Structural and Functional Characterization of New SsoPox Variant Points to the Dimer Interface as a Driver for the Increase in Promiscuous Paraoxonase Activity


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Directed Evolution for the Generation of New Mutant PLL
2.2. Biochemical Characterization of 4Mut Mutant
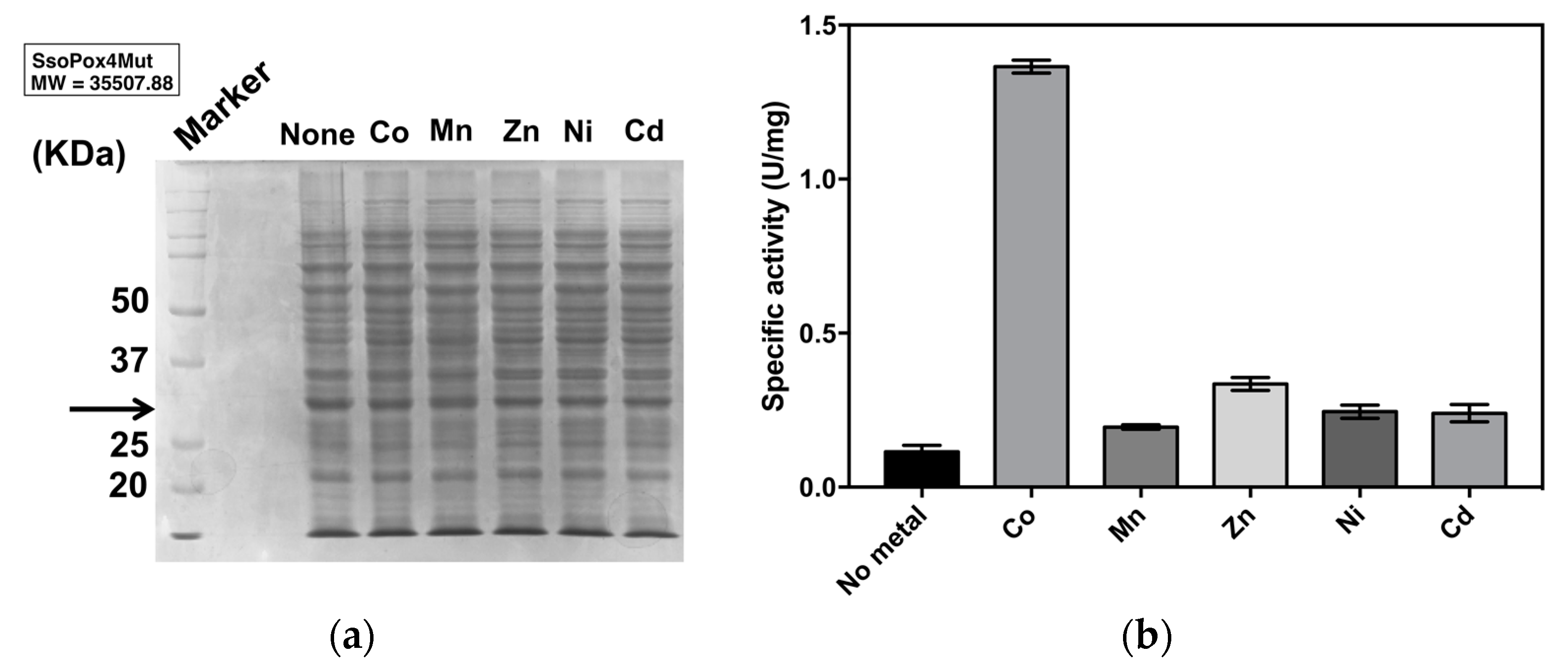
2.2.1. Optimal Metal Ion
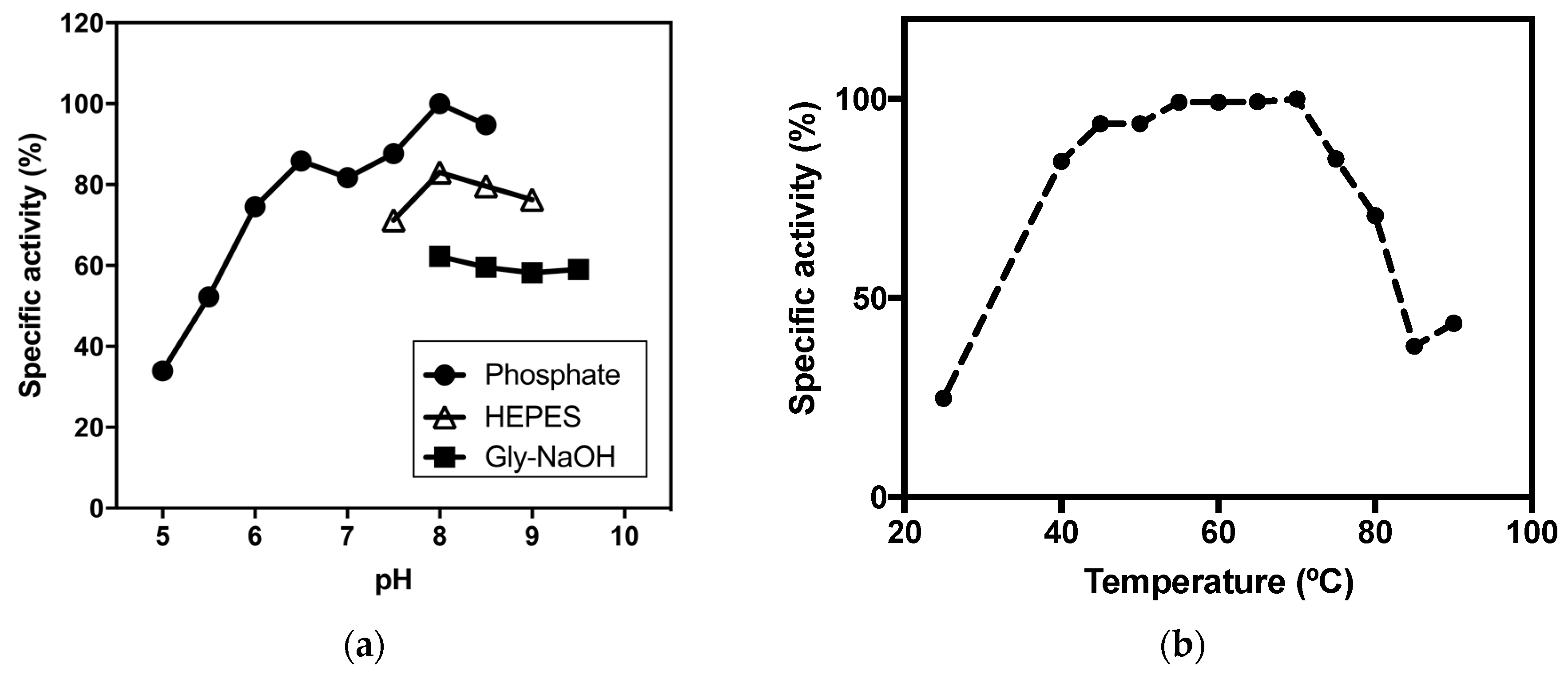
2.2.2. pH Optima
2.2.3. Thermophilicity
2.2.4. Thermal stability of 4Mut
2.3. Enzymatic Characterization of 4Mut
2.3.1. Activity on Phosphotriesters
2.3.2. Effects of SDS on Paraoxonase Activity of 4Mut
2.3.3. Lactonase and Carboxylesterase Activities of 4Mut
2.3.4. Detoxification of Nerve Agents
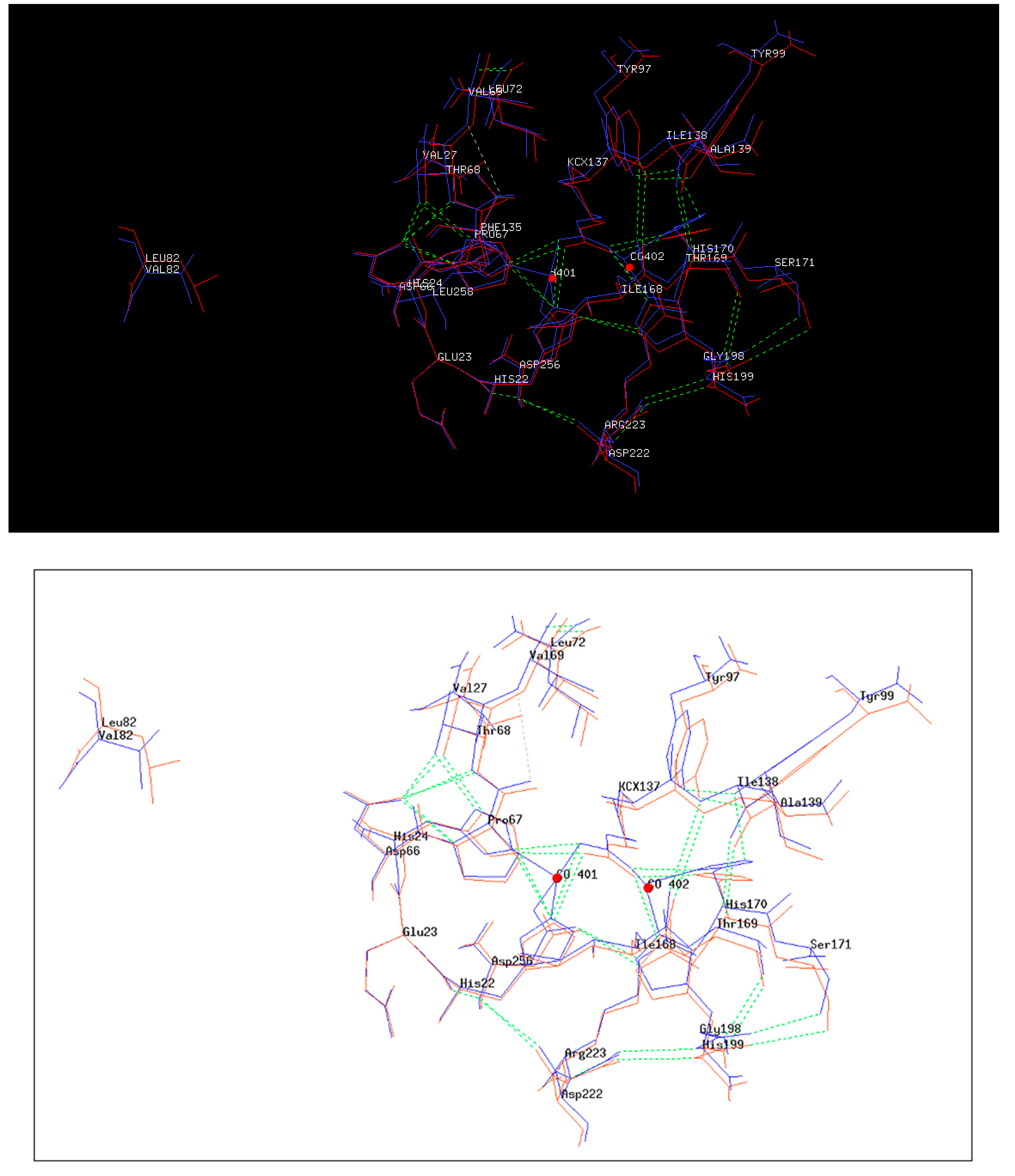
2.4. 3D Structure of 4Mut
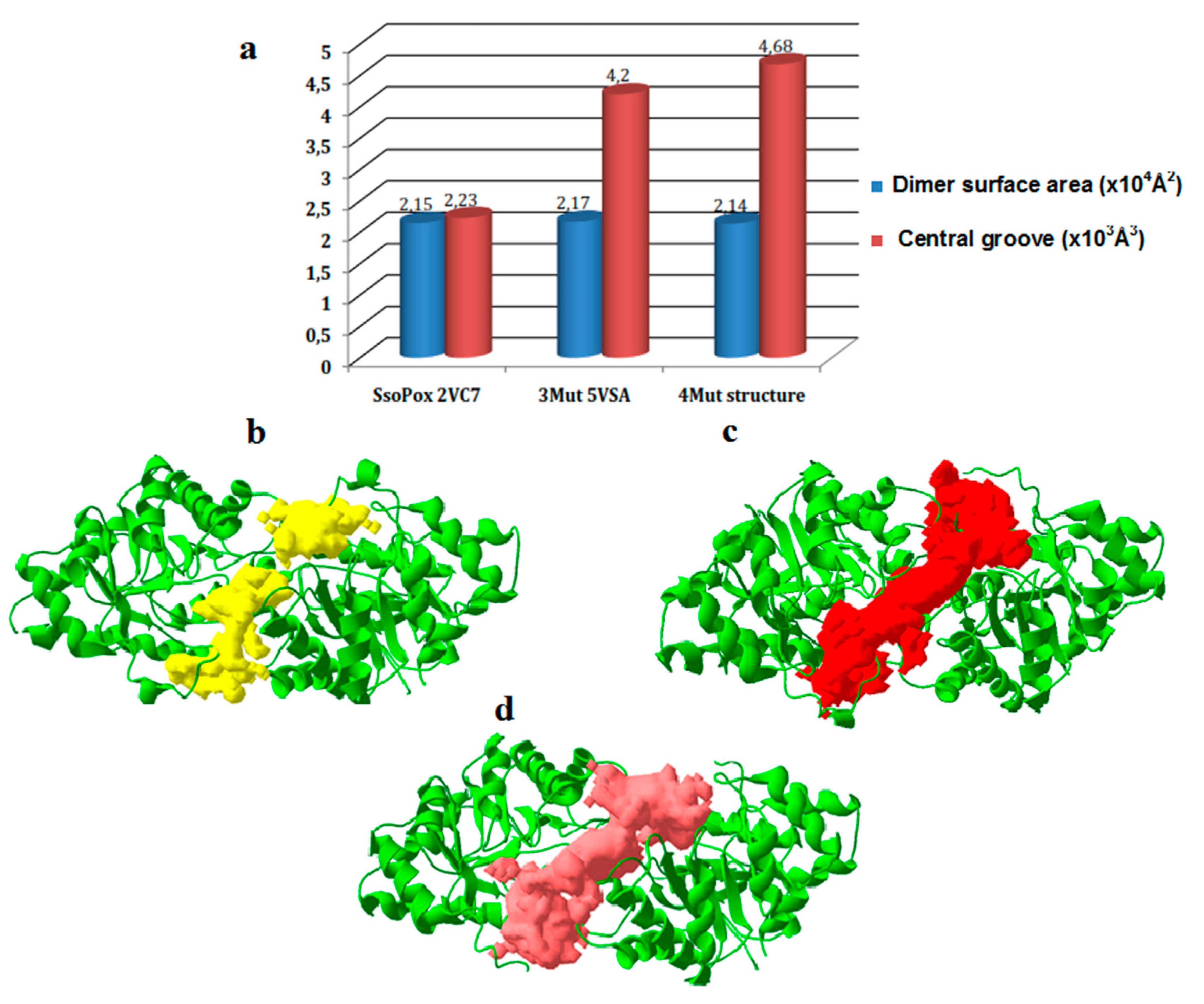
2.5. Structures Comparison
2.6. Circular Dichroism Analyses on 4Mut and 3Mut
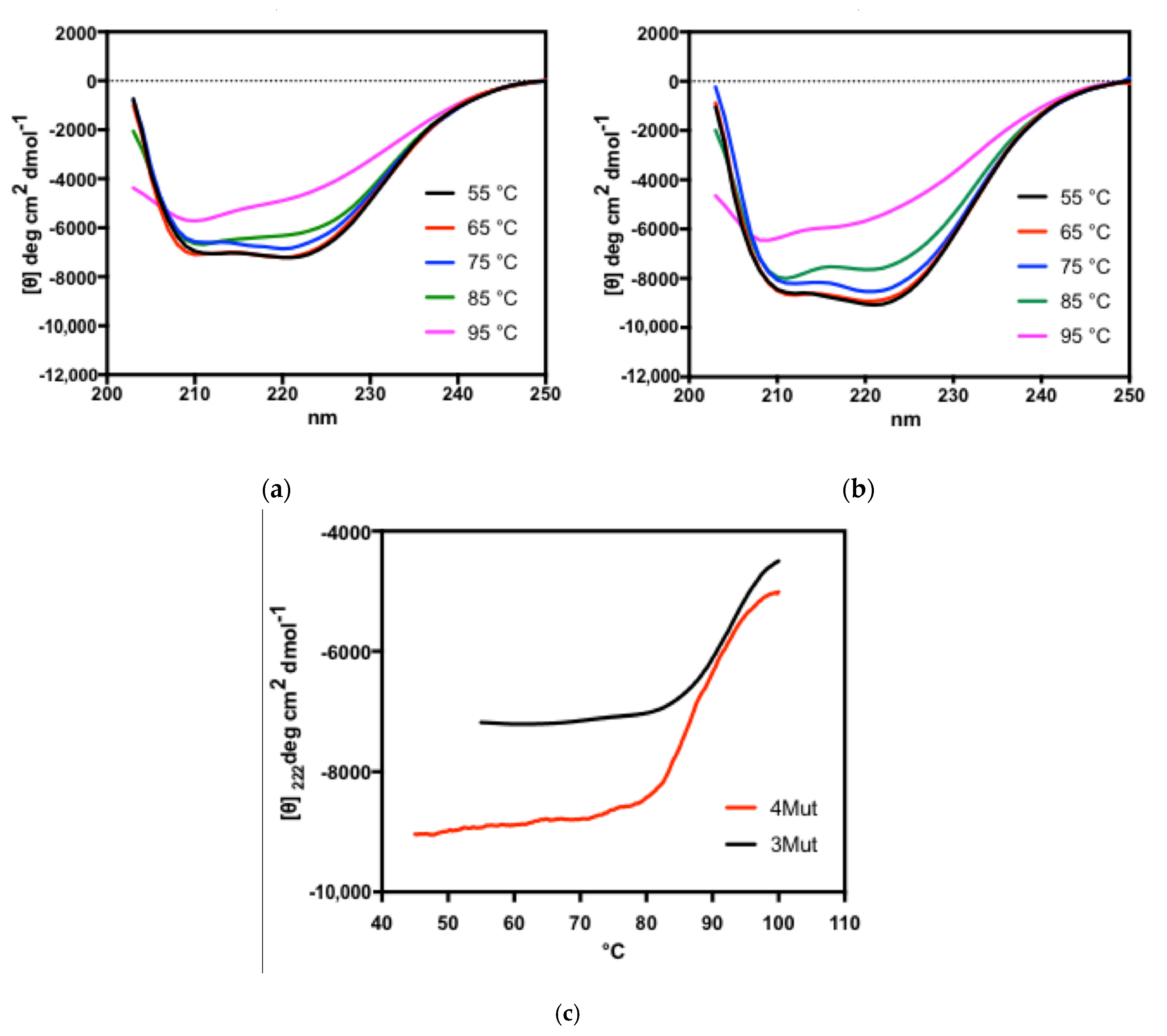
2.6.1. Far-UV CD Spectra for the Analysis of Secondary Structures
2.6.2. Near-UV CD Spectra for the Analysis of Tertiary Structures
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. DNA Manipulation
4.3. Preparation of PLL Genes as Templates for DNA StEP
4.4. Generation of Mutant Library by DNA StEP
4.5. Expression of Mutant Library
4.6. Screening for Paraoxonase Activity of Variants
4.7. Protein Expression and Purification in Large Scale
4.8. Phosphotriesterase Activity Assay
4.9. Determination of Metal Ions for the Enhancement of Paraoxonase Activity
4.10. Determination of Optimal pH, Thermophilicity and Thermal Stability
4.11. Lactonase Activity Assay
4.12. Carboxylesterase Activity Assay
4.13. Preparation of 4Mut Enzyme for Crystallization
4.14. Crystallization, Data Collection, Refinement and Structure Determination
4.15. Metal Contents Analysis
4.16. Detoxification of Nerve Agents
4.17. Model Generation and Analyses
4.18. Circular Dichroism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PLL | Phosphotriesterase-Like-Lactonase |
| StEP | Staggered Extension Process |
| OP | organophosphate |
| CWA | chemical warfare agent |
| bdPTE | phosphotriesterase from Brevundimonas diminuta MG |
| DrOPH/Dr0930 | phosphotriesterase-like-lactonase from Deinococcus radiodurans |
| GKL/GKaP | phosphotriesterase-like-lactonase from Geobacillus kaustophilus |
| StLois | stepwise loop insertion strategy |
| CD | circular dichroism |
| AHL | acyl-homoserine lactone |
| TBBL | 5-thiobutyl butyrolactone |
| C8-HSL | N-octanoyl homoserine lactone |
| C10-HSL | N-decanoyl homoserine lactone |
| ICP-MS | Inductively Coupled Plasma mass spectrometry |
| OpdA | phosphotriesterase from Agrobacterium radiobacter P230 |
| IPTG | Isopropyl β-D-1-thiogalactopyranoside |
| DTNB | 5,5′-Dithiobis(2-nitrobenzoic acid) |
| AChE | acetylcholinesterase |
References
- Raushel, F.M. Catalytic detoxification. Nature 2011, 496, 310–311. [Google Scholar] [CrossRef] [PubMed]
- Enserink, M. U.N. taps special labs to investigate Syrian attack. Science 2013, 341, 1050–1051. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C. Chemical detoxification of nerve agent VX. Acc. Chem. Res. 1999, 32, 109–115. [Google Scholar] [CrossRef]
- Pearson, G.S.; Magee, R.S. Critical evaluation of proven chemical weapon destruction technologies: (IUPAC technical report). Pure Appl. Chem. 2002, 74, 187–316. [Google Scholar] [CrossRef]
- Kim, K.; Tsay, O.G.; Atwood, D.A.; Churchill, D.G. Destruction and detection of chemical warfare agents. Chem. Rev. 2011, 111, 5345–5403. [Google Scholar] [CrossRef]
- Tuorinsky, S.D.; Caneva, D.C.; Sidell, F.R. TRIAGE OF CHEMICAL CASUALTIES, in Medical Aspects of Chemical Warfare; Borden Institute Walter Reed Army Medical Center: Washington, DC, USA, 2008; pp. 511–526. [Google Scholar]
- Tucker, M.D. Reduced weight decontamination formulation for neutralization of chemical and biological warfare agents 2014. U.S. patent US8741174B1, 3 June 2014. [Google Scholar]
- English, J.A., II. Design Aspects of a Low Emission, Two-Stage Incinerator. In Proceedings of the 1974 ASME National Incinerator Conference; ASME: New York, NY, USA, 2015. [Google Scholar]
- Kiss, A.; Virág, D. Photostability and Photodegradation Pathways of Distinctive Pesticides. J. Environ. Qual. 2009, 38, 157–163. [Google Scholar] [CrossRef]
- Reddy, P.V.L.; Kim, K.H. A review of photochemical approaches for the treatment of a wide range of pesticides. J. Hazard. Mater. 2015, 285, 325–335. [Google Scholar] [CrossRef]
- LeJeune, K.E.; Wild, J.E.; Russell, A.J. Nerve agents degrade by enzymatic foams. Nature 1998, 395, 27–28. [Google Scholar] [CrossRef]
- Serdar, C.M.; Gibson, D.T.; Munnecke, D.M.; Lancaster, J.H. Plasmid Involvement in Parathion Hydrolysis by Pseudomonas diminuta. Appl. Environ. Microbiol. 1982, 44, 246–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, D.P.; Caldwell, S.R.; Wild, J.R.; Raushel, F.M. Purification and properties of the phosphotriesterase from Pseudomonas diminuta. J. Biol. Chem. 1989, 264, 19659–19665. [Google Scholar] [PubMed]
- Dumas, D.P.; Durst, H.D.; Landis, W.G.; Raushel, F.M.; Wild, J.R. Inactivation of organophosphorus nerve agents by the phosphotriesterase from Pseudomonas diminuta. Arch. Biochem. Biophys. 1990, 277, 155–159. [Google Scholar] [CrossRef]
- Kolakowski, J.E.; Defrank, J.J.; Harvey, S.P.; Szafraniec, L.L.; Beaudry, W.T.; Lai, K.; Wild, J.R. Enzymatic Hydrolysis of the Chemical Warfare Agent VX and its Neurotoxic Analogues by Organophosphorus Hydrolase. Biocatal. Biotransformation 1997, 15, 297–312. [Google Scholar] [CrossRef]
- Caldwell, S.R.; Newcomb, J.R.; Schlecht, K.A.; Raushel, F.M. Limits of diffusion in the hydrolysis of substrates by the phosphotriesterase from Pseudomonas diminuta. Biochemistry 1991, 30, 7438–7444. [Google Scholar] [CrossRef] [PubMed]
- Afriat, L.; Roodveldt, C.; Manco, G.; Tawfik, D.S. The latent promiscuity of newly identified microbial lactonases is linked to a recently diverged phosphotriesterase. Biochemistry 2006, 45, 13677–13686. [Google Scholar] [CrossRef]
- Merone, L.; Mandrich, L.; Rossi, M.; Manco, G. A thermostable phosphotriesterase from the archaeon Sulfolobus solfataricus: Cloning, overexpression and properties. Extremophiles 2005, 9, 297–305. [Google Scholar] [CrossRef]
- Merone, L.; Mandrich, L.; Porzio, E.; Rossi, M.; Müller, S.; Reiter, G.; Worek, F.; Manco, G. Improving the promiscuous nerve agent hydrolase activity of a thermostable archaeal lactonase. Bioresour. Technol. 2010, 101, 9204–9212. [Google Scholar] [CrossRef]
- Porzio, E.; Merone, L.; Mandrich, L.; Rossi, M.; Manco, G. A new phosphotriesterase from Sulfolobus acidocaldarius and its comparison with the homologue from Sulfolobus solfataricus. Biochimie 2007, 89, 625–636. [Google Scholar] [CrossRef]
- Porzio, E.; Di Gennaro, S.; Palma, A.; Manco, G. Mn2+ modulates the kinetic properties of an archaeal member of the PLL family. Chem. Biol. Interact. 2013, 203, 251–256. [Google Scholar] [CrossRef]
- Del Giudice, I.; Coppolecchia, R.; Merone, L.; Porzio, E.; Carusone, T.M.; Mandrich, L.; Worek, F.; Manco, G. An efficient thermostable organophosphate hydrolase and its application in pesticide decontamination. Biotechnol. Bioeng. 2016, 113, 724–734. [Google Scholar] [CrossRef]
- Hawwa, R.; Larsen, S.D.; Ratia, K.; Mesecar, A.D. Structure-Based and Random Mutagenesis Approaches Increase the Organophosphate-Degrading Activity of a Phosphotriesterase Homologue from Deinococcus radiodurans. J. Mol. Biol. 2009, 393, 36–57. [Google Scholar] [CrossRef]
- Hawwa, R.; Aikens, J.; Turner, R.J.; Santarsiero, B.D.; Mesecar, A.D. Structural basis for thermostability revealed through the identification and characterization of a highly thermostable phosphotriesterase-like lactonase from Geobacillus stearothermophilus. Arch. Biochem. Biophys. 2009, 488, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, J.Y.; Xue, B.; Lee, K.H.; Tung, A.; Wu, L.; Robinson, R.C.; Yew, W.S. Directed evolution of a thermostable quorum-quenching lactonase from the amidohydrolase superfamily. J. Biol. Chem. 2010, 285, 40911–40920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiblot, J.; Gotthard, G.; Chabriere, E.; Elias, M. Structural and Enzymatic characterization of the lactonase SisLac from Sulfolobus islandicus. PLoS ONE 2012, 7, e47028. [Google Scholar] [CrossRef] [PubMed]
- Kallnik, V.; Bunescu, A.; Sayer, C.; Bräsen, C.; Wohlgemuth, R.; Littlechild, J.; Siebers, B. Characterization of a phosphotriesterase-like lactonase from the hyperthermoacidophilic crenarchaeon Vulcanisaeta moutnovskia. J. Biotechnol. 2014, 190, 11–17. [Google Scholar] [CrossRef]
- Hoque, M.A.; Zhang, Y.; Chen, L.; Yang, G.-Y.; Khatun, M.A.; Chen, H.-F.; Hao, L.; Feng, Y. Stepwise Loop Insertion Strategy for Active Site Remodeling to Generate Novel Enzyme Functions. ACS Chem. Biol. 2017, 12, 1188–1193. [Google Scholar] [CrossRef]
- Hiblot, J.; Gotthard, G.; Elias, M.; Chabriere, E. Differential Active Site Loop Conformations Mediate Promiscuous Activities in the Lactonase SsoPox. PLoS ONE 2013, 8, e75272. [Google Scholar] [CrossRef] [Green Version]
- Jacquet, P.; Hiblot, J.; Daudé, D.; Bergonzi, C.; Gotthard, G.; Armstrong, N.; Chabrière, E.; Elias, M. Rational engineering of a native hyperthermostable lactonase into a broad spectrum phosphotriesterase. Sci. Rep. 2017, 7, 16745. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Giver, L.; Shao, Z.; Affholter, J.A.; Arnold, F.H. Molecular evolution by staggered extension process (StEP) in vitro recombination. Nat. Biotechnol. 1998, 16, 258–261. [Google Scholar] [CrossRef]
- Hiblot, J.; Gotthard, G.; Chabriere, E.; Elias, M. Characterisation of the organophosphate hydrolase catalytic activity of SsoPox. Sci. Rep. 2012, 2, 779. [Google Scholar] [CrossRef] [Green Version]
- Porzio, E.; Bettazzi, F.; Mandrich, L.; Del Giudice, I.; Restaino, O.F.; Laschi, S.; Febbraio, F.; De Luca, V.; Borzacchiello, M.G.; Carusone, T.M.; et al. Innovative Biocatalysts as Tools to Detect and Inactivate Nerve Agents. Sci. Rep. 2018, 8, 13773. [Google Scholar] [CrossRef]
- Elias, M.; Dupuy, J.; Merone, L.; Mandrich, L.; Porzio, E.; Moniot, S.; Rochu, D.; Lecomte, C.; Rossi, M.; Masson, P.; et al. Structural Basis for Natural Lactonase and Promiscuous Phosphotriesterase Activities. J. Mol. Biol. 2008, 379, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, P.; Elias, M.; Merone, L.; Graziano, G.; Dupuy, J.; Mandrich, L.; Carullo, P.; Fournier, B.; Rochu, D.; Rossi, M.; et al. Structural determinants of the high thermal stability of SsoPox from the hyperthermophilic archaeon Sulfolobus solfataricus. Extremophiles 2009, 13, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Kaltenbach, M.; Emond, S.; Hollfelder, F.; Tokuriki, N. Functional Trade-Offs in Promiscuous Enzymes Cannot Be Explained by Intrinsic Mutational Robustness of the Native Activity. PLoS Genet. 2016. [Google Scholar] [CrossRef] [PubMed]
- Muga, A.; Arrondo, J.L.R.; Bellon, T.; Sancho, J.; Bernabeu, C. Structural and Functional Studies on the Interaction of Sodium Dodecyl Sulfate with β-Galactosidase. Arch. Biochem. Biophys. 1993, 300, 451–457. [Google Scholar] [CrossRef]
- Tanford, C. The hydrophobic effect and the organization of living matter. Science 1978, 200, 1012–1018. [Google Scholar] [CrossRef]
- D’Auria, S.; Di Cesare, N.; Gryczynski, I.; Rossi, M.; Lakowicz, J.R. On the effect of sodium dodecyl sulfate on the structure of β-galactosidase from Escherichia coli. A fluorescence study. J. Biochem. 2001, 130, 13–18. [Google Scholar] [CrossRef]
- Mandrich, L.; Di Gennaro, S.; Palma, A.; Manco, G. A further biochemical characterization of DrPLL the thermophilic lactonase from Deinococcus radiodurans. Protein Pept Lett 2013, 20, 36–44. [Google Scholar] [CrossRef]
- Leslie, A.G.W.; Powell, H.R. Processing diffraction data with mosflm. In Evolving Methods for Macromolecular Crystallography; Randy J, R., Sussman, J.L., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 41–51. ISBN 978-1-4020-6316-9. [Google Scholar]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D. Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef]
- French, S.; Wilson, K. On the treatment of negative intensity observations. Acta Crystallogr. Sect. A 1978, 517–525. [Google Scholar] [CrossRef]
- McCoy, A.J. Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallogr. Sect. D Biol. Crystallogr. 2007, 63, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Vagin, A.A.; Lebedev, A.; Wilson, K.S.; Dodson, E.J. Efficient anisotropic refinement of macromolecular structures using FFT. Acta Crystallogr. Sect. D Biol. Crystallogr. 1999, D55, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, D60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, R.P.; Long, F.; Murshudov, G.N.; Perrakis, A. The PDB_REDO server for macromolecular structure model optimization. IUCrJ 2014, 1 Pt 4, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66 Pt 1, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Delano, W.L. The PyMOL Molecular Graphics System. DeLano Sci. San Carlos, CA, USA 2002. [Google Scholar]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Foglia, F.; Mandrich, L.; Pezzullo, M.; Graziano, G.; Barone, G.; Rossi, M.; Manco, G.; Del Vecchio, P. Role of the N-terminal region for the conformational stability of esterase 2 from Alicyclobacillus acidocaldarius. Biophys. Chem. 2007, 127, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Roviello, G.N.; Moccia, M.; Sapio, R.; Valente, M.; Bucci, E.M.; Castiglione, M.; Pedone, C.; Perretta, G.; Benedetti, E.; Musumeci, D. Synthesis, characterization and hybridization studies of new nucleo-γ-peptides based on diaminobutyric acid. J. Pept. Sci. 2006, 12, 829–835. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Substrates | Name | λ (nm) | U/mg |
|---|---|---|---|
| Phosphotriester | paraoxon | 405 | 206.4 ± 18.3 |
| m-paraoxon | 405 | 50.9 ± 12.0 | |
| parathion | 405 | N.A | |
| m-parathion | 405 | 0.24 ± 0.01 | |
| diazinon | 228 | N.A | |
| coumaphos | 348 | N.A | |
| dursban | 276 | N.A | |
| malathion | 405 | N.A | |
| phosmet | 405 | N.A | |
| dimethoate | 405 | N.A | |
| tolclophos | 224 | N.A | |
| cyanophos | 246 | N.A | |
| Carboxylester | pNP-butanoate | 405 | N.A |
| β-Naphthyl acetate | 320 | N.A | |
| Lactone | N-octanoyl HSL | 572 | N.A |
| N-decanoyl HSL | 572 | N.A | |
| TBBL | 412 | 63.0 ± 3.4 |
| Enzymes | Substrate | Conditions | Vmax (U/mg) | kcat (s−1) | KM (mM) | kcat/KM (s−1 M−1) |
|---|---|---|---|---|---|---|
| SsoPox[18] | paraoxon | 70 °C | 0.42 ± 0.02 | 0.24 ± 0.01 | 0.060 ± 0.009 | 3.8 ± 0.75 × 103 |
| W263F[19] | paraoxon | 70 °C | 12.8 ± 0.3 | 3.8 ± 0.14 | 0.153 ± 0.039 | 2.4 × 104 |
| W263F[19] | paraoxon | 25 °C | n.d. | n.d. | n.d. | 5.8 × 102 |
| 3Mut[21] | paraoxon | 65 °C | 123.6 ± 12.8 | 71.0 ± 7.27 | 1.59 ± 0.114 | 4.5 ± 0.8 × 104 |
| paraoxon | 25 °C | 8.7 ± 0.6 | 6.91 ± 0.47 | 0.380 ± 0.067 | 1.8 ± 0.5 × 104 | |
| m-paraoxon | 65 °C | 72.9 ± 9.0 | 43.1 ± 5.3 | 0.744 ± 0.22 | 5.8 ± 0.9 × 104 | |
| paraoxon | 65 ° + 0.025% SDS | 55.2 ± 3.2 | 31.7 ± 1.86 | 0.325 ± 0.05 | 1.0 ± 0.2 × 105 | |
| paraoxon | 25 °C + 0.025% SDS | 15.0 ± 0.39 | 8.6 ± 0.24 | 0.107 ± 0.012 | 0.8 ± 0.1 × 105 | |
| 4Mut | paraoxon | 70 °C | 206.4 ± 18.3 | 122.0 ± 10.3 | 0.747 ± 0.02 | 1.6 ± 0.2 × 105 |
| paraoxon | 25 °C | 55.8 ± 1.4 | 33.0 ± 0.8 | 0.645 ± 0.053 | 5.1 ± 0.5 × 104 | |
| m-paraoxon | 70 °C | 50.9 ± 12.0 | 30.1 ± 7.1 | 0.871 ± 0.043 | 3.5 ± 1.0 × 104 | |
| paraoxon | 70 °C +0.025% SDS | 343.8 ± 13.8 | 203.4 ± 8.2 | 0.475 ± 0.051 | 4.3 ± 0.7 × 105 | |
| paraoxon | 25 °C + 0.025% SDS | 61.6 ± 1.0 | 36.4 ± 0.6 | 0.366 ± 0.017 | 1.0 ± 0.1 × 105 |
| Nerve agents | SsoPox 3Mut kcat/KM(M−1 s−1) | SsoPox 4Mut kcat/KM(M−1 s−1) |
|---|---|---|
| Tabun | 6.87 × 101 | 3.2 × 102 |
| Sarin | 2.77 × 102 | 4.2 × 102 |
| Soman | 4.32 × 101 | 4.2 × 102 |
| Cyclosarin | 7.63 × 101 | N.A. |
| VX | N.A. | N.A. |
| Data Collection | SsoPox 4Mut |
|---|---|
| Space group | P212121 |
| Cell dimensions: a,b,c (Å) | 84.138, 105.009, 153.651 |
| Resolution (Å) | 46.03-2.813 |
| Upper resolution shell (Å) | 2.913 -2.813 |
| Measured reflections a | 385,386 (39,037) |
| Unique reflections | 33,783 (3,252) |
| Multiplicity | 11.4 (11.6) |
| Completeness (%) | 99.07 (96.79) |
| Average I/ σ (I) | 14.50 (2.20) |
| Wilson B-factor (Å2) | 62.38 |
| R-pim | 0.0419 (0.332) |
| CC1/2 | 0.998 (0.897) |
| Refinement Statistics | |
| Reflections used in refinement | 33,498 (3,252) |
| Reflections used for R-free | 1,681 (159) |
| R-work | 0.2178 (0.3506) |
| R-free | 0.2498 (0.3709) |
| Number of non-hydrogen atoms | 9424 |
| Number of protein atoms | 9412 |
| Solvent | 4 |
| RMS(bonds) (Å) | 0.011 |
| RMS(angles) (o) | 1.42 |
| Ramachandran favored (%) | 95.42 |
| Ramachandran allowed (%) | 4.50 |
| Ramachandran outliers (%) | 5.73 |
| Average B-factor (Å2) | 70.86 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzumoto, Y.; Dym, O.; Roviello, G.N.; Worek, F.; Sussman, J.L.; Manco, G. Structural and Functional Characterization of New SsoPox Variant Points to the Dimer Interface as a Driver for the Increase in Promiscuous Paraoxonase Activity. Int. J. Mol. Sci. 2020, 21, 1683. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051683
Suzumoto Y, Dym O, Roviello GN, Worek F, Sussman JL, Manco G. Structural and Functional Characterization of New SsoPox Variant Points to the Dimer Interface as a Driver for the Increase in Promiscuous Paraoxonase Activity. International Journal of Molecular Sciences. 2020; 21(5):1683. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051683
Chicago/Turabian StyleSuzumoto, Yoko, Orly Dym, Giovanni N. Roviello, Franz Worek, Joel L. Sussman, and Giuseppe Manco. 2020. "Structural and Functional Characterization of New SsoPox Variant Points to the Dimer Interface as a Driver for the Increase in Promiscuous Paraoxonase Activity" International Journal of Molecular Sciences 21, no. 5: 1683. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051683