Resvega Alleviates Hydroquinone-Induced Oxidative Stress in ARPE-19 Cells

 , , and
, , and 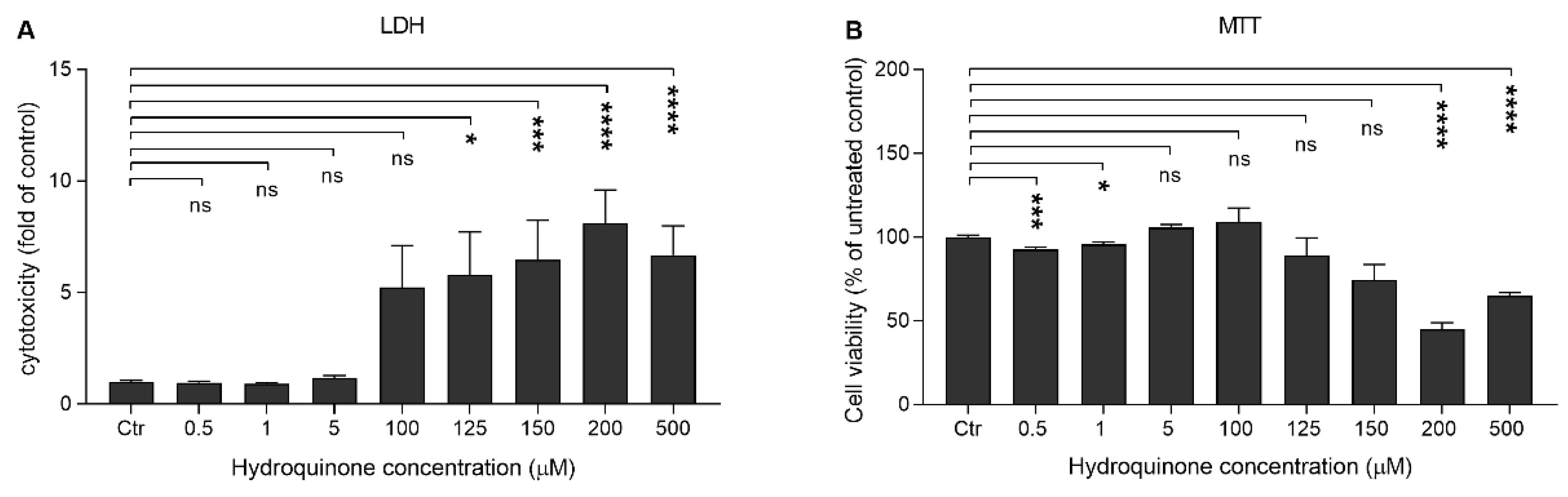{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. High Concentrations of Hydroquinone Reduce the Viability of ARPE-19 Cells
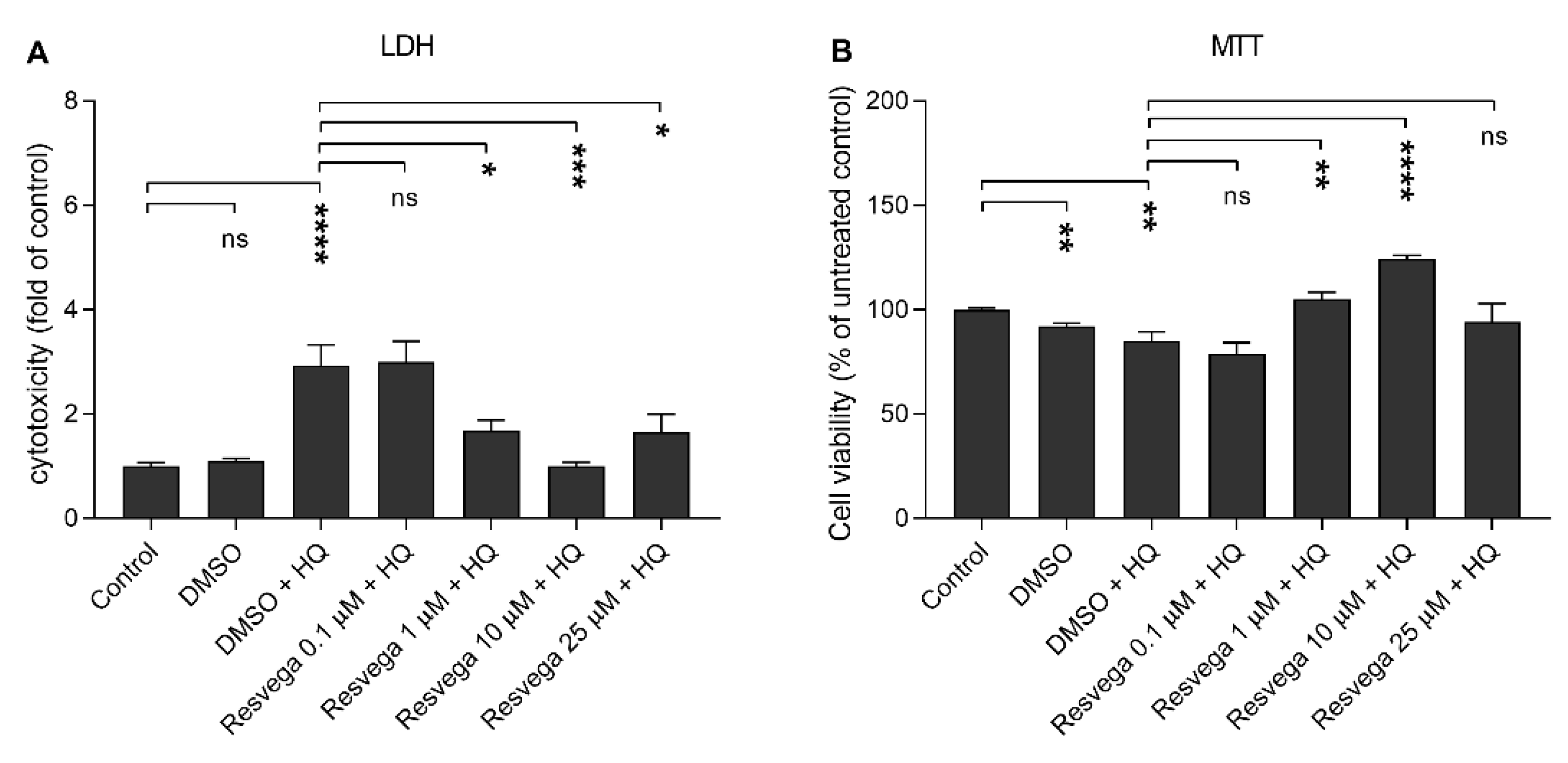
2.2. Resvega Alleviates Hydroquinone-Induced Cytotoxicity
2.3. Resvega Reduces IL-8 and MCP-1 Release but Enhances IL-6 in Comparison to RPE Cells Treated with Hydroquinone Only
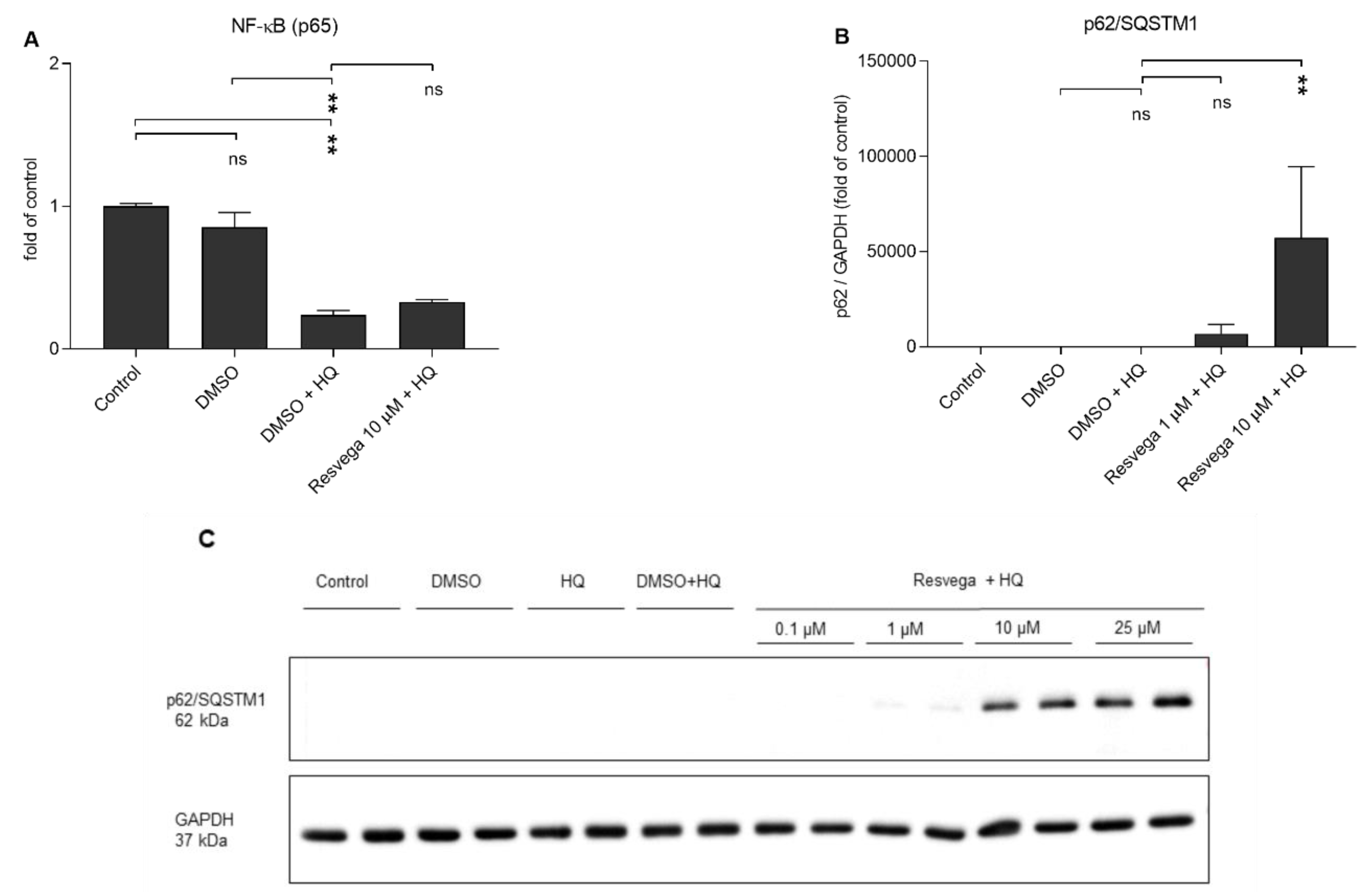
2.4. Hydroquinone Reduces the Activivity of NF-κB and Resvega Increases the Levels of p62/SQSTM1 (p62) Protein
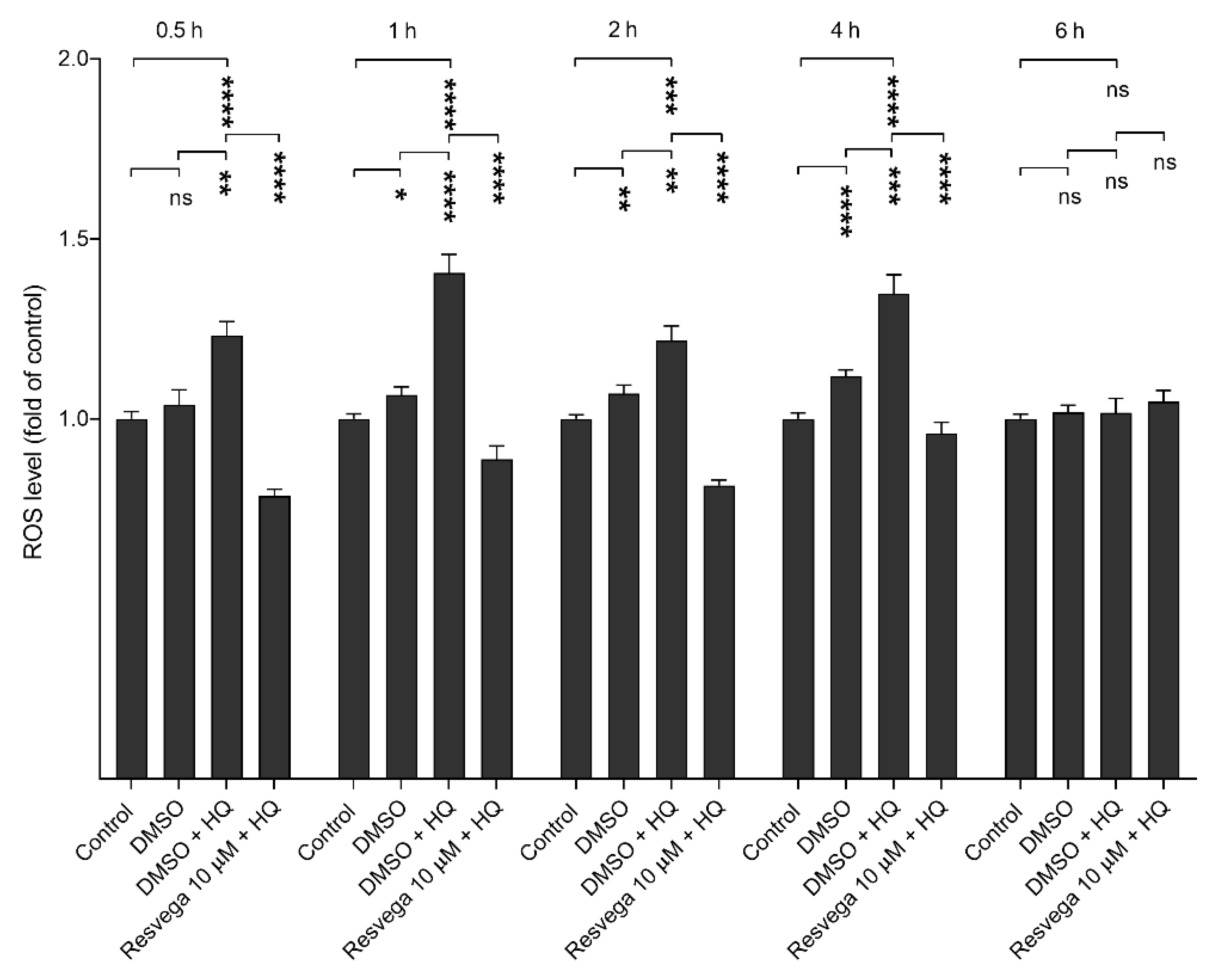
2.5. Resvega Prevents Hydroquinone-Induced ROS Production
2.6. Antioxidants NAC and APDC Prevent the Hydroquinone-Induced ROS Production
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Stimulations and Treatments
4.3. Cell Viability Assays
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blot
4.6. ROS Detection
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gehrs, K.M.; Anderson, D.H.; Johnson, L.V.; Hageman, G.S. Age-Related Macular Degeneration-Emerging Pathogenetic and Therapeutic Concepts. Ann. Med. 2006, 38, 450–471. [Google Scholar] [CrossRef]
- Pennington, K.L.; DeAngelis, M.M. Epidemiology of Age-Related Macular Degeneration (AMD): Associations with Cardiovascular Disease Phenotypes and Lipid Factors. Eye Vis. (Lond.) 2016, 3, 34–54. [Google Scholar] [CrossRef] [Green Version]
- Kaarniranta, K.; Sinha, D.; Blasiak, J.; Kauppinen, A.; Veréb, Z.; Salminen, A.; Boulton, M.E.; Petrovski, G. Autophagy and Heterophagy Dysregulation Leads to Retinal Pigment Epithelium Dysfunction and Development of Age-Related Macular Degeneration. Autophagy 2013, 9, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its Role in Age-Related Macular Degeneration. Cellular and molecular life sciences. Cell. Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y. Suppressive Effect of Hydroquinone, a Benzene Metabolite, on in Vitro Inflammatory Responses Mediated by Macrophages, Monocytes, and Lymphocytes. Mediat. Inflamm. 2008, 2008, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, K.M.; Baglole, C.J.; Phipps, R.P.; Libby, R.T. Molecular Regulation of Cigarette Smoke Induced-Oxidative Stress in Human Retinal Pigment Epithelial Cells: Implications for Age-Related Macular Degeneration. Am. J. Physiol. Cell Physiol. 2009, 297, 1200–1210. [Google Scholar] [CrossRef] [Green Version]
- Kauppinen, A.; Niskanen, H.; Suuronen, T.; Kinnunen, K.; Salminen, A.; Kaarniranta, K. Oxidative Stress Activates NLRP3 Inflammasomes in ARPE-19 Cells-Implications for Age-Related Degeneration (AMD). Immunol. Lett. 2012, 147, 29–33. [Google Scholar] [CrossRef]
- Klein, R.; Knudtson, M.D.; Cruickshanks, K.J.; Klein, B.E.K. Further Observations on the Association between Smoking and the Long-Term Incidence and Progression of Age-Related Macular Degeneration: The Beaver Dam Eye Study. Arch. Ophthalmol. 2008, 126, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Hammond, B.R.; Wooten, B.R.; Snodderly, D.M. Cigarette Smoking and Retinal Carotenoids: Implications for Age-Related Macular Degeneration. Vision Res. 1996, 36, 3003–3009. [Google Scholar] [CrossRef] [Green Version]
- Talhout, R.; Schulz, T.; Florek, E.; van Benthem, J.; Wester, P.; Opperhuizen, A. Hazardous Compounds in Tobacco Smoke. Int. J. Environ. Res. Public Health 2011, 8, 613–628. [Google Scholar] [CrossRef]
- Badham, H.J.; Renaud, S.J.; Wan, J.; Winn, L.M. Benzene-Initiated Oxidative Stress: Effects on Embryonic Signaling Pathways. Chem. Biol. Interact. 2010, 184, 218–221. [Google Scholar] [CrossRef]
- Bolton, J.L.; Trush, M.A.; Penning, T.M.; Dryhurst, G.; Monks, T.J. Role of Quinones in Toxicology. Chem Res. Toxicol. 2000, 13, 135–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, G.; Han, Q.; Wang, J.; Suriguga; Li, Y.; Yu, C.; Li, Y.; Yi, Z. Phenolic Metabolites of Benzene Induced Caspase-dependent Cytotoxicities to K562 Cells Accompanied with Decrease in Cell Surface Sialic Acids. Environ. Toxicol. 2014, 29, 1437–1451. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.C.; Suriguga, Y.L.; Ran, L.Y.; Tang, K.Y.; Liang, J.; Chun, Y.Z. The Role of ROS in Hydroquinone-Induced Inhibition of K562 Cell Erythroid Differentiation. Biomed. Environ. Sci. 2014, 27, 212–214. [Google Scholar] [CrossRef]
- Kim, E.; Kang, B.Y.; Kim, T.S. Inhibition of Interleukin-12 Production in Mouse Macrophages by Hydroquinone, a Reactive Metabolite of Benzene, Via Suppression of Nuclear Factor- ᴋB Binding Activity. Immunol. Lett. 2005, 99, 24–29. [Google Scholar] [CrossRef]
- Moustafa, M.T.; Ramirez, C.; Schneider, K.; Atilano, S.R.; Limb, G.A.; Kuppermann, B.D.; Kenney, M.C. Protective Effects of Memantine on Hydroquinone-Treated Human Retinal Pigment Epithelium Cells and Human Retinal Müller Cells. J. Ocul. Pharmacol. Ther. 2017, 33, 61–619. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, C.; Cáceres-del-Carpio, J.; Chu, J.; Chu, J.; Moustafa, M.T.; Chwa, M.; Limb, G.A.; Kuppermann, B.D.; Kenney, M.C. Brimonidine can Prevent in Vitro Hydroquinone Damage on Retinal Pigment Epithelium Cells and Retinal Müller Cells. J. Ocul. Pharmacol. Ther. 2016, 32, 12–108. [Google Scholar] [CrossRef]
- Strunnikova, N.; Zhang, C.; Teichberg, D.; Cousins, S.W.; Baffi, J.; Becker, K.G.; Csaky, K.G. Survival of Retinal Pigment Epithelium After Exposure to Prolonged Oxidative Injury: A Detailed Gene Expression and Cellular Analysis. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3767–3777. [Google Scholar] [CrossRef] [Green Version]
- Alcazar, O.; Hawkridge, A.M.; Collier, T.S.; Cousins, S.W.; Bhattacharya, S.K.; Muddiman, D.C.; Marin-Castano, M.E. Proteomics Characterization of Cell Membrane Blebs in Human Retinal Pigment Epithelium Cells. Mol. Cell. Proteom. 2009, 8, 2201–2211. [Google Scholar] [CrossRef] [Green Version]
- Pons, M.; Cousins, S.W.; Csaky, K.G.; Striker, G.; Marin-Castaño, M.E. Cigarette Smoke-Related Hydroquinone Induces Filamentous Actin Reorganization and Heat Shock Protein 27 Phosphorylation through p38 and Extracellular Signal-Regulated Kinase 1/2 in Retinal Pigment Epithelium: Implications for Age-Related Macular Degeneration. Am. J. Pathol. 2010, 177, 1198–1213. [Google Scholar] [CrossRef]
- Sharma, A.; Patila Jayaprakash, A.; Gramajo, A.L.; Seigel, G.M.; Kuppermann, B.D.; Kenney Cristina, M. Effects of Hydroquinone on Retinal and Vascular Cells in Vitro. Indian J. Ophthalmol. 2012, 60, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Koskela, A.; Reinisalo, M.; Petrovski, G.; Sinha, D.; Olmiere, C.; Karjalainen, R.; Kaarniranta, K. Nutraceutical with Resveratrol and Omega-3 Fatty Acids Induces Autophagy in ARPE-19 Cells. Nutrients 2016, 8, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querques, G.; Forte, R.; Souied, E.H. Retina and Omega-3. J. Nutr. Metab. 2011, 2011, 748361-12. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.M.; Cho, I.J.; Kim, S.G. Resveratrol Protects Mitochondria Against Oxidative Stress through AMP-Activated Protein Kinase-Mediated Glycogen Synthase Kinase-3 Inhibition Downstream of Poly(ADP-Ribose)Polymerase-LKB1 Pathway. Mol. Pharmacol. 2009, 76, 884–895. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.S.; Xia, C.; Jiang, B.; Stinefelt, B.; Klandorf, H.; Harris, G.K.; Shi, X. Resveratrol Scavenges Reactive Oxygen Species and Effects Radical-Induced Cellular Responses. Biochem. Biophys. Res. Commun. 2003, 309, 1017–1026. [Google Scholar] [CrossRef]
- Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. The Role of Oxidative Stress in the Pathogenesis of Age-Related Macular Degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Walker, G.B.; Wang, X.; Cui, J.Z.; Matsubara, J.A. Altered Cytokine Profiles of Human Retinal Pigment Epithelium: Oxidant Injury and Replicative Senescence. Mol. Vis. 2013, 19, 718–728. [Google Scholar]
- López-García, J.; Lehocký, M.; Humpolíček, P.; Sáha, P. HaCaT Keratinocytes Response on Antimicrobial Atelocollagen Substrates: Extent of Cytotoxicity, Cell Viability and Proliferation. J. Funct. Biomater. 2014, 5, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Yang, E.; Kim, I. Hydroquinone-Induced Apoptosis of Human Lymphocytes through Caspase 9/3 Pathway. Mol. Biol. Rep. 2012, 39, 6737–6743. [Google Scholar] [CrossRef]
- Lee, M.H.; Chung, S.W.; Kang, B.Y.; Kim, K.-; Kim, T.S. Hydroquinone, a Reactive Metabolite of Benzene, Enhances Interleukin-4 Production in CD4+ T Cells and Increases Immunoglobulin E Levels in Antigen-primed Mice. Immunology 2002, 106, 496–502. [Google Scholar] [CrossRef]
- Bunt, S.K.; Clements, V.K.; Hanson, E.M.; Sinha, P.; Ostrand-Rosenberg, S. Inflammation Enhances Myeloid-Derived Suppressor Cell Cross-Talk by Signaling through Toll-Like Receptor 4. J. Leukoc. Biol. 2009, 85, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, F.; Tang, M.; Yuan, M.; Hu, A.; Zhan, Z.; Li, Z.; Li, J.; Ding, X.; Lu, L. Macrophage Polarization in Experimental and Clinical Choroidal Neovascularization. Sci. Rep. 2016, 6, 30933. [Google Scholar] [CrossRef]
- Luzina, I.G.; Keegan, A.D.; Heller, N.M.; Rook, G.A.W.; Shea-Donohue, T.; Atamas, S.P. Regulation of Inflammation by Interleukin-4: A Review of “Alternatives”. J. Leukoc. Biol. 2012, 92, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, A.M.; Clarke, D.L.; Bradbury, D.A.; Corbett, L.M.; Patel, J.A.; Knox, A.J. Transcriptional Regulation of Monocyte Chemotactic Protein-1 Release by Endothelin-1 in Human Airway Smooth Muscle Cells Involves NF-κB and AP-1. Br. J. Pharmacol. 2009, 157, 436–450. [Google Scholar] [CrossRef] [Green Version]
- Jundi, K.; Greene, C.M. Transcription of Interleukin-8: How Altered Regulation can Affect Cystic Fibrosis Lung Disease. Biomolecules 2015, 5, 1386–1398. [Google Scholar] [CrossRef]
- Ebrahem, Q.; Minamoto, A.; Hoppe, G.; Anand-Apte, B.; Sears, J.E. Triamcinolone Acetonide Inhibits IL-6- and VEGF-Induced Angiogenesis Downstream of the IL-6 and VEGF Receptors. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4935. [Google Scholar] [CrossRef]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of Interleukin (IL)-6-Type Cytokine Signalling and its Regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Mendonça Torres, P.M.; de Araujo, E.G. Interleukin-6 Increases the Survival of Retinal Ganglion Cells in Vitro. J. Neuroimmunol. 2001, 117, 43–50. [Google Scholar] [CrossRef]
- Marasco, M.R.; Conteh, A.M.; Reissaus, C.A.; Cupit, J.E.; Appleman, E.M.; Mirmira, R.G.; Linnemann, A.K. Interleukin-6 Reduces Β-Cell Oxidative Stress by Linking Autophagy with the Antioxidant Response. Diabetes 2018, 67, 1576–1588. [Google Scholar] [CrossRef] [Green Version]
- Katsuragi, Y.; Ichimura, Y.; Komatsu, M. Regulation of the Keap1–Nrf2 Pathway by p62/ SQSTM1. Curr. Opin. Toxicol. 2016, 1, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Wruck, C.J.; Streetz, K.; Pavic, G.; Götz, M.E.; Tohidnezhad, M.; Brandenburg, L.; Varoga, D.; Eickelberg, O.; Herdegen, T.; Trautwein, C.; et al. Nrf2 Induces Interleukin-6 (IL-6) Expression Via an Antioxidant Response Element within the IL-6 Promoter. J. Biol. Chem. 2011, 286, 4493–4499. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cano, M.; Handa, J.T. P62 Provides Dual Cytoprotection Against Oxidative Stress in the Retinal Pigment Epithelium. Biochim. Biophys. Acta 1843, 1248–1258. [Google Scholar] [CrossRef] [Green Version]
- Tsujinaka, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Ota, H.; Takeda, M.; Fujimura, T.; Takasawa, S.; Ogata, N. Human Retinal Pigment Epithelial Cell Proliferation by the Combined Stimulation of Hydroquinone and Advanced Glycation End-Products Via Up-Regulation of VEGF Gene. Biochem. Biophys. Rep. 2015, 2, 123–131. [Google Scholar] [CrossRef]
- Piippo, N.; Korhonen, E.; Hytti, M.; Kinnunen, K.; Kaarniranta, K.; Kauppinen, A. Oxidative Stress is the Principal Contributor to Inflammasome Activation in Retinal Pigment Epithelium Cells with Defunct Proteasomes and Autophagy. Cell Physiol. Biochem. 2018, 49, 359–367. [Google Scholar] [CrossRef]
- Koskela, A.; Reinisalo, M.; Hyttinen, J.M.T.; Kaarniranta, K.; Karjalainen, R.O. Pinosylvin-Mediated Protection Against Oxidative Stress in Human Retinal Pigment Epithelial Cells. Mol. Vis. 2014, 20, 760–769. [Google Scholar]
- Wei, W.; Li, L.; Zhang, Y.; Geriletu; Yang, J.; Zhang, Y.; Xing, Y. Vitamin C Protected Human Retinal Pigmented Epithelium from Oxidant Injury Depending on Regulating SIRT1. Sci. World J. 2014, 2014, 750634–750638. [Google Scholar] [CrossRef]
- Hutnik, C.; Liu, H.; Kagan, D.B. Efficacy of various Antioxidants in the Protection of the Retinal Pigment Epithelium from Oxidative Stress. Clin. Ophthalmol. 2012, 6, 1471–1476. [Google Scholar] [CrossRef] [Green Version]
- Lan, F.; Weikel, K.A.; Cacicedo, J.M.; Ido, Y. Resveratrol-Induced AMP-Activated Protein Kinase Activation is Cell-Type Dependent: Lessons from Basic Research for Clinical Application. Nutrients 2017, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Dikalov, S. Crosstalk between Mitochondria and NADPH Oxidases. Free Radic. Biol. Med. 2011, 51, 1289–1301. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Liu, Y.; Ojcius, D.M.; Lee, C.; Chen, R.; Huang, P.; Martel, J.; Young, J.D. Mineral Particles Stimulate Innate Immunity through Neutrophil Extracellular Traps Containing HMGB1. Sci. Rep. 2017, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.V.C.; Lardy, B.; Rousset, F.; Hazane-Puch, F.; Zhang, L.; Trocmé, C.; Serrander, L.; Krause, K.; Morel, F. Quinone Compounds Regulate the Level of ROS Production by the NADPH Oxidase Nox4. Biochem. Pharmacol. 2013, 85, 1644–1654. [Google Scholar] [CrossRef]
- Pyatt, D.W.; Stillman, W.S.; Irons, R.D. Hydroquinone, a Reactive Metabolite of Benzene, Inhibits NF-κB in Primary Human CD4+T Lymphocytes. Toxicol. Appl. Pharmacol. 1998, 149, 178–184. [Google Scholar] [CrossRef]
- World Health Organization; United Nations Environment Programme; International Labour Organization. Hydroquinone Health and Safety Guide, Health and Safety Guide No. 101; World Health Organization: Geneva, Switzerland, 1996; p. 11. [Google Scholar]
- The Age-Related Eye Disease Study (AREDS): Design Implications. AREDS Report no. 1. Control. Clin. Trials 1999, 20, 573–600. [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Sangiovanni, J.P. Lutein + Zeaxanthin and Omega-3 Fatty Acids for Age-Related Macular Degeneration: The Age-Related Eye Disease Study 2 (AREDS2) Randomized Clinical Trial. JAMA 2013, 309. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.; Xu, C.; Lu, Y.; Hu, H.; Qin, B.; Wang, Y.; He, D.; Li, C.; Yu, X.; et al. MitoTEMPO Prevents Oxalate Induced Injury in NRK-52E Cells Via Inhibiting Mitochondrial Dysfunction and Modulating Oxidative Stress. Oxid. Med. Cell. Longev. 2017, 7528090–7528099. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, H.; He, D.; Wang, Y.; Cai, B.; Chen, J.; Ma, J.; Liu, Z.; Wu, Y. Retinal Pigment Epithelium Cell Death is Associated with NLRP3 Inflammasome Activation by all-Trans Retinal. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3034–3045. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattarai, N.; Korhonen, E.; Toppila, M.; Koskela, A.; Kaarniranta, K.; Mysore, Y.; Kauppinen, A. Resvega Alleviates Hydroquinone-Induced Oxidative Stress in ARPE-19 Cells. Int. J. Mol. Sci. 2020, 21, 2066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062066
Bhattarai N, Korhonen E, Toppila M, Koskela A, Kaarniranta K, Mysore Y, Kauppinen A. Resvega Alleviates Hydroquinone-Induced Oxidative Stress in ARPE-19 Cells. International Journal of Molecular Sciences. 2020; 21(6):2066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062066
Chicago/Turabian StyleBhattarai, Niina, Eveliina Korhonen, Maija Toppila, Ali Koskela, Kai Kaarniranta, Yashavanthi Mysore, and Anu Kauppinen. 2020. "Resvega Alleviates Hydroquinone-Induced Oxidative Stress in ARPE-19 Cells" International Journal of Molecular Sciences 21, no. 6: 2066. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062066