Distinct Metabolome Changes during Seed Germination of Lettuce (Lactuca sativa L.) in Response to Thermal Stress as Revealed by Untargeted Metabolomics Analysis


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Country of Origin, Seed Coat Color and Seed Weight of the Lettuce Germplasm Used in The Current Study
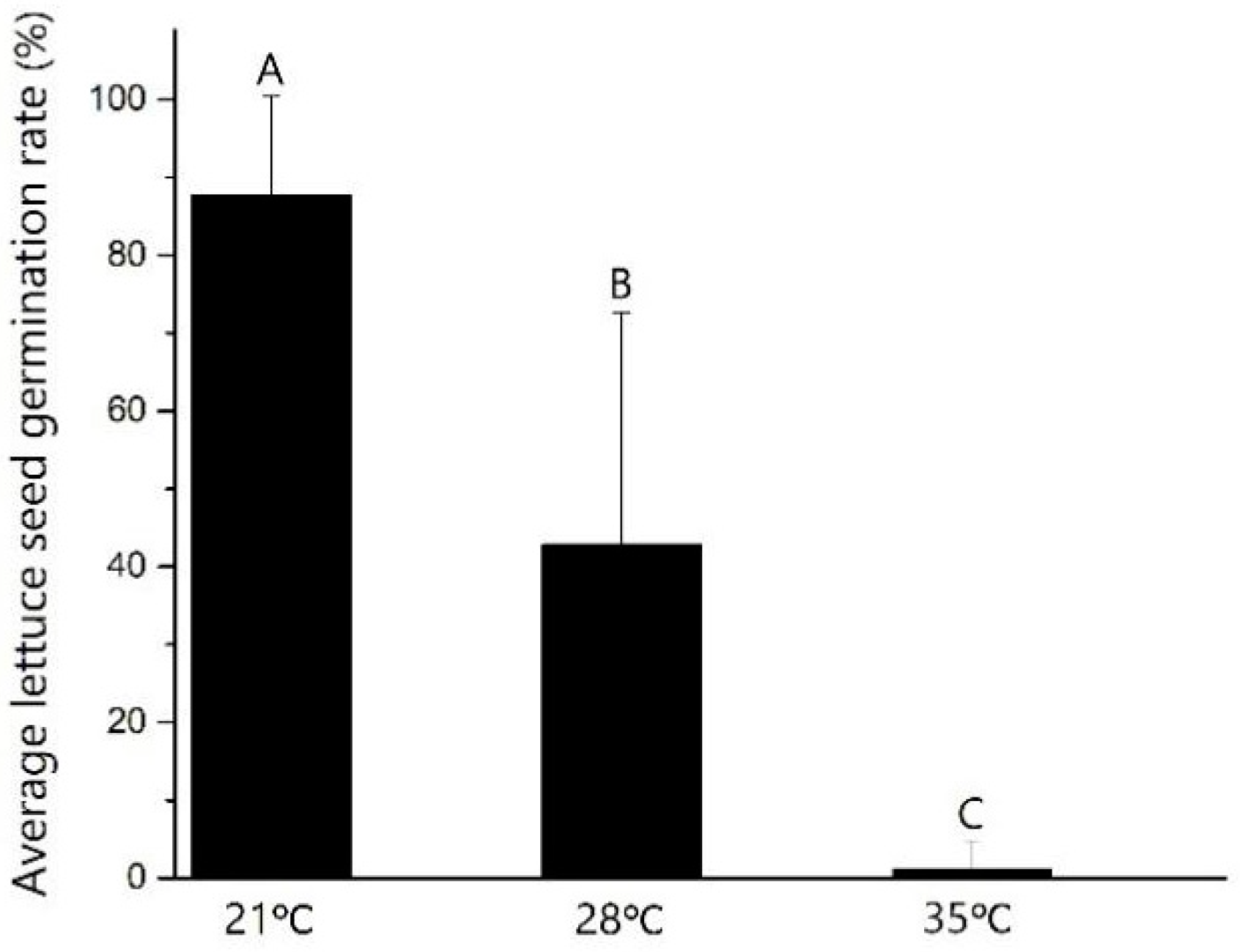
2.2. Evaluation of Seed Germination under Different Temperatures
2.3. The Seed Color, Weight and Origin Region Influence on Germination Rates of Lettuce under Heat Stress
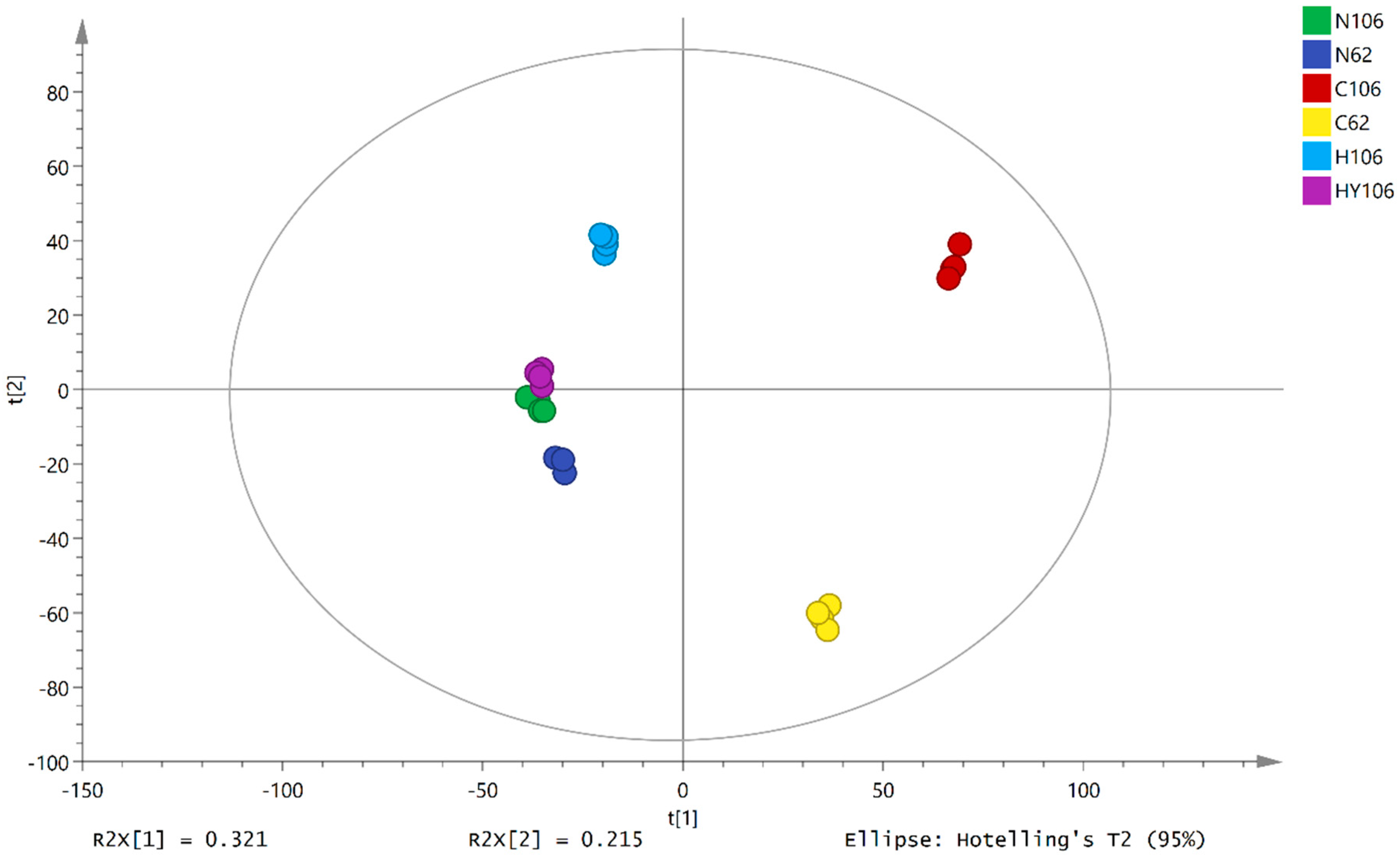
2.4. Metabolic Profiles of Lettuce Seeds
2.5. Identification of Differential Metabolites
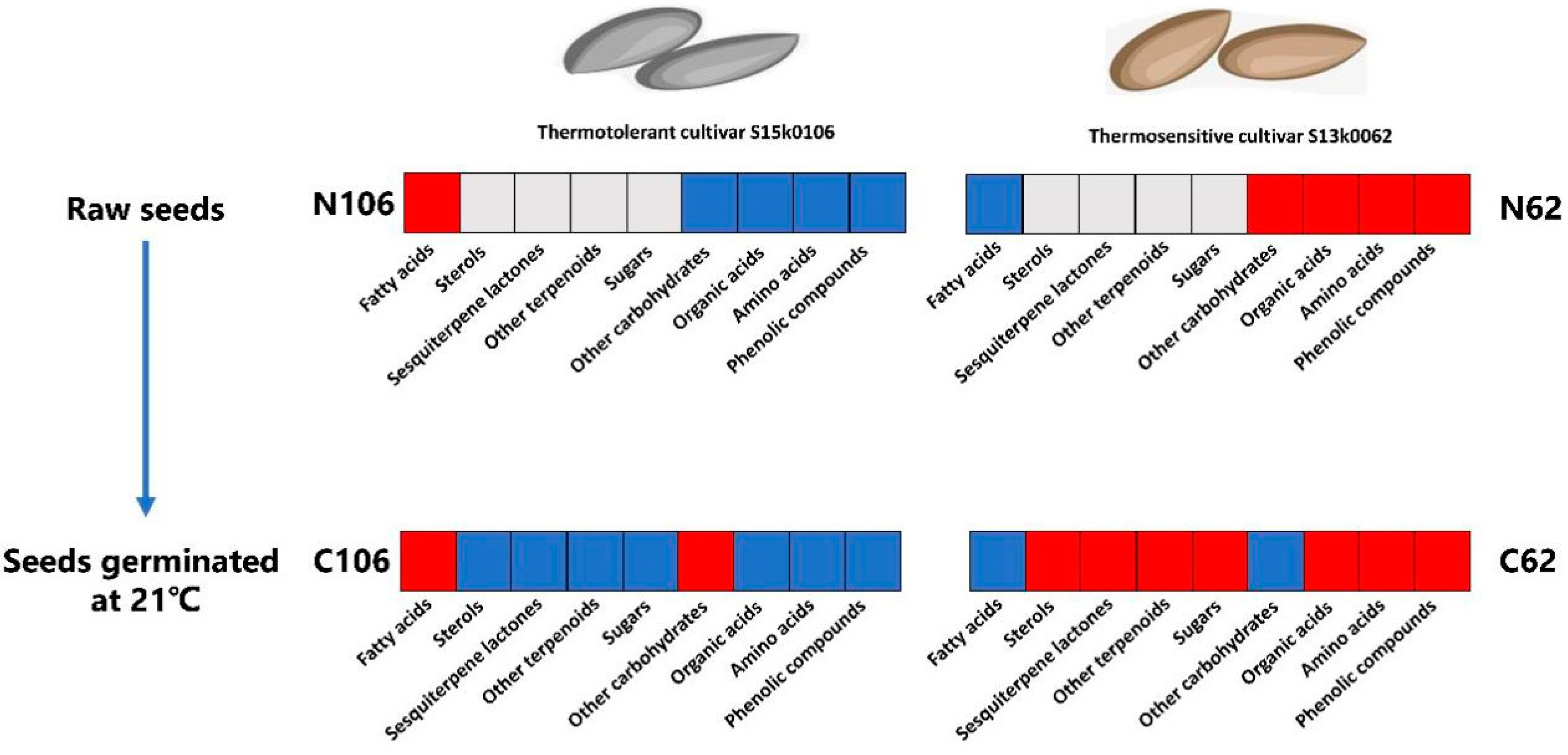
2.6. Metabolite Differences between Thermo-Tolerant and Thermo-Sensitive Seeds
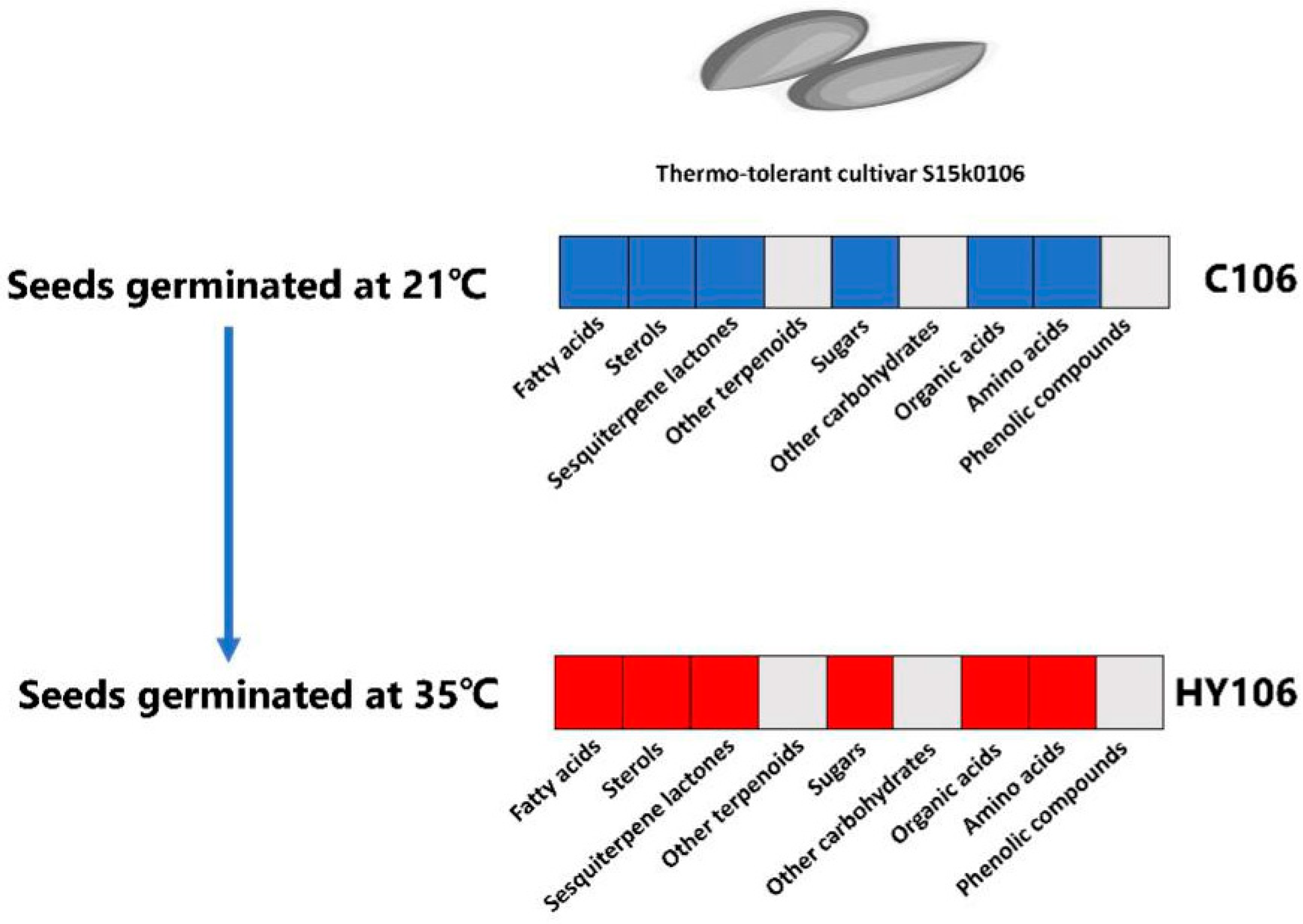
2.7. Metabolite Differences between Thermotolerant Seeds Germinated at High Temperature and Optimal Temperature
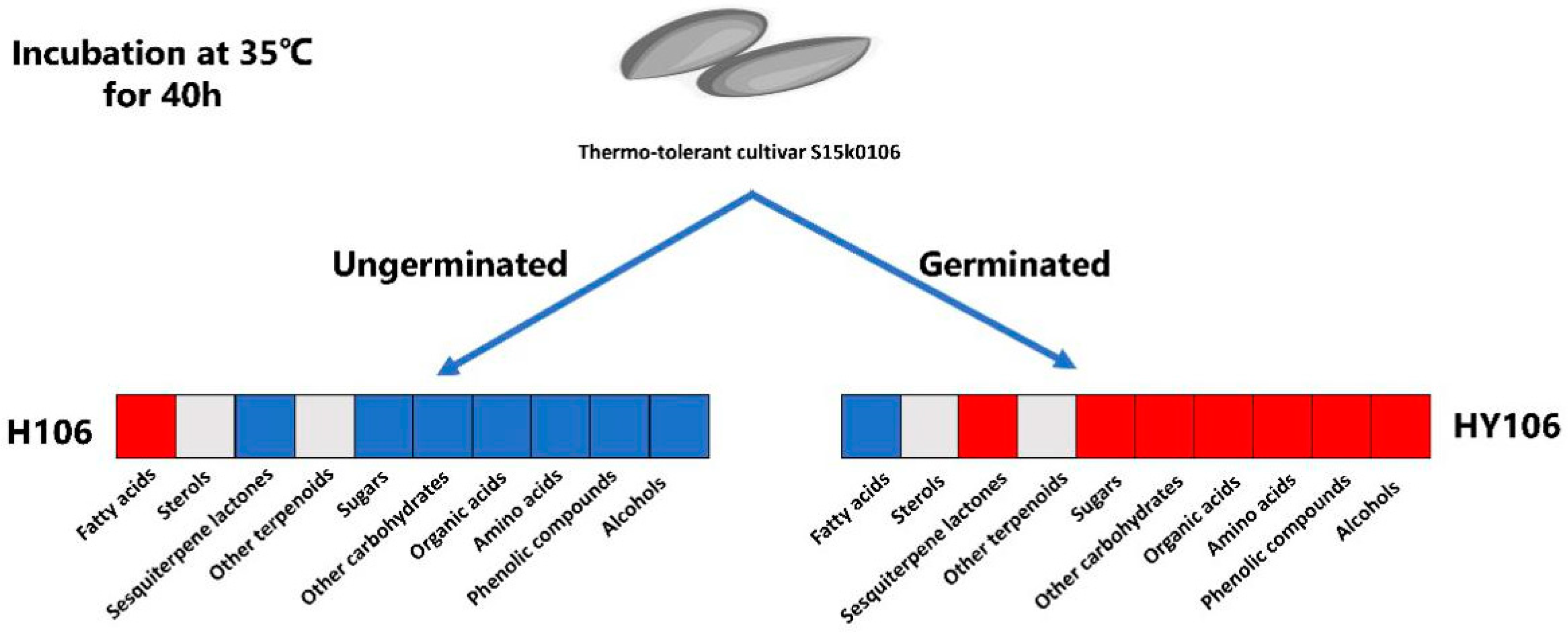
2.8. Metabolite Differences between Non-Germinated Seeds and Germinated Buds of Thermotolerant Cultivar at High Temperature
3. Discussion
3.1. Germination of Lettuce Germplasms is Inhibited by High Temperature
3.2. Metabolic Differences between Thermal-Sensitive and Thermal-Tolerant Cultivars during Germination at Proper Temperature
3.3. Metabolite Requirements for Thermo-Tolerant Seeds Germination under Thermal Stress
4. Materials and Methods
4.1. Evaluate Seed Germination under Different Temperature
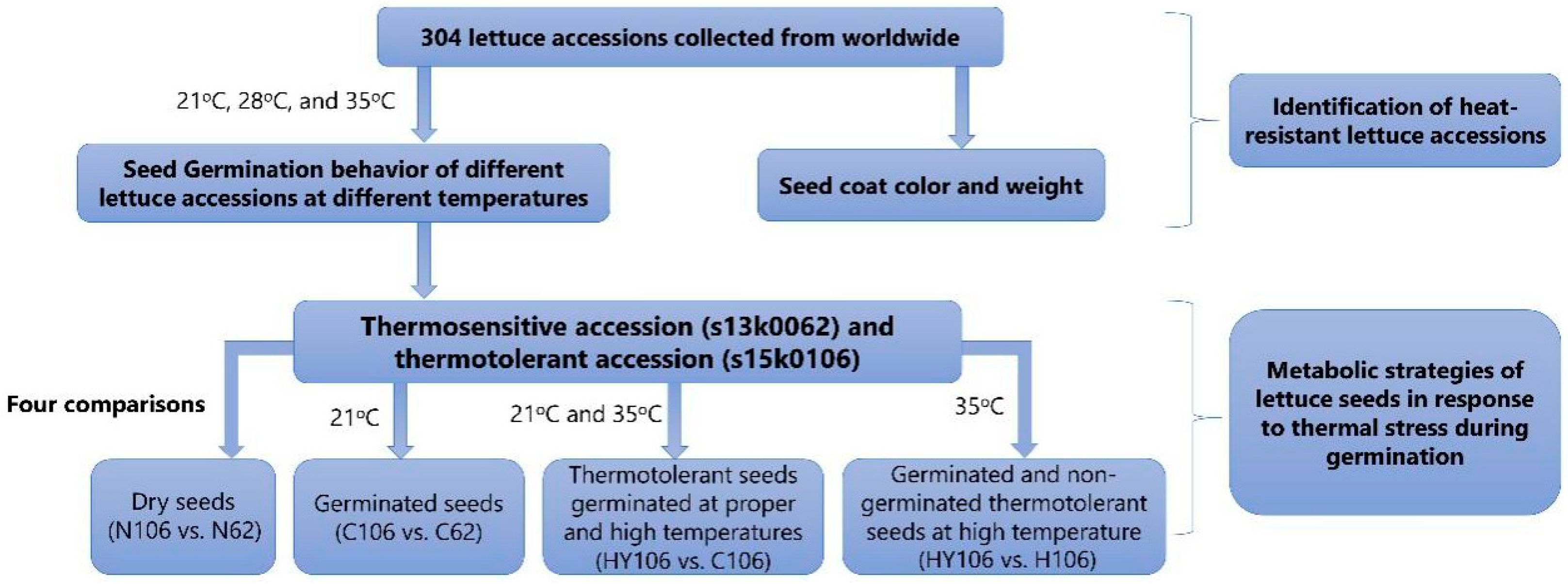
4.2. Two Accessions Are Selected for Metabolic Profiling
4.3. Metabolite Profiling
4.3.1. GC-MS Analysis
4.3.2. UPLC-IMS-QTOF/MS Analysis
4.4. Data Pre-Processing and Normalization
4.5. Data Analysis
4.5.1. Study Design
4.5.2. Univariate and Multivariate Statistical Analysis
4.5.3. Differential Metabolites Screening
4.5.4. Compound Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Quality control | QC |
| Mass spectrometry | MS |
| Shanghai Agricultural Gene Center | SAGC |
| Gas chromatography-mass spectrometry | GC/MS |
| Ultra-performance liquid chromatography-ion mobility spectrometry quadrupole time-of-flight mass spectrometry | UPLC-IMS-QTOF/MS |
| Principal component analysis | PCA |
| Partial least squares-discriminant analysis | PLS-DA |
| Tricarboxylic acid cycle | TCA |
| Variable importance in projection | VIP |
References
- Abeles, F.B.; Lonski, J. Stimulation of lettuce seed germination by ethylene. Plant Physiol. 1969, 44, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.Q.; Song, B.Y.; Deng, Z.J.; Wang, Y.; Liu, S.J.; Moller, I.M.; Song, S.Q. Proteomic analysis of lettuce seed germination and thermoinhibition by sampling of individual seeds at germination and removal of storage proteins by polyethylene glycol fractionation. Plant Physiol. 2015, 167, 1332–1350. [Google Scholar] [CrossRef]
- Huo, H.; Dahal, P.; Kunusoth, K.; McCallum, C.M.; Bradford, K.J. Expression of 9-cis-epoxycarotenoid dioxygenase4 is essential for thermoinhibition of lettuce seed germination but not for seed development or stress tolerance. Plant Cell 2013, 25, 884–900. [Google Scholar] [CrossRef] [Green Version]
- Lafta, A.; Mou, B. Evaluation of lettuce genotypes for seed thermotolerance. Hortscience 2013, 48, 708–714. [Google Scholar] [CrossRef]
- Nascimento, W.M.; Cantliffe, D.J.; Huber, D.J. Thermotolerance in lettuce seeds: Association with ethylene and endo-β-mannanase. J. Am. Soc. Hortic. Sci. 2000, 125, 518–524. [Google Scholar] [CrossRef]
- Contreras, S.; Bennett, M.A.; Metzger, J.D.; Tay, D.; Nerson, H. Red to far-red ratio during seed development affects lettuce seed germinability and longevity. HortScience 2009, 44, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Borthwick, H.A.; Hendricks, S.B.; Parker, M.W.; Toole, E.H.; Toole, V.K. A Reversible photoreaction controlling seed germination. Proc. Natl. Acad. Sci. USA 1952, 38, 662–666. [Google Scholar] [CrossRef] [Green Version]
- Toyomasu, T.; Kawaide, H.; Mitsuhashi, W.; Inoue, Y.; Kamiya, Y. Phytochrome regulates gibberellin biosynthesis during germination of photoblastic lettuce seeds. Plant Physiol. 1998, 118, 1517–1523. [Google Scholar] [CrossRef] [Green Version]
- Argyris, J.; Dahal, P.; Hayashi, E.; Still, D.W.; Bradford, K.J. Genetic variation for lettuce seed thermoinhibition is associated with temperature-sensitive expression of abscisic acid, gibberellin, and ethylene biosynthesis, metabolism, and response genes. Plant Physiol. 2008, 148, 926–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoong, F.Y.; O’Brien, L.K.; Truco, M.J.; Huo, H.; Sideman, R.; Hayes, R.; Michelmore, R.W.; Bradford, K.J. Genetic variation for thermotolerance in lettuce seed germination is associated with temperature-sensitive regulation of ethylene response factor1 (ERF1). Plant Physiol. 2016, 170, 472–488. [Google Scholar] [CrossRef] [Green Version]
- De Vos, R.C.; Moco, S.; Lommen, A.; Keurentjes, J.J.; Bino, R.J.; Hall, R.D. Untargeted large-scale plant metabolomics using liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2007, 2, 778–791. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics - The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Garcia, C.J.; Garcia-Villalba, R.; Gil, M.I.; Tomas-Barberan, F.A. LC-MS untargeted metabolomics to explain the signal metabolites inducing browning in fresh-cut lettuce. J Agric. Food Chem. 2017, 65, 4526–4535. [Google Scholar] [CrossRef]
- García, C.J.; Gil, M.I.; Tomas-Barberan, F.A. LC–MS untargeted metabolomics reveals early biomarkers to predict browning of fresh-cut lettuce. Postharvest Biol. Technol. 2018, 146, 9–17. [Google Scholar] [CrossRef]
- Yang, X.; Feng, L.; Zhao, L.; Liu, X.; Hassani, D.; Huang, D. Effect of glycine nitrogen on lettuce growth under soilless culture: A metabolomics approach to identify the main changes occurred in plant primary and secondary metabolism. J. Sci. Food Agric. 2018, 98, 467–477. [Google Scholar] [CrossRef]
- Yang, X.; Wei, S.; Liu, B.; Guo, D.; Zheng, B.; Feng, L.; Liu, Y.; Tomas-Barberan, F.A.; Luo, L.; Huang, D. A novel integrated non-targeted metabolomic analysis reveals significant metabolite variations between different lettuce (Lactuca sativa, L.) varieties. Hortic. Res. 2018, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Gonai, T.; Kawahara, S.; Tougou, M.; Satoh, S.; Hashiba, T.; Hirai, N.; Kawaide, H.; Kamiya, Y.; Yoshioka, T. Abscisic acid in the thermoinhibition of lettuce seed germination and enhancement of its catabolism by gibberellin. J. Exp. Bot. 2004, 55, 111–118. [Google Scholar] [CrossRef]
- Gray, D. Effects of temperature on the germination and emergence of lettuce (Lactuca sativa, L.) varieties. J. Hortic. Sci. 1975, 50, 349–361. [Google Scholar] [CrossRef]
- Kozarewa, I.; Cantliffe, D.J.; Nagata, R.T.; Stoffella, P.J. High maturation temperature of lettuce seeds during development increased ethylene production and germination at elevated temperatures. J. Am. Soc. Hortic. Sci. 2006, 131, 564–570. [Google Scholar] [CrossRef] [Green Version]
- FAO, U.N. Lettuce and Chicory Production of 2017. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 1 June 2019).
- China Meteorological Administration. China Climate Bulletin 2018. Available online: http://www.cma.gov.cn/root7/auto13139/201903/t20190319_517664.html (accessed on 1 June 2019).
- NPGS, U.A. Grin Gloval Database. Available online: https://npgsweb.ars-grin.gov/gringlobal/accessiondetail.aspx?id=1718215 (accessed on 14 October 2019).
- Penaloza, P.; Ramirez-Rosales, G.; McDonald, M.B.; Bennett, M.A. Lettuce (Lactuca sativa, L.) seed quality evaluation using seed physical attributes, saturated salt accelerated aging and the seed vigour imaging system. Electron. J. Biotechnol. 2005, 8. [Google Scholar] [CrossRef] [Green Version]
- Hoy, D.J.; Gamble, E.E. The effects of seed size and seed density on germination and vigor in soybean (Glycine max (L.) Merr.). Can. J. Plant Sci. 1985, 65, 1–8. [Google Scholar] [CrossRef]
- Lafond, G.; Baker, R. Effects of genotype and seed size on speed of emergence and seedling vigor in nine spring wheat cultivars 1. Crop Sci. 1986, 26, 341–346. [Google Scholar] [CrossRef]
- Grahn, C.M.; Hellier, B.C.; Benedict, C.; Miles, C.A. Screening USDA lettuce (Lactuca sativa, L.) germplasm for ability to germinate under cold conditions. Hortscience 2015, 50, 1155–1159. [Google Scholar] [CrossRef] [Green Version]
- Lindqvist, K. On the origin of cultivated lettuce. Hereditas 1960, 46, 319–350. [Google Scholar] [CrossRef]
- De Vries, I.M. Origin and domestication of Lactuca sativa L. Genet. Resour. Crop Evol. 1997, 44, 165–174. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Zhang, L.; Su, W.Q.; Tao, R.; Zhang, W.Y.; Chen, J.J.; Wu, P.Y.; Yan, C.H.; Jia, Y.; Larkin, R.M.; Lavelle, D.; et al. RNA sequencing provides insights into the evolution of lettuce and the regulation of flavonoid biosynthesis. Nat. Commun. 2017, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Orozco, R.; Piskula, M.K.; Zielinski, H.; Kozlowska, H.; Frias, J.; Vidal-Valverde, C. Germination as a process to improve the antioxidant capacity of Lupinus angustifolius L. var. Zapaton. Eur. Food Res. Technol. 2006, 223, 495–502. [Google Scholar] [CrossRef]
- Dueñas, M.; Hernández, T.; Estrella, I.; Fernández, D. Germination as a process to increase the polyphenol content and antioxidant activity of lupin seeds (Lupinus angustifolius, L.). Food Chem. 2009, 117, 599–607. [Google Scholar] [CrossRef]
- Guo, X.; Li, T.; Tang, K.; Liu, R.H. Effect of germination on phytochemical profiles and antioxidant activity of mung bean sprouts (Vigna radiata). J. Agric. Food Chem. 2012, 60, 11050–11055. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef] [Green Version]
- Sripriya, G.; Antony, U.; Chandra, T.S. Changes in carbohydrate, free amino acids, organic acids, phytate and HCl extractability of minerals during germination and fermentation of finger millet (Eleusine coracana). Food Chem. 1997, 58, 345–350. [Google Scholar] [CrossRef]
- Wang, W.B.; Kim, Y.H.; Lee, H.S.; Kim, K.Y.; Deng, X.P.; Kwak, S.S. Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 2009, 47, 570–577. [Google Scholar] [CrossRef]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.-Q.; Shi, Z.; Jiang, Z.-P.; Qi, L.-W.; Sun, X.-M.; Li, C.-X.; Liu, J.-F.; Xiao, W.-F.; Zhang, S.-G. Effects of exogenous GABA on gene expression of Caragana intermedia roots under NaCl stress: Regulatory roles for H2O2 and ethylene production. Plant Cell Environ. 2010, 33, 149–162. [Google Scholar] [CrossRef]
- Joshi, V.; Joung, J.-G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef]
- Araujo, W.L.; Tohge, T.; Ishizaki, K.; Leaver, C.J.; Fernie, A.R. Protein degradation—An alternative respiratory substrate for stressed plants. Trends Plant Sci. 2011, 16, 489–498. [Google Scholar] [CrossRef]
- Urano, K.; Maruyama, K.; Ogata, Y.; Morishita, Y.; Takeda, M.; Sakurai, N.; Suzuki, H.; Saito, K.; Shibata, D.; Kobayashi, M.; et al. Characterization of the ABA-regulated global responses to dehydration in Arabidopsis by metabolomics. Plant J. 2009, 57, 1065–1078. [Google Scholar] [CrossRef]
- Pujar, A.; Jaiswal, P.; Kellogg, E.A.; Ilic, K.; Vincent, L.; Avraham, S.; Stevens, P.; Zapata, F.; Reiser, L.; Rhee, S.Y.; et al. Whole-plant growth stage ontology for angiosperms and its application in plant biology. Plant Physiol. 2006, 142, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Gigon, A.; Matos, A.R.; Laffray, D.; Zuily-Fodil, Y.; Pham-Thi, A.T. Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (ecotype Columbia). Ann. Bot. 2004, 94, 345–351. [Google Scholar] [CrossRef]
- Schaller, H. The role of sterols in plant growth and development. Prog. Lipid Res. 2003, 42, 163–175. [Google Scholar] [CrossRef]
- Wang, J.; Juliani, H.R.; Jespersen, D.; Huang, B. Differential profiles of membrane proteins, fatty acids, and sterols associated with genetic variations in heat tolerance for a perennial grass species, hard fescue (Festuca Trachyphylla). Environ. Exp. Bot. 2017, 140, 65–75. [Google Scholar] [CrossRef]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. Biochim. Biophys. Acta Biomembr. 2004, 1666, 62–87. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.; Ursell, T.; Wiggins, P.; Sens, P. Emerging roles for lipids in shaping membrane-protein function. Nature 2009, 459, 379–385. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Log2 (FC N106/N62) | p-Value | VIP Value |
|---|---|---|---|---|
| GC_95 | Lactic acid | −1.90 | 1.70 × 10−5 | 1.26 |
| GC_361 | L-Methionine | −1.12 | 1.70 × 10−5 | 1.26 |
| GC_453 | Asparagine | −1.75 | 3.58 × 10−3 | 1.17 |
| GC_484 | Arabinitol | −1.03 | 3.31 × 10−5 | 1.25 |
| GC_537 | L-Ornithine | −1.19 | 1.73 × 10−5 | 1.26 |
| GC_554 | Quininic acid | −1.01 | 9.85 × 10−5 | 1.25 |
| GC_686 | 9,12-Octadecadienoic acid (Z, Z)- | 1.00 | 2.69 × 10−4 | 1.23 |
| GC_689 | 9-Octadecenoic acid, (E)- | 1.56 | 4.09 × 10−5 | 1.25 |
| GC_692 | Oleic acid, (Z)- | 1.11 | 1.00 × 10−3 | 1.21 |
| GC_786 | Arachidic acid | 1.06 | 9.46 × 10−4 | 1.21 |
| LC_4594 | Caffeoyl-hexose 2 | −1.55 | 1.55 × 10−5 | 1.26 |
| No. | Compound | Log2 (FC C106/C62) | p-Value | VIP Value |
|---|---|---|---|---|
| GC_89 | Pyruvic acid | −1.83 | 4.86 × 10−5 | 1.12 |
| GC_115 | l-Alanine | −2.13 | 5.94 × 10−4 | 1.09 |
| GC_185 | l-Valine | −2.34 | 4.27 × 10−4 | 1.09 |
| GC_200 | 4-Hydroxybutanoic acid | −3.46 | 8.85 × 10−5 | 1.11 |
| GC_233 | l-Isoleucine | −1.83 | 2.10 × 10−5 | 1.13 |
| GC_236 | l-Proline | −5.33 | 6.79 × 10−5 | 1.12 |
| GC_249 | Butanedioic acid | −2.69 | 5.53 × 10−8 | 1.14 |
| GC_258 | Glyceric acid | −1.85 | 2.17 × 10−5 | 1.13 |
| GC_290 | l-Threonine | −2.01 | 2.15 × 10−8 | 1.14 |
| GC_349 | Malic acid | −6.45 | 1.42 × 10−9 | 1.14 |
| GC_359 | meso-Erythritol | 1.56 | 9.35 × 10−6 | 1.13 |
| GC_362 | l-5-Oxoproline | −1.92 | 4.36 × 10−7 | 1.14 |
| GC_364 | l-Aspartic acid | −2.40 | 4.96 × 10−7 | 1.14 |
| GC_368 | 4-Aminobutanoic acid | −1.01 | 2.27 × 10−4 | 1.10 |
| GC_391 | l-Threonic acid | −2.19 | 7.27 × 10−7 | 1.14 |
| GC_425 | l-Ornithine | −5.37 | 1.53 × 10−4 | 1.11 |
| GC_427 | l-Glutamic acid | −2.28 | 1.70 × 10−6 | 1.14 |
| GC_430 | Phenylalanine | −1.27 | 4.23 × 10−8 | 1.14 |
| GC_554 | Quininic acid | −1.88 | 1.27 × 10−6 | 1.14 |
| GC_557 | Fructose | −2.91 | 6.58 × 10−6 | 1.13 |
| GC_569 | Glucose | −1.67 | 3.44 × 10−5 | 1.12 |
| GC_606 | Caffeic acid | −1.25 | 4.12 × 10−6 | 1.13 |
| GC_627 | Palmitic acid | 2.91 | 7.42 × 10−8 | 1.14 |
| GC_689 | 9-Octadecenoic acid, (E)- | 3.21 | 5.83 × 10−6 | 1.13 |
| GC_692 | Oleic acid, (Z)- | −44.26 | 5.41 × 10−15 | 1.14 |
| GC_702 | Stearic acid | 3.77 | 1.55 × 10−8 | 1.14 |
| GC_786 | Arachidic acid | 3.84 | 1.33 × 10−9 | 1.14 |
| GC_874 | 1-Monolinolein | 1.43 | 2.11 × 10−7 | 1.14 |
| GC_897 | Lignoceric acid | 1.17 | 1.18 × 10−4 | 1.11 |
| GC_948 | α-Tocopherol | −1.56 | 1.19 × 10−4 | 1.11 |
| GC_965 | Campesterol | −1.23 | 8.03 × 10−5 | 1.12 |
| GC_968 | Stigmasterol | −1.35 | 6.34 × 10−5 | 1.12 |
| LC_443 | Caffeoylquinic acid | −2.65 | 2.06 × 10−4 | 1.10 |
| LC_1072 | Dihydrocaffeic acid hexose | −3.52 | 6.67 × 10−10 | 1.14 |
| LC_1096 | Dihydroxybenzoic acid | −2.25 | 1.85 × 10−7 | 1.14 |
| LC_1152 | Caffeoyl-hexose 1 | −3.97 | 7.57 × 10−9 | 1.14 |
| LC_1736 | Lactucopicrin | −2.28 | 8.49 × 10−6 | 1.13 |
| LC_1777 | Dihydroxybenzoic acid hexoside | −2.01 | 1.65 × 10−7 | 1.14 |
| LC_4473 | Caffeoylquinic acid hexoside | −3.15 | 9.40 × 10−8 | 1.14 |
| No. | Compound | Log2 (FC HY106/C106) | p-Value | VIP Value |
|---|---|---|---|---|
| GC_89 | Pyruvic acid | 8.18 | 1.16 × 10−7 | 1.13 |
| GC_95 | Lactic acid | 5.11 | 5.76 × 10−8 | 1.13 |
| GC_103 | Glycolic acid | 5.10 | 3.68 × 10−7 | 1.13 |
| GC_115 | l-Alanine | 7.60 | 8.95 × 10−7 | 1.12 |
| GC_122 | Glycine | 6.06 | 9.84 × 10−6 | 1.12 |
| GC_185 | l-Valine | 7.74 | 8.89 × 10−7 | 1.12 |
| GC_200 | 4-Hydroxybutanoic acid | 9.13 | 2.47 × 10−7 | 1.13 |
| GC_221 | l-Leucine | 8.36 | 1.41 × 10−6 | 1.12 |
| GC_233 | l-Isoleucine | 6.80 | 8.26 × 10−8 | 1.13 |
| GC_236 | l-Proline | 8.13 | 5.74 × 10−6 | 1.12 |
| GC_249 | Butanedioic acid | 6.81 | 1.79 × 10−8 | 1.13 |
| GC_258 | Glyceric acid | 7.88 | 3.10 × 10−8 | 1.13 |
| GC_270 | Fumaric acid | 9.83 | 2.10 × 10−8 | 1.13 |
| GC_290 | l-Threonine | 7.59 | 1.23 × 10−8 | 1.13 |
| GC_349 | Malic acid | 12.77 | 9.02 × 10−10 | 1.13 |
| GC_359 | meso-Erythritol | -2.49 | 5.77 × 10−6 | 1.12 |
| GC_361 | l-Methionine | 6.21 | 6.76 × 10−8 | 1.13 |
| GC_362 | l-5-Oxoproline | 7.61 | 3.01 × 10−8 | 1.13 |
| GC_364 | l-Aspartic acid | 8.73 | 2.22 × 10−8 | 1.13 |
| GC_368 | 4-Aminobutanoic acid | 6.54 | 5.38 × 10−6 | 1.12 |
| GC_425 | l-Ornithine | 11.77 | 2.35 × 10−6 | 1.12 |
| GC_427 | l-Glutamic acid | 8.47 | 5.08 × 10−8 | 1.13 |
| GC_430 | Phenylalanine | 6.41 | 2.81 × 10−8 | 1.13 |
| GC_439 | Tartaric acid | 8.50 | 2.50 × 10−8 | 1.13 |
| GC_457 | Arabinose | 4.81 | 3.56 × 10−6 | 1.12 |
| GC_468 | Xylitol | 4.18 | 2.86 × 10−7 | 1.13 |
| GC_484 | Arabinitol | -2.81 | 5.47 × 10−6 | 1.12 |
| GC_538 | Citric acid | 6.14 | 1.22 × 10−7 | 1.13 |
| GC_554 | Quininic acid | 4.25 | 4.31 × 10−7 | 1.13 |
| GC_557 | Fructose | 8.55 | 1.27 × 10−6 | 1.12 |
| GC_606 | Caffeic acid | 3.72 | 6.53 × 10−4 | 1.07 |
| GC_627 | Palmitic acid | 1.81 | 4.62 × 10−5 | 1.10 |
| GC_686 | 9,12-Octadecadienoic acid (Z, Z)- | 4.60 | 3.84 × 10−7 | 1.13 |
| GC_702 | Stearic acid | 1.56 | 7.71 × 10−5 | 1.10 |
| GC_786 | Arachidic acid | 1.60 | 1.01 × 10−4 | 1.10 |
| GC_863 | 2-Linoleoylglycerol | 4.88 | 7.03 × 10−7 | 1.12 |
| GC_897 | Lignoceric acid | 4.64 | 3.60 × 10−7 | 1.13 |
| GC_965 | Campesterol | 6.54 | 1.48 × 10−7 | 1.13 |
| GC_968 | Stigmasterol | 6.98 | 1.29 × 10−7 | 1.13 |
| LC_1736 | Lactucopicrin | 5.17 | 5.46 × 10−7 | 1.12 |
| LC_4802 | 1-(sn-Glycero-3-phospho)-1D-myo-inositol | 6.97 | 2.13 × 10−4 | 1.09 |
| No. | Compound | Log2 (FC H106/HY106) | p-Value | VIP Value |
|---|---|---|---|---|
| GC_103 | Glycolic acid | −2.29 | 2.12 × 10−5 | 1.23 |
| GC_108 | Pyruvic acid | −2.40 | 1.56 × 10−5 | 1.23 |
| GC_115 | L-Alanine | −3.14 | 2.64 × 10−5 | 1.23 |
| GC_122 | Glycine | −2.68 | 1.02 × 10−4 | 1.22 |
| GC_146 | R-3-Hydroxybutyric acid | 1.78 | 5.14 × 10−3 | 1.12 |
| GC_177 | Propanedioic acid | −34.26 | 5.88 × 10−12 | 1.24 |
| GC_185 | L-Valine | −3.42 | 2.20 × 10−5 | 1.23 |
| GC_200 | 4-Hydroxybutanoic acid | −5.27 | 4.09 × 10−3 | 1.13 |
| GC_223 | Glycerol | −1.07 | 3.67 × 10−3 | 1.14 |
| GC_233 | L-Isoleucine | −3.66 | 1.03 × 10−5 | 1.23 |
| GC_236 | L-Proline | −4.53 | 2.99 × 10−7 | 1.24 |
| GC_249 | Butanedioic acid | −1.14 | 4.45 × 10−3 | 1.13 |
| GC_258 | Glyceric acid | −3.90 | 1.87 × 10−6 | 1.24 |
| GC_261 | Uracil | 2.41 | 2.39 × 10−3 | 1.15 |
| GC_270 | Fumaric acid | −4.54 | 1.01 × 10−5 | 1.23 |
| GC_290 | L-Threonine | −3.72 | 5.43 × 10−6 | 1.24 |
| GC_349 | Malic acid | −6.99 | 1.50 × 10−8 | 1.24 |
| GC_359 | meso-Erythritol | −1.73 | 4.82 × 10−4 | 1.19 |
| GC_361 | L-Methionine | −2.00 | 6.25 × 10−4 | 1.19 |
| GC_362 | L-5-Oxoproline | −4.81 | 1.07 × 10−7 | 1.24 |
| GC_364 | L-Aspartic acid | −3.44 | 8.99 × 10−6 | 1.23 |
| GC_391 | L-Threonic acid | −3.20 | 1.49 × 10−5 | 1.23 |
| GC_425 | L-Ornithine | −2.72 | 2.15 × 10−6 | 1.24 |
| GC_427 | L-Glutamic acid | −3.80 | 3.32 × 10−6 | 1.24 |
| GC_430 | Phenylalanine | −3.35 | 7.81 × 10−6 | 1.23 |
| GC_439 | Tartaric acid | −5.25 | 6.12 × 10−8 | 1.24 |
| GC_468 | Xylitol | −2.46 | 9.96 × 10−5 | 1.22 |
| GC_484 | Arabinitol | −1.37 | 1.75 × 10−4 | 1.21 |
| GC_513 | Ribonic acid | −2.90 | 4.47 × 10−5 | 1.22 |
| GC_538 | Citric acid | −2.77 | 1.19 × 10−4 | 1.22 |
| GC_554 | Quininic acid | −1.95 | 3.91 × 10−5 | 1.23 |
| GC_557 | Fructose | −1.80 | 8.22 × 10−3 | 1.10 |
| GC_627 | Palmitic acid | 1.68 | 6.45 × 10−5 | 1.22 |
| GC_702 | Stearic acid | 1.94 | 3.32 × 10−5 | 1.23 |
| GC_786 | Arachidic acid | 2.00 | 2.69 × 10−5 | 1.23 |
| GC_874 | 1-Monolinolein | 1.54 | 1.22 × 10−4 | 1.21 |
| GC_917 | 1-Hexacosanol | −2.44 | 4.29 × 10−6 | 1.24 |
| LC_1072 | Dihydrocaffeic acid hexose | −1.69 | 7.35 × 10−5 | 1.22 |
| LC_1239 | 15-deoxylactucin-8-sulfate | −4.41 | 3.54 × 10−7 | 1.24 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.; Yang, X.; Huo, G.; Ge, G.; Liu, H.; Luo, L.; Hu, J.; Huang, D.; Long, P. Distinct Metabolome Changes during Seed Germination of Lettuce (Lactuca sativa L.) in Response to Thermal Stress as Revealed by Untargeted Metabolomics Analysis. Int. J. Mol. Sci. 2020, 21, 1481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041481
Wei S, Yang X, Huo G, Ge G, Liu H, Luo L, Hu J, Huang D, Long P. Distinct Metabolome Changes during Seed Germination of Lettuce (Lactuca sativa L.) in Response to Thermal Stress as Revealed by Untargeted Metabolomics Analysis. International Journal of Molecular Sciences. 2020; 21(4):1481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041481
Chicago/Turabian StyleWei, Shiwei, Xiao Yang, Guotao Huo, Guojun Ge, Hongyan Liu, Lijun Luo, Jinguo Hu, Danfeng Huang, and Ping Long. 2020. "Distinct Metabolome Changes during Seed Germination of Lettuce (Lactuca sativa L.) in Response to Thermal Stress as Revealed by Untargeted Metabolomics Analysis" International Journal of Molecular Sciences 21, no. 4: 1481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041481