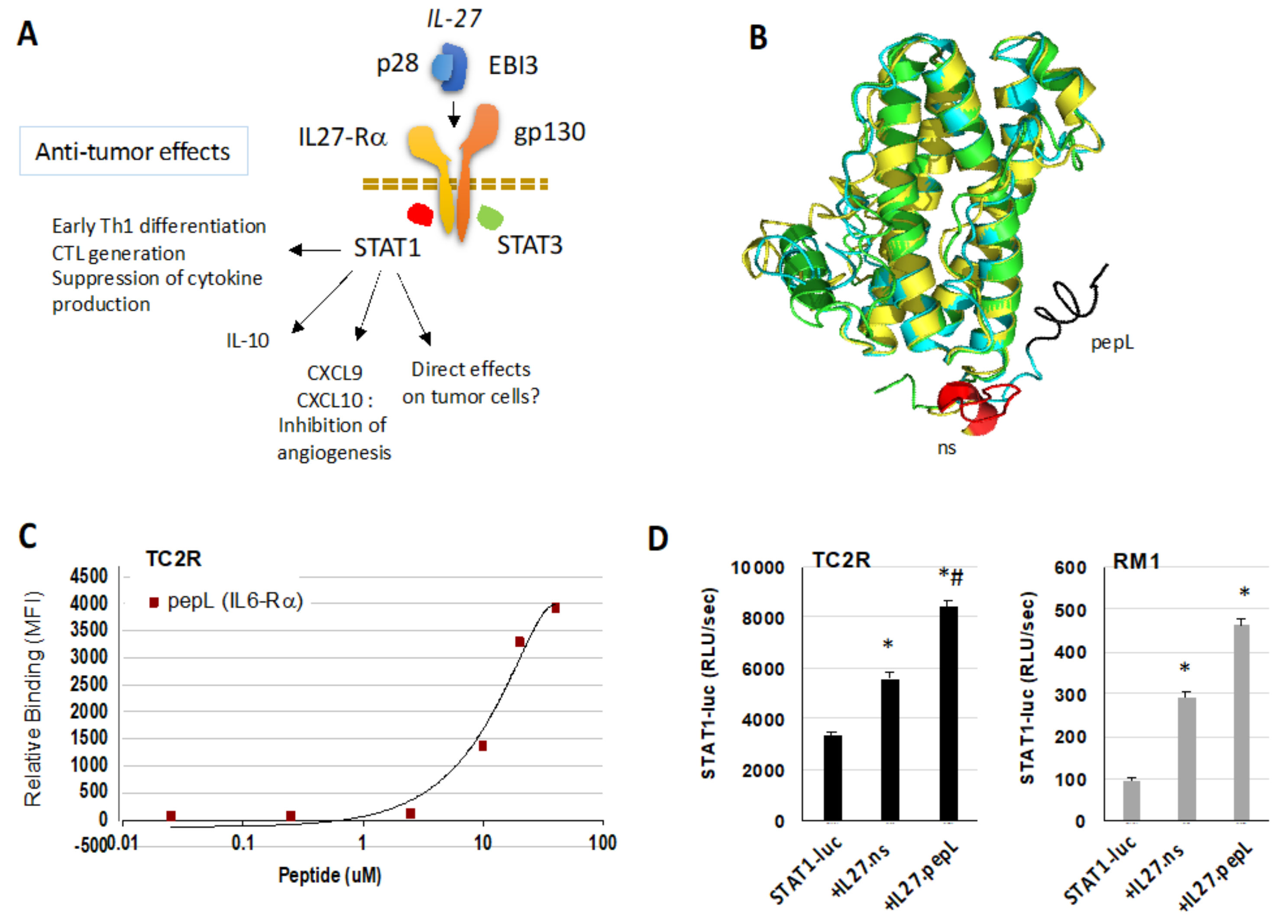{kind=link}
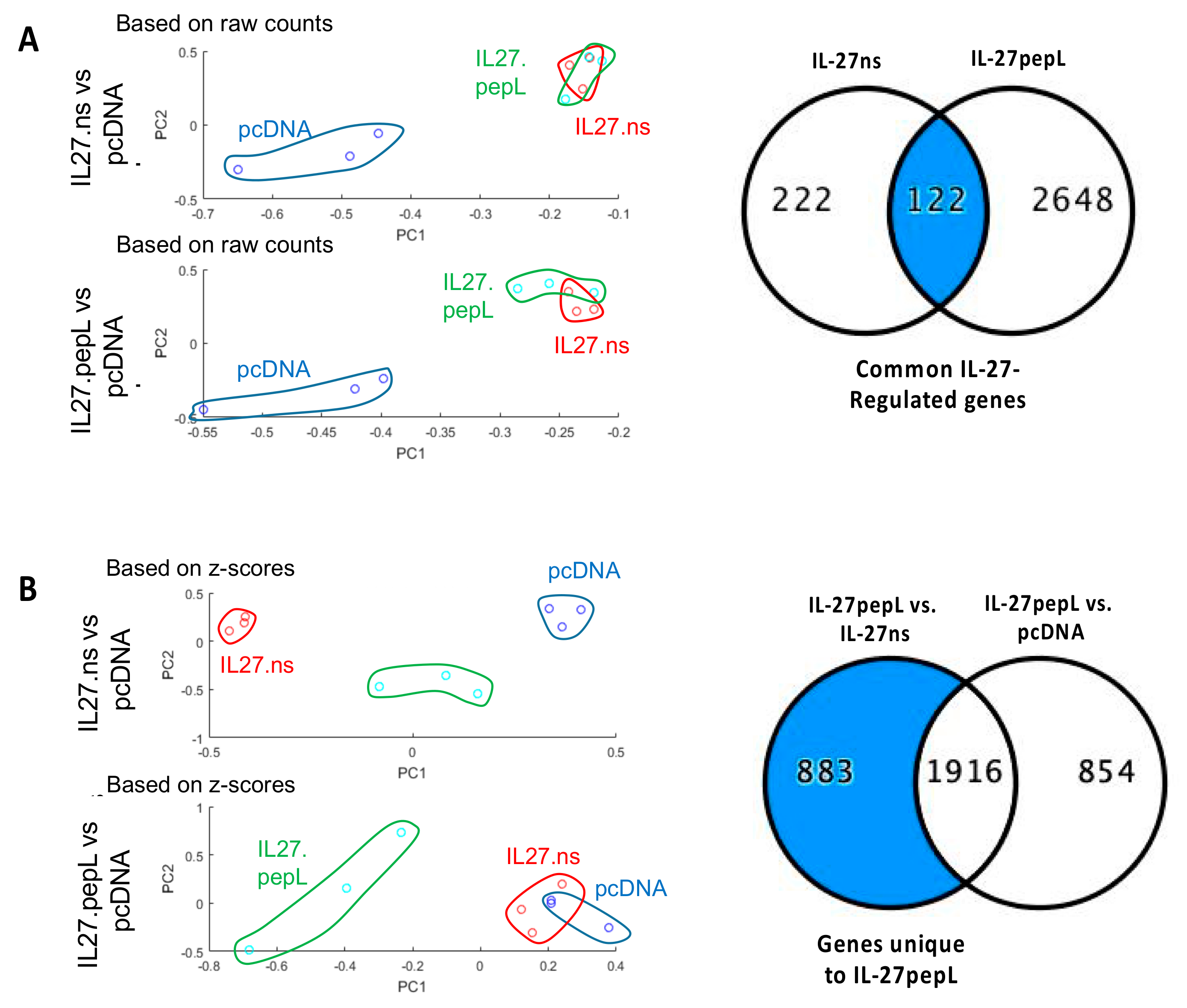{kind=link}
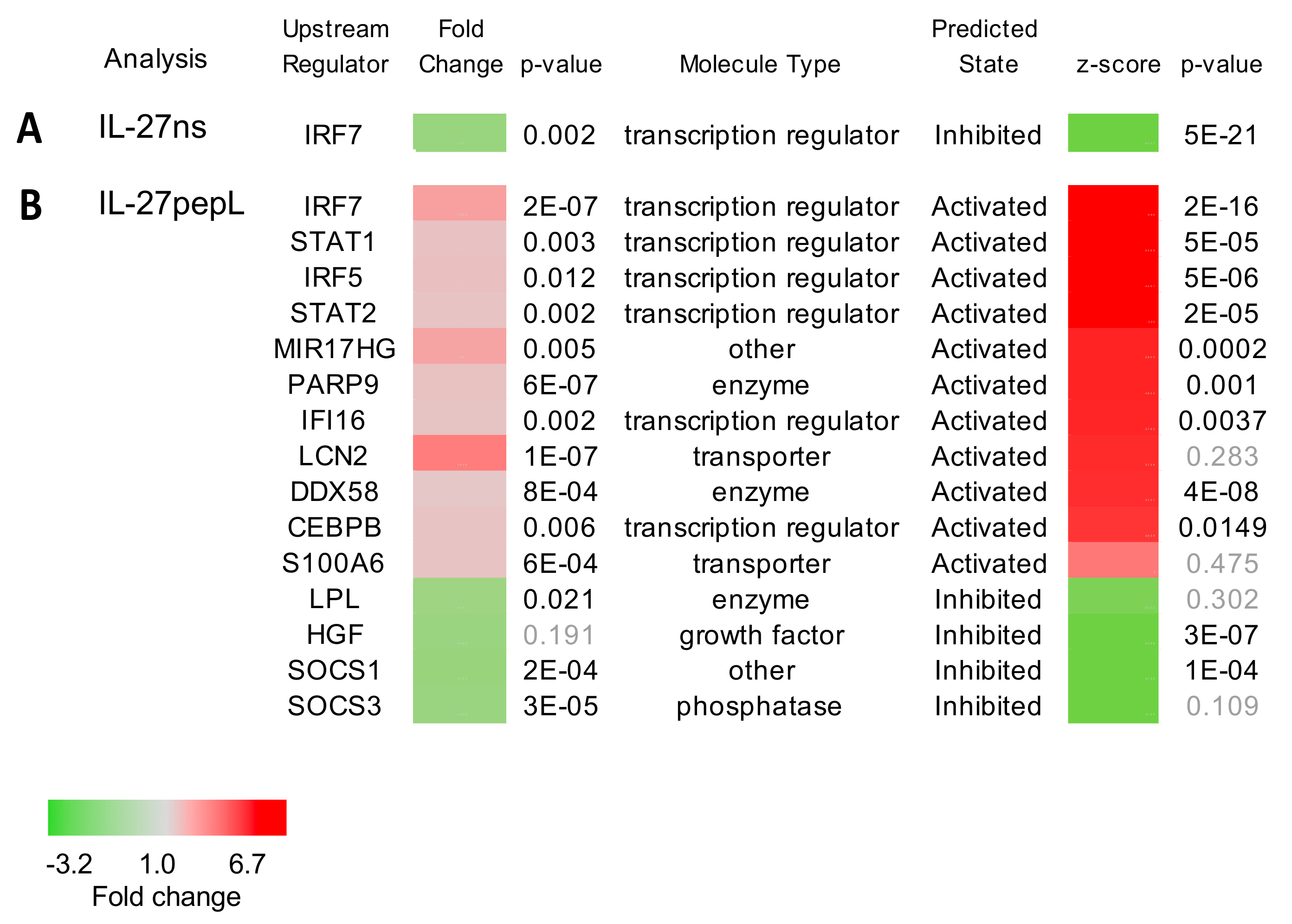{kind=link}
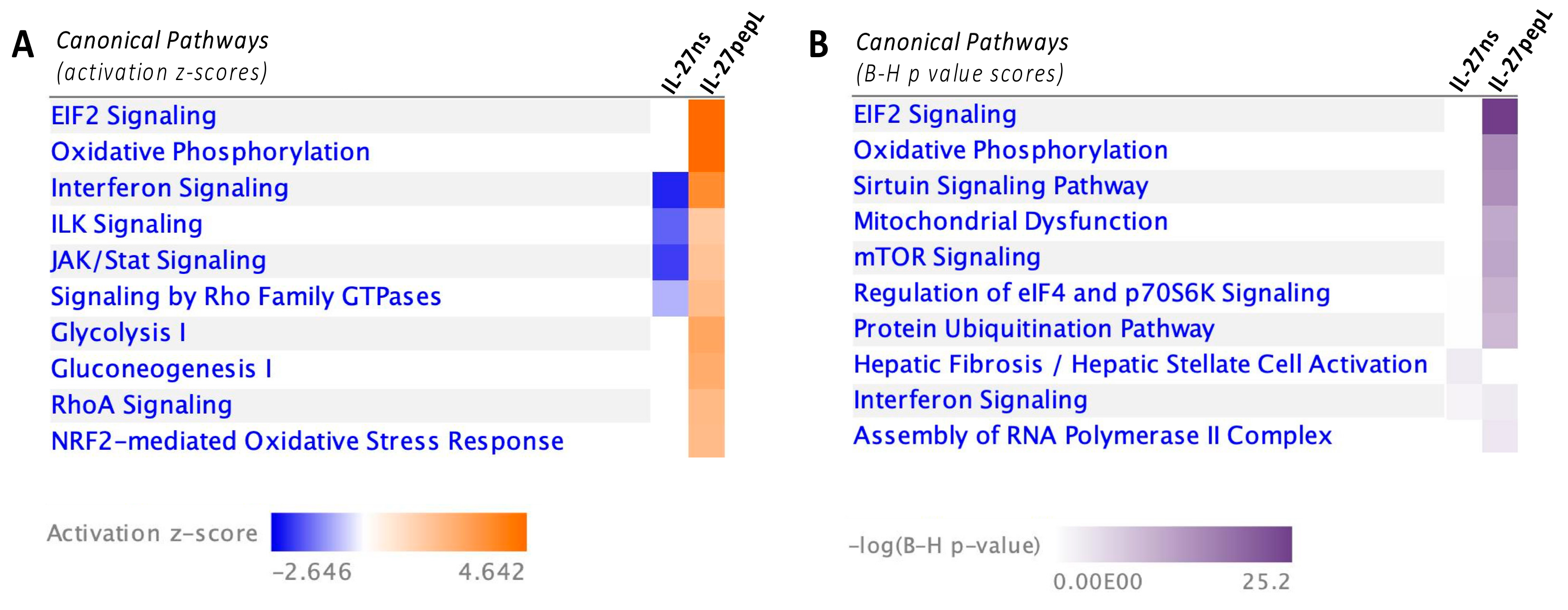{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Different Global Gene Expression Analyses Following RNAseq Data Collection Showed Commonalities and Differences for These Gene Therapy-Based IL-27 Therapeutic Candidates
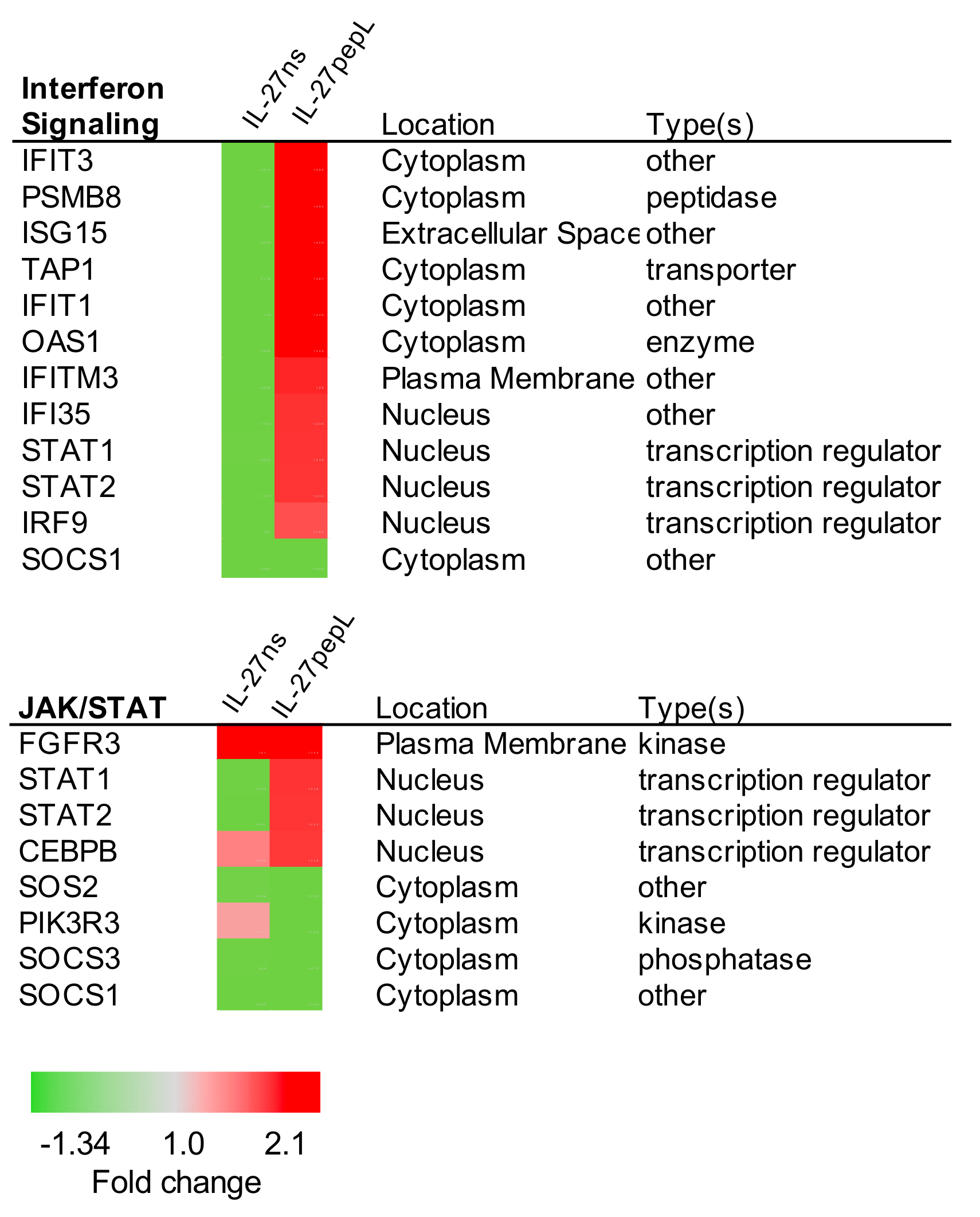
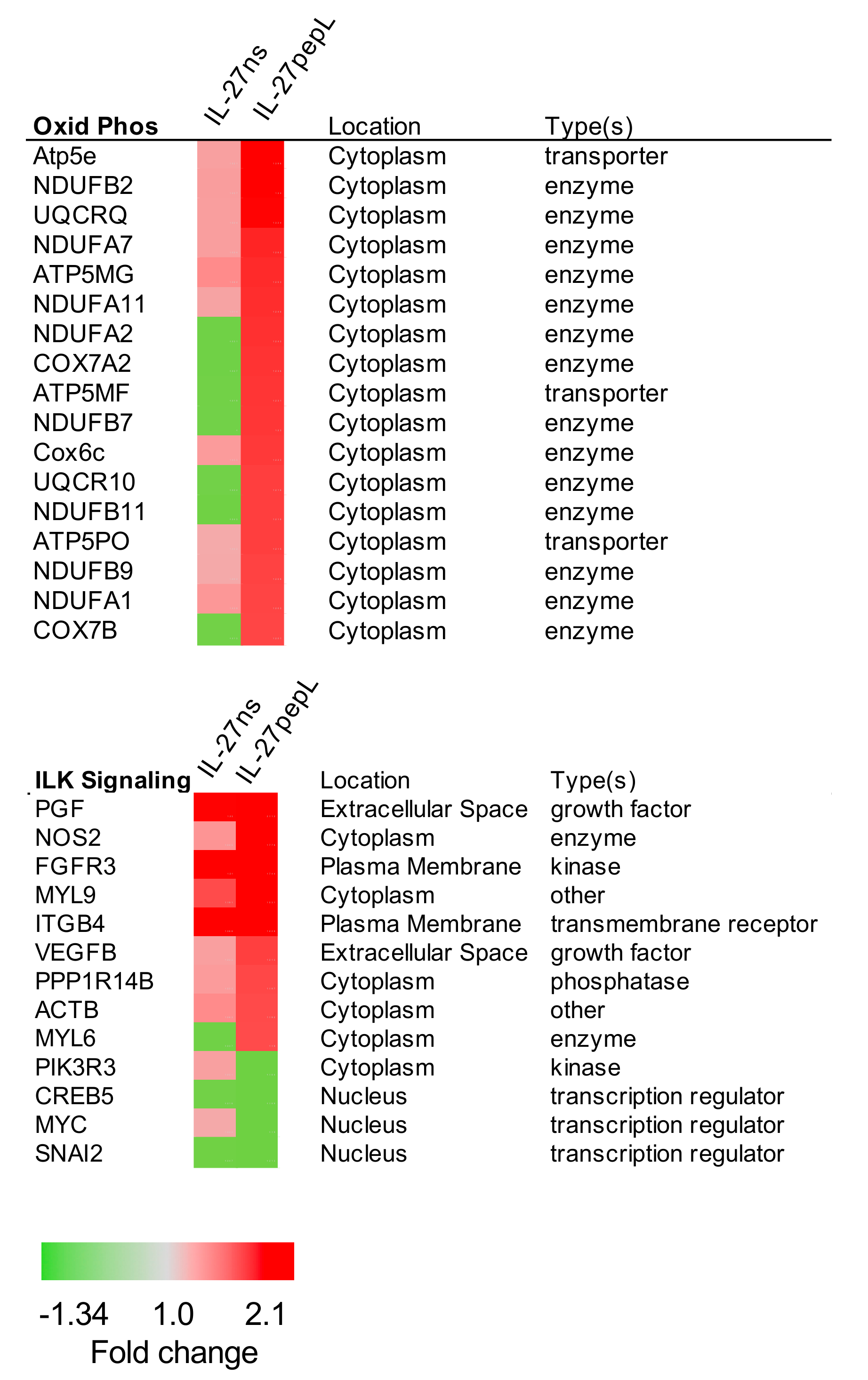
2.2. Ingenuity Pathway Analyses (IPA) Reported Specific Upstream Regulators and Canonical Pathways Differentially Modulated by IL-27pepL
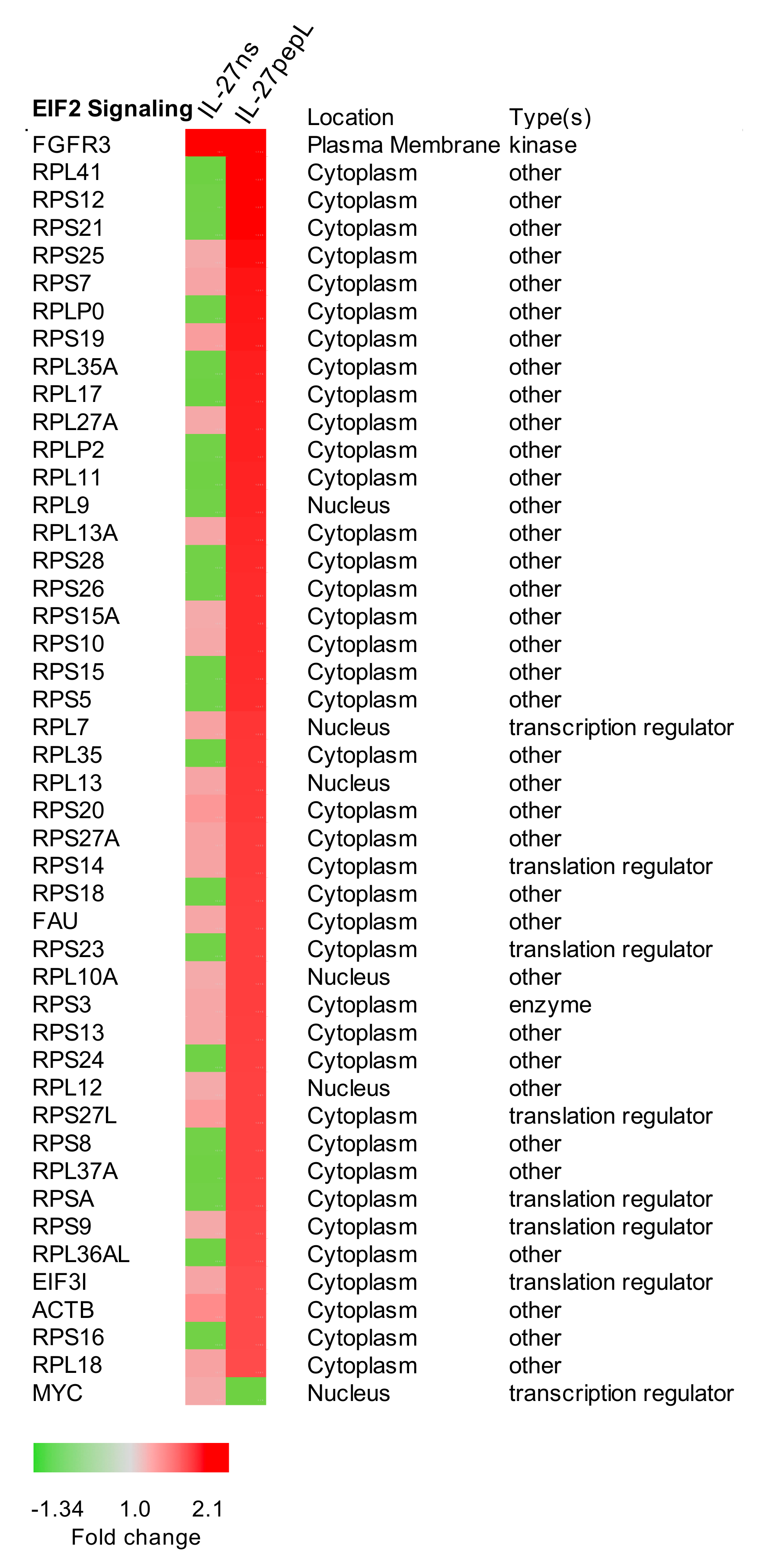
2.3. Expanded Analyses of the Top Five Canonical Pathways Differentially Activated by Targeted IL-27 Relative to Controls
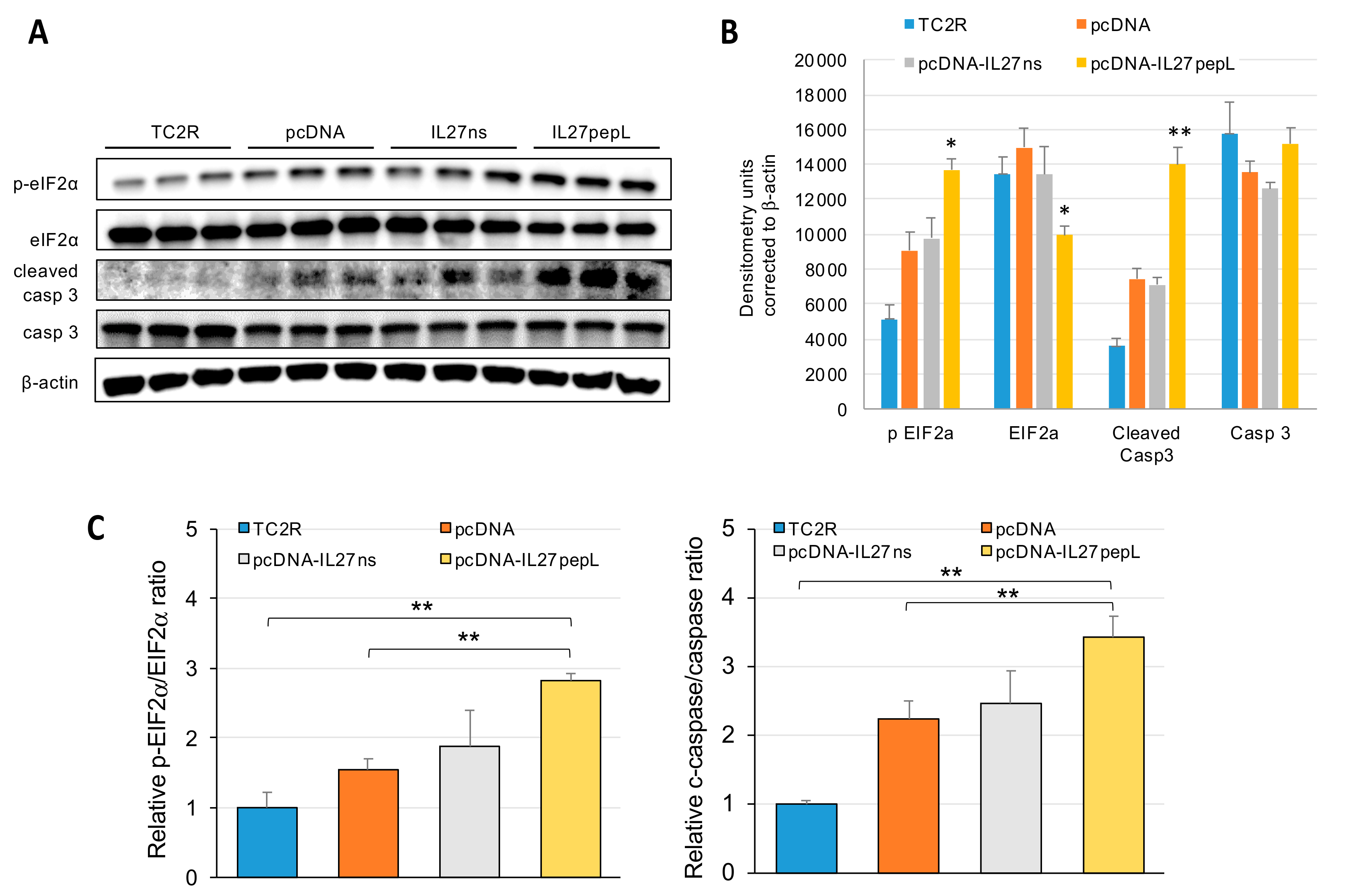
2.4. The Top Novel IL-27pepL-activated EIF2 Pathway Could Be Validated at the Protein Level
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Vectors, Modeling, and Transfection Assays
4.3. RNAseq and Principal Component Analysis (PCA)
4.4. Ingenuity Pathway Analyses (IPA)
4.5. Western Blots
4.6. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| eIF2α | eukaryotic translation initiation factor 2 alpha |
| ER | endoplasmic reticulum |
| DOAJ | directory of open access journals |
| IL-27 | interleukin-27 |
| IL-6-Rα | interleukin-6 receptor alpha |
| IPA | ingenuity pathway analyses |
| MDPI | Multidisciplinary Digital Publishing Institute |
| PCA | principal component analysis |
| TC2R | TRAMPC2-Ras murine prostate adenocarcinoma cells |
| RM1 | Ras/Myc murine prostate adenocarcinoma cells |
| RNAseq | RNA sequencing |
References
- Figueiredo Neto, M.; Figueiredo, M.L. Combination of Interleukin-27 and MicroRNA for Enhancing Expression of Anti-Inflammatory and Proosteogenic Genes. Arthritis 2017, 2017, 6365857. [Google Scholar] [CrossRef] [PubMed]
- Zolochevska, O.; Xia, X.; Williams, B.J.; Ramsay, A.; Li, S.; Figueiredo, M.L. Sonoporation delivery of Interleukin 27 gene therapy efficiently reduces prostate tumor cell growth in vivo. Human Gene Ther. 2011, 22, 1537–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolochevska, O.; Diaz-Quiñones, A.O.; Ellis, J.; Figueiredo, M.L. Interleukin-27 expression modifies prostate cancer cell crosstalk with bone and immune cells in vitro. J. Cell Physiol. 2013, 228, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Zolochevska, O.; Ellis, J.; Parelkar, S.; Chan-Seng, D.; Emrick, T.; Wei, J.; Patrikeev, I.; Motamedi, M.; Figueiredo, M.L. Interleukin-27 gene delivery for modifying malignant interactions between prostate tumor and bone. Hum. Gene Ther. 2013, 24, 970–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibra, D.; Cutrera, J.J.; Xia, X.; Birkenbach, M.P.; Li, S. Expression of WSX1 in tumors sensitizes IL-27 signaling-independent natural killer cell surveillance. Cancer Res. 2009, 69, 5505–5513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larousserie, F.; Bsiri, L.; Dumaine, V.; Dietrich, C.; Audebourg, A.; Radenen-Bussière, B.; Anract, P.; Vacher-Lavenu, M.C.; Devergne, O. Frontline Science: Human bone cells as a source of IL-27 under inflammatory conditions: Role of TLRs and cytokines. J. Leukoc. Biol. 2017, 101, 1289–1300. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Morishima, N.; Mizoguchi, I.; Shimizu, M.; Nagai, H.; Oniki, S.; Oka, M.; Nishigori, C.; Mizuguchi, J. Antiproliferative activity of IL-27 on melanoma. J. Immunol. 2008, 180, 6527–6535. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, A.; Lambert, Q.T.; Reuther, G.W. Transformation of hematopoietic cells and activation of JAK2-V617F by IL-27R, a component of a heterodimeric type I cytokine receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 18502–18507. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.; Ghilardi, N.; Li, J.; de Sauvage, F.J. IL-27 regulates IL-12 responsiveness of naive CD4+ T cells through Stat1-dependent and -independent mechanisms. Proc. Natl. Acad. Sci. USA 2003, 100, 15047–15052. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Hamano, S.; Yamanaka, A.; Hanada, T.; Ishibashi, T.; Mak, T.W.; Yoshimura, A.; Yoshida, H. Cutting edge: Role of IL-27/WSX-1 signaling for induction of T-bet through activation of STAT1 during initial Th1 commitment. J. Immunol. 2003, 170, 4886–4890. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Lee, D.A.; Li, S. IL-12 and IL-27 sequential gene therapy via intramuscular electroporation delivery for eliminating distal aggressive tumors. J. Immunol. 2010, 184, 2348–2354. [Google Scholar] [CrossRef]
- Figueiredo, M.; Figueiredo Neto, M.; Salameh, J.; Decker, R.; Letteri, R.; Chan-Seng, D.; Emrick, T. Ligand-mediated targeting of cytokine Interleukin-27 enhances its bioactivity in vivo. Mol. Ther. Methods Clin. Develop. 2020. [Google Scholar]
- Don-Doncow, N.; Marginean, F.; Coleman, I.; Nelson, P.S.; Ehrnström, R.; Krzyzanowska, A.; Morrissey, C.; Hellsten, R.; Bjartell, A. Expression of STAT3 in Prostate Cancer Metastases. Eur. Urol. 2017, 71, 313–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.L.; Lai, K.P.; Chen, C.A.; Yang, C.Y.; Chen, P.S.; Chang, C.C.; Chou, C.H.; Hu, C.L.; Kuo, M.L.; Hsieh, C.Y.; et al. A novel peptide specifically binding to interleukin-6 receptor (gp80) inhibits angiogenesis and tumor growth. Cancer Res. 2005, 65, 4827–4835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Morley, M.; Brandimarto, J.; Hannenhalli, S.; Hu, Y.; Ashley, E.A.; Tang, W.H.; Moravec, C.S.; Margulies, K.B.; Cappola, T.P.; et al. RNA-Seq identifies novel myocardial gene expression signatures of heart failure. Genomics 2015, 105, 83–89. [Google Scholar] [CrossRef]
- Wang, C.; Gong, B.; Bushel, P.R.; Thierry-Mieg, J.; Thierry-Mieg, D.; Xu, J.; Fang, H.; Hong, H.; Shen, J.; Su, Z.; et al. The concordance between RNA-seq and microarray data depends on chemical treatment and transcript abundance. Nat. Biotechnol. 2014, 32, 926–932. [Google Scholar] [CrossRef]
- Zhao, S.; Fung-Leung, W.P.; Bittner, A.; Ngo, K.; Liu, X. Comparison of RNA-Seq and microarray in transcriptome profiling of activated T cells. PLoS ONE 2014, 9, e78644. [Google Scholar] [CrossRef]
- Su, Q.; Wang, S.; Baltzis, D.; Qu, L.K.; Raven, J.F.; Li, S.; Wong, A.H.; Koromilas, A.E. Interferons induce tyrosine phosphorylation of the eIF2alpha kinase PKR through activation of Jak1 and Tyk2. EMBO Rep. 2007, 8, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.G.; Conn, C.S.; Kye, Y.; Xue, L.; Forester, C.M.; Cowan, J.E.; Hsieh, A.C.; Cunningham, J.T.; Truillet, C.; Tameire, F.; et al. Development of a stress response therapy targeting aggressive prostate cancer. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Liu, L.; He, X.; Wang, H.; Lin, W.; Yoon, S.O.; Wood, T.L.; Lu, Q.R. Regulation of PERK-eIF2α signalling by tuberous sclerosis complex-1 controls homoeostasis and survival of myelinating oligodendrocytes. Nat. Commun. 2016, 7, 12185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Hamamura, K.; Jiang, C.; Zhao, L.; Yokota, H. Salubrinal promotes healing of surgical wounds in rat femurs. J. Bone Miner. Metab. 2012, 30, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Hamamura, K.; Zhang, P.; Chen, Y.; Yokota, H. Systems analysis of bone remodelling as a homeostatic regulator. IET Syst. Biol. 2010, 4, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, M.; Ruan, H.; Qiu, W.; Xu, X.; Zhang, L.; Yu, J. Co-targeting translation and proteasome rapidly kills colon cancer cells with mutant RAS/RAF via ER stress. Oncotarget 2017, 8, 9280–9292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umbaugh, C.S.; Diaz-Quiñones, A.; Neto, M.F.; Shearer, J.J.; Figueiredo, M.L. A dock derived compound against laminin receptor (37 LR) exhibits anti-cancer properties in a prostate cancer cell line model. Oncotarget 2018, 9, 5958–5978. [Google Scholar] [CrossRef] [Green Version]
- Sockolosky, J.T.; Kivimäe, S.; Szoka, F.C. Fusion of a short peptide that binds immunoglobulin G to a recombinant protein substantially increases its plasma half-life in mice. PLoS ONE 2014, 9, e102566. [Google Scholar] [CrossRef]
- Manfredini, R.; Tenedini, E.; Siena, M.; Tagliafico, E.; Montanari, M.; Grande, A.; Zanocco-Marani, T.; Poligani, C.; Zini, R.; Gemelli, C.; et al. Development of an IL-6 antagonist peptide that induces apoptosis in 7TD1 cells. Peptides 2003, 24, 1207–1220. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Bluyssen, H.A.; Levy, D.E. Stat2 is a transcriptional activator that requires sequence-specific contacts provided by stat1 and p48 for stable interaction with DNA. J. Biol. Chem. 1997, 272, 4600–4605. [Google Scholar] [CrossRef] [Green Version]
- Raychaudhuri, S.; Stuart, J.M.; Altman, R.B. Principal components analysis to summarize microarray experiments: Application to sporulation time series. Pac. Symp. Biocomput. 2000, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Gopurappilly, R.; Bhonde, R. Transcriptional profiling and functional network analyses of islet-like clusters (ILCs) generated from pancreatic stem cells in vitro. Genomics 2015, 105, 211–219. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueiredo Neto, M.; Liu, S.; Salameh, J.W.; Yokota, H.; Figueiredo, M.L. Interleukin-27 Gene Delivery Targeting IL-6Rα-Expressing Cells as a Stress Response Therapy. Int. J. Mol. Sci. 2020, 21, 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031108
Figueiredo Neto M, Liu S, Salameh JW, Yokota H, Figueiredo ML. Interleukin-27 Gene Delivery Targeting IL-6Rα-Expressing Cells as a Stress Response Therapy. International Journal of Molecular Sciences. 2020; 21(3):1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031108
Chicago/Turabian StyleFigueiredo Neto, Manoel, Shengzhi Liu, Janelle Wes Salameh, Hiroki Yokota, and Marxa Leão Figueiredo. 2020. "Interleukin-27 Gene Delivery Targeting IL-6Rα-Expressing Cells as a Stress Response Therapy" International Journal of Molecular Sciences 21, no. 3: 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031108