An Arabidopsis Mutant Over-Expressing Subtilase SBT4.13 Uncovers the Role of Oxidative Stress in the Inhibition of Growth by Intracellular Acidification

 , , , , and
, , , , and 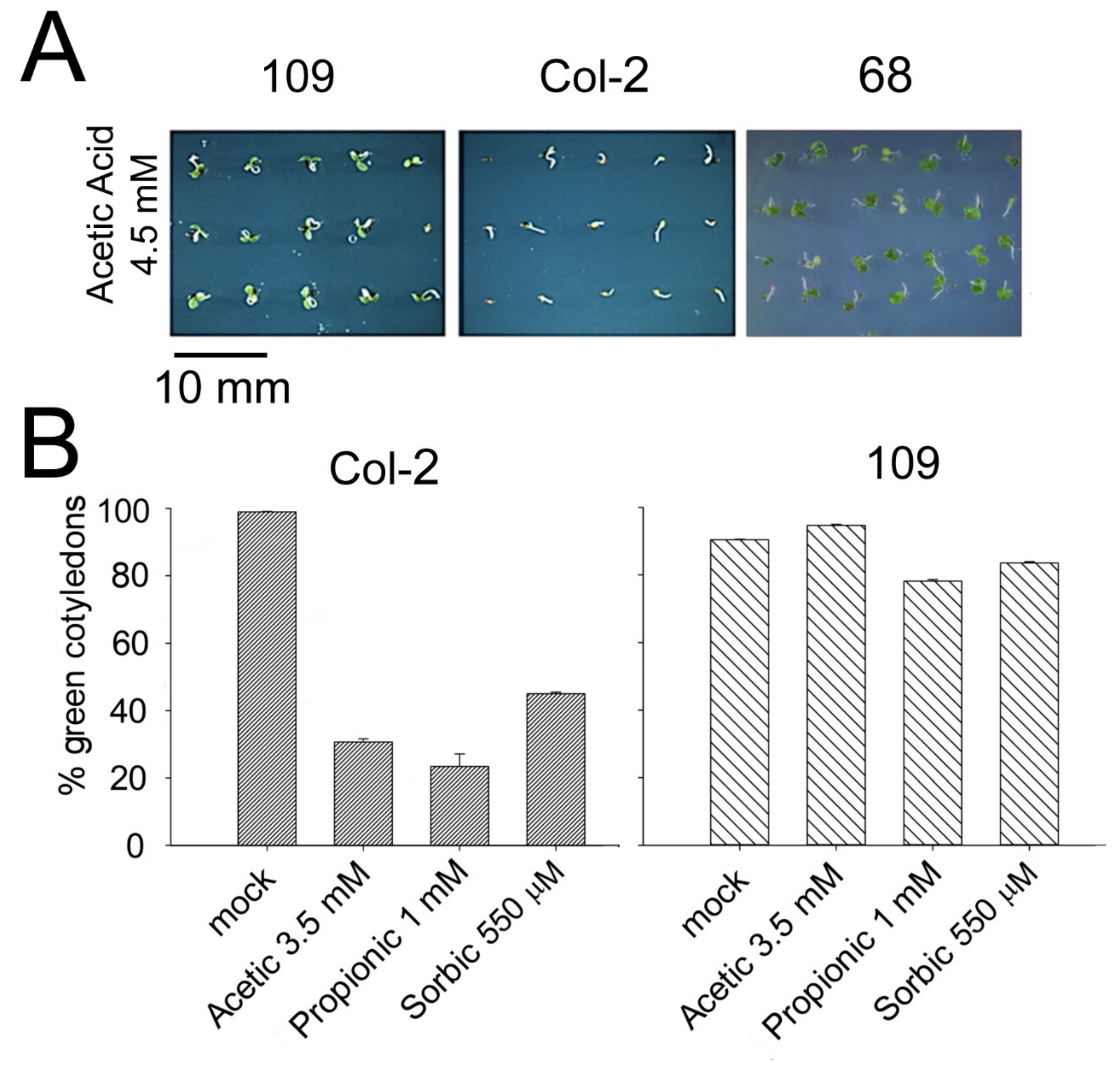{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
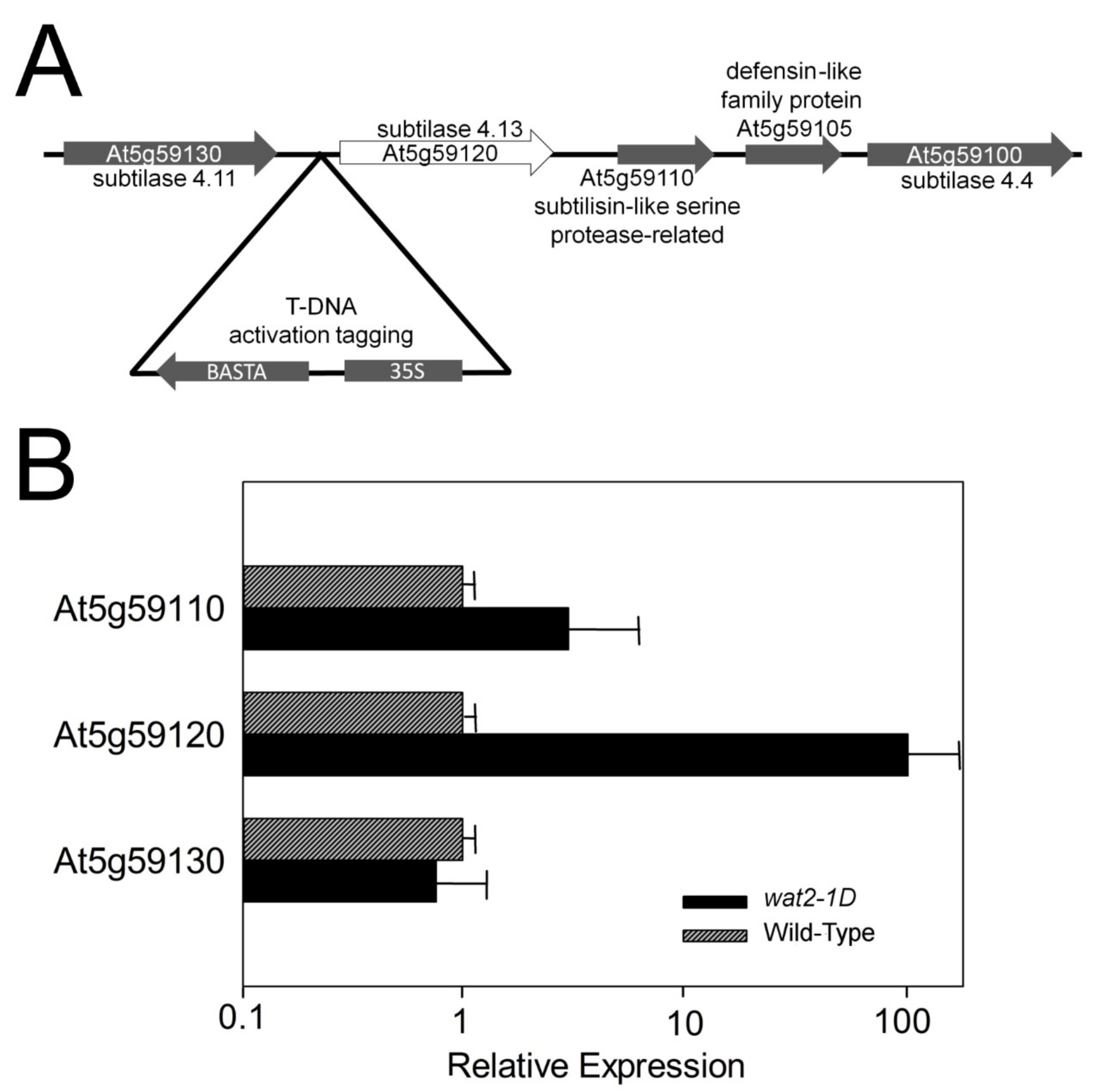
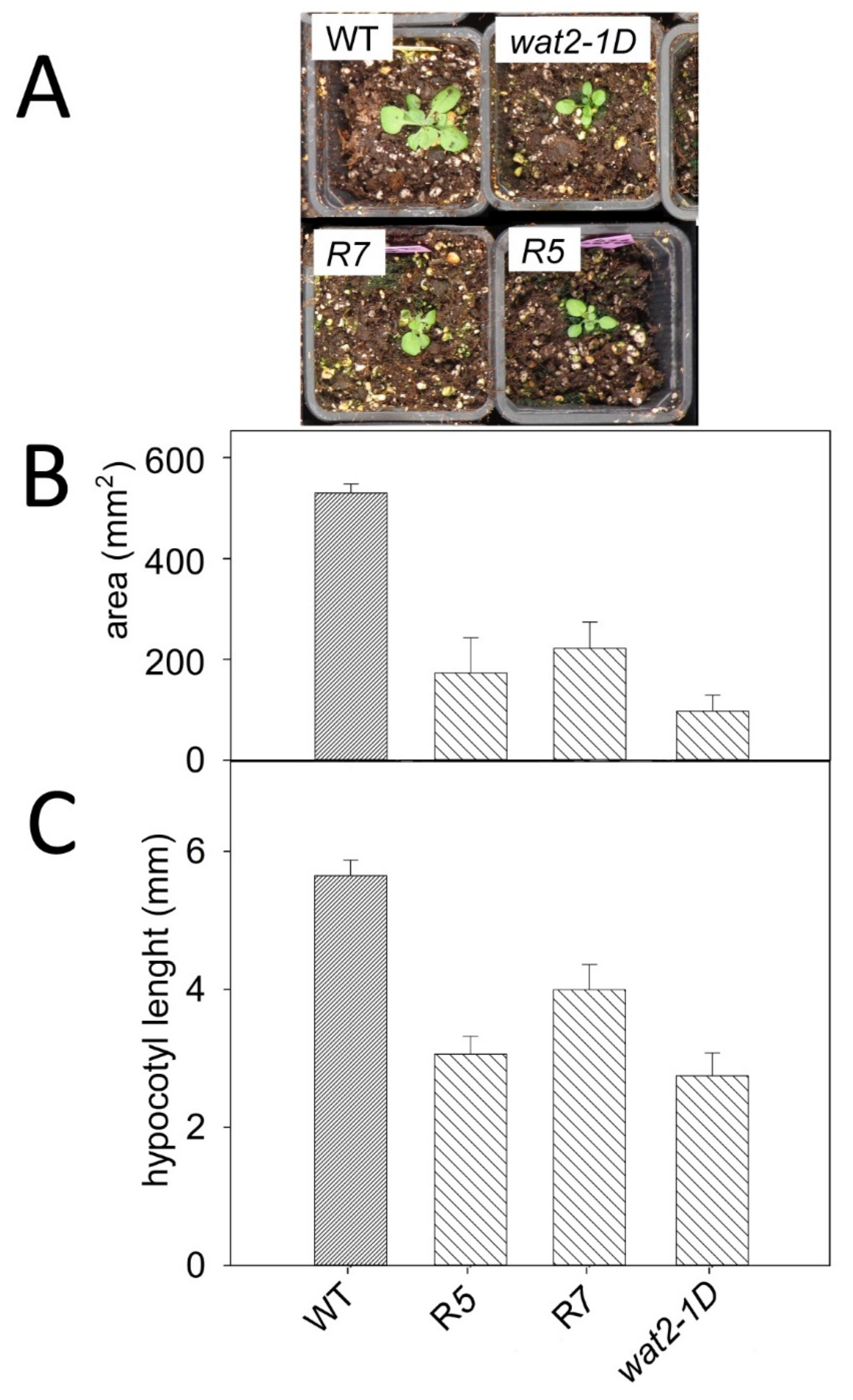
2.1. A Novel Mutant Tolerant to Weak Organic Acids Over-Expresses Subtilase SBT4.13
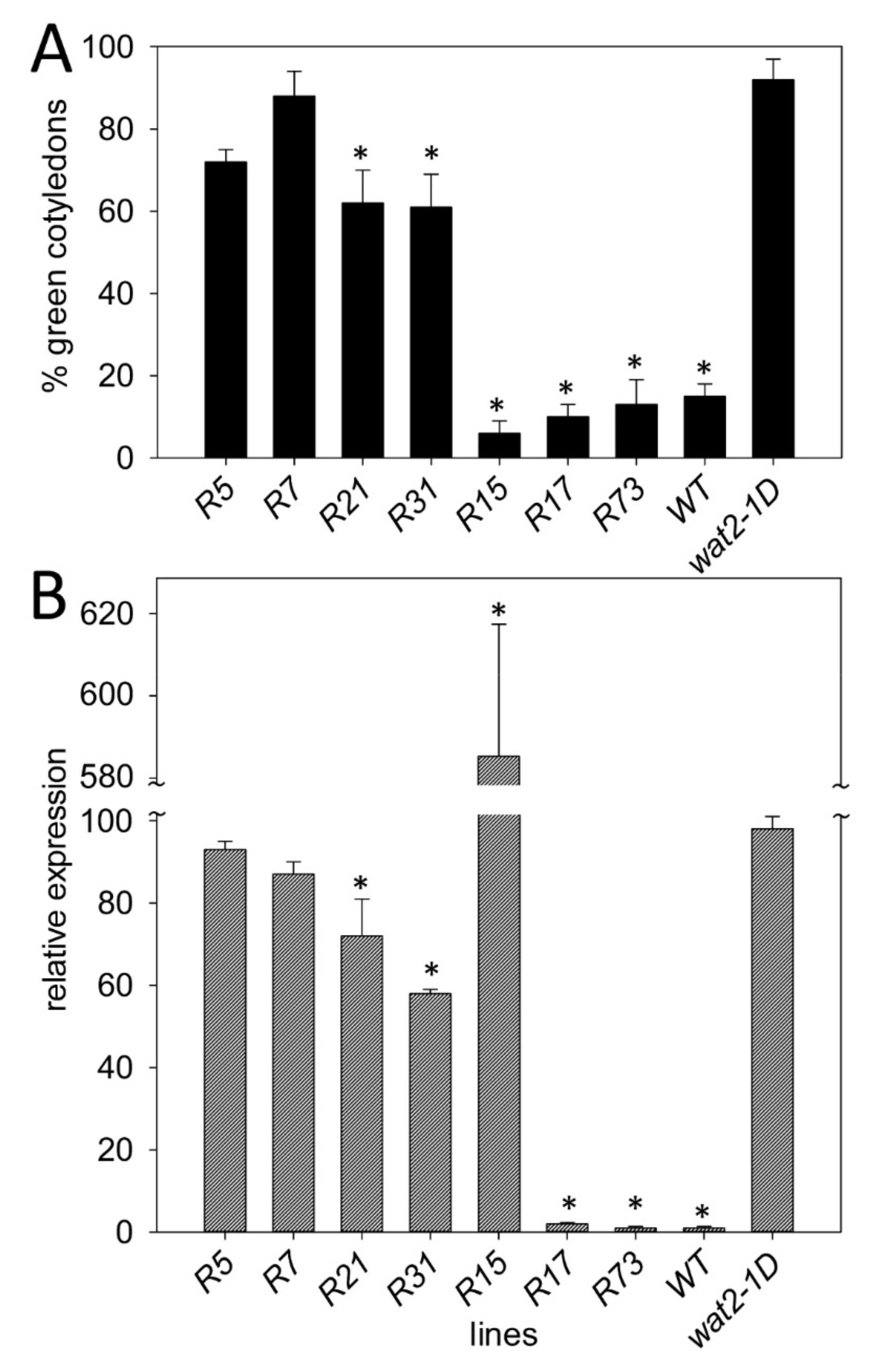
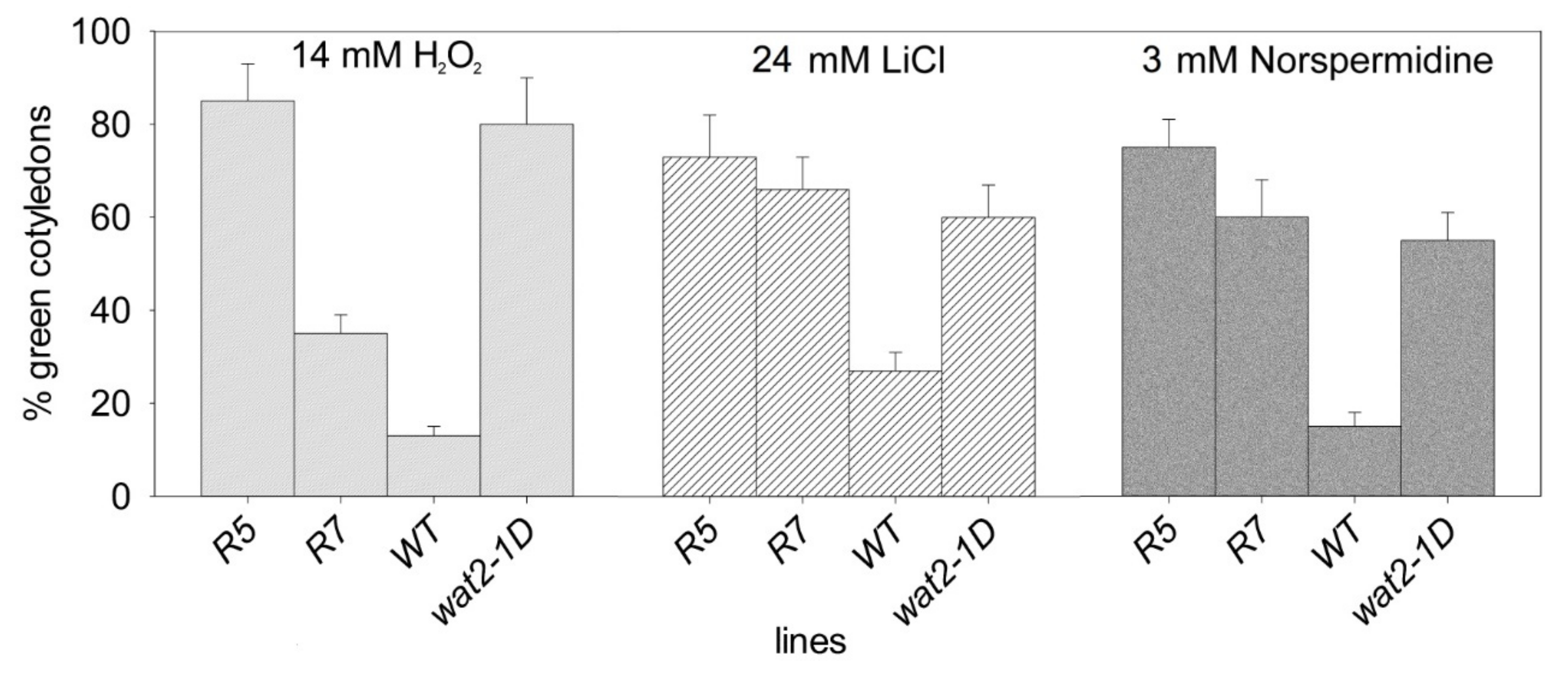
2.2. Recapitulation of Phenotypic Features of the Wat2-1D Mutant by Transgenic Plants Over-Expressing the SBT4.13 Gene
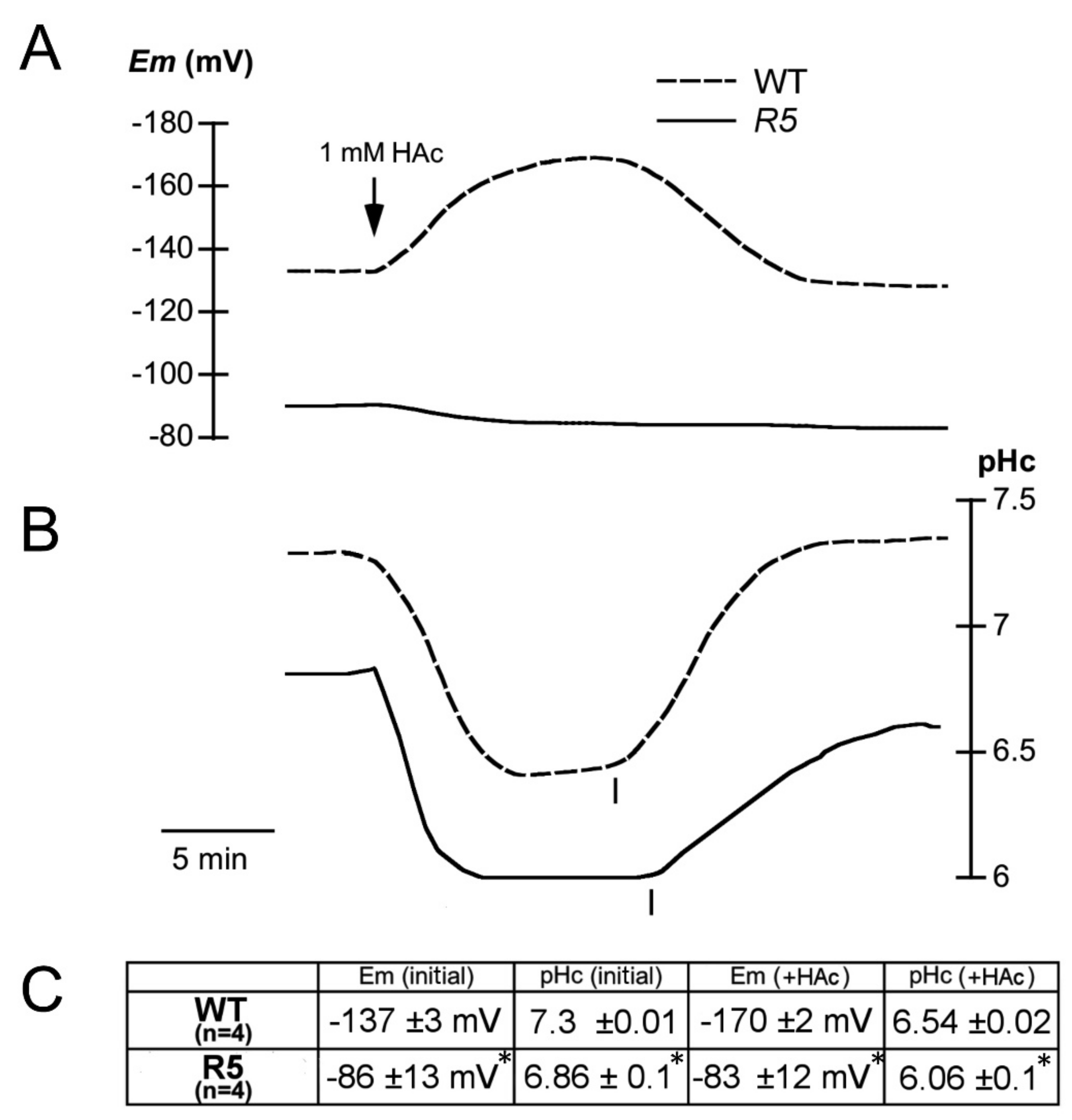
2.3. Electrophysiological Characterization of the sbt4.13-1D Mutant
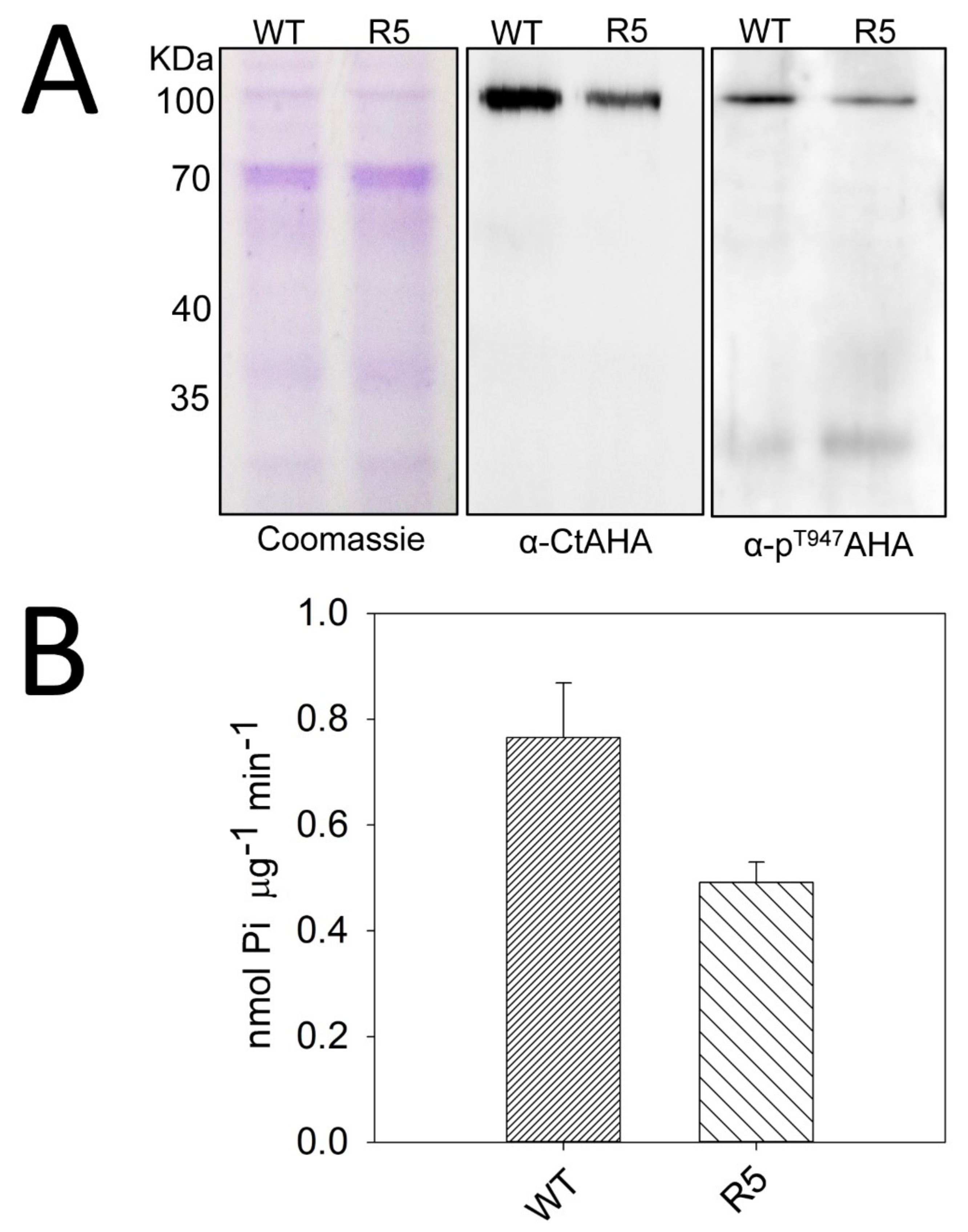
2.4. The sbt4.13-1D Mutant is Defective in Plasma Membrane H+-ATPase
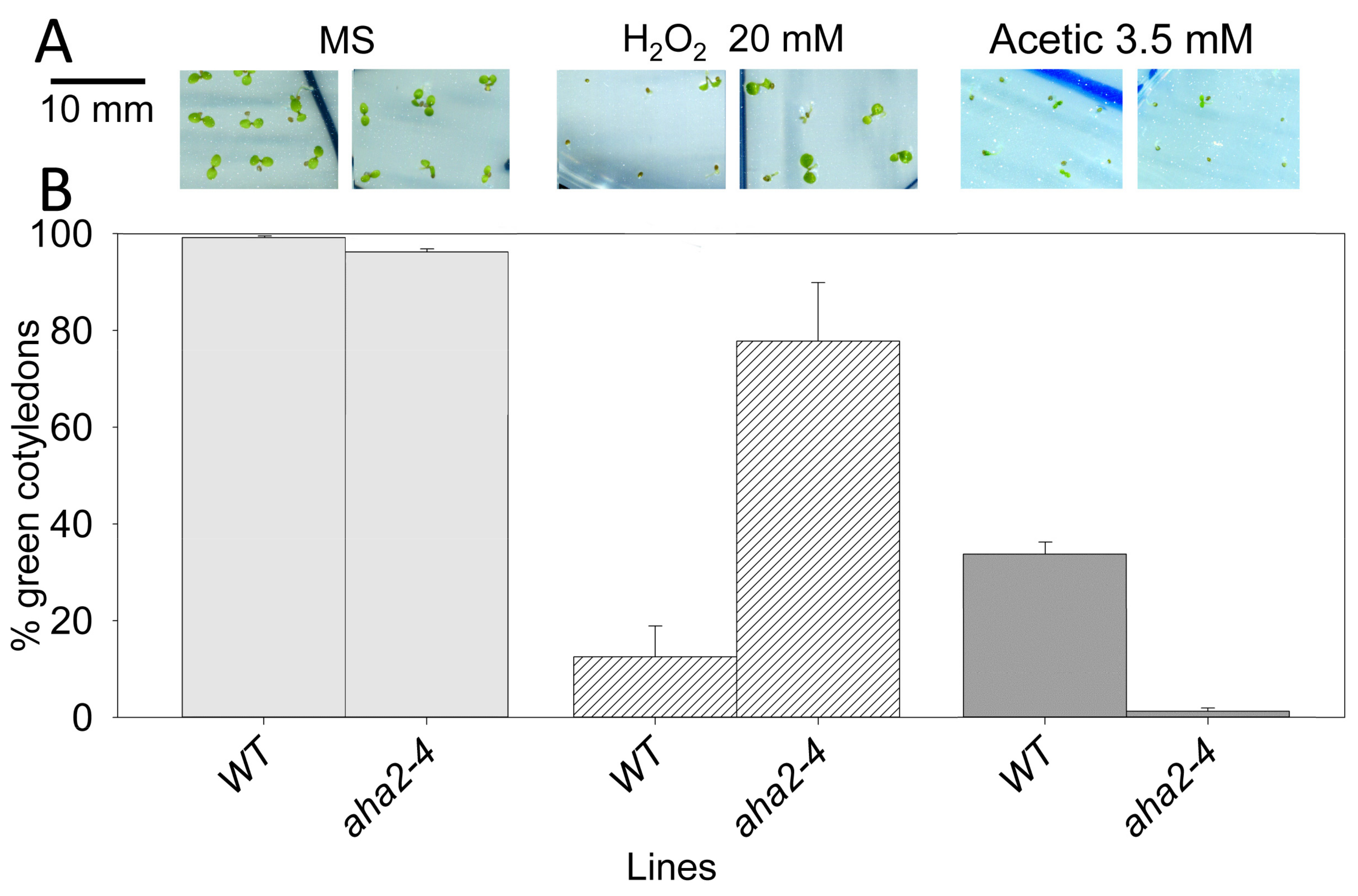
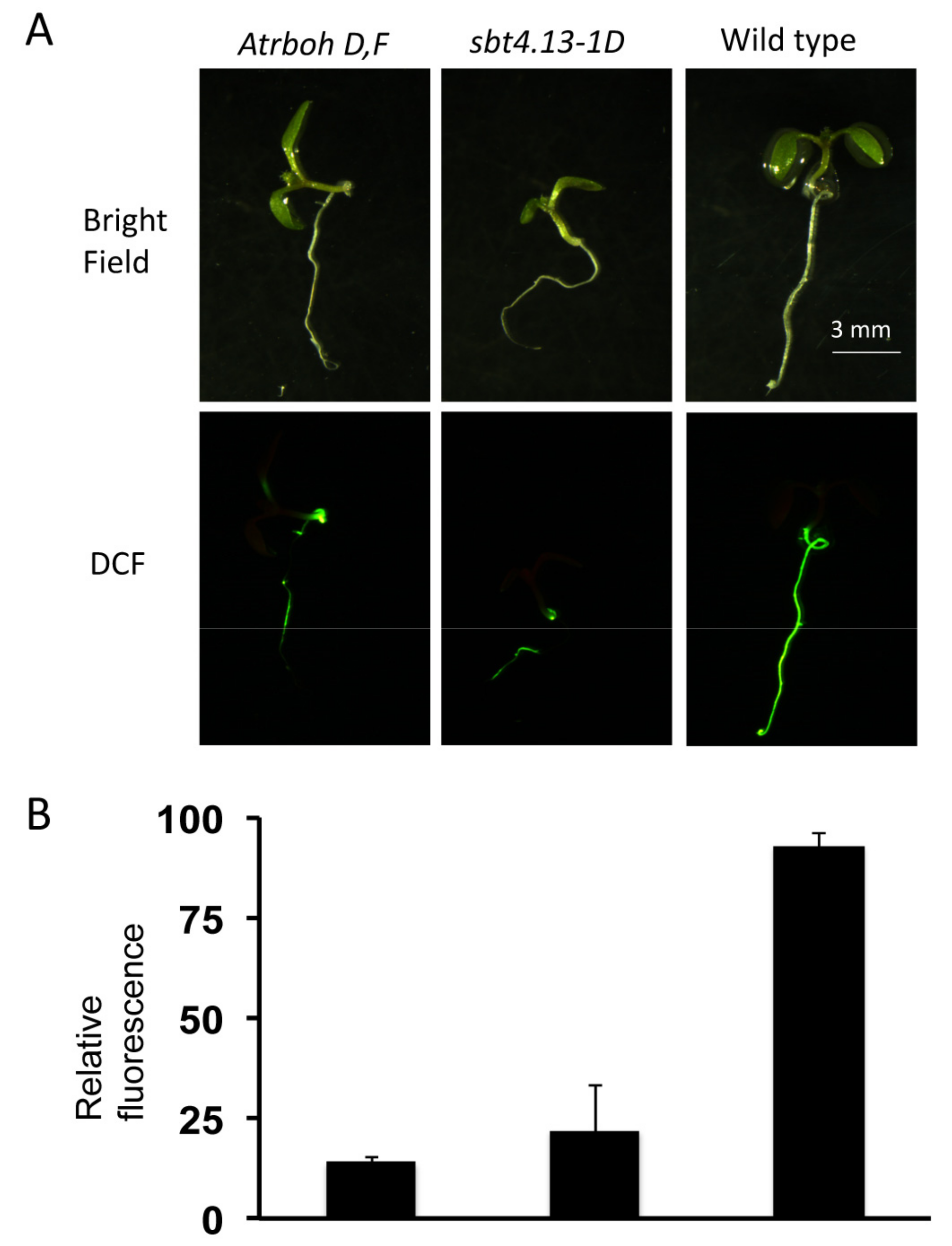
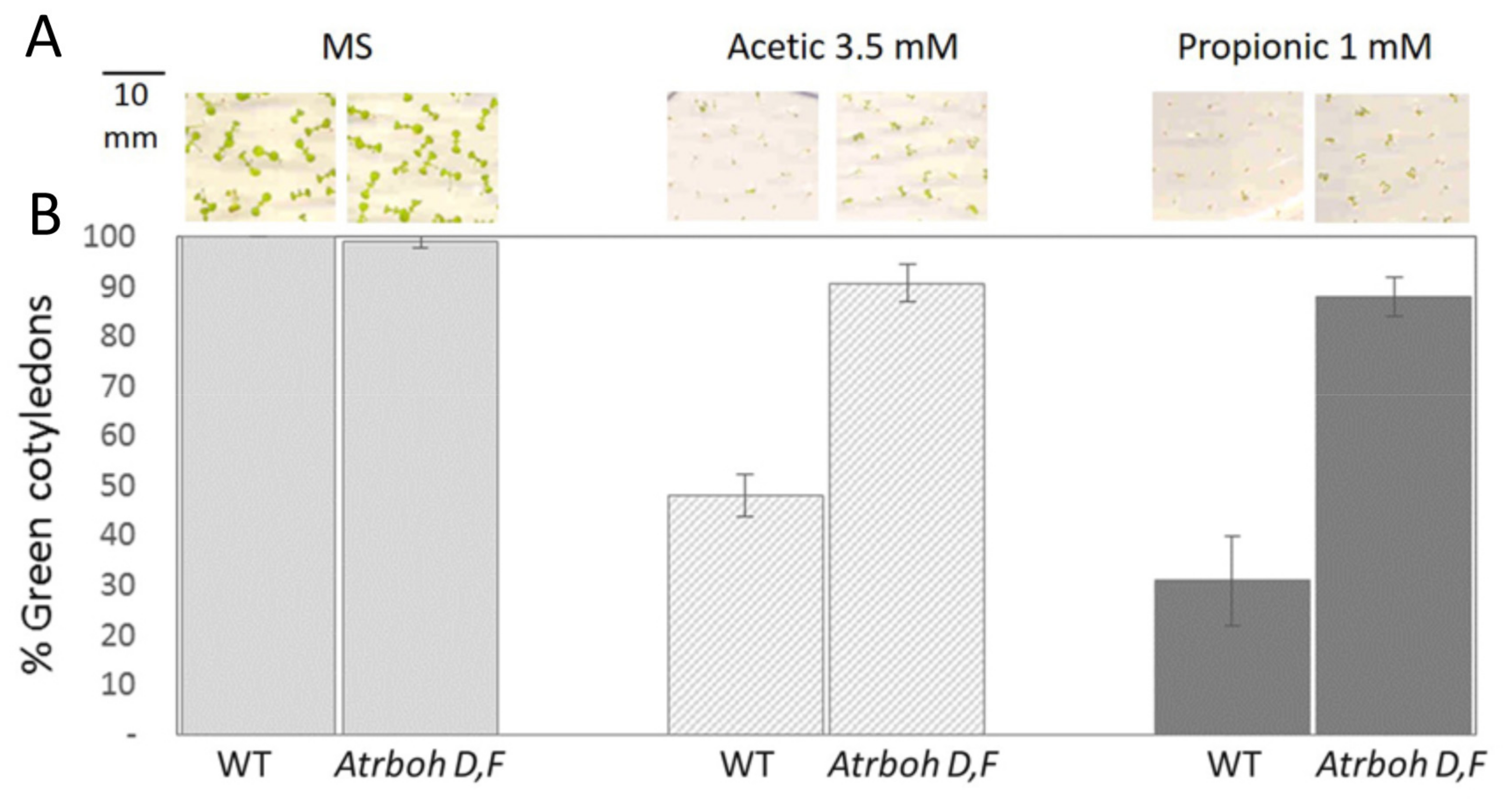
2.5. Intracellular Acidification Induces ROS Production by RBOH-D,F in Wild Type but not in sbt4.13-1D and the Atrboh-D,F Mutant is Acid Tolerant
3. Discussion
3.1. A novel Arabidopsis Mutant Tolerant to Acids Over-Expresses Subtilase SBT4.13 and Has Less Plasma Membrane H+-ATPase (PMA)
3.2. Intracellular Acidification Induces ROS Production Mediated by NADPH Oxidases (NOX), Explaining Growth Inhibition by Organic Acids
3.3. PMA and a Protein-Mediating Acid Activation of NOX are Probably Degraded by the Over-Expressed Subtilase of the sbt4.13-1D Mutant
3.4. Regulation of NADPH Oxidases by pH
3.5. A Novel Pathway for Growth Inhibition by Organic Acids through Activation of NOX
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Isolation and Genetic Characterization of the Wat2-1D Mutant
4.3. Molecular Characterization of the sbt4.13 Mutant
4.4. Construction of Transgenic Plants to Recapitulate the Mutant Phenotype
4.5. Transient Expression of SBT4.13-GFP in Nicotiana Benthamiana Leaves
4.6. Measurement of ROS Content in Plants with H2DCFDA
4.7. Transcriptomic Analysis by Microarrays
4.8. Other Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nuccitelli, R.; Heiple, J.M. Summary of the evidence and discussion concerning the involvement of pHi in the control of cellular functions. In Intracellular pH: Its Measurement, Regulation, and Utilization in Cellular Functions; Nuccitelli, R., Deamer, D.W., Eds.; Alan R. Liss: New York, NY, USA, 1982; pp. 567–586. [Google Scholar]
- Gottlieb, R.A.; Nordberg, J.; Skowronski, E.; Babior, B.M. Apoptosis induced in Jurkat cells by several agents is preceded by intracellular acidification. Proc. Natl. Acad. Sci. USA 1996, 93, 654–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obara, K.; Kuriyama, H.; Fukuda, H. Direct evidence of active and rapid nuclear degradation triggered by vacuole rupture during programmed cell death in Zinnia. Plant Physiol. 2001, 125, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, B.; Wightman, R.; Blanvillain, R.; Purcel, S.B.; Gallois, P. pH-sensitivity of YFP provides an intracellular indicator of programmed cell death. Plant Methods 2010, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuchi, S.; Koyama, H.; Iuchi, A.; Kobayashi, Y.; Kitabayashi, S.; Kobayashi, Y.; Ikka, T.; Hirayama, T.; Shinozaki, K.; Kobayashi, M. Zinc finger protein STOP1 is critical for proton tolerance in Arabidopsis and coregulates a key gene in aluminum tolerance. Proc. Natl. Acad. Sci. USA 2007, 104, 9900–9905. [Google Scholar] [CrossRef] [Green Version]
- Weitzel, G.; Pilatus, U.; Rensing, L. The cytoplasmic pH, ATP content and total protein synthesis rate during heat-shock protein inducing treatments in yeast. Exp. Cell Res. 1987, 170, 64–79. [Google Scholar] [CrossRef]
- Kiang, J.G.; McKinney, L.C.; Gallin, E.K. Heat induces intracellular acidification in human A-431 cells: Role of Na(+)-H+ exchange and metabolism. Am. J. Physiol. 1990, 259, C727–C737. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.J.; Black, C.R.; Miller, A.J. The role of cytosolic potassium and pH in the growth of barley roots. Plant Physiol. 1998, 118, 957–964. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Smith, F.A. The evolution of chemiosmotic energy coupling. J. Theor. Biol. 1976, 57, 301–312. [Google Scholar] [CrossRef]
- Kulichikhin, K.Y.; Chirkova, T.V.; Fagerstedt, K.V. Intracellular pH in rice and wheat root tips under hypoxic and anoxic conditions. Plant Signal. Behav. 2008, 3, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.D. The fine control of cytosolic pH. Physiol. Plant. 1986, 67, 702–706. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Palmgren, M.G.; Schumacher, K. Plant proton pumps. FEBS Lett. 2007, 581, 2204–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Planes, M.D.; Niñoles, R.; Rubio, L.; Bissoli, G.; Bueso, E.; García-Sánchez, M.J.; Alejandro, S.; Gonzalez-Guzmán, M.; Hedrich, R.; Rodriguez, P.L.; et al. A mechanism of growth inhibition by abscisic acid in germinating seeds of Arabidopsis thaliana based on inhibition of plasma membrane H+-ATPase and decreased cytosolic pH, K+, and anions. J. Exp. Bot. 2015, 66, 813–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawaki, Y.; Iuchi, S.; Kobayashi, Y.; Ikka, T.; Sakurai, N.; Fujita, M.; Shinozaki, K.; Shibata, D.; Kobayashi, M.; Koyama, H. STOP1 regulates multiple genes that protect Arabidopsis from proton and aluminum toxicities. Plant Physiol. 2009, 150, 281–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.; Lou, H.Q.; Yang, J.L.; Zheng, S.J. The roles of STOP1-like transcription factors in aluminum and proton tolerance. Plant Signal. Behav. 2016, 11, e1131371. [Google Scholar] [CrossRef] [PubMed]
- Niñoles, R.; Rubio, L.; García-Sánchez, M.J.; Fernández, J.A.; Bueso, E.; Alejandro, S.; Serrano, R. A dominant-negative form of Arabidopsis AP-3 β-adaptin improves intracellular pH homeostasis. Plant J. 2013, 74, 557–568. [Google Scholar] [CrossRef]
- Bissoli, G.; Niñoles, R.; Fresquet, S.; Palombieri, S.; Bueso, E.; Rubio, L.; García-Sánchez, M.J.; Fernández, J.A.; Mulet, J.M.; Serrano, R. Peptidyl-prolyl cis-trans isomerase ROF2 modulates intracellular pH homeostasis in Arabidopsis. Plant J. 2012, 70, 704–716. [Google Scholar] [CrossRef] [Green Version]
- Alejandro, S.; Rodriguez, P.L.; Bellés, J.M.; Yenush, L.; García-Sanchez, M.J.; Fernández, J.A.; Serrano, R. An Arabidopsis quiescin-sulphydryl oxidase regulates cation homeostasis at the root symplast-xylem interface. EMBO J. 2007, 26, 3203–3215. [Google Scholar] [CrossRef] [Green Version]
- Bobik, K.; Boutry, M.; Duby, G. Activation of the plasma membrane H+-ATPase by acid stress. Plant Signal. Behav. 2010, 5, 681–683. [Google Scholar] [CrossRef] [Green Version]
- Haruta, H.; Sussman, M.R. The effect of genetically reduced plasma membrane protonmotive force on vegetative growth of Arabidopsis. Plant Physiol. 2012, 158, 1158–1171. [Google Scholar] [CrossRef] [Green Version]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.L.; Li, W.Y.; Miao, H.; Yang, S.Q.; Li, R.; Wang, X.; Li, W.Q.; Chen, K.M. Comprehensive genomic analysis and expression profiling of the NOX gene families under abiotic stresses and hormones in plants. Genome Biol. Evol. 2016, 8, 791–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, J.M.; Mori, I.C.; Pei, Z.-M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bdde, S.; Jones, J.D.G.; Schroeder, J.L. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. [Google Scholar] [CrossRef]
- Keller, T.; Damude, H.G.; Werner, D.; Doerner, P.; Dixon, R.A.; Lamb, C. A plant homolog of the neutrophil NADPH oxidase gp91phox subunit gene encodes a plasma membrane protein with Ca2+ binding motifs. Plant Cell 1998, 10, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flinck, M.; Kramer, S.H.; Pedersen, S.F. Roles of pH in control of cell proliferation. Acta Physiol. 2018, 223, e13068. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Hirai, Y. Good and bad protons: Genetic aspects of acidity stress responses in plants. J. Exp. Bot. 2016, 67, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechant, R.; Saad, S.; Ibañez, A.J.; Peter, M. Cytosolic pH regulates cell growth through distinct GTPases, Arf1 and Gtr1, to promote ras/PKA and TORC1 activity. Mol. Cell. 2014, 55, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Balgi, A.D.; Diering, G.H.; Donohue, E.; Lam, K.K.; Fonseca, B.D.; Zimmerman, C.; Numata, M.; Roberge, M. Regulation of mTORC1 signaling by pH. PLoS ONE 2011, 6, 6–11. [Google Scholar] [CrossRef]
- Lucas, C.A.; Gillies, R.J.; Olson, J.E.; Giuliano, K.A.; Martinez, R.; Sneider, J.M. Intracellular acidification inhibits the proliferative response in BALB/c-3T3 cells. J. Cell Physiol. 1988, 136, 161–167. [Google Scholar] [CrossRef]
- Yan, L.; Du, C.; Riaz, M.; Jiang, C. Boron mitigates citrus root injuries by regulating intracellular pH and reactive oxygen species to resist H+-toxicity. Environ. Pollut. 2019, 255, 113254. [Google Scholar] [CrossRef]
- Riemann, A.; Schneider, B.; Ihling, A.; Nowak, M.; Sauvant, C.; Thews, O.; Gekle, M. Acidic environment leads to ROS-induced MAPK signaling in cancer cells. PLoS ONE 2011, 6, e22445. [Google Scholar] [CrossRef]
- Abramov, A.Y.; Jacobson, J.; Wientjes, F.; Hothersall, J.; Canevari, L.; Duchen, M.R. Expression and modulation of an NADPH oxidase in mammalian astrocytes. J. Neurosci. 2005, 25, 9176–9184. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A.; Stintzi, A.; Rivas, S.; Serrano, I.; Chichkova, N.V.; Vartapetian, A.B.; Martínez, D.; Guiamét, J.J.; Sueldo, D.J.; van der Hoorn, R.A.L.; et al. From structure to function—A family portrait of plant subtilases. New Phytol. 2018, 218, 901–915. [Google Scholar] [CrossRef] [PubMed]
- Schardon, K.; Hohl, M.; Graff, L.; Pfannstiel, J.; Schulze, W.; Stintzi, A.; Schaller, A. Precursor processing for plant peptide hormone maturation by subtilisin-like serine proteinases. Science 2016, 354, 1594–1597. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Xu, X.; Gao, M.; Li, J.; Guo, C.; Song, J.; Wang, X. Genome-wide analysis of respiratory burst oxidase homologs in grape (Vitis vinifera L.). Int. J. Mol. Sci. 2013, 14, 24169–24186. [Google Scholar] [CrossRef]
- Henderson, L.M.; Chapell, J.B.; Jones, O.T.G. The superoxide-generating NADPH oxidase of human neutrophils is electrogenic and associated with an H+ channel. Biochem. J. 1987, 246, 325–329. [Google Scholar] [CrossRef]
- Hoeberichts, F.A.; Perez-Valle, J.; Montesinos, C.; Mulet, J.M.; Planes, M.D.; Hueso, G.; Yenush, L.; Sharma, S.C.; Serrano, R. The role of K+ and H+ transport systems during glucose- and H2O2-induced cell death in Saccharomyces cerevisiae. Yeast 2010, 27, 713–725. [Google Scholar] [CrossRef]
- Wall, S.B.; Oh, J.Y.; Diers, A.R.; Landar, A. Oxidative modification of proteins: An emerging mechanism of cell signaling. Front. Physiol. 2012, 3, 369. [Google Scholar] [CrossRef] [Green Version]
- Felle, H.H. pH: Signal and messenger in plant cells. Plant Biol. 2001, 3, 577–591. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Qu, Y.; Yan, M.; Zhang, Q. Functional regulation of plant NADPH oxidase and its role in signaling. Plant Signal. Behav. 2017, 12, e1356970. [Google Scholar] [CrossRef] [Green Version]
- Han, J.P.; Köster, P.; Drerup, M.M.; Scholz, M.; Li, S.; Edel, K.H.; Hashimoto, K.; Kuchitsu, K.; Hippler, M.; Kudla, J. Fine-tuning of RBOHF activity is achieved by differential phosphorylation and Ca2+ binding. New Phytol. 2019, 221, 1935–1949. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Shang, Z.; Shin, R.; Thompson, E.; Rubio, L.; Laohavisit, A.; Mortimer, J.C.; Chivasa, S.; Slabas, A.R.; Glover, B.J.; et al. Plant extracellular ATP signalling by plasma membrane NADPH oxidase and Ca2+ channels. Plant J. 2009, 58, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-dependent ROS synthesis and ROS scavenging by plant specialized metabolites to modulate plant development and stress responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef] [PubMed]
- Felle, H.H. Cytoplasmic free calcium in Riccia fluitans L. and Zea mays L.: Interaction of Ca2+ and pH? Planta 1988, 176, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Henderson, L.M.; Chappell, J.B. NADPH oxidase of neutrophils. Biochim. Biophys. Acta 1996, 1273, 87–107. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa, T.; Nakazawa, M.; Kawashima, M.; Iizumi, H.; Kuroda, H.; Kondou, Y.; Tuhara, Y.; Suzuki, K.; Ishikawa, A.; Seki, M.; et al. The FOX hunting system: An alternative gain-of-function gene hunting technique. Plant J. 2006, 48, 974–985. [Google Scholar] [CrossRef]
- Brewer, T.F.; Garcia, F.J.; Onak, C.S.; Carroll, K.S.; Chang, C.J. Chemical approaches to discovery and study of sources and targets of hydrogen peroxide redox signaling through NADPH oxidase proteins. Annu. Rev. Biochem. 2015, 84, 765–790. [Google Scholar] [CrossRef]
- Brachi, B.; Morris, G.P.; Borevitz, J.O. Genome-wide association studies in plants: The missing heritability is in the field. Genome Biol. 2011, 12, 232. [Google Scholar] [CrossRef] [Green Version]
- Togninalli, M.; Seren, Ü.; Freudenthal, J.A.; Monroe, J.G.; Meng, D.; Nordborg, M.; Weigel, D.; Borgwardt, K.; Korte, A.; Grimm, D.G. AraPheno and the AraGWAS Catalog 2020: A major database update including RNA-Seq and knockout mutation data for Arabidopsis thaliana. Nucleic Acids Res. 2020, 48, D1063–D1068. [Google Scholar] [CrossRef]
- Haruta, M.; Burch, H.L.; Nelson, R.B.; Barrett-Wilt, G.; Kline, K.G.; Mohsin, S.B.; Young, J.C.; Otegui, M.S.; Sussman, M.R. Molecular characterization of mutant Arabidopsis plants with reduced plasma membrane proton pump activity. J. Biol. Chem. 2010, 285, 17918–17929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, M.; Carninci, P.; Nishiyama, Y.; Hayashizaki, Y.; Shinozaki, K. High-efficiency cloning of Arabidopsis full-length cDNA by biotinylated CAP trapper. Plant J. 1998, 15, 707–720. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Kamiya, A.; Ishida, J.; Satou, M.; Sakurai, T.; Nakajima, M.; Enju, A.; Akiyama, K.; Oono, Y.; et al. Functional annotation of a full-length Arabidopsis cDNA collection. Science 2002, 296, 141–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, M.D.; Grossniklaus, U. A Gateway cloning vector set for high throughput functional analysis of genes in plants. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.; Chaban, C.; Schütze, K.; Batistic, O.; Weckermann, K.; Näke, C.; Blazevic, D.; Grefen, C.; Schumacher, K.; Oecking, C.; et al. Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 2004, 40, 428–438. [Google Scholar] [CrossRef]
- Voinnet, O.; Rivas, S.; Mestre, P.; Baulcombe, D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003, 33, 949–956. [Google Scholar] [CrossRef] [Green Version]
- Grondin, A.; Rodrigues, O.; Verdoucq, L.; Merlot, S.; Leonhardt, N.; Maurel, C. Aquaporins contribute to ABA-triggered stomatal closure through OST1-mediated phosphorylation. Plant Cell 2015, 27, 1945–1954. [Google Scholar] [CrossRef] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bissoli, G.; Muñoz-Bertomeu, J.; Bueso, E.; Sayas, E.; Vilcara, E.A.; Felipo, A.; Niñoles, R.; Rubio, L.; Fernández, J.A.; Serrano, R. An Arabidopsis Mutant Over-Expressing Subtilase SBT4.13 Uncovers the Role of Oxidative Stress in the Inhibition of Growth by Intracellular Acidification. Int. J. Mol. Sci. 2020, 21, 1173. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031173
Bissoli G, Muñoz-Bertomeu J, Bueso E, Sayas E, Vilcara EA, Felipo A, Niñoles R, Rubio L, Fernández JA, Serrano R. An Arabidopsis Mutant Over-Expressing Subtilase SBT4.13 Uncovers the Role of Oxidative Stress in the Inhibition of Growth by Intracellular Acidification. International Journal of Molecular Sciences. 2020; 21(3):1173. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031173
Chicago/Turabian StyleBissoli, Gaetano, Jesús Muñoz-Bertomeu, Eduardo Bueso, Enric Sayas, Edgardo A. Vilcara, Amelia Felipo, Regina Niñoles, Lourdes Rubio, José A. Fernández, and Ramón Serrano. 2020. "An Arabidopsis Mutant Over-Expressing Subtilase SBT4.13 Uncovers the Role of Oxidative Stress in the Inhibition of Growth by Intracellular Acidification" International Journal of Molecular Sciences 21, no. 3: 1173. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031173