Overexpression of a Malus baccata NAC Transcription Factor Gene MbNAC25 Increases Cold and Salinity Tolerance in Arabidopsis

Abstract
:1. Introduction
2. Results
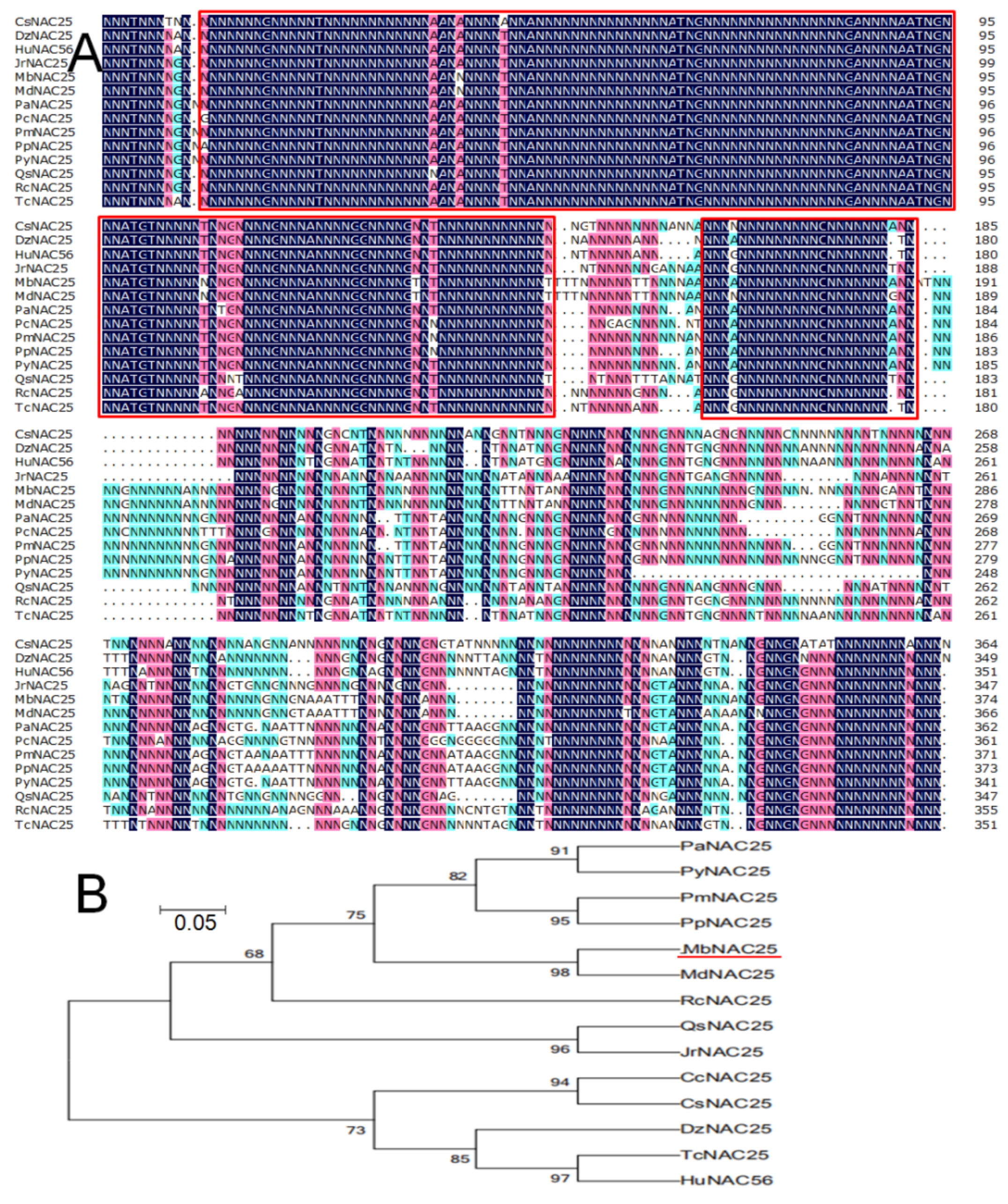
2.1. Isolation and Phylogenetic Relationship of MbNAC25
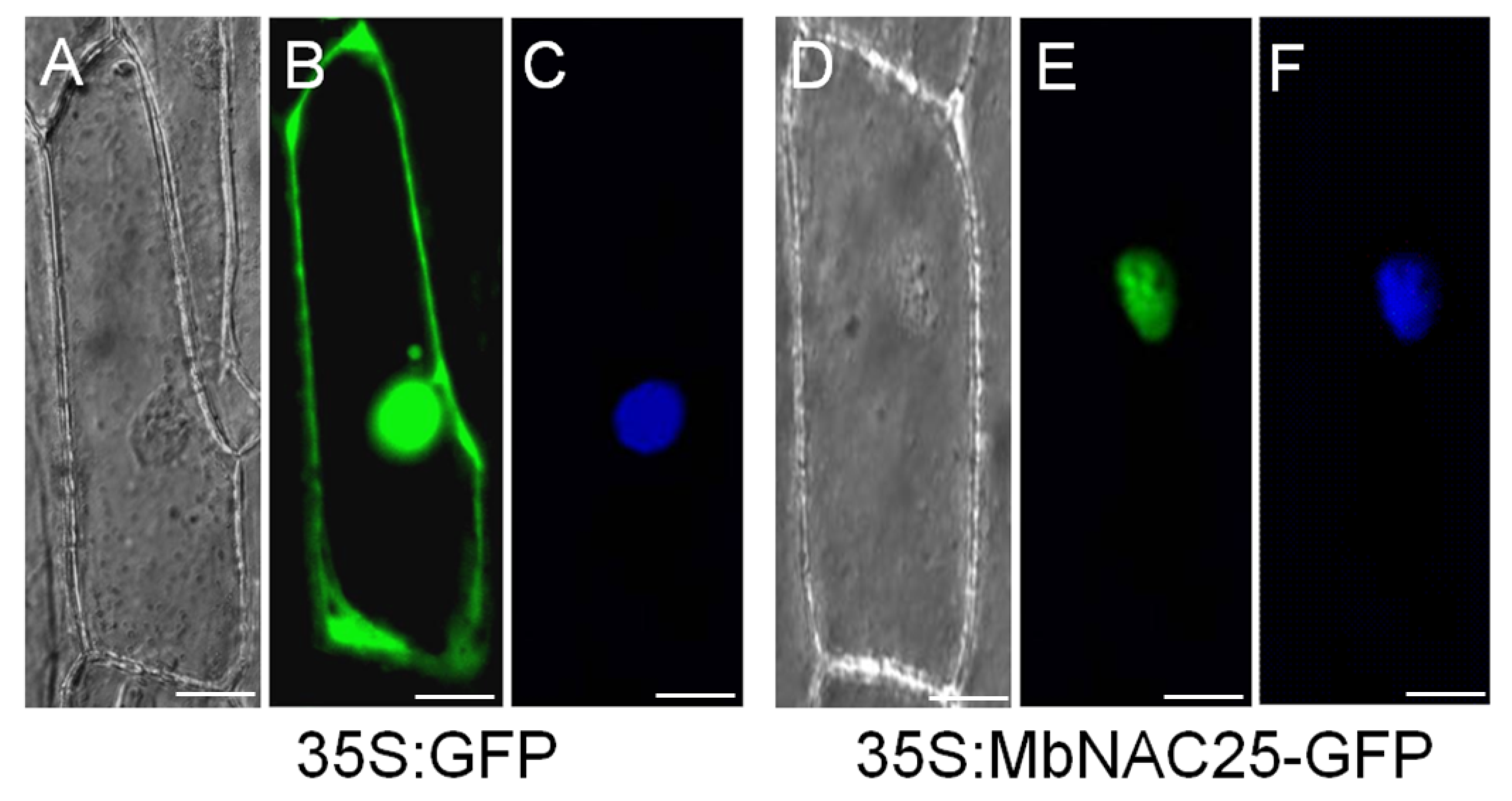
2.2. Subcellular Localization of MbNAC25 Protein
2.3. Expression Analysis of MbNAC25 in M. baccata
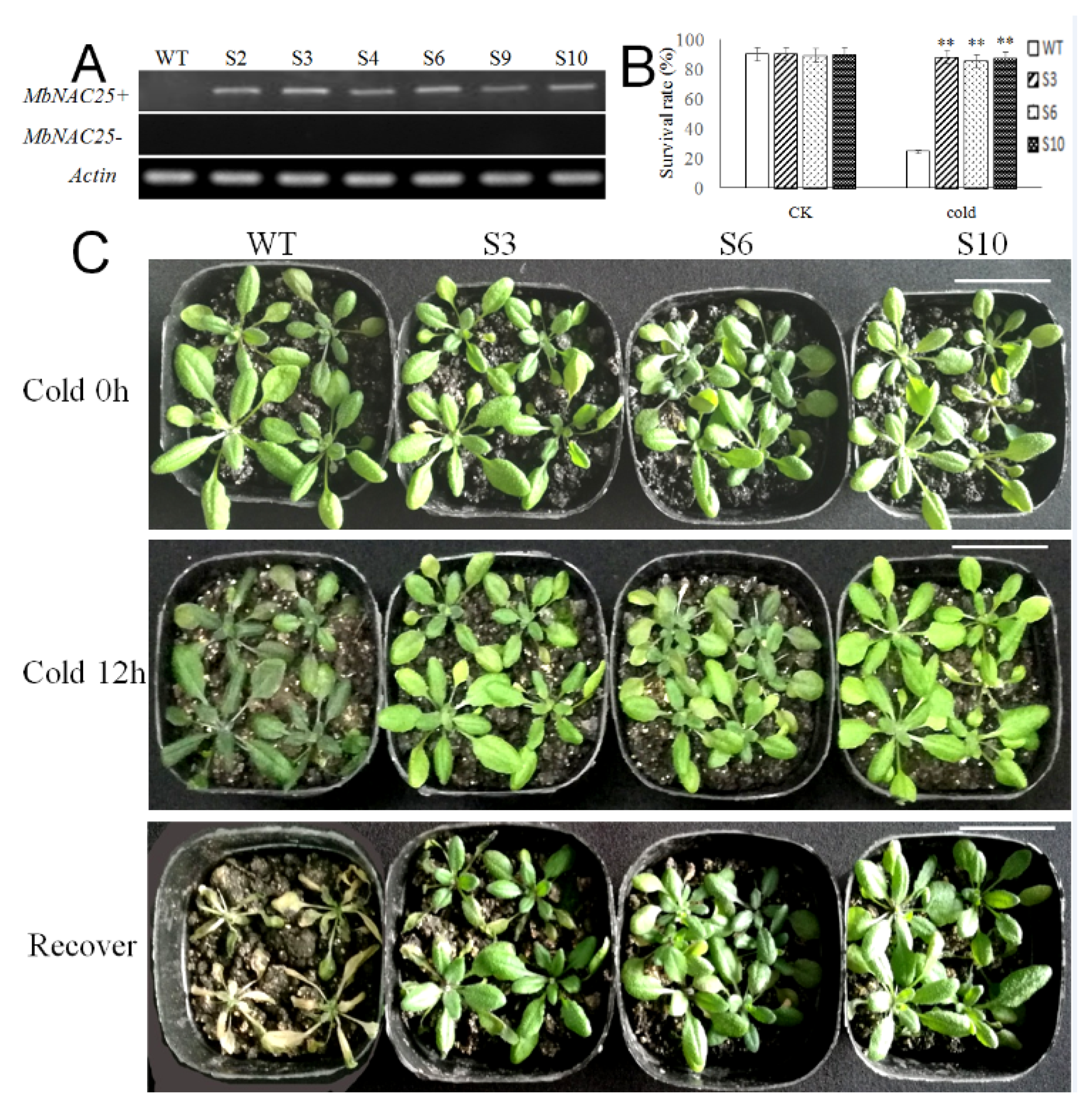
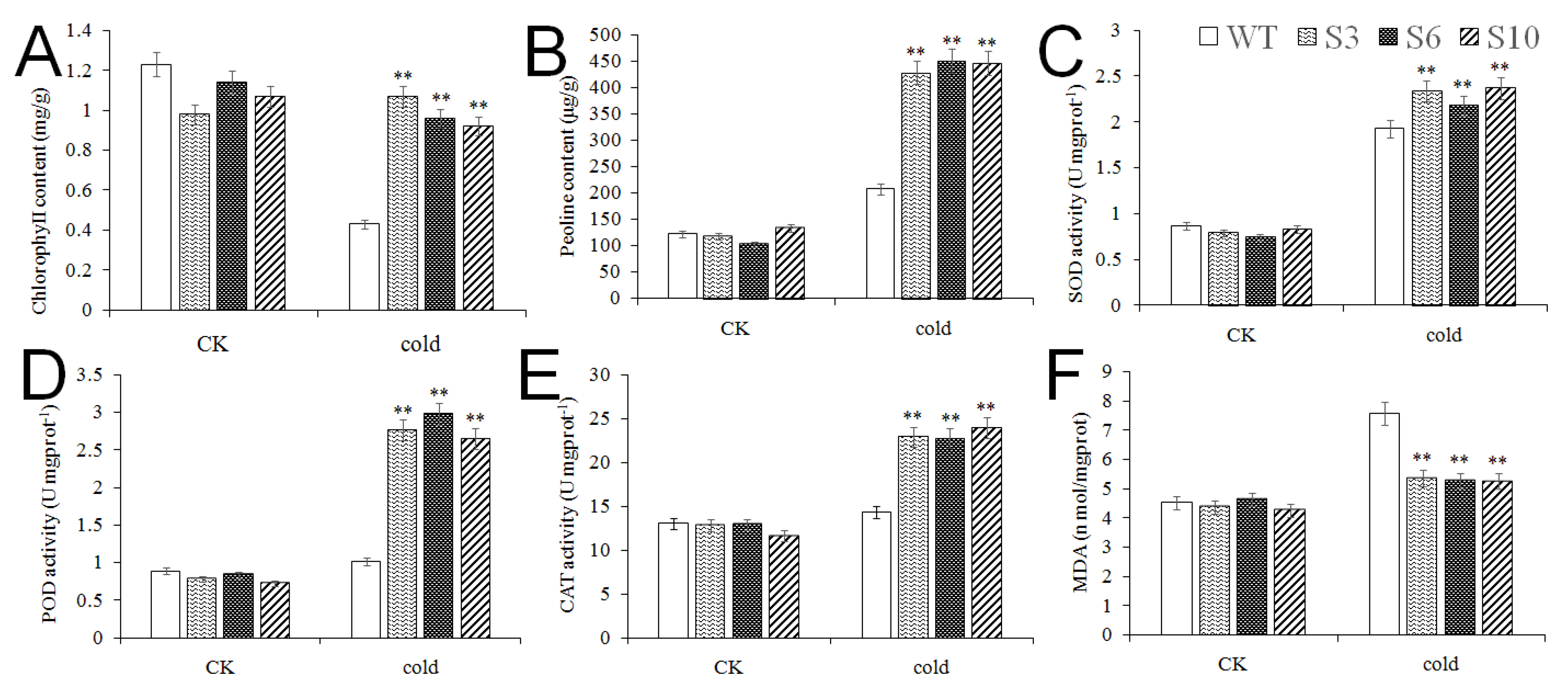
2.4. Overexpression of MbNAC25 in Arabidopsis Enhances Cold Tolerance
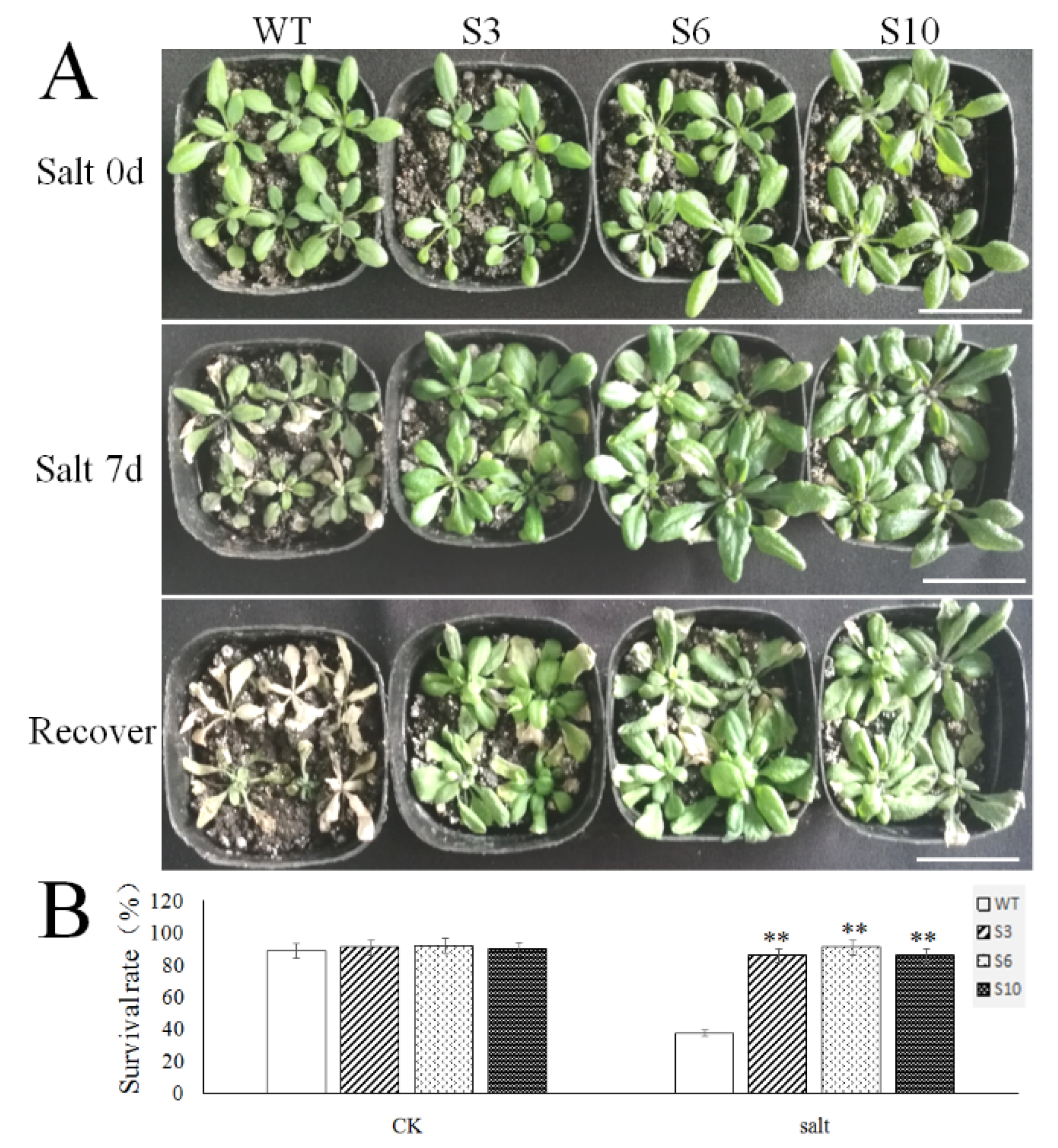
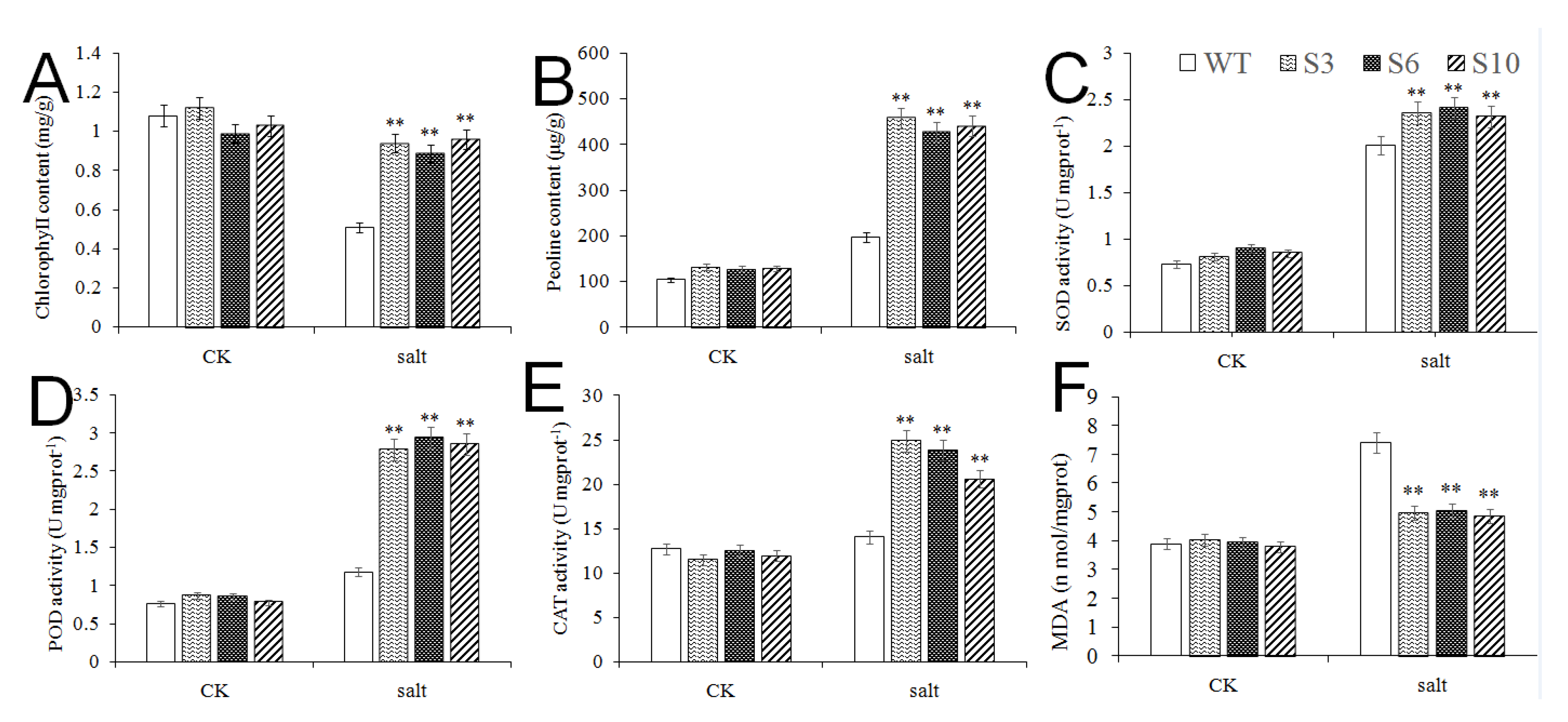
2.5. Overexpression of MbNAC25 in Transgenic Arabidopsis Increased High Salt Tolerance
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Isolation and Cloning of MbNAC25
4.3. Subcellular Localization Analysis of the MbNAC25 Protein
4.4. Quantitative Real-Time PCR Analysis
4.5. Vector Construction and Agrobacterium-Mediated Arabidopsis Transformation
4.6. Determination of Related Physiological Indexes
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor osnap confers abiotic stress response through the aba pathway. Plant Cell Physiol. 2013, 55, 604–619. [Google Scholar] [CrossRef] [Green Version]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.; Zhang, H.; Brannock, J.F.; Chen, X.; Laudeman, T.W.; Timko, M.P. Tobacco transcription factors: Novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef] [Green Version]
- Pereirasantana, A.; Alcaraz, L.D.; Castaño, E.; Sanchezcalderon, L.; Sanchezteyer, F.; Rodriguezzapata, L. Comparative genomics of nac transcriptional factors in angiosperms: Implications for the adaptation and diversification of flowering plants. PLoS ONE 2015, 10, e0141866. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Dugas, D.V.; Bartel, D.P.; Bartel, B. Micro RNA regulation of NAC-domain targets is required for proper formation and separation of adjacent embryonic, vegetative, and floral organs. Curr. Biol. 2014, 14, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.S.; Xie, Q.; Fei, J.F.; Chua, N.H. Micro RNA directs mRNA cleavage of the transcription factor NAC1 to downregulate auxin signals for Arabidopsis lateral root development. Plant Cell 2005, 17, 1376–1386. [Google Scholar] [CrossRef] [Green Version]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical M eristem gene of Petunia is required for pattern form ation in embryos and flowers and is expressed at meristem and prim ordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Gallego-Giraldo, L.; Wang, H.; Zeng, Y.; Ding, S.Y.; Chen, F.; Dixon, R.A. An NAC transcription factor orchestrates multiple features of cell wall development in Medicago truncatula. Plant J. 2001, 63, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Ding, W.; Wu, Y.; Yu, J.; He, X.; Shou, H.; Wu, P. Overexpression of a NAC-domain protein promotes shoot branching in rice. New Phytol. 2007, 176, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Furutani, M.; Vernoux, T.; Traas, J.; Kato, T.; Tasaka, M.; Aida, M. PIN-FORMED1 and PINOID regulate boundary formation and cotyledon development in Arabidopsis embryogenesis. Development 2004, 131, 5021–5030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2014, 14, 3024–3036. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Z.; Baig, M.N.R.; Fan, R.; Ye, J.-L.; Cao, Y.-C.; Deng, X.-X. Identification and expression pattern of a novel NAM, ATAF, and CUC-like gene from Citrus sinensis Osbeck. Plant Mol. Biol. Report. 2009, 27, 292–297. [Google Scholar] [CrossRef]
- Hao, H.Y.; Song, Q.X.; Chen, H.W.; Zou, H.F.; Wei, W.; Kang, X.S.; Ma, B.; Zhang, W.K.; Zhang, J.S.; Chen, S.Y. Plant NAC-type transcription factor proteins contain a NARD domain for repression of transcriptional activation. Planta 2010, 232, 1033–1043. [Google Scholar] [CrossRef]
- Xin, X.; Zhang, Q.; Wu, J. Research of reactive oxygen species in plants and its application on stress tolerance. Biotechnol. Bull. 2013, 36, 6–11. [Google Scholar]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean nac transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Mao, X.; Zhang, H.; Qian, X.; Li, A.; Zhao, G.; Jing, R. TaNAC2, a NAC-type wheat transcription factor conferring enhanced multiple abiotic stress tolerances in arabidopsis. J. Exp. Bot. 2012, 63, 2933–2946. [Google Scholar] [CrossRef]
- Zheng, X.; Chen, B.; Lu, G.; Han, B. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem. Biophys. Res. 2009, 379, 985–989. [Google Scholar] [CrossRef]
- Ding, A.; Li, S.; Li, W.; Hao, Q.; Wan, X.; Wang, K.; Liu, Q.C.; Liu, Q.H.; Jiang, X. RhNAC31, a novel rose NAC transcription factor, enhances tolerance to multiple abiotic stresses in Arabidopsis. Acta Physiol. Plant. 2019, 41, 75. [Google Scholar] [CrossRef]
- Jin, H.; Huang, F.; Cheng, H.; Song, H.; Yu, D. Overexpression of the gmNAC2 gene, an NAC transcription factor, reduces abiotic stress tolerance in tobacco. Plant Mol. Biol. Report. 2013, 31, 435–442. [Google Scholar] [CrossRef]
- Han, D.G.; Ding, H.B.; C, L.J.; Liu, W.; Zhang, Z.Y.; Hou, Y.J.; Yang, G.H. Isolation and characterization of mbwrky1, a wrky transcription factor gene from malus baccata, (L.) borkh involved in drought tolerance. Can. J. Plant Sci. 2018, 98, 1023–1034. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.S.; Van Nguyen, D.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involvedin a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.S.P.; Quach, T.N.; Guttikonda, S.K.; Aldrich, D.L.; Kumar, R.; Neelakandan, A.; Valliyodan, B.; Nguyen, H.T. Molecular characterization of stress-induciblegmnacgenes in soybean. Mol. Genet. Genom. 2009, 281, 647–664. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated nac transcription factor (slnac35) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant. 2016, 158, 45–64. [Google Scholar] [CrossRef]
- Ma, N.N.; Zuo, Y.Q.; Liang, X.Q.; Yin, B.; Wang, G.D.; Meng, Q.W. The multiple stress-responsive transcription factor SlNAC1 improves the chilling tolerance of tomato. Physiol. Plant. 2013, 149, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y. Effects of low temperature on leaf structure and physiological and biochemical characteristics of the Oreorchis patens in Northeast China. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2018; pp. 5–6. [Google Scholar]
- Nguyen, K.H.; Mostofa, M.G.; Li, W.; Ha, C.V.; Watanabe, Y.; Le, D.T.; Thao, N.P.; Tran, L.-S.P. The soybean transcription factor gmnac085 enhances drought tolerance in, arabidopsis. Environ. Exp. Bot. 2018, 151, 12–20. [Google Scholar] [CrossRef]
- Yang, W.; Wang, J.Q.; Liu, Y.; Bai, H.Y.; Wu, Q.; He, Y.F.; Shi, B.A. Research progress on plant salt stress. Hortic. Seedl. 2018, 5, 55–57. [Google Scholar]
- Han, D.G.; Hou, Y.J.; Ding, H.B.; Zhou, Z.Y.; Li, H.X.; Yang, G.H. Isolation and preliminary functional analysis of MbWRKY4 gene involved in salt tolerance in transgenic tobacco. Int. J. Agric. Biol. 2018, 20, 2045–2052. [Google Scholar]
- Zuo, W.; Chao, Q.; Zhang, N.; Ye, J.; Tan, G.; Li, B.; Xing, Y.; Zhang, B.; Liu, H.; Fengler, K.A.; et al. A maize wall-associated kinase confers quantitative resistance to head smut. Nat. Genet. 2015, 47, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Shen, Y.H.; Han, M.J.; Cao, Y.F.; Zhang, Z. An Adaptive Transposable Element Insertion in the Regulatory Region of the EO Gene in the Domesticated Silkworm, Bombyx mori. Mol. Biol. Evol. 2014, 31, 3302–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, X.; Huang, X.; Wang, H.; Huang, L. pTC plasmids from sulfolobus species in the geothermal area of Tengchong, China: Genomic conservation and naturally-occurring variations as a result of transposition by mobile genetic elements. Life 2015, 5, 506–520. [Google Scholar] [CrossRef] [Green Version]
- Han, D.G.; Shi, Y.; Wang, B.; Liu, W.; Yu, Z.Y.; Lv, B.Y.; Yang, G.H. Isolation and preliminary functional analysis of MxCS2: A gene encoding a citrate synthase in Malus xiaojinensis. Plant Mol. Biol. Report. 2015, 33, 133–142. [Google Scholar] [CrossRef]
- Yang, G.H.; Li, J.; Liu, W.; Yu, Z.Y.; Shi, Y.; Lv, B.Y.; Wang, B.; Han, D.G. Molecular cloning and characterization of MxNAS2, a gene encodingnicotianamine synthase in Malus xiaojinensis, with functions intolerance to iron stress and misshapen flower in transgenic tobacco. Sci. Horti. 2015, 183, 77–86. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, J.S.; Zhou, H.L.; Li, J.; Zhang, Z.G.; Wang, D.W.; Chen, S.Y. Serine/threonine kinase activity in the putative histidine kinase-like ethylene receptor NTHK1 from tobacco. Plant J. 2003, 33, 385–393. [Google Scholar] [CrossRef]
- Han, D.G.; Wang, Y.F.; Zhang, Z.Y.; Pu, Q.Q.; Ding, H.B.; Han, J.X.; Fan, T.T.; Bai, X.; Yang, G.H. Isolation and functional analysis of MxCS3: A gene encoding a citrate synthase in Malus xiaojinensis, with functions in tolerance to iron stress and abnormal flower in transgenic Arabidopsis thaliana. Plant Growth Regul. 2017, 82, 479–489. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- An, G.; Watson, B.D.; Chiang, C.C. Transformation of tobacco, tomato, potato, and Arabidopsis thaliana using a binary ti vector system. Plant Physiol. 1986, 81, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.X.; Liu, S.Y.; Liu, Y.F.; Xu, J.; Liu, T.; Dong, S.Z. Effectiveness of lysozyme coatings and 1-MCP treatments on storage and preservation of kiwifruit. Food Chem. 2019, 288, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.H.; Qian, X.C.; Jiang, T.J.; Zheng, X.L. Effect of eugenol fumigation treatment on chilling injury and CBF gene expression in eggplant fruit during cold storage. Food Chem. 2019, 292, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Yuan, H.W.; Kumar, V.; Ramakrishnan, M.; Kohli, S.K.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Zheng, B.S. Castasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Li, L.; Lin, G.J.; Gu, H.Z.; Yu, L.; Ni, C.W. Effects of Srxn1 on growth and Notch signalling of astrocyte induced by hydrogen peroxide. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1917–1923. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Yang, H.; Wu, M.Y.; Yi, H.L. Enhanced drought tolerance of foxtail millet seedlings by sulfur dioxide fumigation. Ecotoxicol. Environ. Saf. 2019, 178, 9–16. [Google Scholar] [CrossRef]
- Jiang, Y.; Lin, L.L.; Zhong, S.; Cai, Y.J.; Zhang, F.; Wang, X.B.; Miao, R.R.; Zhang, B.D.; Gao, S.M.; Hu, X.Q. Overexpression of novel lncRNA NLIPMT inhibits metastasis by reducing phosphorylated glycogen synthase kinase 3 in breast cancer. J. Cell. Physiol. 2019, 234, 10689–10708. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′→3′) | Purpose |
|---|---|---|
| MbNAC25-F | ATGGAGAGCACAGATTCATC | full-length cDNA of MbNAC25 |
| MbNAC25-R | CTATGAATTCCAGTTCATGCTT | full-length cDNA of MbNAC25 |
| MbNAC25-qF | CAATGCGCAAAGGCCTACGA | qPCR |
| MbNAC25-qR | AGCAGACCCTATCGATCCCA | qPCR |
| Actin-F | ACACGGGGAGGTAGTGACAA | qPCR |
| Actin-R | CCTCCAATGGATCCTCGTTAT | qPCR |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, D.; Du, M.; Zhou, Z.; Wang, S.; Li, T.; Han, J.; Xu, T.; Yang, G. Overexpression of a Malus baccata NAC Transcription Factor Gene MbNAC25 Increases Cold and Salinity Tolerance in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1198. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041198
Han D, Du M, Zhou Z, Wang S, Li T, Han J, Xu T, Yang G. Overexpression of a Malus baccata NAC Transcription Factor Gene MbNAC25 Increases Cold and Salinity Tolerance in Arabidopsis. International Journal of Molecular Sciences. 2020; 21(4):1198. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041198
Chicago/Turabian StyleHan, Deguo, Man Du, Zhengyi Zhou, Shuang Wang, Tiemei Li, Jiaxin Han, Tianlong Xu, and Guohui Yang. 2020. "Overexpression of a Malus baccata NAC Transcription Factor Gene MbNAC25 Increases Cold and Salinity Tolerance in Arabidopsis" International Journal of Molecular Sciences 21, no. 4: 1198. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041198