Aberrant Expression of RAD52, Its Prognostic Impact in Rectal Cancer and Association with Poor Survival of Patients

Abstract
:1. Introduction
2. Results
2.1. Study Population
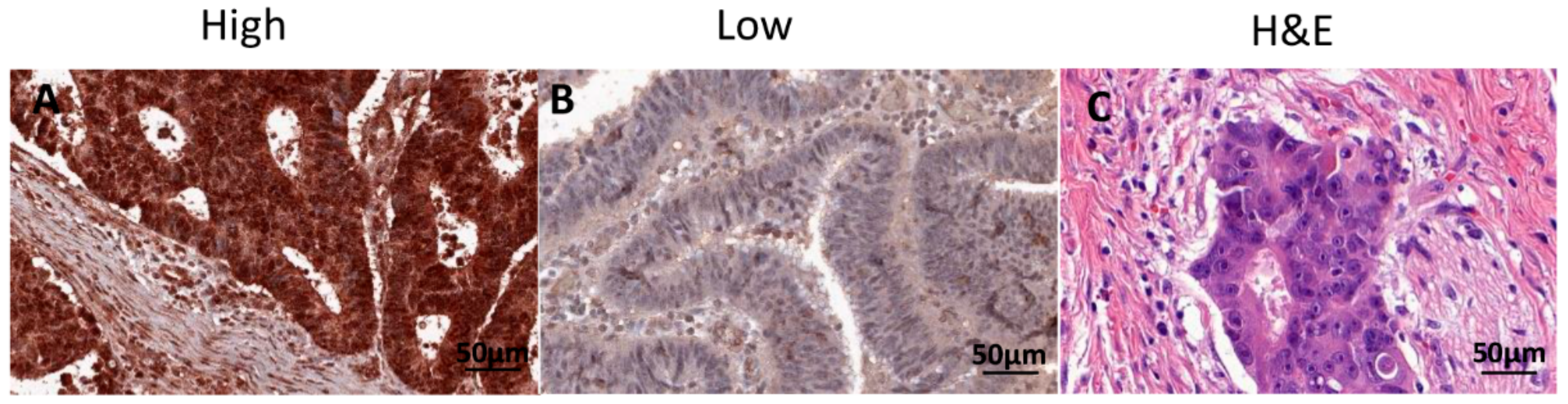
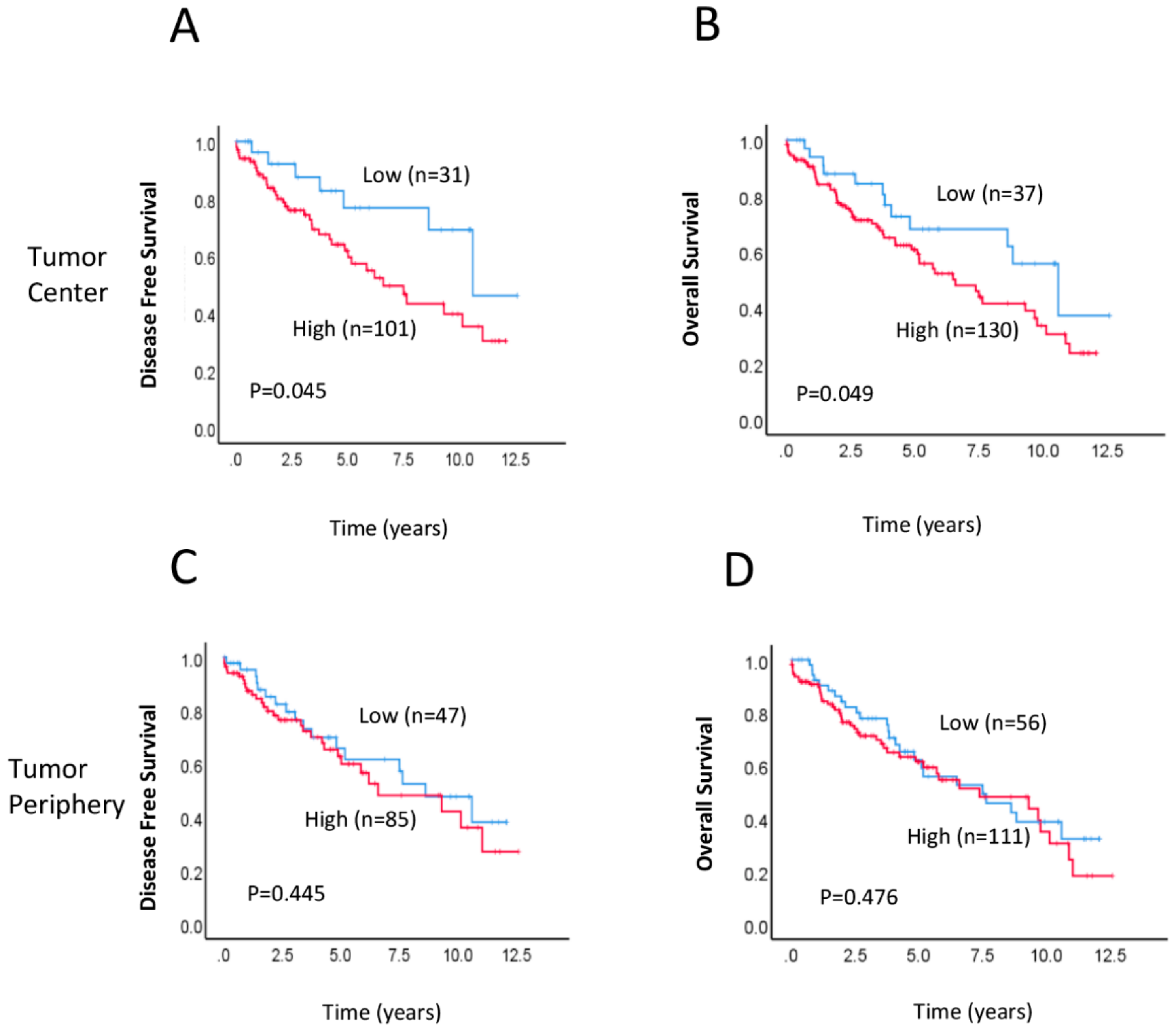
2.2. Association between RAD52 Expression and Clinicopathological Features and Prognosis
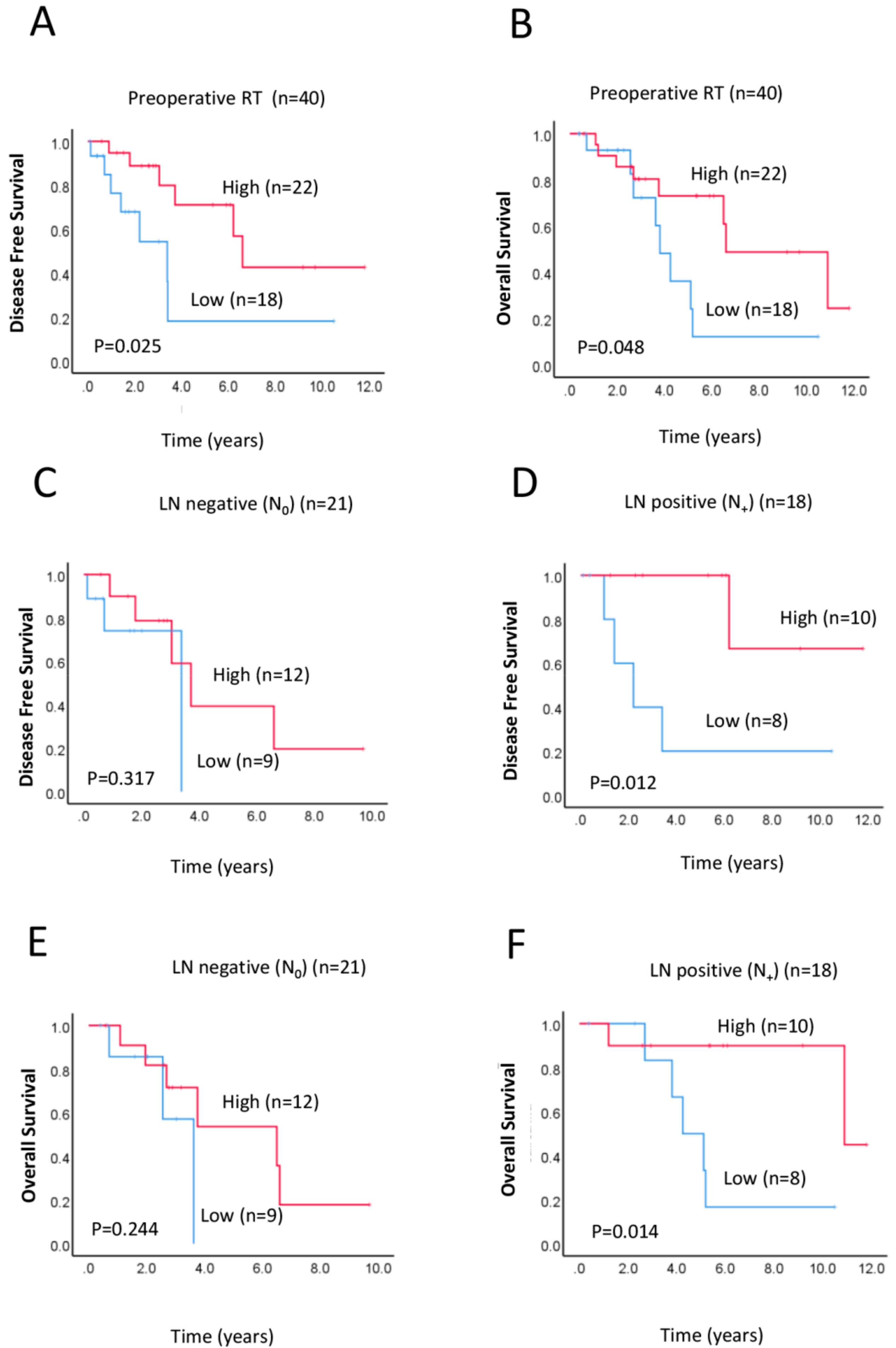
2.3. Correlation of RAD52 Expression with Survival Outcomes after Preoperative Neoadjuvant Therapy
2.4. Prognostic Implications of RAD52 Expression in Lymph Node (LN)-Positive Subgroup
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Response and Outcomes of Interest
4.3. Sample Preparation and Tissue Microarrays
4.4. Immunohistochemistry
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [Green Version]
- Hong, T.S.; Clark, J.W.; Haigis, K.M. Cancers of the colon and rectum: Identical or fraternal twins? Cancer Dis. 2012, 2, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.J.; Hathout, L.; Malhotra, U.; Maloney-Patel, N.; Kilic, S.; Poplin, E.; Jabbour, S.K. Decision-making strategy for rectal cancer management using radiation therapy for elderly or comorbid patients. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 926–944. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Skibber, J.M.; Rodriguez-Bigas, M.A.; Feig, B.W.; Chang, G.J.; Wolff, R.A.; Eng, C.; Krishnan, S.; Janjan, N.A.; Crane, C.H. Predictors of tumor response and downstaging in patients who receive preoperative chemoradiation for rectal cancer. Cancer 2007, 109, 1750–1755. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Wang, J. Molecular effectors of radiation resistance in colorectal cancer. Precis. Radiat. Oncol. 2017, 1, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Ruo, L.; Tickoo, S.; Klimstra, D.S.; Minsky, B.D.; Saltz, L.; Mazumdar, M.; Paty, P.B.; Wong, W.D.; Larson, S.M.; Cohen, A.M.; et al. Long-term prognostic significance of extent of rectal cancer response to preoperative radiation and chemotherapy. Annu. Surg. 2002, 236, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Tennstedt, P.; Fresow, R.; Simon, R.; Marx, A.; Terracciano, L.; Petersen, C.; Sauter, G.; Dikomey, E.; Borgmann, K. Rad51 overexpression is a negative prognostic marker for colorectal adenocarcinoma. Int. J. Cancer 2013, 132, 2118–2126. [Google Scholar] [CrossRef]
- Bottarelli, L.; De Angelis, G.L.; Azzoni, C.; Di Mario, F.; De’ Angelis, N.; Leandro, G.; Fornaroli, F.; Gaiani, F.; Negri, F. Potential predictive biomarkers in locally advanced rectal cancer treated with preoperative chemo-radiotherapy. Acta. Bio-Med. 2018, 89, 102–106. [Google Scholar]
- Dayde, D.; Tanaka, I.; Jain, R.; Tai, M.C.; Taguchi, A. Predictive and prognostic molecular biomarkers for response to neoadjuvant chemoradiation in rectal cancer. Int. J. Mol. Sci. 2017, 18, 573. [Google Scholar] [CrossRef]
- Ho, V.; Chung, L.; Revoltar, M.; Lim, S.H.; Tut, T.G.; Abubakar, A.; Henderson, C.J.; Chua, W.; Ng, W.; Lee, M.; et al. Mre11 and atm expression levels predict rectal cancer survival and their association with radiotherapy response. PLoS ONE 2016, 11, e0167675. [Google Scholar] [CrossRef]
- Peluso, G.; Incollingo, P.; Calogero, A.; Tammaro, V.; Rupealta, N.; Chiacchio, G.; Sandoval Sotelo, M.L.; Minieri, G.; Pisani, A.; Riccio, E.; et al. Current tissue molecular markers in colorectal cancer: A literature review. Biomed Res. Int. 2017, 2017, 2605628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacante, M.; Borzi, A.M.; Basile, F.; Biondi, A. Biomarkers in colorectal cancer: Current clinical utility and future perspectives. World J. Clin. Cases 2018, 6, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.C.; Yeo, S.G. Clinical utility of pretreatment prediction of chemoradiotherapy response in rectal cancer: A review. EPMA J. 2017, 8, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Povirk, L.F. Biochemical mechanisms of chromosomal translocations resulting from DNA double-strand breaks. DNA Repair 2006, 5, 1199–1212. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Wu, W.; Wang, M.; Terzoudi, G.I.; Pantelias, G.E. Backup pathways of nonhomologous end joining may have a dominant role in the formation of chromosome aberrations. In Chromosomal Alterations; Obe, G.V., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 67–85. [Google Scholar]
- Adamsen, B.L.; Kravik, K.L.; De Angelis, P.M. DNA signaling in response to 5-fluorouracil in three colorectal cancer cell lines with different mismatch repair and TP53 status. Int. J. Oncol. 2011, 39, 673–682. [Google Scholar] [PubMed] [Green Version]
- Lieber, M.R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasin, M.; Rothstein, R. Repair of strand breaks by homologous recombination. Cold Spring Harb. Perspect. Biol. 2013, 5, a012740. [Google Scholar] [CrossRef]
- Bhargava, R.; Onyango, D.O.; Stark, J.M. Regulation of single-strand annealing and its role in genome maintenance. Trends Genet. 2016, 32, 566–575. [Google Scholar] [CrossRef] [Green Version]
- Mladenov, E.; Magin, S.; Soni, A.; Iliakis, G. DNA double-strand break repair as determinant of cellular radiosensitivity to killing and target in radiation therapy. Front. Oncol. 2013, 3, 113. [Google Scholar] [CrossRef] [Green Version]
- Guo, G.S.; Zhang, F.M.; Gao, R.J.; Delsite, R.; Feng, Z.H.; Powell, S.N. DNA repair and synthetic lethality. Int. J. Oral Sci. 2011, 3, 176–179. [Google Scholar] [CrossRef]
- Morales, M.E.; White, T.B.; Streva, V.A.; DeFreece, C.B.; Hedges, D.J.; Deininger, P.L. The contribution of alu elements to mutagenic DNA double-strand break repair. PLoS Genet. 2015, 11, e1005016. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.B.; Carreira, A.; Kowalczykowski, S.C. Purified human brca2 stimulates rad51-mediated recombination. Nature 2010, 467, 678–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Doty, T.; Gibson, B.; Heyer, W.D. Human brca2 protein promotes rad51 filament formation on rpa-covered single-stranded DNA. Nat. Struct. Mol. Biol. 2010, 17, 1260–1262. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Scott, S.P.; Bussen, W.; Sharma, G.G.; Guo, G.; Pandita, T.K.; Powell, S.N. Rad52 inactivation is synthetically lethal with brca2 deficiency. Proc. Natl. Acad. Sci. USA 2011, 108, 686–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman, R.; You, M. Corrupting the DNA damage response: A critical role for rad52 in tumor cell survival. Aging 2017, 9, 1647–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naccarati, A.; Rosa, F.; Vymetalkova, V.; Barone, E.; Jiraskova, K.; Di Gaetano, C.; Novotny, J.; Levy, M.; Vodickova, L.; Gemignani, F.; et al. Double-strand break repair and colorectal cancer: Gene variants within 3’ utrs and micrornas binding as modulators of cancer risk and clinical outcome. Oncotarget 2016, 7, 23156–23169. [Google Scholar] [CrossRef] [Green Version]
- Sotiriou, S.K.; Kamileri, I.; Lugli, N.; Evangelou, K.; Da-Ré, C.; Huber, F.; Padayachy, L.; Tardy, S.; Nicati, N.L.; Barriot, S.; et al. Mammalian RAD52 functions in break-induced replication repair of collapsed DNA replication forks. Mol. Cell. 2016, 64, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Belickova, M.; Merkerova, M.D.; Stara, E.; Vesela, J.; Sponerova, D.; Mikulenkova, D.; Brdicka, R.; Neuwirtova, R.; Jonasova, A.; Cermak, J. DNA repair gene variants are associated with an increased risk of myelodysplastic syndromes in a Czech population. J. Hematol. Oncol. 2013, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Tassone, P.; Di Martino, M.T.; Ventura, M.; Pietragalla, A.; Cucinotto, I.; Calimeri, T.; Bulotta, A.; Neri, P.; Caraglia, M.; Tagliaferri, P. Loss of BRCA1 function increases the antitumor activity of cisplatin against human breast cancer xenografts in vivo. Cancer Biol. Ther. 2009, 8, 648–653. [Google Scholar] [CrossRef] [Green Version]
- Fayaz, S.; Karimmirza, M.; Tanhaei, S.; Fathi, M.; Torbati, P.M.; Fard-Esfahani, P. Increased risk of differentiated thyroid carcinoma with combined effects of homologous recombination repair gene polymorphisms in an Iranian population. Asian Pac. J. Cancer Prev. 2014, 14, 6727–6731. [Google Scholar] [CrossRef] [Green Version]
- Li, H.M.; Yuan, P.; Yu, D.K.; Ma, F.; Tan, W.W.; Feng, T.; Yang, J.; Huang, Y.; Lin, D.X.; Xu, B.H.; et al. Genetic variation in DNA repair gene RAD52 is associated with the response to platinum-based chemotherapy in SCLC patients. Zhonghua Zhong Liu Za Zhi 2016, 38, 504–509. [Google Scholar] [PubMed]
- De Azevêdo Silva, J.; Pancotto, J.A.; Donadi, E.A.; Crovella, S.; Sandrin-Garcia, P. LIG4 and RAD52 DNA repair genes polymorphisms and systemic lupus erythematosus. Mol. Biol. Rep. 2014, 41, 2249–2256. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Yang, X.; Ming, H.; Li, P. Expression and significance of gene Rad52 in nasopharyngeal carcinoma. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2014, 28, 486–489. [Google Scholar] [PubMed]
- Lieberman, R.; Xiong, D.; James, M.; Han, Y.; Amos, C.I.; Wang, L.; You, M. Functional characterization of RAD52 as a lung cancer susceptibility gene in the 12p13.33 locus. Mol. Carcinog. 2016, 55, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Xu, Y.; Zhang, Q.; Li, Y.; Jia, W.; Wang, X.; Xie, Z.; Liu, J.; Zhao, D.; Shao, M.; et al. Evaluating the role of RAD52 and its interactors as novel potential molecular targets for hepatocellular carcinoma. Cancer Cell. Int. 2019, 19, 279. [Google Scholar] [CrossRef]
- Bartkova, J.; Hamerlik, P.; Stockhausen, M.T.; Ehrmann, J.; Hlobilkova, A.; Laursen, H.; Kalita, O.; Kolar, Z.; Poulsen, H.S.; Broholm, H.; et al. Replication stress and oxidative damage contribute to aberrant constitutive activation of DNA damage signalling in human gliomas. Oncogene 2010, 29, 5095–5102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorgoulis, V.G.; Vassiliou, L.V.; Karakaidos, P.; Zacharatos, P.; Kotsinas, A.; Liloglou, T.; Venere, M.; Ditullio, R.A., Jr.; Kastrinakis, N.G.; Levy, B.; et al. Activation of the DNA damage checkpoint and genomic instability in human precancerous lesions. Nature 2005, 434, 907–913. [Google Scholar] [CrossRef]
- Oka, K.; Tanaka, T.; Enoki, T.; Yoshimura, K.; Ohshima, M.; Kubo, M.; Murakami, T.; Gondou, T.; Minami, Y.; Takemoto, Y.; et al. DNA damage signaling is activated during cancer progression in human colorectal carcinoma. Cancer Biol. Ther. 2010, 9, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Khanna, A. DNA damage in cancer therapeutics: A boon or a curse? Cancer Res. 2015, 75, 2133–2138. [Google Scholar] [CrossRef] [Green Version]
- Stark, J.M.; Pierce, A.J.; Oh, J.; Pastink, A.; Jasin, M. Genetic steps of mammalian homologous repair with distinct mutagenic consequences. Mol. Cell. Biol. 2004, 24, 9305–9316. [Google Scholar] [CrossRef] [Green Version]
- Belancio, V.P.; Roy-Engel, A.M.; Deininger, P.L. All y’all need to know ’bout retroelements in cancer. Semin. Cancer Biol. 2010, 20, 200–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Edwards, A.; Fan, W.; Deininger, P.; Zhang, K. Alu distribution and mutation types of cancer genes. BMC Genomics 2011, 12, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deininger, P.L.; Batzer, M.A. Alu repeats and human disease. Mol. Genet. Metab. 1999, 67, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, B.; Richardson, C.; Jasin, M. Chromosomal translocation mechanisms at intronic alu elements in mammalian cells. Mol. Cell. 2005, 17, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Manthey, G.M.; Bailis, A.M. Rad51 inhibits translocation formation by non-conservative homologous recombination in saccharomyces cerevisiae. PLoS ONE 2010, 5, e11889. [Google Scholar] [CrossRef] [Green Version]
- Berglund, E.; Maaskola, J.; Schultz, N.; Friedrich, S.; Marklund, M.; Bergenstrahle, J.; Tarish, F.; Tanoglidi, A.; Vickovic, S.; Larsson, L.; et al. Spatial maps of prostate cancer transcriptomes reveal an unexplored landscape of heterogeneity. Nat. Commun. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Munthe, S.; Petterson, S.A.; Dahlrot, R.H.; Poulsen, F.R.; Hansen, S.; Kristensen, B.W. Glioma cells in the tumor periphery have a stem cell phenotype. PLoS ONE 2016, 11, e0155106. [Google Scholar] [CrossRef]
- Chakraborty, U.; George, C.M.; Lyndaker, A.M.; Alani, E. A delicate balance between repair and replication factors regulates recombination between divergent DNA sequences in saccharomyces cerevisiae. Genetics 2016, 202, 525–540. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, N.; Goldfarb, T.; Studamire, B.; Alani, E.; Haber, J.E. Heteroduplex rejection during single-strand annealing requires sgs1 helicase and mismatch repair proteins msh2 and msh6 but not pms1. Proc. Natl. Acad. Sci. USA 2004, 101, 9315–9320. [Google Scholar] [CrossRef] [Green Version]
- Bennardo, N.; Gunn, A.; Cheng, A.; Hasty, P.; Stark, J.M. Limiting the persistence of a chromosome break diminishes its mutagenic potential. PLoS Genet. 2009, 5, e1000683. [Google Scholar] [CrossRef]
- Schadt, L.; Sparano, C.; Schweiger, N.A.; Silina, K.; Cecconi, V.; Lucchiari, G.; Yagita, H.; Guggisberg, E.; Saba, S.; Nascakova, Z.; et al. Cancer-Cell Intrinsic cGAS expression mediates tumor immunogenicity. Cell Rep. 2019, 29, 1236–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quek, H.; Luff, J.; Cheung, K.; Kozlov, S.; Gatei, M.; Lee, C.S.; Bellingham, M.C.; Noakes, P.G.; Lim, Y.C.; Barnett, N.L.; et al. A rat model of ataxia-telangiectasia: Evidence for a neurodegenerative phenotype. Hum. Mol. Genet. 2017, 26, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Falahat, R.; Perez-Villarroel, P.; Mailloux, A.W.; Zhu, G.; Pilon-Thomas, S.; Barber, G.N.; Mulé, J.J. STING Signaling in Melanoma Cells Shapes Antigenicity and Can Promote Antitumor T-cell Activity. Cancer Immunol. Res. 2019, 7, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.; Kastan, M.B. The DNA damage response: Implications for tumor responses to radiation and chemotherapy. Annu. Rev. Med. 2015, 66, 129–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.M.; Fan, S.; Rockwell, S.; Goldberg, I.D. The molecular and cellular basis of radiosensitivity: Implications for understanding how normal tissues and tumors respond to therapeutic radiation. Cancer Invest. 1999, 17, 56–72. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.J.; Song, X.M.; Chen, Z.H.; Ren, X.Q.; Xu, K.W.; Jing, H.; He, Y.L. Xrcc2 as a predictive biomarker for radioresistance in locally advanced rectal cancer patients undergoing preoperative radiotherapy. Oncotarget 2015, 6, 32193–32204. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.; Chung, L.; Singh, A.; Lea, V.; Revoltar, M.; Lim, S.H.; Tut, T.G.; Ng, W.; Lee, M.; de Souza, P.; et al. Early postoperative low expression of rad50 in rectal cancer patients associates with disease-free survival. Cancers 2017, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Ho, V.; Chung, L.; Singh, A.; Lea, V.; Abubakar, A.; Lim, S.H.; Ng, W.; Lee, M.; de Souza, P.; Shin, J.S.; et al. Overexpression of the mre11-rad50-nbs1 (mrn) complex in rectal cancer correlates with poor response to neoadjuvant radiotherapy and prognosis. BMC Cancer 2018, 18, 869. [Google Scholar] [CrossRef]
- Forker, L.J.; Choudhury, A.; Kiltie, A.E. Biomarkers of tumour radiosensitivity and predicting benefit from radiotherapy. Clin. Oncol. 2015, 27, 561–569. [Google Scholar] [CrossRef]
- Curtin, N.J. DNA repair dysregulation from cancer driver to therapeutic target. Nat. Rev. Cancer 2012, 12, 801–817. [Google Scholar] [CrossRef]
- Kan, Y.; Batada, N.N.; Hendrickson, E.A. Human somatic cells deficient for RAD52 are impaired for viral integration and compromised for most aspects of homology-directed repair. DNA Repair 2017, 55, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.-Y.; Yang, G.; Tu, X.-Y.; Yang, J.-M.; Qian, J.; Wu, X.-H.; Zhou, X.Y.; Cheng, X.; Wei, Q. RAD52 Variants Predict Platinum Resistance and Prognosis of Cervical Cancer. PLoS ONE 2012, 7, e50461. [Google Scholar] [CrossRef] [PubMed]
- Edge, S.B.; Compton, C.C. The American joint committee on cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | All Patients (%) | Preoperative Neoadjuvant-Therapy Group |
|---|---|---|
| Total, n | 179 | 40 |
| Age median | 71 | 67 |
| Gender | ||
| Male | 119 (66.5) | 27 (67.5) |
| Female | 60 (33.5) | 13 (32.5) |
| Tumour stage | ||
| T1, T2 | 58/177 (32.8) | 13/40 (32.5) |
| T3, T4 | 121/177 (67.2) | 27/40 (67.5) |
| Node stage | ||
| N0 | 92/173 (53.2) | 23/40 (57.5) |
| N1, N2 | 81/173 (46.8) | 17/40 (42.5) |
| Metastasis stage | ||
| M0 | 152/161(94.4) | 38/39 (97.4) |
| M1 | 9/161 (5.9) | 1/39 (2.6) |
| Grade | ||
| 1, 2 | 167/179 (93.3) | 38/40 (92.7) |
| 3 | 12/179 (6.7) | 2/40 (7.3) |
| Vascular invasion | ||
| Absent | 137/177 (77.4) | 35/40 (87.5) |
| Present | 40/177 (22.6) | 5/40 (12.5) |
| Perineural invasion | ||
| Absent | 155/177 (87.6) | 33/40 (82.5) |
| Present | 22/177 (12.4) | 7/40 (17.5) |
| Chemoradiotherapy | ||
| Total | 61/177 (34.5) | - |
| Neoadjuvant | 40/61 (65.6) | - |
| Adjuvant | 21/61 (34.4) | 0/40 (0) |
| Recurrence | ||
| Absent | 93/148 (62.8) | 22/38 (57.9) |
| Present | 55/148 (37.2) | 16/38 (42.1) |
| Tumour regression grade | ||
| 0, 2 (good response) | N/A | 6/37 (16.2) |
| 3 (poor response) | N/A | 31/37 (83.8) |
| Variables | Subgroups | Tumour Centre | Tumour Periphery | ||||
|---|---|---|---|---|---|---|---|
| Low (%) | High (%) | p value | Low (%) | High (%) | p value | ||
| Sex | Male | 71.4 | 64.4 | 0.709 | 64.4 | 67.2 | 0.405 |
| Female | 28.6 | 35.6 | 35.6 | 32.8 | |||
| Age | ≤71 | 31.0 | 51.9 | 0.489 | 44.1 | 49.6 | 0.018 |
| >71 | 69.0 | 48.1 | 55.9 | 50.4 | |||
| Tumour stage | T1, T2 | 45.0 | 29.3 | 0.065 | 41.4 | 28.4 | 0.134 |
| T3, T4 | 55.0 | 70.7 | 58.6 | 71.6 | |||
| Node stage | Negative | 58.5 | 49.6 | 0.028 | 63.8 | 46.1 | 0.320 |
| Positive | 41.5 | 50.4 | 36.2 | 53.9 | |||
| Metastasis stage | M0 | 94.4 | 93.7 | 0.453 | 96.3 | 93.4 | 0.984 |
| M1 | 5.6 | 6.3 | 3.7 | 6.6 | |||
| Grade | 1, 2 | 88.1 | 94.1 | 0.673 | 91.5 | 93.3 | 0.196 |
| 3 | 11.9 | 5.9 | 8.5 | 6.7 | |||
| Vascular invasion | Absent | 87.8 | 73.1 | 0.184 | 82.8 | 73.7 | 0.053 |
| Present | 12.2 | 26.9 | 17.2 | 26.3 | |||
| Perineural invasion | Absent | 92.7 | 85.1 | 0.842 | 86.2 | 87.3 | 0.208 |
| Present | 7.3 | 14.9 | 13.8 | 12.7 | |||
| Adjuvant therapy | No | 75.0 | 68.1 | 0.456 | 52.4 | 71.4 | 0.458 |
| Yes | 25.0 | 31.9 | 47.6 | 28.6 | |||
| Neoadjuvant therapy | No | 70.3 | 77.5 | 0.165 | 64.9 | 82.7 | 0.365 |
| Yes | 29.7 | 22.5 | 35.1 | 17.3 | |||
| MSH6 | Negative | 0 | 2.2 | 0.198 | 3.6 | 0.8 | 0.354 |
| Positive | 100 | 97.8 | 96.4 | 98.2 | |||
| PMS2 | Negative | 7.9 | 4.7 | 0.088 | 7.3 | 4.4 | 0.445 |
| Positive | 92.1 | 95.3 | 92.7 | 95.6 | |||
| Variables | Univariate | Multivariate * | |||||
|---|---|---|---|---|---|---|---|
| n (%) | HR | 95% CI | p Value | HR | 95% | p Value | |
| RAD52 | |||||||
| High | 76.5 | 1.711 | 0.915–3.200 | 0.040 | 1.525 | 0.788–2.950 | 0.046 |
| Low | 23.5 | ||||||
| Sex | |||||||
| Male | 66.1 | 0.912 | 0.549–1.574 | 0.721 | |||
| Female | 33.9 | ||||||
| Age | |||||||
| ≤71 | 46.9 | 1.192 | 0.726–1.951 | 0.484 | |||
| >71 | 53.1 | ||||||
| Tumour stage | |||||||
| T1–2 | 32.9 | 1.962 | 1.118–3.411 | 0.091 | |||
| T3–4 | 67.1 | ||||||
| Node stage | |||||||
| Negative | 51.7 | 1.075 | 0.663–1.741 | 0.077 | |||
| Positive | 48.3 | ||||||
| Metastasis stage | |||||||
| M0 | 94.4 | 4.652 | 1.922–11.260 | 0.001 | 3.215 | 1.265–8.169 | 0.014 |
| M1 | 5.6 | ||||||
| Grade | |||||||
| 1, 2 | 92.7 | 1.003 | 0.403–2.496 | 0.995 | |||
| 3 | 7.3 | ||||||
| Vascular invasion | |||||||
| Absent | 76.6 | 2.659 | 1.577–4.482 | 0.001 | 2.315 | 1.314–4.075 | 0.004 |
| Present | 23.4 | ||||||
| Perineural invasion | |||||||
| Absent | 86.9 | 1.794 | 0.933–3.448 | 0.08 | |||
| Present | 13.1 | ||||||
| Adjuvant therapy | |||||||
| No | 69.7 | 0.482 | 0.222–1.047 | 0.065 | |||
| Yes | 30.3 | ||||||
| Neoadjuvant therapy | |||||||
| No | 75.9 | 0.437 | 0.546–1.682 | 0.588 | |||
| Yes | 24.1 | ||||||
| LN-negative † | 53.8 | 0.869 | 0.025–0.503 | 0.939 | |||
| LN-positive † | 46.2 | 0.473 | 0.029–0.905 | 0.021 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, V.; Chung, L.; Singh, A.; Lea, V.; Abubakar, A.; Lim, S.H.; Chua, W.; Ng, W.; Lee, M.; Roberts, T.L.; et al. Aberrant Expression of RAD52, Its Prognostic Impact in Rectal Cancer and Association with Poor Survival of Patients. Int. J. Mol. Sci. 2020, 21, 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051768
Ho V, Chung L, Singh A, Lea V, Abubakar A, Lim SH, Chua W, Ng W, Lee M, Roberts TL, et al. Aberrant Expression of RAD52, Its Prognostic Impact in Rectal Cancer and Association with Poor Survival of Patients. International Journal of Molecular Sciences. 2020; 21(5):1768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051768
Chicago/Turabian StyleHo, Vincent, Liping Chung, Amandeep Singh, Vivienne Lea, Askar Abubakar, Stephanie H. Lim, Wei Chua, Weng Ng, Mark Lee, Tara L. Roberts, and et al. 2020. "Aberrant Expression of RAD52, Its Prognostic Impact in Rectal Cancer and Association with Poor Survival of Patients" International Journal of Molecular Sciences 21, no. 5: 1768. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051768