Interleukin-17A and Keratinocytes in Psoriasis


1
Department of Dermatology, Graduate School of Medical Sciences, Kyushu University, Maidashi 3-1-1, Higashiku, Fukuoka 812-8582, Japan
2
Research and Clinical Center for Yusho and Dioxin, Kyushu University, Maidashi 3-1-1, Higashiku, Fukuoka 812-8582, Japan
3
Division of Skin Surface Sensing, Graduate School of Medical Sciences, Kyushu University, Maidashi 3-1-1, Higashiku, Fukuoka 812-8582, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(4), 1275; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041275
Submission received: 22 January 2020
/
Revised: 11 February 2020
/
Accepted: 11 February 2020
/
Published: 13 February 2020
(This article belongs to the Special Issue Therapy and Prevention of Atopic Dermatitis and Psoriasis)
{kind=link}
{kind=link}
Abstract
:The excellent clinical efficacy of anti-interleukin 17A (IL-17A) biologics on psoriasis indicates a crucial pathogenic role of IL-17A in this autoinflammatory skin disease. IL-17A accelerates the proliferation of epidermal keratinocytes. Keratinocytes produce a myriad of antimicrobial peptides and chemokines, such as CXCL1, CXCL2, CXCL8, and CCL20. Antimicrobial peptides enhance skin inflammation. IL-17A is capable of upregulating the production of these chemokines and antimicrobial peptides in keratinocytes. CXCL1, CXCL2, and CXCL8 recruit neutrophils and CCL20 chemoattracts IL-17A-producing CCR6+ immune cells, which further contributes to forming an IL-17A-rich milieu. This feed-forward pathogenic process results in characteristic histopathological features, such as epidermal hyperproliferation, intraepidermal neutrophilic microabscess, and dermal CCR6+ cell infiltration. In this review, we focus on IL-17A and keratinocyte interaction regarding psoriasis pathogenesis.
Keywords:
psoriasis; keratinocytes; interleukin-17A; Th17; ILC3; CCL20; CXCL1; CXCL8; Koebner phenomenon; antimicrobial peptides1. Introduction
Psoriasis is an immune-mediated chronic skin disease characterized by epidermal hyperproliferation, an intraepidermal accumulation of neutrophils, and dermal inflammatory cell infiltrates that are composed of dendritic cells and T cells [1]. It has an estimated global prevalence of 2–4% [1,2]. Males are twice as likely as females to be affected [3,4]. As desquamative erythema can affect any skin site, psoriasis profoundly impairs the patients’ quality of life, treatment satisfaction and adherence, and socioeconomic stability [5,6,7]. The skin lesion usually appears on the sites with frequent trauma such as elbows and knees [8,9]. This injury-induced development of psoriasis is called Koebner phenomenon [8,9]. Environmental factors, such as smoking, also trigger or exacerbate psoriatic lesions [10,11]. Genetic factors are critically involved in the development of psoriasis [12,13]. In addition to skin eruption, approximately 30% of patients with psoriasis manifest psoriatic arthritis [14,15,16,17,18,19]. Psoriasis is also significantly comorbid with other autoimmune diseases, such as bullous pemphigoid [20,21,22,23,24]. Psoriasis is frequently associated with cardiovascular diseases, metabolic diseases, and renal disorders [25,26,27,28,29,30,31,32,33,34,35,36]. Cancer risk is slightly higher in patients with psoriasis [37]. The topical application of steroids and vitamin D3 analogues inhibits psoriatic inflammation and normalizes epidermal differentiation [38,39]. Systemic treatments, such as methotrexate, cyclosporine, phototherapy, and the phosphodiesterase 4 inhibitor apremilast, are useful for patients with extensive lesions [40,41,42,43,44,45].
That the tumor necrosis factor-α (TNF-α) and IL-23/IL-17A axes appear to be major drivers in the pathogenesis of psoriasis is underscored by the excellent response of psoriasis to biologics targeting TNF-α, IL-23, and IL-17A, although a difference exists in their efficacy [46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61]. Anti-TNF-α/IL-23/IL-17A biologics successfully improve psoriatic arthritis [18,56,62,63,64]. Reductions in comorbid cardiovascular events and systemic inflammation have been reported in patients with psoriasis treated with anti-TNF/IL23/IL17 biologics [65,66].
As the clinical response to anti-IL-23/IL-17A biologics seems better than that to anti-TNF-α biologics in psoriasis, the IL-23/IL-17A axis likely plays a more crucial role than the TNF-α axis in the development of psoriasis [67,68,69,70,71]. Under the regulation of IL-23p19, IL-17A-producing CD4+ helper T (Th17) cells create a self-amplifying, feed-forward inflammatory response that is markedly augmented in the presence of TNF-α [68,72,73]. In addition, Th17 cells produce high amounts of TNF-α [74,75]. Therefore, IL-17A inhibition by the anti-IL-17A biologic results in early clinical, histopathologic, and molecular resolution of psoriasis [69].
In addition, various murine psoriasis models stress a pivotal role of the IL-23/IL-17A axis in experimental psoriasis [76,77,78,79,80]. Multiple animal studies have indicated that the interaction between IL-17A and keratinocyte is the key issue in the development of psoriasis [76,77,78,79]. In parallel, the importance of IL-17A and keratinocyte interaction is reinforced in patients with psoriasis who are successfully treated with the anti-IL-17A biologic [69]. In this review, we will focus on the multifaceted biological response in keratinocytes stimulated by IL-17A with regard to psoriatic pathogenesis.
2. IL-17A Signaling System
The IL-17 family plays a critical role in the immune response in infectious, inflammatory, autoimmune, and neoplastic disorders [81,82]. The IL-17 family and its receptors are evolutionally ancient, and they are present in species as early as lamprey and sea urchins [83,84]. IL-17 family members comprise IL-17A, IL-17B, IL-17C, IL-17D, IL-17E, IL-17F, and IL-17AF [81,82]. IL-17AF is a hybrid heterodimer of IL-17A and IL-17F. IL-17E is also called IL-25 and is related to the type 2 immune response and allergies [81,82,85]. Among IL-17 family members, IL-17A has been the most strongly implicated in human health and disease. IL-17A is produced from hematopoietic cells, including Th17, CD8+ cytotoxic T cell (Tc17), γδ T cell, natural killer cell, group 3 innate lymphoid cell (ILC3), and “natural” Th17 cell [86,87,88,89], but IL-17B, IL-17C, and IL-25 are preferentially produced from nonhematopoietic cells, including keratinocytes [81,82,90]. Both hematopoietic cells and keratinocytes produce IL-17F [81,82,90]. Keratinocyte-derived IL-17C is capable of stimulating Th17 cells to secrete more IL-17A [91].
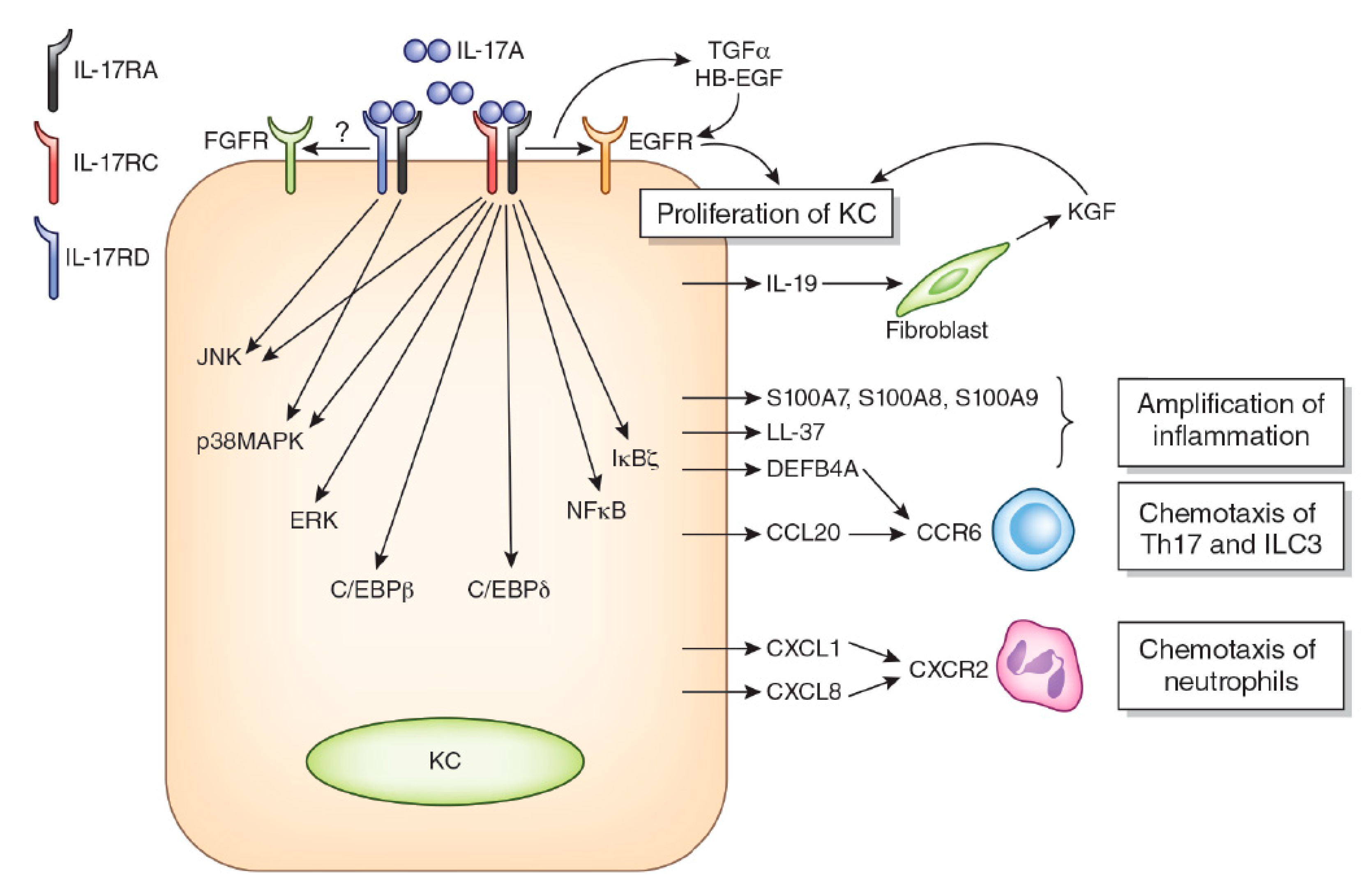
The IL-17 receptor (IL-17R) is composed of five members: IL-17RA, IL-17RB, IL-17RC, IL-17RD, and IL-17RE [81,82,90]. IL-17A, IL-17F, and IL-17AF all share the same IL-17R comprising IL-17RA and IL-17RC heterodimers [81,82,90]. IL-17C binds to IL-17RA and IL-17RE heterodimers [92]. IL-25 ligates IL-17RA and IL-17RB [81,82,90]. A recent study showed that IL-17A also activates IL-17RA and IL-17RD heterodimers [93] (Figure 1). IL-17B binds IL-17RB, but another heterodimeric IL-17R member remains unidentified [82]. Keratinocytes express both IL-17RA/IL-17RC and IL-17RA/IL-17RD, and the binding of IL-17A induces the transcription of differential gene sets [93]. Initial subcellular events in the ligation of IL-17RA/RC by IL-17A are the recruitment and activation of ACT1 encoded by the TRAF3IP2 gene, TRAF6 and CARMA2 complexes, and the downstream activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and MAPKs [93,94,95,96,97]. The ligation of IL-17RA/IL-17RC by IL-17A induces the activation of NF-κB, ERK, p38 MAPK, and JNK, while that of IL-17RA/IL-17-RD mainly activates p38 MAPK and JNK and barely affects NF-κB and ERK [93]. In addition, IL-17RA physically and functionally interacts with and transactivates epidermal growth factor (EGFR) [98]. IL-17RD potentially interacts with and transactivates fibroblast growth factor 2 receptor [82,99].
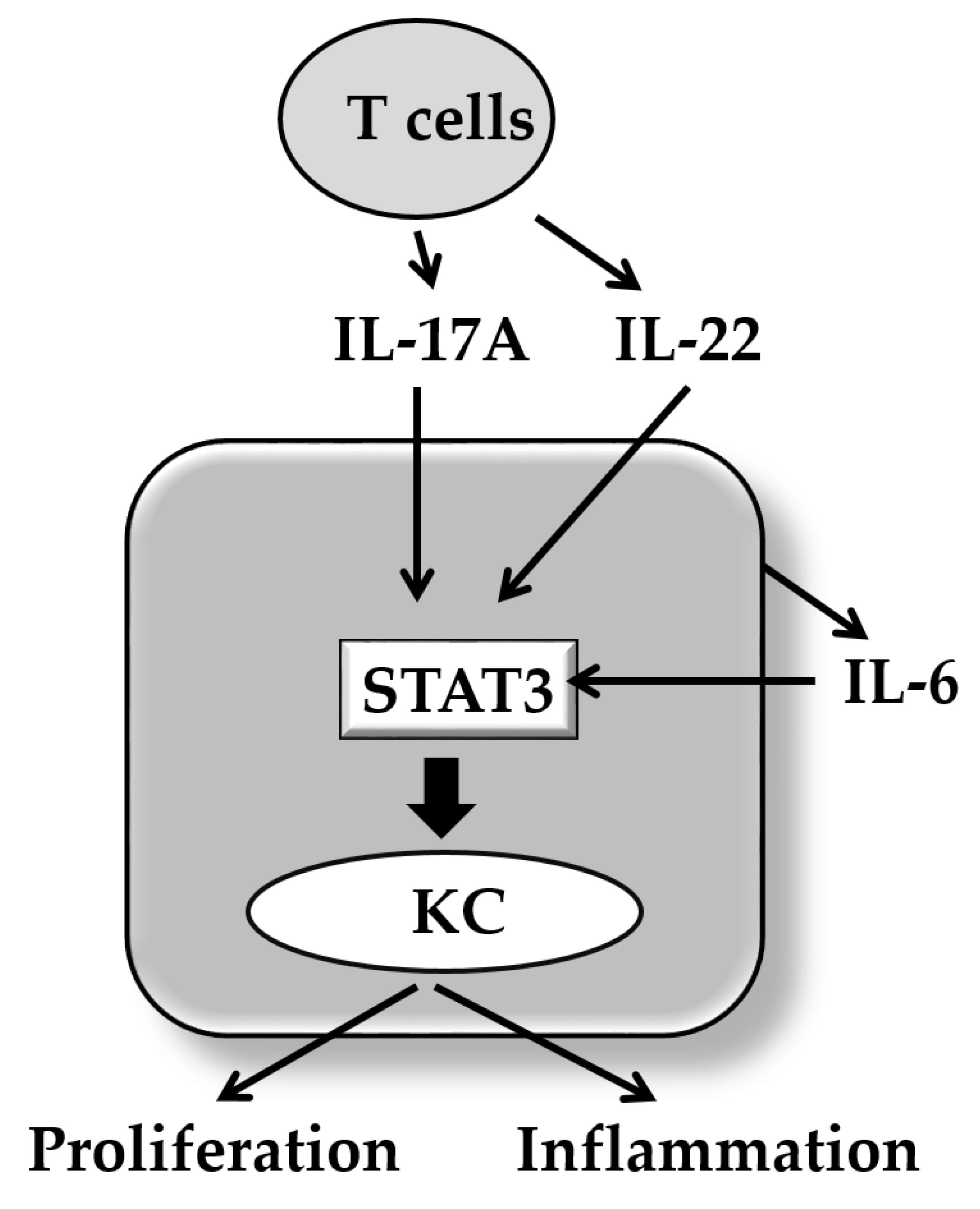
In addition to the above-mentioned signaling cascades, IL-17A activates various other signal molecules including signal transducer and activator of transcription 3 (STAT3) in keratinocytes [100]. STAT3 is a very crucial signaling molecule in the development of psoriasis because transgenic mice with keratinocytes expressing a constitutively active Stat3 (K5.Stat3C mice) develop a skin phenotype either spontaneously, or in response to wounding, that closely resembles psoriasis [101]. Moreover, a STAT3 inhibitor STA-21 inhibits the generation of skin lesion in these psoriatic mice [102]. IL-17A is known to activate STAT3 via receptor-interacting protein 4 (RIP4) activation and upregulates the CCL20 expression [103]. IL-17A also upregulates keratin 17 expression via STAT1 and STAT3 activation [104]. IL-6 and IL-22 also play a synergistic role in development of psoriasis with IL-17A [68]. Notably, both IL-6 and IL-22 are potent STAT3 activators [105]. In accordance, biological or natural molecules such as indirubin and its derivatives useful for inactivating STAT3 exhibit therapeutic potential for psoriasis [106] (Figure 2). It reveals that IL-17 and IL-22 promote keratinocyte stemness and potentiate its regeneration [107]. IL-6 is produced from keratinocytes in response to IL-17A [108]. IL-22 is produced from Th17/22 cells, Th22 cells, and other immune cells [109,110].
In humans, impairment of the IL-17 signal causes infectious diseases, especially by Candida albicans, which is a ubiquitous fungus and commensal yeast of the intestines and skin [96]. Notably, deficiency of the IL17RA, IL17RC, IL17F, or TRAF3IP2 genes is implicated in chronic mucocutaneous candidiasis disease (CMCD), which is characterized by recurrent or persistent infection affecting the nails, skin, and oral and genital mucosae caused by the Candida species, often C. albicans [96,111,112,113]. Impairment of the IL-17 signal is evident in other immunocompromised inborn errors, including autosomal-dominant hyper IgE syndrome, autosomal dominant STAT1 gain-of-function, autosomal-recessive autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED), autosomal-recessive CARD9 deficiency, IL12RB1 deficiency, IL12B deficiency, and RORC deficiency [96]. However, these inborn errors seem to exhibit more complicated immune defects beyond IL-17 dysfunction and manifest CMCD together with other types of infection, including Mycobacterium, Staphylococcus, and viral disorders [96]. Of note, mice lacking Traf3ip2 (Act1) or Il17ra manifest similar clinical phenotype as human CMCD patients lacking IL17RA or IL17RC [114,115,116,117]. A recent murine study by Sparber et al. also indicates that Malassezia infection selectively triggers the IL-17A-induced immune response [118]. These findings indicate a crucial role of IL-17A in anti-fungal immunity in humans and mice.
Mice overexpressing IL-17A in keratinocytes (K14-IL-17Aind/+ mice) exhibit severe psoriasiform skin inflammation and vascular dysfunction in conjunction with infiltration of the vasculature by inflammatory myeloid cells [108]. The K14-IL-17Aind/+ mice acquire the highest local and systemic IL-17A levels and exhibit a particularly severe psoriasis-like skin phenotype [108]. Homozygous CD11c-IL-17Aind/ind mice and heterozygous CD11c-IL-17A ind/+ mice show a delayed onset of moderate to severe psoriasis-like skin disease associated with reduced amounts of cutaneous IL-17A compared with K14-IL-17Aind/+ mice. In agreement with elevated skin and a stepwise increase in systemic IL-17A [119], homozygous CD11c-IL-17A ind/ind mice develop earlier and more severe skin lesions, as well as more pronounced vascular inflammation and dysfunction than heterozygous CD11c-IL-17A ind/+ mice [76,119]. These experimental models indicate that IL-17A induces psoriasis-like lesions in a dose-dependent manner, irrespective of its cellular source [76,119].
3. IL-17A and Psoriasis
In the early 2000s, IL-23 was found to induce the production of IL-17A by activated CD4+ T cells, which were later named Th17 cells [74,120]. These cells express RORγt (RORC) as the master transcription factor [121] and promoted the anti-infectious defense in the mucosa and skin [96], whereas excessive exposure to IL-23 induces their transformation into autoimmune or autoinflammatory immune cells [122,123]. Accordingly, the intradermal injection of IL-23 induces murine psoriasis-like dermatitis with epidermal acanthosis, neutrophil recruitment, and the infiltration of IL-17-producing T cells [78,79,124].
Psoriasis is one of the most typical IL-23/IL-17A-driven human diseases [68,72,73]. Th17 cells, Tc17 cells, IL-17-producing γδ T cells, and ILC3 can all be found in psoriatic skin [89,125,126]. IL-23 is composed of IL-23p40 and IL-23p19 subunits, and the expression of both is upregulated in the lesional skin compared to the nonlesional skin in psoriasis [127]. Dermal dendritic cells and monocytes are major sources of IL-23 production [128]. However, immunohistological staining also proves that IL23p40 and IL23p19 are expressed in normal and psoriatic keratinocytes [127,128,129]. Moreover, the epidermal expression of IL-23p40 and IL-23p19 is stronger in psoriasis lesions than in healthy controls [127,128,129]. Murine keratinocytes also produce biologically active IL-23 [130]. IL-23 can promote keratinocyte growth and histological acanthosis via the JAK2/AKT/STAT3/LMO4 signaling pathway [131]. LIM-domain only protein 4 (LMO4) is a LIM-domain protein that regulates keratinocyte proliferation and differentiation during embryogenesis [131]. In addition, TNF-α is an active stimulator of IL-23 production from keratinocytes in mice and possibly in humans [131]. In parallel, the anti-IL-23p40 antibody (Ustekinumab) [132,133] and anti-IL-23p19 antibodies (guselkumab, risankizumab, and tildrakizumab) [133,134,135] exhibit a high efficacy to psoriasis with a superiority of risankizumab over ustekinumab [133], and guselkumab over the anti-TNF-α antibody adalimumab [134].
The expression of IL17A, IL17F, and IL17C is upregulated in psoriatic lesional skin [136,137,138]. The intradermal injection of IL17C induces epidermal thickening and neutrophil infiltration [139]. As significant efficacy of the anti-IL17C antibody has recently been noted in murine psoriasis and atopic dermatitis [140], its human use has been suggested [141]. However, IL-17A is believed to be the most pathogenic IL-17 family member in psoriasis [68,69,72,73,142]. The lesional skin of psoriasis harbors IL-17A-producing immune cells [89,143], and transcriptomic analysis indicates a clear predominance of IL-17A signals [69,142]. Subsets of IL-17A-producing T cells are reactive to autoantigens, such as LL-37 (cathelicidin), a disintegrin and metalloprotease domain containing thrombospondin type 1 motif-like 5 (ADAMTSL5), keratin 17, and neolipid antigens generated by phospholipase A2 group IV D (PLA2G4D) [20,125,126,144,145,146,147]. IL-17A-dominant immune activation is also detected in the nonlesional skin of psoriasis [148].
There are three commercially available anti-IL17A biologics: secukinumab (fully human IgG1κ anti-IL17A antibody), ixekizumab (fully human IgG4 anti-IL17A antibody), and brodalumab (fully human IgG2 anti-IL17RA antibody). The severity of psoriatic skin lesions is generally evaluated by the Psoriasis Area and Severity Index (PASI) [149]. The efficacy of a therapeutic agent is assessed by the rate of responders who achieve 75%, 90%, and 100% reduction of PASI, namely, PASI75, PASI90, and PASI100, respectively. All three anti-IL-17A biologics exbibit remarkable therapeutic efficacy for psoriasis. The PASI90 scores of secukinumab, ixekizumab, and brodalumab are reported to be as high as 60% to 70% at 12 weeks post treatment [150,151,152].
Regarding histology, the psoriatic epidermis is acanthotic and the proliferating capacity of keratinocyte is increased as determined by Ki67 positivity and cytokeratin 16 expression [68,153,154]. In accordance with the accumulation of neutrophils in psoriasis, the gene expression of neutrophilic cytochemokines CXCL1, CXCL2, CXCL8, and IL-36 is upregulated in the lesional skin of psoriasis [68,155,156,157]. Psoriatic keratinocytes produce a large amount of CCL20 [158,159], which is a potent chemoattractant for CCR6+ IL-17A-producing Th17 [89,143,160], ILC3 [161,162], Tc17 [163,164], and γδ T cells [78,165]. Interestingly, IL-17A actively upregulates the production of these key cytochemokines as well as proliferative capacity in keratinocytes.
4. IL-17A and Keratinocyte Proliferation/Differentiation
Psoriatic keratinocytes simultaneously exhibit increased proliferation (Ki67+ or cytokeratin 16+ cell number) and differentiation [involucrin (IVL)+ cell number] [69,166,167,168]. Accordingly, IL17A is an active promoter of keratinocyte proliferation and differentiation [168,169,170,171]. In psoriasis patients, the neutralization of IL-17A by ixekizumab [168] or secukinumab [69] significantly decreases histological acanthosis, the number of Ki-67+-proliferating keratinocytes, and epidermal cytokeratin 16 expression. In the murine imiquimod-induced psoriasis model, topical imiquimod induces epidermal thickening that is significantly alleviated in Traf3ip2 (Act1)-deficient mice in which IL-17 signaling is blocked [170]. The molecular mechanisms of the IL-17A-mediated acceleration of keratinocyte proliferation and differentiation are not yet fully understood.
IL-17A alone is likely to be insufficient to evoke a significant inflammatory response and may cooperatively or synergistically accelerate the proinflammatory cascade in combination with other cytokines, such as TNF-α, IL-23, IL-1β, IL-6, IL-22 and transforming growth factor-β (TGF-β) [68,172]. Transcriptomic analysis reveals a clear additive or synergistic gene regulation by IL-17A and TNF-α in human keratinocytes [173]. This gene regulation is likely attributable to two sets of transcription factors: NF-κB and the C/CAAT-enhancer-binding proteins (C/EBP), C/EBPβ, or C/EBPδ [169,173,174]. TNF-α is a strong inducer of active NF-κB, while IL-17A activates C/EBPβ or C/EBPδ and to a lesser extent NF-κB [169,173,174]. Therefore, the IL-17A blockade reduces the expression of these responsive genes to a greater extent than TNF-α inhibition [168].
The C/EBP family members are highly expressed transcription factors in epidermal keratinocytes and sebocytes in mice and humans and accelerate their differentiation [175]. The expression of C/EBPβ protein is located in the upper spinous layer and is strongly upregulated in the lesional skin of psoriasis [169]. Together with the elevated gene expression of CEBPB, the expression of keratinocyte-terminal differentiation genes, such as IVL, FLG2, and TGM1, is upregulated in the lesional skin of psoriasis [169]. In the promoter region of the IVL gene, there is a binding site for C/EBP, and the C/EBP transcription factor is necessary for the appropriate and continuous production of IVL protein [176]. These results suggest that IL-17A accelerates keratinocyte differentiation by increasing C/EBPβ protein in keratinocytes [168,169,173].
Regarding keratinocyte proliferation, IL-17A stimulates keratinocytes to produce IL-19 [173,177]. The combination of IL-17 and TNF-α results in the synergistic expression of IL-19 in keratinocytes. IL-17 alone promotes IL-19 expression by approximately 1.79-fold, and TNF-α alone slightly reduces IL-19 expression, whereas a combination of IL-17 and TNF-α promotes expression by 54.6-fold [173,177]. IL-19 itself promotes keratinocyte migration but not proliferation [178]. However, it activates fibroblasts to produce keratinocyte growth factor, which upregulates keratinocyte proliferation [178]. Additionally, the upregulation of cell cycle-related genes, such as CCNE1, CDCA5, and CDCA25A, suggests a direct contribution of IL-17 to epidermal KC proliferation [169]. The expression of cytokeratin 16 is upregulated in the lesional epidermis of psoriasis patients and in keratinocytes incubated with IL-17A [69,168,179]. The expression of cytokeratin 16 increases the proliferative capacity of keratinocytes via EGFR phosphorylation [180].
Other studies also underpin a possibility that IL-17A activates EGFR [98,181,182]. Chen et al. have demonstrated that IL-17A transactivates EGFR in keratinocyte stem cells [98]. IL-17A accelerates the enzymatic cleavage of amphiregulin, which activates EGFR in airway epithelial cells [181]. In the lesional skin of psoriasis, the expression of EGFR ligands, such as heparin-binding EGF, transforming growth factor-α, and amphiregulin, is overexpressed [183]. Consistent with these notions, the inhibition of EGFR by erlotinib or cetuximab successfully improves severe psoriasis [184,185,186,187]. These studies suggest that EGFR activation may partly explain the IL-17A-mediated upregulation of keratinocyte proliferation.
5. IL-17A and Cyto/Chemokines in Keratinocytes
In keratinocytes, IL-17A upregulates the expression of various psoriasis-related cyto/chemokines and antimicrobial peptides, such as CXCL1, CXCL8, IL-36G, CCL20, IL-19, IL-17C, S100A7, S100A8, S100A9, LL-37, and defensin β 4A (DEFB4A) [157,169,173,188,189,190]. The expression of these molecules is upregulated in the lesional skin of psoriasis and is downregulated to normal levels by biologics targeting TNF-α [128] or IL-17A [69,168]. Some are upregulated even in the nonlesional skin of patients with psoriasis [148]. The IL-17A-induced upregulation of these molecules is further amplified in the presence of TNF-α and IL-19 [173,177].
CXCL1 and CXCL8 are potent chemoattractants for neutrophils [191,192,193]. IL-36G induces CXCL1 and CXCL8 expression in an autocrine fashion in keratinocytes and recruits neutrophils [194]. Full-length IL-36G is cleaved by cathepsin G released from infiltrated neutrophils, and the cleaved IL-36G exhibits a more potent functional activity than the full-length one [192,195,196]. Moreover, IL-17A upregulates IL-36G production more potently in human psoriasis-derived keratinocytes than in healthy keratinocytes [197]. Therefore, the IL-17A, IL-36G, CXCL1/CXCL8, and neutrophils form a feed-forward vicious cycle, and an intraepidermal neutrophilic microabscess (Munro’s microabscess or Kogoj’s spongiform pustule) may develop [156,198,199]. The pathogenic significance of IL-36 is stressed more in pustular rather than plaque psoriasis [156,198,199]. In parallel, inhibition of the IL-36 pathway is efficacious for the treatment of pustular psoriasis [200].
While most chemokines redundantly bind to multiple receptors, CCL20 has only one known receptor, CCR6 [191]. CCR6 is expressed on dendritic cells [159] and IL-17-producing immune cells, including Th17 [89,143,201], ILC3 [161,162], Tc17 [163,164], and γδ T cells [78,165]. CCR6 is now considered a representative marker for Th17 cells [202,203]. The human psoriatic epidermis expresses abundant CCL20 with the dermal infiltration of CCR6+ dendritic cells and skin-homing T cells [143,158,159]. Although dermal dendritic cells and T cells express CCL20 in the psoriatic dermis [159], the expression of CCL20 is largely confined to the psoriatic epidermis, suggesting that epidermal keratinocytes are the major source of CCL20 production in psoriatic lesions [158,159]. CCL20 is constitutively expressed in cultured keratinocytes [201,204], and its production is upregulated by TNF-α and IL-17A [182,204]. Scratch injury upregulates the gene expression of CCL20, CXCL8, and IL36G, which may be related to the scratch-induced Koebner phenomenon frequently observed in patients with psoriasis [201]. Preclinical studies have revealed that the humanized anti-CCR6 antibody efficiently inhibits the cutaneous infiltration of CCR6+ T cells in human and murine models [205,206]. The development of small molecule inhibitors against CCR6 is also ongoing [207,208]. Targeting the CCL20/CCR6 axis may be a potential therapeutic strategy for the treatment of psoriasis.
6. IL-17A and Anti-Microbial Peptides in Keratinocytes
S100A7, S100A8, S100A9, LL-37, and DEFB4A are anti-microbial peptides, and their expression is upregulated by IL-17A in keratinocytes [189,209,210]. S100A7 (psoriasin) has a multifaceted role in keratinocyte pathophysiology, including wound healing, keratinocyte differentiation, nucleocytoplasmic transport, and chemotaxis for CD4+ T cells, neutrophils, and monocytes [211]. The production of S100A7 is augmented in the presence of IL-19 [212], IL-36G [188], and TNF-α [213]. S100A8, S100A9, and S100A12 are alternatively known as calgranulin A, B, and C, respectively, and exhibit antimicrobial activity [214]. All are elevated in psoriatic plaque and decreased by treatment with anti-IL-17A antibody secukinumab [69,168,215]. The S100 family members primarily form homodimers or higher-order oligomers, but S100A8 and S100A9 uniquely form heterodimeric complexes, which are known as calprotectin [214]. The chelation of Zn2+ and Mn2+ by extracellular S100A8/A9 is a proposed mechanism of antimicrobial activity [214]. S100A12 also has a chemotactic activity for mast cells and monocytes [216]. Pure human S100A8/A9 has broad spectrum antimicrobial activities against microorganisms, including Capnocytophaga sputigena, C. albicans, Escherichia coli, Staphylococcus aureus, S. epidermis, Borrelia burgdorferi, and Listeria monocytogenes, in vitro, and S100A12 has antimicrobial activity against filarial parasites [214]. The receptor for advanced glycation end-products (RAGE) may serve as a common receptor for S100 proteins, including S100A7, S100A8, S100A9, and S100A12 [214]. S100A8/S100A9 augments the production of CXCL1, CXCL2, CXCL3, CXCL8, CCL20, IL-6, and TNF-α in keratinocytes [217], and it enhances keratinocyte proliferation [217]. The transcriptional co-activator IκBζ, encoded by the NFKBIZ gene, plays a critical role in IL-17A-, IL-17F-, and IL-17A/F-mediated signaling, such as the gene expression of S100A7 and CCL20 [218,219,220].
LL-37 is an antimicrobial peptide of human cathelicidin that is produced when keratinocytes are injured by a broad range of bacteria, viruses, and fungi [221,222]. In addition to antimicrobial peptide activity, LL-37 exhibits “alarmin” function, affects adenosine triphosphate-receptor P2X7 and Toll-like receptor (TLR) signaling, and EGFR transactivation or intracellular Ca2+ mobilization [223,224,225]. The released LL-37 binds to the infiltrated neutrophils [221]. Neutrophils are a rich source of extracellular DNA due to their neutrophil extracellular traps [226]. Upon stimulation with complexes of host DNA and LL-37, plasmacytoid dendritic cells produce large amounts of IFN-α [227]. Notably, LL-37 induced the proliferation of circulating CD3+ T cells in 24 out of 52 patients with psoriasis (46%) [144]; therefore, LL-37 is effective for autoantigens. In total, 50 LL-37-reactive CD3+ T cells, including both CD4+ and CD8+ T cells, express the skin-homing receptor cutaneous lymphocyte antigen [144]. LL-37 peptides bind to HLA-DR in dendritic cells and are presented to CD4+ T cells, while LL-37 peptides and the HLA-C*0602 complex activate CD8+ T cells [144,228]. The majority of LL-37-reactive CD3+ T cells produce IL-17, and the capacity of their IL-17 production is associated with disease severity [144]. Interestingly, the LL-37-specific IL-17-producing T cells are exclusively CD4+, whereas the LL-37-specific CD8+ T cells do not produce IL-17 [144]. A recent study by Takahashi et al. revealed that LL-37 can bind to self-RNA and stimulate macrophages to produce IL-6 via a scavenger receptor [229].
DEFB4A is highly expressed in psoriasis plaques and is the most psoriasis-specific antimicrobial peptide [230,231]. In contrast, DEFB4A is expressed at negligible or low levels in normal skin and skin lesions of eczema [230,231,232]. The expression of DEFB4A is upregulated by IL-17A and synergistically by IL-17A and TNF-α [215,220]. The serum levels of DEFB4A are highly specific biomarkers for disease activity in patients with psoriasis [215]. The number of neutrophil extracellular traps increase in psoriasis and upregulate the expression of DEFB4A [226]. Notably, DEFB4A is a functional (non-chemokine) ligand for CCR6 and feasibly attracts Th17 cells [233,234].
Although the biological implications of increased antimicrobial peptides in psoriasis remain obscure, they are intimately associated with IL-17A-rich milieu. Therefore, the upregulated expression of these antimicrobial peptides is rapidly normalized by the neutralization of IL-17A by ixekizumab [168] or secukinumab [69].
7. Conclusions
IL-17A is a multifunctional cytokine produced from adaptive and innate immune cells, such as Th17 and ILC3s. It orchestrates and promotes the peripheral tissue defense system against microbial insult, especially fungal infection. Psoriasis is a major inflammatory skin disease in which the interaction between IL-17A and epidermal keratinocytes plays a critical pathogenic role. IL-17A stimulates the proliferation of keratinocytes. Keratinocytes also produce a variety of antimicrobial peptides and cytochemokines in response to IL-17A. The antimicrobial peptides further exacerbate skin inflammation. CCL20 produced from IL-17A-stimulated keratinocytes recruits IL-17A-producing Th17 cells and ILC3s and accelerates the feed-forward vicious cycle, which causes fully developed psoriasis. This pathogenetic scheme has been verified with a high clinical efficacy of anti-IL-17A biologics. Therefore, psoriasis is considered an excellent human model of how IL-17A works with target peripheral tissues, and it provides in-depth insight into human autoinflammatory diseases.
Author Contributions
M.F. wrote the first draft. K.F., G.T. and T.N. reviewed the draft. M.F. finalized the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partly supported by a grant from The Ministry of Health, Labor, and Welfare in Japan (H30-Shokuhin-Shitei-005).
Conflicts of Interest
The authors have no conflicts of interest.
References
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S.M. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Takahashi, H.; Kawada, A.; Iizuka, H.; Nakagawa, H. Epidemiological survey from 2009 to 2012 of psoriatic patients in Japanese Society for Psoriasis Research. J. Dermatol. 2018, 45, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, E.; Okuyama, R.; Seki, T.; Kobayashi, A.; Oiso, N.; Muto, M.; Nakagawa, H.; Kawada, A. Epidemiological survey of patients with psoriasis in Matsumoto city, Nagano Prefecture, Japan. J. Dermatol. 2018, 45, 314–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiyama, S.; Ito, M.; Funasaka, Y.; Abe, M.; Nishida, E.; Muramatsu, S.; Nishihara, H.; Kato, H.; Morita, A.; Imafuku, S.; et al. Assessment of medication adherence and treatment satisfaction in Japanese patients with psoriasis of various severities. J. Dermatol. 2018, 45, 727–731. [Google Scholar] [CrossRef]
- Souza, C.S.; de Castro, C.C.S.; Carneiro, F.R.O.; Pinto, J.M.N.; Fabricio, L.H.Z.; Azulay-Abulafia, L.; Romiti, R.; Cestari, T.F.; Suzuki, C.E.; Biegun, P.M.; et al. Metabolic syndrome and psoriatic arthritis among patients with psoriasis vulgaris: Quality of life and prevalence. J. Dermatol. 2019, 46, 3–10. [Google Scholar] [CrossRef]
- Takahashi, H.; Satoh, K.; Takagi, A.; Iizuka, H. Cost-efficacy and pharmacoeconomics of psoriatic patients in Japan: Analysis from a single outpatient clinic. J. Dermatol. 2019, 46, 478–481. [Google Scholar] [CrossRef]
- Miller, R.A. The Koebner phenomenon. Int. J. Dermatol. 1982, 21, 192–197. [Google Scholar] [CrossRef]
- Weiss, G.; Shemer, A.; Trau, H. The Koebner phenomenon: Review of the literature. J. Eur. Acad. Dermatol. Venereol. 2002, 16, 241–248. [Google Scholar] [CrossRef]
- Dai, Y.X.; Wang, S.C.; Chou, Y.J.; Chang, Y.T.; Chen, T.J.; Li, C.P.; Wu, C.Y. Smoking, but not alcohol, is associated with risk of psoriasis in a Taiwanese population-based cohort study. J. Am. Acad. Dermatol. 2019, 80, 727–734. [Google Scholar] [CrossRef]
- Kamiya, K.; Kishimoto, M.; Sugai, J.; Komine, M.; Ohtsuki, M. Risk factors for the development of psoriasis. Int. J. Mol. Sci. 2019, 20, 4347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rielly, D.D.; Jani, M.; Rahman, P.; Elder, J.T. The Genetics of Psoriasis and Psoriatic Arthritis. J. Rheumatol Suppl. 2019, 95, 46–50. [Google Scholar] [PubMed]
- Elder, J.T. Expanded genome-wide association study meta-analysis of psoriasis expands the catalog of common psoriasis-associated variants. J. Investig. Dermatol. Symp. Proc. 2018, 19, S77–S78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diani, M.; Perego, S.; Sansoni, V.; Bertino, L.; Gomarasca, M.; Faraldi, M.; Pigatto, P.D.M.; Damiani, G.; Banfi, G.; Altomare, G.; et al. Differences in osteoimmunological biomarkers predictive of psoriatic arthritis among a large Italian cohort of psoriatic patients. Int. J. Mol. Sci. 2019, 20, 5617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mease, P.J.; Palmer, J.B.; Hur, P.; Strober, B.E.; Lebwohl, M.; Karki, C.; Reed, G.W.; Etzel, C.J.; Greenberg, J.D.; Helliwell, P.S. Utilization of the validated psoriasis epidemiology screening tool to identify signs and symptoms of psoriatic arthritis among those with psoriasis: A cross-sectional analysis from the US-based Corrona Psoriasis Registry. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 886–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Kawada, A. Clinical characteristics of Japanese patients with psoriatic arthritis: Comparison with East Asian countries. J. Dermatol. 2018, 45, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Ohtsuki, M.; Sano, S.; Morita, A.; Igarashi, A.; Okuyama, R.; Kawada, A. Late-onset psoriatic arthritis in Japanese patients. J. Dermatol. 2019, 46, 169–170. [Google Scholar] [CrossRef]
- Yamamoto, T.; Ohtsuki, M.; Sano, S.; Morita, A.; Igarashi, A.; Okuyama, R.; Kawada, A. Switching biologics in the treatment of psoriatic arthritis in Japan. J. Dermatol. 2019, 46, e113–e114. [Google Scholar] [CrossRef]
- Umezawa, Y.; Yanaba, K.; Asahina, A.; Nakagawa, H.; Fukuda, T.; Fukuda, K. Usefulness of dual-energy computed tomography for the evaluation of psoriatic arthritis accompanied by knee osteoarthritis. J. Dermatol. 2019, 46, e30–e32. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Tsuji, G.; Kadono, T.; Nakahara, T.; Furue, M. Autoimmunity and autoimmune co-morbidities in psoriasis. Immunology 2018, 154, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Ho, Y.H.; Hu, H.Y.; Chang, Y.T.; Li, C.P.; Wu, C.Y. Psoriasis is associated with increased risk of bullous pemphigoid: A nationwide population-based cohort study in Taiwan. J. Dermatol. 2019, 46, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Ichiyama, S.; Hoashi, T.; Kanda, N.; Hashimoto, H.; Matsushita, M.; Nozawa, K.; Ueno, T.; Saeki, H. Psoriasis vulgaris associated with systemic lupus erythematosus successfully treated with apremilast. J. Dermatol. 2019, 46, e219–e221. [Google Scholar] [CrossRef] [PubMed]
- Chujo, S.; Asahina, A.; Itoh, Y.; Kobayashi, K.; Sueki, H.; Ishiji, T.; Umezawa, Y.; Nakagawa, H. New onset of psoriasis during nivolumab treatment for lung cancer. J. Dermatol. 2018, 45, e55–e56. [Google Scholar] [CrossRef] [PubMed]
- Kamata, M.; Asano, Y.; Shida, R.; Maeda, N.; Yoshizaki, A.; Miyagaki, T.; Kawashima, T.; Tada, Y.; Sato, S. Secukinumab decreased circulating anti-BP180-NC16a autoantibodies in a patient with coexisting psoriasis vulgaris and bullous pemphigoid. J. Dermatol. 2019, 46, e216–e217. [Google Scholar] [CrossRef]
- Bayaraa, B.; Imafuku, S. Relationship between environmental factors, age of onset and familial history in Japanese patients with psoriasis. J. Dermatol. 2018, 45, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.Y.; Chang, W.L.; Shiu, M.N.; Huang, W.F.; Tsai, T.F. Psoriasis is associated with a greater risk for cardiovascular procedure and surgery in patients with hypertension: A nationwide cohort study. J. Dermatol. 2018, 45, 1381–1388. [Google Scholar] [CrossRef]
- Furue, M.; Kadono, T. “Inflammatory skin march” in atopic dermatitis and psoriasis. Inflamm. Res. 2017, 66, 833–842. [Google Scholar] [CrossRef]
- Momose, M.; Asahina, A.; Fukuda, T.; Sakuma, T.; Umezawa, Y.; Nakagawa, H. Evaluation of epicardial adipose tissue volume and coronary artery calcification in Japanese patients with psoriasis vulgaris. J. Dermatol. 2018, 45, 1349–1352. [Google Scholar] [CrossRef]
- Takamura, S.; Takahashi, A.; Inoue, Y.; Teraki, Y. Effects of tumor necrosis factor-α, interleukin-23 and interleukin-17A inhibitors on bodyweight and body mass index in patients with psoriasis. J. Dermatol. 2018, 45, 1130–1134. [Google Scholar] [CrossRef]
- Wang, C.C.; Tang, C.H.; Huang, K.C.; Huang, S.Y.; Sue, Y.M. Increased risk of incident psoriasis in end-stage renal disease patients on chronic hemodialysis: A nationwide population-based cohort study. J. Dermatol. 2018, 45, 1063–1070. [Google Scholar] [CrossRef]
- Han, J.H.; Lee, J.H.; Han, K.D.; Kim, H.N.; Bang, C.H.; Park, Y.M.; Lee, J.Y.; Kim, T.Y. Increased risk of psoriasis in subjects with abdominal obesity: A nationwide population-based study. J. Dermatol. 2019, 46, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.J.; Kim, T.G.; Lee, J.W.; Lee, M.; Oh, J.; Lee, S.E.; Chang, H.J.; Jee, S.H.; Lee, M.G. Increased risk of atherosclerotic cardiovascular disease among patients with psoriasis in Korea: A 15-year nationwide population-based cohort study. J. Dermatol. 2019, 46, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Masaki, S.; Bayaraa, B.; Imafuku, S. Prevalence of inflammatory bowel disease in Japanese psoriatic patients. J. Dermatol. 2019, 46, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Tokuyama, M.; Shimizu, T.; Yamada, T.; Kondoh, A.; Mabuchi, T. Case of psoriasis vulgaris with atrial fibrillation, heart failure and chronic kidney disease which were found accidentally through blood examination during apremilast treatment. J. Dermatol. 2019, 46, e239–e240. [Google Scholar] [CrossRef]
- Yamazaki, F.; Takehana, K.; Tamashima, M.; Okamoto, H. Improvement in abnormal coronary arteries estimated by coronary computed tomography angiography after secukinumab treatment in a Japanese psoriatic patient. J. Dermatol. 2019, 46, e51–e52. [Google Scholar] [CrossRef]
- Wójcik, P.; Biernacki, M.; Wroński, A.; Łuczaj, W.; Waeg, G.; Žarković, N.; Skrzydlewska, E. Altered lipid metabolism in blood mononuclear cells of psoriatic patients indicates differential changes in psoriasis vulgaris and psoriatic arthritis. Int. J. Mol. Sci. 2019, 20, 4249. [Google Scholar]
- Lee, J.H.; Kim, H.J.; Han, K.D.; Kim, H.N.; Park, Y.M.; Lee, J.Y.; Park, Y.G.; Lee, Y.B. Cancer risk in 892089 patients with psoriasis in Korea: A nationwide population-based cohort study. J. Dermatol. 2019, 46, 95–102. [Google Scholar] [CrossRef]
- Hsu, S.; Papp, K.A.; Lebwohl, M.G.; Bagel, J.; Blauvelt, A.; Duffin, K.C.; Crowley, J.; Eichenfield, L.F.; Feldman, S.R.; Fiorentino, D.F.; et al. Consensus guidelines for the management of plaque psoriasis. Arch. Dermatol. 2012, 148, 95–102. [Google Scholar] [CrossRef]
- Imafuku, S.; Zheng, M.; Tada, Y.; Zhang, X.; Theng, C.; Thevarajah, S.; Zhao, Y.; Song, H.J. Asian consensus on assessment and management of mild to moderate plaque psoriasis with topical therapy. J. Dermatol. 2018, 45, 805–811. [Google Scholar] [CrossRef]
- Meephansan, J.; Subpayasarn, U.; Ponnikorn, S.; Chakkavittumrong, P.; Juntongjin, P.; Komine, M.; Ohtsuki, M.; Poovorawan, Y. Methotrexate, but not narrowband ultraviolet B radiation, suppresses interleukin-33 mRNA levels in psoriatic plaques and protein levels in serum of patients with psoriasis. J. Dermatol. 2018, 45, 322–325. [Google Scholar] [CrossRef]
- Kishimoto, M.; Komine, M.; Hioki, T.; Kamiya, K.; Sugai, J.; Ohtsuki, M. Real-world use of apremilast for patients with psoriasis in Japan. J. Dermatol. 2018, 45, 1345–1348. [Google Scholar] [CrossRef] [PubMed]
- Morita, A. Current developments in phototherapy for psoriasis. J. Dermatol. 2018, 45, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, E.H.; Koh, W.S.; Shin, J.M.; Kim, J.E.; Ko, J.Y.; Ro, Y.S. Clinical experience of cyclosporin treatment in patients with psoriasis and psoriatic arthritis. J. Dermatol. 2018, 45, 329–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pongparit, K.; Chularojanamontri, L.; Limphoka, P.; Silpa-Archa, N.; Wongpraparat, C. Effectiveness of and factors associated with clinical response to methotrexate under daily life conditions in Asian patients with psoriasis: A retrospective cohort study. J. Dermatol. 2018, 45, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, S.; Osawa, R.; Nakajima, H.; Nakajima, K.; Sano, S. Favorable response to apremilast in a patient with refractory psoriasis verrucosa. J. Dermatol. 2019, 46, 544–547. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Furue, M. Differential efficacy of biologic treatments targeting the TNF-α/IL-23/IL-17 axis in psoriasis and psoriatic arthritis. Cytokine 2018, 111, 182–188. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Tsuji, G.; Kadono, T.; Furue, M. Psoriasis and the TNF/IL23/IL17 axis. G. Ital. Dermatol. Venereol. 2019, 154, 418–424. [Google Scholar] [CrossRef]
- Sawyer, L.M.; Malottki, K.; Sabry-Grant, C.; Yasmeen, N.; Wright, E.; Sohrt, A.; Borg, E.; Warren, R.B. Assessing the relative efficacy of interleukin-17 and interleukin-23 targeted treatments for moderate-to-severe plaque psoriasis: A systematic review and network meta-analysis of PASI response. PLoS ONE 2019, 14, e0220868. [Google Scholar] [CrossRef] [Green Version]
- Kamata, M.; Tada, Y. Safety of biologics in psoriasis. J. Dermatol. 2018, 45, 279–286. [Google Scholar] [CrossRef]
- Momose, M.; Asahina, A.; Umezawa, Y.; Nakagawa, H. Long-term clinical efficacy and safety of secukinumab for Japanese patients with psoriasis: A single-center experience. J. Dermatol. 2018, 45, 318–321. [Google Scholar]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Dermatol. 2018, 45, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsuki, M.; Kubo, H.; Morishima, H.; Goto, R.; Zheng, R.; Nakagawa, H. Guselkumab, an anti-interleukin-23 monoclonal antibody, for the treatment of moderate to severe plaque-type psoriasis in Japanese patients: Efficacy and safety results from a phase 3, randomized, double-blind, placebo-controlled study. J. Dermatol. 2018, 45, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Bayaraa, B.; Imafuku, S. Sustainability and switching of biologics for psoriasis and psoriatic arthritis at Fukuoka University Psoriasis Registry. J. Dermatol. 2019, 46, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Karakawa, M.; Komine, M.; Kishimoto, M.; Sugai, J.; Ohtsuki, M. Results of a retrospective study on the efficacy and safety of adalimumab 80 mg administrated every other week in patients with psoriasis at a single Japanese institution. J. Dermatol. 2019, 46, 199–205. [Google Scholar] [CrossRef]
- Komatsu-Fujii, T.; Honda, T.; Otsuka, A.; Kabashima, K. Inverse responses of the skin and nail lesions of psoriatic arthritis to an anti-interleukin-17A antibody and an anti-tumor necrosis factor-α antibody. J. Dermatol. 2019, 46, e440–e441. [Google Scholar] [CrossRef]
- Komatsu-Fujii, T.; Honda, T.; Otsuka, A.; Kabashima, K. Improvement of nail lesions in a patient with psoriatic arthritis by switching the treatment from an anti-interleukin-17A antibody to an anti-tumor necrosis factor-α antibody. J. Dermatol. 2019, 46, e158–e160. [Google Scholar] [CrossRef]
- Lee, M.G.; Huang, Y.H.; Lee, J.H.; Lee, S.C.; Kim, T.G.; Aw, D.C.; Bao, W.; Dee, C.M.A.; Guana, A.; Tsai, T.F. Secukinumab demonstrates superior efficacy and a faster response in clearing skin in Asian subjects with moderate to severe plaque psoriasis compared with ustekinumab: Subgroup analysis from the CLEAR study. J. Dermatol. 2019, 46, 752–758. [Google Scholar] [CrossRef]
- Okubo, Y.; Ohtsuki, M.; Morita, A.; Yamaguchi, M.; Shima, T.; Tani, Y.; Nakagawa, H. Long-term efficacy and safety of secukinumab in Japanese patients with moderate to severe plaque psoriasis: 3-year results of a double-blind extension study. J. Dermatol. 2019, 46, 186–192. [Google Scholar] [CrossRef]
- Shibata, T.; Muto, J.; Takama, H.; Yanagishita, T.; Ito, T.; Watanabe, D. Case of psoriatic erythroderma induced by the discontinuation of the chronic use of topical steroid after dialysis initiation and successfully treated with secukinumab. J. Dermatol. 2019, 46, e119–e120. [Google Scholar] [CrossRef]
- Tada, Y.; Ishii, K.; Kimura, J.; Hanada, K.; Kawaguchi, I. Patient preference for biologic treatments of psoriasis in Japan. J. Dermatol. 2019, 46, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Tsuruta, N.; Narisawa, Y.; Imafuku, S.; Ito, K.; Yamaguchi, K.; Miyagi, T.; Takahashi, K.; Fukamatsu, H.; Morizane, S.; Koketsu, H.; et al. Cross-sectional multicenter observational study of psoriatic arthritis in Japanese patients: Relationship between skin and joint symptoms and results of treatment with tumor necrosis factor-α inhibitors. J. Dermatol. 2019, 46, 193–198. [Google Scholar] [CrossRef]
- Veale, D.J.; Fearon, U. The pathogenesis of psoriatic arthritis. Lancet 2018, 391, 2273–2284. [Google Scholar] [CrossRef]
- Assefa, G.T.; Kaneko, S.; Oguro, H.; Morita, E. Treatment of psoriasis and psoriatic arthritis with secukinumab after unsatisfactory response to ustekinumab in multiple sclerosis patient. J. Dermatol. 2019, 46, e112–e113. [Google Scholar] [CrossRef] [PubMed]
- Mourad, A.; Gniadecki, R. Treatment of dactylitis and enthesitis in psoriatic arthritis with biologic agents: A systematic review and metaanalysis. J. Rheumatol. 2019, 47, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Korman, N.J. Management of psoriasis as a systemic disease: What is the evidence? Br. J. Dermatol. 2019. [Google Scholar] [CrossRef]
- Lockshin, B.; Balagula, Y.; Merola, J.F. Interleukin 17, inflammation, and cardiovascular risk in patients with psoriasis. J. Am. Acad. Dermatol. 2018, 79, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Erichsen, C.Y.; Jensen, P.; Kofoed, K. Biologic therapies targeting the interleukin (IL)-23/IL-17 immune axis for the treatment of moderate-to-severe plaque psoriasis: A systematic review and meta-analysis. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Krueger, J.G.; Brunner, P.M. Interleukin-17 alters the biology of many cell types involved in the genesis of psoriasis, systemic inflammation and associated comorbidities. Exp. Dermatol. 2018, 27, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Krueger, J.G.; Wharton, K.A., Jr.; Schlitt, T.; Suprun, M.; Torene, R.I.; Jiang, X.; Wang, C.Q.; Fuentes-Duculan, J.; Hartmann, N.; Peters, T.; et al. IL-17A inhibition by secukinumab induces early clinical, histopathologic, and molecular resolution of psoriasis. J. Allergy Clin. Immunol. 2019, 144, 750–763. [Google Scholar] [CrossRef] [Green Version]
- Mease, P.J.; Smolen, J.S.; Behrens, F.; Nash, P.; Liu Leage, S.; Li, L.; Tahir, H.; Gooderham, M.; Krishnan, E.; Liu-Seifert, H.; et al. A head-to-head comparison of the efficacy and safety of ixekizumab and adalimumab in biological-naïve patients with active psoriatic arthritis: 24-week results of a randomised, open-label, blinded-assessor trial. Ann. Rheum. Dis. 2020, 79, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.B.; Barker, J.; Finlay, A.Y.; Burden, A.D.; Kirby, B.; Armendariz, Y.; Williams, R.; Hatchard, C.; Khare, S.; Griffiths, C.E.M. Secukinumab for patients failing previous TNFα-inhibitor therapy: Results of a randomised open-label study (Signature). Br. J. Dermatol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkes, J.E.; Yan, B.Y.; Chan, T.C.; Krueger, J.G. Discovery of the IL-23/IL-17 signaling pathway and the treatment of psoriasis. J. Immunol. 2018, 201, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Shiga, T.; Sato, K.; Kataoka, S.; Sano, S. TNF inhibitors directly target Th17 cells via attenuation of autonomous TNF/TNFR2 signalling in psoriasis. J. Dermatol. Sci. 2015, 77, 79–81. [Google Scholar] [CrossRef]
- Schüler, R.; Brand, A.; Klebow, S.; Wild, J.; Veras, F.P.; Ullmann, E.; Roohani, S.; Kolbinger, F.; Kossmann, S.; Wohn, C.; et al. Antagonization of IL-17A attenuates skin inflammation and vascular dysfunction in mouse models of psoriasis. J. Investig. Dermatol. 2019, 139, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Moos, S.; Mohebiany, A.N.; Waisman, A.; Kurschus, F.C. Imiquimod-induced psoriasis in mice depends on the IL-17 signaling of keratinocytes. J. Investig. Dermatol. 2019, 139, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, T.; Takekoshi, T.; Hwang, S.T. Epidermal CCR6+ γδ T cells are major producers of IL-22 and IL-17 in a murine model of psoriasiform dermatitis. J. Immunol. 2011, 187, 5026–5031. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, T.; Singh, T.P.; Takekoshi, T.; Jia, G.F.; Wu, X.; Kao, M.C.; Weiss, I.; Farber, J.M.; Hwang, S.T. CCR6 is required for epidermal trafficking of γδ-T cells in an IL-23-induced model of psoriasiform dermatitis. J. Investig. Dermatol. 2013, 133, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.; Sano, S. Mouse models of psoriasis and their relevance. J. Dermatol. 2018, 45, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Brembilla, N.C.; Senra, L.; Boehncke, W.H. The IL-17 family of cytokines in psoriasis: IL-17A and beyond. Front. Immunol. 2018, 9, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 family of cytokines in health and disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Buckley, K.M.; Ho, E.C.H.; Hibino, T.; Schrankel, C.S.; Schuh, N.W.; Wang, G.; Rast, J.P. IL17 factors are early regulators in the gut epithelium during inflammatory response to Vibrio in the sea urchin larva. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Das, S.; Hirano, M.; Holland, S.J.; McCurley, N.; Guo, P.; Rosenberg, C.S.; Boehm, T.; Cooper, M.D. Characterization of lamprey IL-17 family members and their receptors. J. Immunol. 2015, 195, 5440–5451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktar, M.K.; Kido-Nakahara, M.; Furue, M.; Nakahara, T. Mutual upregulation of endothelin-1 and IL-25 in atopic dermatitis. Allergy 2015, 70, 846–854. [Google Scholar] [CrossRef]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, Z.; Slinger, E.; Bongers, G.; Lapenda, T.L.S.; Pacer, M.E.; Jiao, J.; Beltrao, M.F.; Soto, A.J.; Harpaz, N.; et al. IL-23 activates innate lymphoid cells to promote neonatal intestinal pathology. Mucosal Immunol. 2015, 8, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Shen, X.; Ding, C.; Qi, C.; Li, K.; Li, X.; Jala, V.R.; Zhang, H.G.; Wang, T.; Zheng, J.; et al. Pivotal role of dermal IL-17-producing γδ T cells in skin inflammation. Immunity 2011, 35, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Villanova, F.; Flutter, B.; Tosi, I.; Grys, K.; Sreeneebus, H.; Perera, G.K.; Chapman, A.; Smith, C.H.; Di Meglio, P.; Nestle, F.O. Characterization of innate lymphoid cells in human skin and blood demonstrates increase of NKp44+ ILC3 in psoriasis. J. Investig. Dermatol. 2014, 134, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, G.; Umemura, M. Interleukin-17 family cytokines in protective immunity against infections: Role of hematopoietic cell-derived and non-hematopoietic cell-derived interleukin-17s. Microbiol. Immunol. 2018, 62, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.H.; Reynolds, J.M.; Pappu, B.P.; Chen, G.; Martinez, G.J.; Dong, C. Interleukin-17C promotes Th17 cell responses and autoimmune disease via interleukin-17 receptor E. Immunity 2011, 35, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Ota, N.; Peng, I.; Refino, C.J.; Danilenko, D.M.; Caplazi, P.; Ouyang, W. IL-17RC is required for IL-17A- and IL-17F-dependent signaling and the pathogenesis of experimental autoimmune encephalomyelitis. J. Immunol. 2010, 184, 4307–4316. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Huang, J.; Zhao, X.; Lu, H.; Wang, W.; Yang, X.O.; Shi, Y.; Wang, X.; Lai, Y.; Dong, C. Interleukin-17 receptor D constitutes an alternative receptor for interleukin-17A important in psoriasis-like skin inflammation. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Liu, C.; Swaidani, S.; Qian, W.; Kang, Z.; Sun, P.; Han, Y.; Wang, C.; Gulen, M.F.; Yin, W.; Zhang, C.; et al. A CC’ loop decoy peptide blocks the interaction between Act1 and IL-17RA to attenuate IL-17- and IL-25-induced inflammation. Sci.Signal 2011, 4, ra72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sønder, S.U.; Saret, S.; Tang, W.; Sturdevant, D.E.; Porcella, S.F.; Siebenlist, U. IL-17-induced NF-kappaB activation via CIKS/Act1: Physiologic significance and signaling mechanisms. J. Biol. Chem. 2011, 286, 12881–12890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, S.; Puel, A.; Casanova, J.L.; Kobayashi, M. Chronic mucocutaneous candidiasis disease associated with inborn errors of IL-17 immunity. Clin. Transl. Immunology 2016, 5, e114. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, S.; Zheng, G.; Huang, J.; Songyang, Z.; Zhao, X.; Lin, X. Gain-of-function mutation of Card14 leads to spontaneous psoriasis-like skin inflammation through enhanced keratinocyte response to IL-17A. Immunity 2018, 49, 66–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Cai, G.; Liu, C.; Zhao, J.; Gu, C.; Wu, L.; Hamilton, T.A.; Zhang, C.J.; Ko, J.; Zhu, L.; et al. IL-17R-EGFR axis links wound healing to tumorigenesis in Lrig1(+) stem cells. J. Exp. Med. 2019, 216, 195–214. [Google Scholar] [CrossRef] [Green Version]
- Tsang, M.; Friesel, R.; Kudoh, T.; Dawid, I.B. Identification of Sef, a novel modulator of FGF signalling. Nat. Cell Biol. 2002, 4, 165–169. [Google Scholar] [CrossRef]
- Calautti, E.; Avalle, L.; Poli, V. Psoriasis: A STAT3-centric view. Int. J. Mol. Sci. 2018, 19, 171. [Google Scholar] [CrossRef] [Green Version]
- Sano, S.; Chan, K.S.; Carbajal, S.; Clifford, J.; Peavey, M.; Kiguchi, K.; Itami, S.; Nickoloff, B.J.; DiGiovanni, J. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model. Nat. Med. 2005, 11, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, K.; Takaishi, M.; Nakajima, K.; Ikeda, M.; Kanda, T.; Tarutani, M.; Iiyama, T.; Asao, N.; DiGiovanni, J.; Sano, S. Stat3 as a therapeutic target for the treatment of psoriasis: A clinical feasibility study with STA-21, a Stat3 inhibitor. J. Investig. Dermatol. 2011, 131, 108–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.C.; Jeong, S.H.; Kim, J.H.; Lee, H.; Ryu, W.I.; Kim, M.G.; Son, E.D.; Lee, T.R.; Son, S.W. RIP4 upregulates CCL20 expression through STAT3 signalling in cultured keratinocytes. Exp. Dermatol. 2018, 27, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Jin, L.; Dang, E.; Chang, T.; Feng, Z.; Liu, Y.; Wang, G. IL-17A upregulates keratin 17 expression in keratinocytes through STAT1- and STAT3-dependent mechanisms. J. Investig. Dermatol. 2011, 131, 2401–2408. [Google Scholar] [CrossRef] [Green Version]
- Honma, M.; Minami-Hori, M.; Takahashi, H.; Iizuka, H. Podoplanin expression in wound and hyperproliferative psoriatic epidermis: Regulation by TGF-β and STAT-3 activating cytokines, IFN-γ, IL-6, and IL-22. J. Dermatol. Sci. 2012, 65, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, K.; Takaishi, M.; Digiovanni, J.; Sano, S. Attenuation of psoriasis-like skin lesion in a mouse model by topical treatment with indirubin and its derivastive E804. J. Dermatol. Sci. 2012, 65, 70–72. [Google Scholar] [CrossRef]
- Ekman, A.K.; Bivik Eding, C.; Rundquist, I.; Enerbäck, C. IL-17 and IL-22 promote keratinocyte stemness in the germinative compartment in psoriasis. J. Investig. Dermatol. 2019, 139, 1564–1573. [Google Scholar] [CrossRef] [Green Version]
- Karbach, S.; Croxford, A.L.; Oelze, M.; Schüler, R.; Minwegen, D.; Wegner, J.; Koukes, L.; Yogev, N.; Nikolaev, A.; Reißig, S.; et al. Interleukin 17 drives vascular inflammation, endothelial dysfunction, and arterial hypertension in psoriasis-like skin disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2658–2668. [Google Scholar] [CrossRef] [Green Version]
- Cordoro, K.M.; Hitraya-Low, M.; Taravati, K.; Sandoval, P.M.; Kim, E.; Sugarman, J.; Pauli, M.L.; Liao, W.; Rosenblum, M.D. Skin-infiltrating, interleukin-22-producing T cells differentiate pediatric psoriasis from adult psoriasis. J. Am. Acad. Dermatol. 2017, 77, 417–424. [Google Scholar] [CrossRef]
- Diani, M.; Altomare, G.; Reali, E. T cell responses in psoriasis and psoriatic arthritis. Autoimmun. Rev. 2015, 14, 286–292. [Google Scholar] [CrossRef]
- Puel, A.; Cypowyj, S.; Bustamante, J.; Wright, J.F.; Liu, L.; Lim, H.K.; Migaud, M.; Israel, L.; Chrabieh, M.; Audry, M.; et al. Chronic mucocutaneous candidiasis in humans with inborn errors of interleukin-17 immunity. Science 2011, 332, 65–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisson, B.; Wang, C.; Pedergnana, V.; Wu, L.; Cypowyj, S.; Rybojad, M.; Belkadi, A.; Picard, C.; Abel, L.; Fieschi, C.; et al. An ACT1 mutation selectively abolishes interleukin-17 responses in humans with chronic mucocutaneous candidiasis. Immunity 2013, 39, 676–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Y.; Cypowyj, S.; Aytekin, C.; Galicchio, M.; Camcioglu, Y.; Nepesov, S.; Ikinciogullari, A.; Dogu, F.; Belkadi, A.; Levy, R.; et al. Inherited IL-17RC deficiency in patients with chronic mucocutaneous candidiasis. J. Exp. Med. 2015, 212, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.C.; Whibley, N.; Mamo, A.J.; Siebenlist, U.; Chan, Y.R.; Gaffen, S.L. Interleukin-17-induced protein lipocalin 2 is dispensable for immunity to oral candidiasis. Infect. Immun. 2014, 82, 1030–1035. [Google Scholar] [CrossRef] [Green Version]
- Conti, H.R.; Shen, F.; Nayyar, N.; Stocum, E.; Sun, J.N.; Lindemann, M.J.; Ho, A.W.; Hai, J.H.; Yu, J.J.; Jung, J.W.; et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. J. Exp. Med. 2009, 206, 299–311. [Google Scholar] [CrossRef]
- Conti, H.R.; Gaffen, S.L. IL-17-mediated immunity to the opportunistic fungal pathogen Candida albicans. J. Immunol. 2015, 195, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Vinh, D.C.; Casanova, J.L.; Puel, A. Inborn errors of immunity underlying fungal diseases in otherwise healthy individuals. Curr. Opin. Microbiol. 2017, 40, 46–57. [Google Scholar] [CrossRef]
- Sparber, F.; De Gregorio, C.; Steckholzer, S.; Ferreira, F.M.; Dolowschiak, T.; Ruchti, F.; Kirchner, F.R.; Mertens, S.; Prinz, I.; Joller, N.; et al. The skin commensal yeast Malassezia triggers a type 17 response that coordinates anti-fungal immunity and exacerbates skin inflammation. Cell Host Microbe 2019, 25, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Wohn, C.; Brand, A.; van Ettinger, K.; Brouwers-Haspels, I.; Waisman, A.; Laman, J.D.; Clausen, B.E. Gradual development of psoriatic skin lesions by constitutive low-level expression of IL-17A. Cell. Immunol. 2016, 308, 57–65. [Google Scholar] [CrossRef]
- Aggarwal, S.; Ghilardi, N.; Xie, M.H.; de Sauvage, F.J.; Gurney, A.L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 2003, 278, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffen, S.L. Structure and signalling in the IL-17 receptor family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, J.A.; Bayer, A.C.; Ider, K.; Bismuth, J.; Truffault, F.; Roussin, R.; Santelmo, N.; Le Panse, R.; Berrih-Aknin, S.; Dragin, N. Il-23/Th17 cell pathway: A promising target to alleviate thymic inflammation maintenance in myasthenia gravis. J. Autoimmun. 2019, 98, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.R.; Blumenschein, W.; Murphy, E.; Diveu, C.; Wiekowski, M.; Abbondanzo, S.; Lucian, L.; Geissler, R.; Brodie, S.; Kimball, A.B.; et al. IL-23 stimulates epidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasis pathogenesis. J. Exp. Med. 2006, 203, 2577–2587. [Google Scholar] [CrossRef]
- Furue, M.; Kadono, T. The contribution of IL-17 to the development of autoimmunity in psoriasis. Innate Immun. 2019, 25, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, A.; Siewert, K.; Stöhr, J.; Besgen, P.; Kim, S.M.; Rühl, G.; Nickel, J.; Vollmer, S.; Thomas, P.; Krebs, S.; et al. Melanocyte antigen triggers autoimmunity in human psoriasis. J. Exp. Med. 2015, 212, 2203–2212. [Google Scholar] [CrossRef]
- Lee, E.; Trepicchio, W.L.; Oestreicher, J.L.; Pittman, D.; Wang, F.; Chamian, F.; Dhodapkar, M.; Krueger, J.G. Increased expression of interleukin 23 p19 and p40 in lesional skin of patients with psoriasis vulgaris. J. Exp. Med. 2004, 199, 125–130. [Google Scholar] [CrossRef]
- Zaba, L.C.; Cardinale, I.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Novitskaya, I.; Khatcherian, A.; Bluth, M.J.; Lowes, M.A.; et al. Amelioration of epidermal hyperplasia by TNF inhibition is associated with reduced Th17 responses. J. Exp. Med. 2007, 204, 3183–3194. [Google Scholar] [CrossRef]
- Piskin, G.; Sylva-Steenland, R.M.; Bos, J.D.; Teunissen, M.B. In vitro and in situ expression of IL-23 by keratinocytes in healthy skin and psoriasis lesions: Enhanced expression in psoriatic skin. J. Immunol. 2006, 176, 1908–1915. [Google Scholar] [CrossRef]
- Li, H.; Yao, Q.; Mariscal, A.G.; Wu, X.; Hülse, J.; Pedersen, E.; Helin, K.; Waisman, A.; Vinkel, C.; Thomsen, S.F.; et al. Epigenetic control of IL-23 expression in keratinocytes is important for chronic skin inflammation. Nat. Commun. 2018, 9, 1420. [Google Scholar] [CrossRef]
- Tu, Z.; Zhang, S.; Zhou, G.; Zhou, L.; Xiang, Q.; Chen, Q.; Zhao, P.; Zhan, H.; Zhou, H.; Sun, L. LMO4 is a disease-provocative transcription coregulator activated by IL-23 in psoriatic keratinocytes. J. Investig. Dermatol. 2018, 138, 1078–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McInnes, I.B.; Kavanaugh, A.; Gottlieb, A.B.; Puig, L.; Rahman, P.; Ritchlin, C.; Brodmerkel, C.; Li, S.; Wang, Y.; Mendelsohn, A.M.; et al. Efficacy and safety of ustekinumab in patients with active psoriatic arthritis: 1 year results of the phase 3, multicentre, double-blind, placebo-controlled PSUMMIT 1 trial. Lancet 2013, 382, 780–789. [Google Scholar] [CrossRef]
- Papp, K.A.; Blauvelt, A.; Bukhalo, M.; Gooderham, M.; Krueger, J.G.; Lacour, J.P.; Menter, A.; Philipp, S.; Sofen, H.; Tyring, S.; et al. Risankizumab versus ustekinumab for moderate-to-severe plaque psoriasis. N. Engl. J. Med. 2017, 376, 1551–1560. [Google Scholar] [CrossRef] [PubMed]
- Blauvelt, A.; Papp, K.A.; Griffiths, C.E.; Randazzo, B.; Wasfi, Y.; Shen, Y.K.; Li, S.; Kimball, A.B. Efficacy and safety of guselkumab, an anti-interleukin-23 monoclonal antibody, compared with adalimumab for the continuous treatment of patients with moderate to severe psoriasis: Results from the phase III, double-blinded, placebo- and active comparator-controlled VOYAGE 1 trial. J. Am. Acad. Dermatol. 2017, 76, 405–417. [Google Scholar]
- Reich, K.; Papp, K.A.; Blauvelt, A.; Tyring, S.K.; Sinclair, R.; Thaçi, D.; Nograles, K.; Mehta, A.; Cichanowitz, N.; Li, Q.; et al. Tildrakizumab versus placebo or etanercept for chronic plaque psoriasis (reSURFACE 1 and reSURFACE 2): Results from two randomised controlled, phase 3 trials. Lancet 2017, 390, 276–288. [Google Scholar] [CrossRef]
- Johansen, C.; Usher, P.A.; Kellerup, R.B.; Lundsgaard, D.; Iversen, L.; Kragballe, K. Characterization of the interleukin-17 isoforms and receptors in lesional psoriatic skin. Br. J. Dermatol. 2009, 160, 319–324. [Google Scholar] [CrossRef]
- Johnston, A.; Fritz, Y.; Dawes, S.M.; Diaconu, D.; Al-Attar, P.M.; Guzman, A.M.; Chen, C.S.; Fu, W.; Gudjonsson, J.E.; McCormick, T.S.; et al. Keratinocyte overexpression of IL-17C promotes psoriasiform skin inflammation. J. Immunol. 2013, 190, 2252–2262. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.A.; Towne, J.E.; Kricorian, G.; Klekotka, P.; Gudjonsson, J.E.; Krueger, J.G.; Russell, C.B. The emerging role of IL-17 in the pathogenesis of psoriasis: Preclinical and clinical findings. J. Investig. Dermatol. 2013, 133, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Carrozzi, V.; Sambandam, A.; Luis, E.; Lin, Z.; Jeet, S.; Lesch, J.; Hackney, J.; Kim, J.; Zhou, M.; Lai, J.; et al. IL-17C regulates the innate immune function of epithelial cells in an autocrine manner. Nat. Immunol. 2011, 12, 1159–1166. [Google Scholar] [CrossRef]
- Vandeghinste, N.; Klattig, J.; Jagerschmidt, C.; Lavazais, S.; Marsais, F.; Haas, J.D.; Auberval, M.; Lauffer, F.; Moran, T.; Ongenaert, M.; et al. Neutralization of IL-17C reduces skin inflammation in mouse models of psoriasis and atopic dermatitis. J. Investig. Dermatol. 2018, 138, 1555–1563. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Krueger, J.G. IL-17C: A unique epithelial cytokine with potential for targeting across the spectrum of atopic dermatitis and psoriasis. J. Investig. Dermatol. 2018, 138, 1467–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsoi, L.C.; Rodriguez, E.; Degenhardt, F.; Baurecht, H.; Wehkamp, U.; Volks, N.; Szymczak, S.; Swindell, W.R.; Sarkar, M.K.; Raja, K.; et al. Atopic dermatitis is an IL-13-dominant disease with greater molecular heterogeneity compared to psoriasis. J. Investig. Dermatol. 2019, 139, 1480–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pène, J.; Chevalier, S.; Preisser, L.; Vénéreau, E.; Guilleux, M.H.; Ghannam, S.; Molès, J.P.; Danger, Y.; Ravon, E.; Lesaux, S.; et al. Chronically inflamed human tissues are infiltrated by highly differentiated Th17 lymphocytes. J. Immunol. 2008, 180, 7423–7430. [Google Scholar] [CrossRef] [Green Version]
- Lande, R.; Botti, E.; Jandus, C.; Dojcinovic, D.; Fanelli, G.; Conrad, C.; Chamilos, G.; Feldmeyer, L.; Marinari, B.; Chon, S.; et al. The antimicrobial peptide LL37 is a T-cell autoantigen in psoriasis. Nat. Commun. 2014, 5, 5621. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Duculan, J.; Bonifacio, K.M.; Hawkes, J.E.; Kunjravia, N.; Cueto, I.; Li, X.; Gonzalez, J.; Garcet, S.; Krueger, J.G. Autoantigens ADAMTSL5 and LL37 are significantly upregulated in active Psoriasis and localized with keratinocytes, dendritic cells and other leukocytes. Exp. Dermatol. 2017, 26, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Yunusbaeva, M.; Valiev, R.; Bilalov, F.; Sultanova, Z.; Sharipova, L.; Yunusbayev, B. Psoriasis patients demonstrate HLA-Cw*06:02 allele dosage-dependent T cell proliferation when treated with hair follicle-derived keratin 17 protein. Sci. Rep. 2018, 8, 6098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, K.L.; Jarrett, R.; Subramaniam, S.; Salimi, M.; Gutowska-Owsiak, D.; Chen, Y.L.; Hardman, C.; Xue, L.; Cerundolo, V.; Ogg, G. Psoriatic T cells recognize neolipid antigens generated by mast cell phospholipase delivered by exosomes and presented by CD1a. J. Exp. Med. 2016, 213, 2399–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Cueto, I.; Li, K.; Tian, S.; Brodmerkel, C.; Krueger, J.G. Increased expression of interleukin-17 pathway genes in nonlesional skin of moderate-to-severe psoriasis vulgaris. Br. J. Dermatol. 2016, 174, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisman, S.; Pollack, C.R.; Gottschalk, R.W. Psoriasis disease severity measures: Comparing efficacy of treatments for severe psoriasis. J. Dermatolog. Treat. 2003, 14, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.G.; Elewski, B.E.; Lebwohl, M.; Reich, K.; Griffiths, C.E.; Papp, K.; Puig, L.; Nakagawa, H.; Spelman, L.; Sigurgeirsson, B.; et al. Secukinumab in plaque psoriasis--results of two phase 3 trials. N. Engl. J. Med. 2014, 371, 326–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, K.B.; Blauvelt, A.; Papp, K.A.; Langley, R.G.; Luger, T.; Ohtsuki, M.; Reich, K.; Amato, D.; Ball, S.G.; Braun, D.K.; et al. Phase 3 trials of ixekizumab in moderate-to-severe plaque psoriasis. N. Engl. J. Med. 2016, 375, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Lebwohl, M.; Strober, B.; Menter, A.; Gordon, K.; Weglowska, J.; Puig, L.; Papp, K.; Spelman, L.; Toth, D.; Kerdel, F.; et al. Phase 3 studies comparing brodalumab with ustekinumab in psoriasis. N. Engl. J. Med. 2015, 373, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Paramio, J.M.; Casanova, M.L.; Segrelles, C.; Mittnacht, S.; Lane, E.B.; Jorcano, J.L. Modulation of cell proliferation by cytokeratins K10 and K16. Mol. Cell Biol. 1999, 19, 3086–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, J.G.; Ferris, L.K.; Menter, A.; Wagner, F.; White, A.; Visvanathan, S.; Lalovic, B.; Aslanyan, S.; Wang, E.E.; Hall, D.; et al. Anti-IL-23A mAb BI 655066 for treatment of moderate-to-severe psoriasis: Safety, efficacy, pharmacokinetics, and biomarker results of a single-rising-dose, randomized, double-blind, placebo-controlled trial. J. Allergy Clin. Immunol. 2015, 136, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Kulke, R.; Tödt-Pingel, I.; Rademacher, D.; Röwert, J.; Schröder, J.M.; Christophers, E. Co-localized overexpression of GRO-alpha and IL-8 mRNA is restricted to the suprapapillary layers of psoriatic lesions. J. Investig. Dermatol. 1996, 106, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, K.; Yamamura, K.; Tsuji, G.; Mitoma, C.; Uchi, H.; Nakahara, T.; Kido-Nakahara, M.; Kadono, T.; Furue, M. Highlighting interleukin-36 signalling in plaque psoriasis and pustular psoriasis. Acta Derm. Venereol. 2018, 98, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, A.; Xing, X.; Guzman, A.M.; Riblett, M.; Loyd, C.M.; Ward, N.L.; Wohn, C.; Prens, E.P.; Wang, F.; Maier, L.E.; et al. IL-1F5, -F6, -F8, and -F9: A novel IL-1 family signaling system that is active in psoriasis and promotes keratinocyte antimicrobial peptide expression. J. Immunol. 2011, 186, 2613–2622. [Google Scholar] [CrossRef] [Green Version]
- Homey, B.; Dieu-Nosjean, M.C.; Wiesenborn, A.; Massacrier, C.; Pin, J.J.; Oldham, E.; Catron, D.; Buchanan, M.E.; Müller, A.; de Waal Malefyt, R.; et al. Up-regulation of macrophage inflammatory protein-3 alpha/CCL20 and CC chemokine receptor 6 in psoriasis. J. Immunol. 2000, 164, 6621–6632. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.G.; Jee, H.; Fuentes-Duculan, J.; Wu, W.H.; Byamba, D.; Kim, D.S.; Kim, D.Y.; Lew, D.H.; Yang, W.I.; Krueger, J.G.; et al. Dermal clusters of mature dendritic cells and T cells are associated with the CCL20/CCR6 chemokine system in chronic psoriasis. J. Investig. Dermatol. 2014, 134, 1462–1465. [Google Scholar] [CrossRef] [Green Version]
- Furue, K.; Ito, T.; Tsuji, G.; Nakahara, T.; Furue, M. The CCL20 and CCR6 axis in psoriasis. Scand. J. Immunol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Bando, J.K.; Gilfillan, S.; Song, C.; McDonald, K.G.; Huang, S.C.; Newberry, R.D.; Kobayashi, Y.; Allan, D.S.J.; Carlyle, J.R.; Cella, M.; et al. The tumor necrosis factor superfamily member RANKL suppresses effector cytokine production in group 3 innate lymphoid cells. Immunity 2018, 48, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Talayero, P.; Mancebo, E.; Calvo-Pulido, J.; Rodríguez-Muñoz, S.; Bernardo, I.; Laguna-Goya, R.; Cano-Romero, F.L.; García-Sesma, A.; Loinaz, C.; Jiménez, C.; et al. Innate lymphoid cells groups 1 and 3 in the epithelial compartment of functional human intestinal allografts. Am. J. Transplant. 2016, 16, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Diani, M.; Casciano, F.; Marongiu, L.; Longhi, M.; Altomare, A.; Pigatto, P.D.; Secchiero, P.; Gambari, R.; Banfi, G.; Manfredi, A.A.; et al. Increased frequency of activated CD8(+) T cell effectors in patients with psoriatic arthritis. Sci. Rep. 2019, 9, 10870. [Google Scholar] [CrossRef] [Green Version]
- Steel, K.J.A.; Srenathan, U.; Ridley, M.; Durham, L.E.; Wu, S.Y.; Ryan, S.E.; Hughes, C.D.; Chan, E.; Kirkham, B.W.; Taams, L.S. Synovial IL-17A+ CD8+ T cells display a polyfunctional, pro-inflammatory and tissue-resident memory phenotype and function in psoriatic arthritis. Arthritis Rheumatol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.J.; Ebsworth, K.; Ertl, L.S.; McMahon, J.P.; Newland, D.; Wang, Y.; Liu, S.; Miao, Z.; Dang, T.; Zhang, P.; et al. IL-17-secreting γδ T Cells are completely dependent upon CCR6 for homing to inflamed skin. J. Immunol. 2017, 199, 3129–3136. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Yamamoto, A.; Iizuka, H. Differences in involucrin immunolabeling within cornified cell envelopes in normal and psoriatic epidermis. J. Investig. Dermatol. 1995, 104, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, C.J.; Hobbs, C.; McKee, P.H. The relationship of Ki67 and involucrin expression in proliferative, pre-neoplastic and neoplastic skin. Clin. Exp. Dermatol. 1997, 22, 11–16. [Google Scholar] [CrossRef]
- Krueger, J.G.; Fretzin, S.; Suárez-Fariñas, M.; Haslett, P.A.; Phipps, K.M.; Cameron, G.S.; McColm, J.; Katcherian, A.; Cueto, I.; White, T.; et al. IL-17A is essential for cell activation and inflammatory gene circuits in subjects with psoriasis. J. Allergy Clin. Immunol. 2012, 130, 145–154.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiricozzi, A.; Nograles, K.E.; Johnson-Huang, L.M.; Fuentes-Duculan, J.; Cardinale, I.; Bonifacio, K.M.; Gulati, N.; Mitsui, H.; Guttman-Yassky, E.; Suárez-Fariñas, M.; et al. IL-17 induces an expanded range of downstream genes in reconstituted human epidermis model. PLoS ONE 2014, 9, e90284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.L.; Wang, H.; Pisitkun, P.; Kim, J.C.; Tassi, I.; Tang, W.; Morasso, M.I.; Udey, M.C.; Siebenlist, U. IL-17 drives psoriatic inflammation via distinct, target cell-specific mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, E3422–E3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.Y.; Jia, K.; Zhang, Y. IL-17 promotes keratinocyte proliferation via the downregulation of C/EBPα. Exp. Ther. Med. 2016, 11, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontol. 2000. 2015, 69, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Fariñas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- House, J.S.; Zhu, S.; Ranjan, R.; Linder, K.; Smart, R.C. C/EBPalpha and C/EBPbeta are required for Sebocyte differentiation and stratified squamous differentiation in adult mouse skin. PLoS ONE 2010, 5, e9837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crish, J.F.; Gopalakrishnan, R.; Bone, F.; Gilliam, A.C.; Eckert, R.L. The distal and proximal regulatory regions of the involucrin gene promoter have distinct functions and are required for in vivo involucrin expression. J. Investig. Dermatol. 2006, 126, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, E.; Kokolakis, G.; Witte, K.; Philipp, S.; Doecke, W.D.; Babel, N.; Wittig, B.M.; Warszawska, K.; Kurek, A.; Erdmann-Keding, M.; et al. IL-19 is a component of the pathogenetic IL-23/IL-17 cascade in psoriasis. J. Investig. Dermatol. 2014, 134, 2757–2767. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.P.; Yeh, C.H.; So, E.; Wang, L.Y.; Wei, T.S.; Chang, M.S.; Hsing, C.H. Interleukin (IL)-19 promoted skin wound healing by increasing fibroblast keratinocyte growth factor expression. Cytokine 2013, 62, 360–368. [Google Scholar] [CrossRef]
- Yang, L.; Fan, X.; Cui, T.; Dang, E.; Wang, G. Nrf2 promotes keratinocyte proliferation in psoriasis through up-regulation of keratin 6, keratin 16, and keratin 17. J. Investig. Dermatol. 2017, 137, 2168–2176. [Google Scholar] [CrossRef] [Green Version]
- Paladini, R.D.; Coulombe, P.A. Directed expression of keratin 16 to the progenitor basal cells of transgenic mouse skin delays skin maturation. J. Cell Biol. 1998, 142, 1035–1051. [Google Scholar] [CrossRef] [Green Version]
- Acciani, T.H.; Suzuki, T.; Trapnell, B.C.; Le Cras, T.D. Epidermal growth factor receptor signalling regulates granulocyte-macrophage colony-stimulating factor production by airway epithelial cells and established allergic airway disease. Clin. Exp. Allergy 2016, 46, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, K.; Ito, T.; Tanaka, Y.; Hashimoto-Hachiya, A.; Takemura, M.; Murata, M.; Kido-Nakahara, M.; Tsuji, G.; Nakahara, T.; Furue, M. The EGFR-ERK/JNK-CCL20 pathway in scratched keratinocytes may underpin koebnerization in psoriasis atients. Int. J. Mol. Sci. 2020, 21, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, A.; Gudjonsson, J.E.; Aphale, A.; Guzman, A.M.; Stoll, S.W.; Elder, J.T. EGFR and IL-1 signaling synergistically promote keratinocyte antimicrobial defenses in a differentiation-dependent manner. J. Investig. Dermatol. 2011, 131, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goepel, L.; Jacobi, A.; Augustin, M.; Radtke, M.A. Rapid improvement of psoriasis in a patient with lung cancer after treatment with erlotinib. J. Eur. Acad. Dermatol. Venereol. 2018, 32, e311–e313. [Google Scholar] [CrossRef] [PubMed]
- Overbeck, T.R.; Griesinger, F. Two cases of psoriasis responding to erlotinib: Time to revisiting inhibition of epidermal growth factor receptor in psoriasis therapy? Dermatology 2012, 225, 179–182. [Google Scholar] [CrossRef]
- Trivin, F.; Boucher, E.; Raoul, J.L. Complete sustained regression of extensive psoriasis with cetuximab combination chemotherapy. Acta Oncol. 2004, 43, 592–593. [Google Scholar] [CrossRef]
- Neyns, B.; Meert, V.; Vandenbroucke, F. Cetuximab treatment in a patient with metastatic colorectal cancer and psoriasis. Curr. Oncol. 2008, 15, 196–197. [Google Scholar] [CrossRef] [Green Version]
- Carrier, Y.; Ma, H.L.; Ramon, H.E.; Napierata, L.; Small, C.; O’Toole, M.; Young, D.A.; Fouser, L.A.; Nickerson-Nutter, C.; Collins, M.; et al. Inter-regulation of Th17 cytokines and the IL-36 cytokines in vitro and in vivo: Implications in psoriasis pathogenesis. J. Investig. Dermatol. 2011, 131, 2428–2437. [Google Scholar] [CrossRef] [Green Version]
- Gläser, R.; Meyer-Hoffert, U.; Harder, J.; Cordes, J.; Wittersheim, M.; Kobliakova, J.; Fölster-Holst, R.; Proksch, E.; Schröder, J.M.; Schwarz, T. The antimicrobial protein psoriasin (S100A7) is upregulated in atopic dermatitis and after experimental skin barrier disruption. J. Investig. Dermatol. 2009, 129, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Nograles, K.E.; Zaba, L.C.; Guttman-Yassky, E.; Fuentes-Duculan, J.; Suárez-Fariñas, M.; Cardinale, I.; Khatcherian, A.; Gonzalez, J.; Pierson, K.C.; White, T.R.; et al. Th17 cytokines interleukin (IL)-17 and IL-22 modulate distinct inflammatory and keratinocyte-response pathways. Br. J. Dermatol. 2008, 159, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Baggiolini, M. Chemokines in pathology and medicine. J. Intern. Med. 2001, 250, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.; Xing, X.; Wolterink, L.; Barnes, D.H.; Yin, Z.; Reingold, L.; Kahlenberg, J.M.; Harms, P.W.; Gudjonsson, J.E. IL-1 and IL-36 are dominant cytokines in generalized pustular psoriasis. J. Allergy Clin. Immunol. 2017, 140, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Postepy. Dermatol. Alergol. 2014, 31, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yamasaki, K.; Saito, R.; Fukushi-Takahashi, S.; Shimada-Omori, R.; Asano, M.; Aiba, S. Alarmin function of cathelicidin antimicrobial peptide LL37 through IL-36γ induction in human epidermal keratinocytes. J. Immunol. 2014, 193, 5140–5148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.M.; Sullivan, G.P.; Clancy, D.M.; Afonina, I.S.; Kulms, D.; Martin, S.J. Neutrophil-derived proteases escalate inflammation through activation of IL-36 family cytokines. Cell Rep. 2016, 14, 708–722. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, C.M.; Marquardt, Y.; Fietkau, K.; Baron, J.M.; Lüscher, B. The psoriasis-associated IL-17A induces and cooperates with IL-36 cytokines to control keratinocyte differentiation and function. Sci. Rep. 2017, 7, 15631. [Google Scholar] [CrossRef]
- Muhr, P.; Zeitvogel, J.; Heitland, I.; Werfel, T.; Wittmann, M. Expression of interleukin (IL)-1 family members upon stimulation with IL-17 differs in keratinocytes derived from patients with psoriasis and healthy donors. Br. J. Dermatol. 2011, 165, 189–193. [Google Scholar] [CrossRef]
- Gabay, C.; Towne, J.E. Regulation and function of interleukin-36 cytokines in homeostasis and pathological conditions. J. Leukoc. Biol. 2015, 97, 645–652. [Google Scholar] [CrossRef]
- Madonna, S.; Girolomoni, G.; Dinarello, C.A.; Albanesi, C. The significance of IL-36 hyperactivation and IL-36R targeting in psoriasis. Int. J. Mol. Sci. 2019, 20, 3318. [Google Scholar] [CrossRef] [Green Version]
- Bachelez, H.; Choon, S.E.; Marrakchi, S.; Burden, A.D.; Tsai, T.F.; Morita, A.; Turki, H.; Hall, D.B.; Shear, M.; Baum, P.; et al. Inhibition of the interleukin-36 pathway for the treatment of generalized pustular psoriasis. N. Engl. J. Med. 2019, 380, 981–983. [Google Scholar] [CrossRef]
- Furue, K.; Ito, T.; Tanaka, Y.; Yumine, A.; Hashimoto-Hachiya, A.; Takemura, M.; Murata, M.; Yamamura, K.; Tsuji, G.; Furue, M. Cyto/chemokine profile of in vitro scratched keratinocyte model: Implications of significant upregulation of CCL20, CXCL8 and IL36G in Koebner phenomenon. J. Dermatol. Sci. 2019, 94, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maecker, H.T.; McCoy, J.P.; Nussenblatt, R. Standardizing immunophenotyping for the Human Immunology Project. Nat. Rev. Immunol. 2012, 12, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P.; Zhang, H.H.; Foley, J.F.; Hedrick, M.N.; Farber, J.M. Human T cells that are able to produce IL-17 express the chemokine receptor CCR6. J. Immunol. 2008, 180, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Harper, E.G.; Guo, C.; Rizzo, H.; Lillis, J.V.; Kurtz, S.E.; Skorcheva, I.; Purdy, D.; Fitch, E.; Iordanov, M.; Blauvelt, A. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: Implications for psoriasis pathogenesis. J. Investig. Dermatol. 2009, 129, 2175–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouma, G.; Zamuner, S.; Hicks, K.; Want, A.; Oliveira, J.; Choudhury, A.; Brett, S.; Robertson, D.; Felton, L.; Norris, V.; et al. CCL20 neutralization by a monoclonal antibody in healthy subjects selectively inhibits recruitment of CCR6(+) cells in an experimental suction blister. Br. J. Clin. Pharmacol. 2017, 83, 1976–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, R.; Ang, C.; Sun, G.; Juglair, L.; Lim, E.X.; Mason, L.J.; Payne, N.L.; Bernard, C.C.; Mackay, C.R. Essential role for CCR6 in certain inflammatory diseases demonstrated using specific antagonist and knockin mice. JCI. Insight 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Tawaraishi, T.; Sakauchi, N.; Hidaka, K.; Yoshikawa, K.; Okui, T.; Kuno, H.; Chisaki, I.; Aso, K. Identification of a novel series of potent and selective CCR6 inhibitors as biological probes. Bioorg. Med. Chem. Lett. 2018, 28, 3067–3072. [Google Scholar] [CrossRef]
- Campbell, J.J.; Ebsworth, K.; Ertl, L.S.; McMahon, J.P.; Wang, Y.; Yau, S.; Mali, V.R.; Chhina, V.; Kumamoto, A.; Liu, S.; et al. Efficacy of chemokine receptor inhibition in treating IL-36α-induced psoriasiform inflammation. J. Immunol. 2019, 202, 1687–1692. [Google Scholar] [CrossRef] [Green Version]
- Mose, M.; Kang, Z.; Raaby, L.; Iversen, L.; Johansen, C. TNFα- and IL-17A-mediated S100A8 expression is regulated by p38 MAPK. Exp. Dermatol. 2013, 22, 476–481. [Google Scholar] [CrossRef]
- Liang, S.C.; Tan, X.Y.; Luxenberg, D.P.; Karim, R.; Dunussi-Joannopoulos, K.; Collins, M.; Fouser, L.A. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J. Exp. Med. 2006, 203, 2271–2279. [Google Scholar] [CrossRef]
- D’Amico, F.; Skarmoutsou, E.; Granata, M.; Trovato, C.; Rossi, G.A.; Mazzarino, M.C. S100A7: A rAMPing up AMP molecule in psoriasis. Cytokine Growth Factor Rev. 2016, 32, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Sa, S.M.; Valdez, P.A.; Wu, J.; Jung, K.; Zhong, F.; Hall, L.; Kasman, I.; Winer, J.; Modrusan, Z.; Danilenko, D.M.; et al. The effects of IL-20 subfamily cytokines on reconstituted human epidermis suggest potential roles in cutaneous innate defense and pathogenic adaptive immunity in psoriasis. J. Immunol. 2007, 178, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, Z.; Zwicker, S.; Bureik, D.; Peric, M.; Koglin, S.; Batycka-Baran, A.; Prinz, J.C.; Ruzicka, T.; Schauber, J.; Wolf, R. Vitamin D analog calcipotriol suppresses the Th17 cytokine-induced proinflammatory S100 “alarmins” psoriasin (S100A7) and koebnerisin (S100A15) in psoriasis. J. Investig. Dermatol. 2012, 132, 1416–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.; Champaiboon, C.; Guenther, B.D.; Sorenson, B.S.; Khammanivong, A.; Ross, K.F.; Geczy, C.L.; Herzberg, M.C. Anti-infective protective properties of S100 calgranulins. Antiinflamm. Antiallergy Agents Med. Chem. 2009, 8, 290–305. [Google Scholar] [CrossRef] [Green Version]
- Kolbinger, F.; Loesche, C.; Valentin, M.A.; Jiang, X.; Cheng, Y.; Jarvis, P.; Peters, T.; Calonder, C.; Bruin, G.; Polus, F.; et al. β-Defensin 2 is a responsive biomarker of IL-17A-driven skin pathology in patients with psoriasis. J. Allergy Clin. Immunol. 2017, 139, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.X.; Armishaw, C.; Goyette, J.; Yang, Z.; Cai, H.; Alewood, P.; Geczy, C.L. Mast cell and monocyte recruitment by S100A12 and its hinge domain. J. Biol. Chem. 2008, 283, 13035–13043. [Google Scholar] [CrossRef] [Green Version]
- Nukui, T.; Ehama, R.; Sakaguchi, M.; Sonegawa, H.; Katagiri, C.; Hibino, T.; Huh, N.H. S100A8/A9, a key mediator for positive feedback growth stimulation of normal human keratinocytes. J. Cell Biochem. 2008, 104, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Muta, T. IkappaB-zeta: An inducible regulator of nuclear factor-kappaB. Vitam. Horm. 2006, 74, 301–316. [Google Scholar]
- Bertelsen, T.; Iversen, L.; Johansen, C. The human IL-17A/F heterodimer regulates psoriasis-associated genes through IκBζ. Exp. Dermatol. 2018, 27, 1048–1052. [Google Scholar] [CrossRef]
- Muromoto, R.; Hirao, T.; Tawa, K.; Hirashima, K.; Kon, S.; Kitai, Y.; Matsuda, T. IL-17A plays a central role in the expression of psoriasis signature genes through the induction of IκB-ζ in keratinocytes. Int. Immunol. 2016, 28, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Dorschner, R.A.; Pestonjamasp, V.K.; Tamakuwala, S.; Ohtake, T.; Rudisill, J.; Nizet, V.; Agerberth, B.; Gudmundsson, G.H.; Gallo, R.L. Cutaneous injury induces the release of cathelicidin anti-microbial peptides active against group A Streptococcus. J. Investig. Dermatol. 2001, 117, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinholz, M.; Ruzicka, T.; Schauber, J. Cathelicidin LL-37: An antimicrobial peptide with a role in inflammatory skin disease. Ann. Dermatol. 2012, 24, 126–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasinsig, L.; Pizzirani, C.; Skerlavaj, B.; Pellegatti, P.; Gulinelli, S.; Tossi, A.; Di Virgilio, F.; Zanetti, M. The human cathelicidin LL-37 modulates the activities of the P2X7 receptor in a structure-dependent manner. J. Biol. Chem. 2008, 283, 30471–30481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Nardo, A.; Braff, M.H.; Taylor, K.R.; Na, C.; Granstein, R.D.; McInturff, J.E.; Krutzik, S.; Modlin, R.L.; Gallo, R.L. Cathelicidin antimicrobial peptides block dendritic cell TLR4 activation and allergic contact sensitization. J. Immunol. 2007, 178, 1829–1834. [Google Scholar] [CrossRef] [Green Version]
- Tokumaru, S.; Sayama, K.; Shirakata, Y.; Komatsuzawa, H.; Ouhara, K.; Hanakawa, Y.; Yahata, Y.; Dai, X.; Tohyama, M.; Nagai, H.; et al. Induction of keratinocyte migration via transactivation of the epidermal growth factor receptor by the antimicrobial peptide LL-37. J. Immunol. 2005, 175, 4662–4668. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.C.; Yu, H.S.; Yen, F.L.; Lin, C.L.; Chen, G.S.; Lan, C.C. Neutrophil extracellular trap formation is increased in psoriasis and induces human β-defensin-2 production in epidermal keratinocytes. Sci. Rep. 2016, 6, 31119. [Google Scholar] [CrossRef] [Green Version]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef]
- Mabuchi, T.; Hirayama, N. Binding affinity and interaction of LL-37 with HLA-C*06:02 in psoriasis. J. Investig. Dermatol. 2016, 136, 1901–1903. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Kulkarni, N.N.; Lee, E.Y.; Zhang, L.J.; Wong, G.C.L.; Gallo, R.L. Cathelicidin promotes inflammation by enabling binding of self-RNA to cell surface scavenger receptors. Sci. Rep. 2018, 8, 4032. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- de Jongh, G.J.; Zeeuwen, P.L.; Kucharekova, M.; Pfundt, R.; van der Valk, P.G.; Blokx, W.; Dogan, A.; Hiemstra, P.S.; van de Kerkhof, P.C.; Schalkwijk, J. High expression levels of keratinocyte antimicrobial proteins in psoriasis compared with atopic dermatitis. J. Investig. Dermatol. 2005, 125, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Nomura, I.; Goleva, E.; Howell, M.D.; Hamid, Q.A.; Ong, P.Y.; Hall, C.F.; Darst, M.A.; Gao, B.; Boguniewicz, M.; Travers, J.B.; et al. Cytokine milieu of atopic dermatitis, as compared to psoriasis, skin prevents induction of innate immune response genes. J. Immunol. 2003, 171, 3262–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; et al. Beta-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Ghannam, S.; Dejou, C.; Pedretti, N.; Giot, J.P.; Dorgham, K.; Boukhaddaoui, H.; Deleuze, V.; Bernard, F.X.; Jorgensen, C.; Yssel, H.; et al. CCL20 and β-defensin-2 induce arrest of human Th17 cells on inflamed endothelium in vitro under flow conditions. J. Immunol. 2011, 186, 1411–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Simplified effects of anti-interleukin 17A (IL-17A) on keratinocyte (KC) with regard to psoriasis pathogenesis. IL-17A homodimers bind to IL-17 receptor A (IL-17RA) and IL-17RC or IL-17RA and IL-17RD heterodimers. The ligation of IL-17RA/IL-17RC activates epidermal growth factor receptor (EGFR) directly or by transforming growth factor-α (TGF-α) and heparin-binding EGF-like growth factor (HB-EGF) and promotes keratinocyte proliferation. The ligation of IL-17RA/IL-17RC activates various signal transduction molecules, including ERK, p38 MAPK, JNK, nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB), IκBζ, C/CAAT-enhancer-binding protein β (C/EBPβ), and C/EBPδ. In contrast, the ligation of IL-17RA/IL-17RD preferentially activates JNK and p38 MAPK pathways. IL-17RA/IL-17RD is estimated to transactivate fibroblast growth factor receptor (FGFR); however, this is not conclusive. IL-17RA/IL-17RC signaling stimulates KCs to produce IL-19, which induces the production of keratinocyte growth factor (KGF) from fibroblasts. KGF also enhances the proliferation of KCs. IL-17A also induces the production of antimicrobial peptides, including S100A7, S100A8, S100A9, LL-37, and defensin β 4A (DEFB4A). These antimicrobial peptides amplify the local inflammatory process. Chemokines, such as CCL20, CXCL1, and CXCL8, are also produced from keratinocytes by IL-17RA/IL-17RC ligation. CCL20 is a key chemokine for the recruitment of CCR6+ Th17 cells and group 3 innate lymphoid cells (ILC3). These CCR6+ cells produce large amounts of IL-17A. DEFB4A also exhibits a chemotactic activity by binding to CCR6. CXCL1 and CXCL2 are potent chemoattractants for CXCR2+ neutrophils. Therefore, IL-17A is associated with all of the histopathologic features of psoriasis.
Figure 1.
Simplified effects of anti-interleukin 17A (IL-17A) on keratinocyte (KC) with regard to psoriasis pathogenesis. IL-17A homodimers bind to IL-17 receptor A (IL-17RA) and IL-17RC or IL-17RA and IL-17RD heterodimers. The ligation of IL-17RA/IL-17RC activates epidermal growth factor receptor (EGFR) directly or by transforming growth factor-α (TGF-α) and heparin-binding EGF-like growth factor (HB-EGF) and promotes keratinocyte proliferation. The ligation of IL-17RA/IL-17RC activates various signal transduction molecules, including ERK, p38 MAPK, JNK, nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB), IκBζ, C/CAAT-enhancer-binding protein β (C/EBPβ), and C/EBPδ. In contrast, the ligation of IL-17RA/IL-17RD preferentially activates JNK and p38 MAPK pathways. IL-17RA/IL-17RD is estimated to transactivate fibroblast growth factor receptor (FGFR); however, this is not conclusive. IL-17RA/IL-17RC signaling stimulates KCs to produce IL-19, which induces the production of keratinocyte growth factor (KGF) from fibroblasts. KGF also enhances the proliferation of KCs. IL-17A also induces the production of antimicrobial peptides, including S100A7, S100A8, S100A9, LL-37, and defensin β 4A (DEFB4A). These antimicrobial peptides amplify the local inflammatory process. Chemokines, such as CCL20, CXCL1, and CXCL8, are also produced from keratinocytes by IL-17RA/IL-17RC ligation. CCL20 is a key chemokine for the recruitment of CCR6+ Th17 cells and group 3 innate lymphoid cells (ILC3). These CCR6+ cells produce large amounts of IL-17A. DEFB4A also exhibits a chemotactic activity by binding to CCR6. CXCL1 and CXCL2 are potent chemoattractants for CXCR2+ neutrophils. Therefore, IL-17A is associated with all of the histopathologic features of psoriasis.

Figure 2.
Pivotal role of signal transducer and activator of transcription 3 (STAT3) in psoriasis. The activation of STAT3 promotes keratinocyte (KC) proliferation and inflammatory response. IL-17A and IL-22 induce the STAT3 activation. IL-6 produced from KC also induces STAT3 activation.
Figure 2.
Pivotal role of signal transducer and activator of transcription 3 (STAT3) in psoriasis. The activation of STAT3 promotes keratinocyte (KC) proliferation and inflammatory response. IL-17A and IL-22 induce the STAT3 activation. IL-6 produced from KC also induces STAT3 activation.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Furue, M.; Furue, K.; Tsuji, G.; Nakahara, T. Interleukin-17A and Keratinocytes in Psoriasis. Int. J. Mol. Sci. 2020, 21, 1275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041275
AMA Style
Furue M, Furue K, Tsuji G, Nakahara T. Interleukin-17A and Keratinocytes in Psoriasis. International Journal of Molecular Sciences. 2020; 21(4):1275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041275
Chicago/Turabian StyleFurue, Masutaka, Kazuhisa Furue, Gaku Tsuji, and Takeshi Nakahara. 2020. "Interleukin-17A and Keratinocytes in Psoriasis" International Journal of Molecular Sciences 21, no. 4: 1275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041275
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.