STIP1/HOP Regulates the Actin Cytoskeleton through Interactions with Actin and Changes in Actin-Binding Proteins Cofilin and Profilin

 , , , , , , and
, , , , , , and 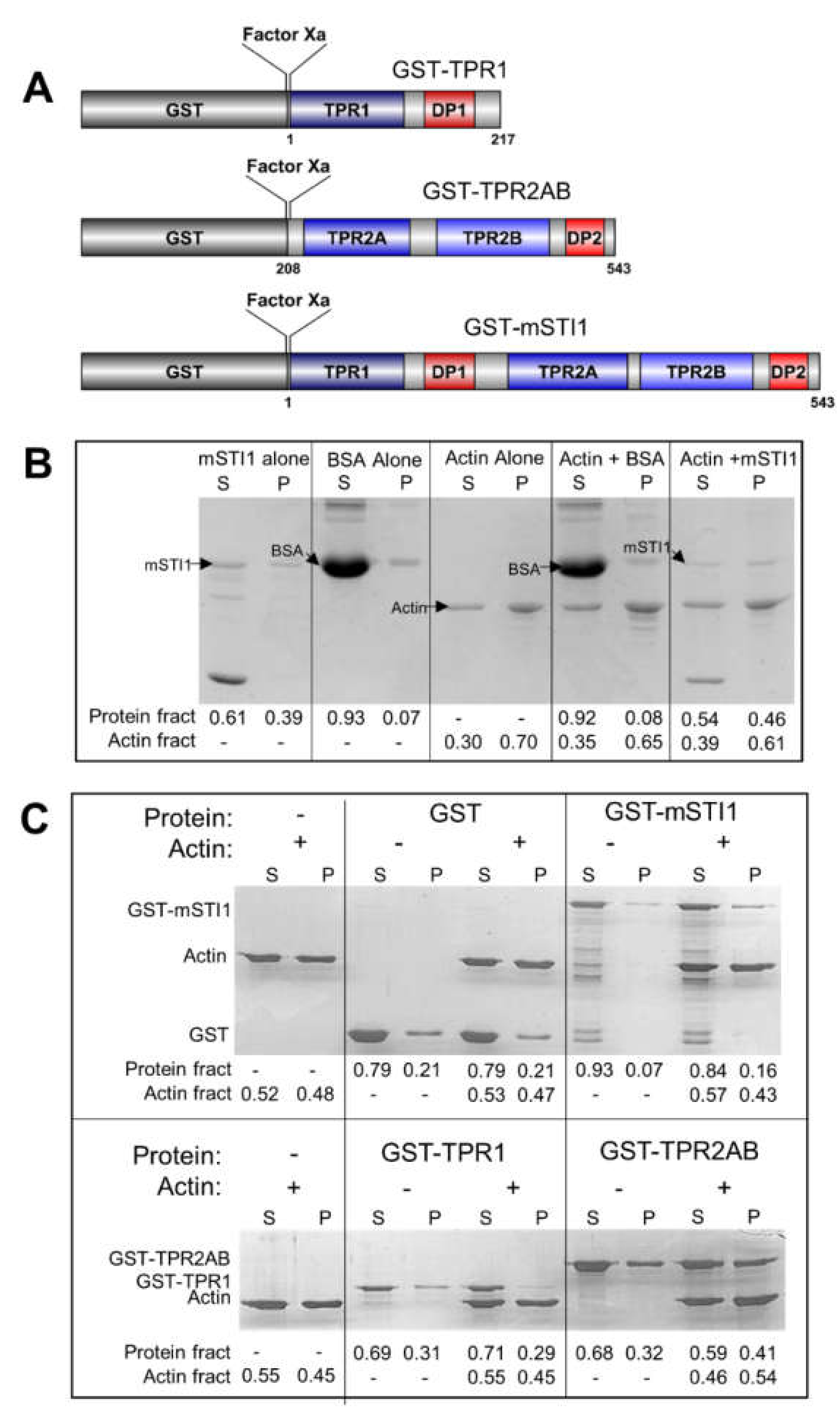{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
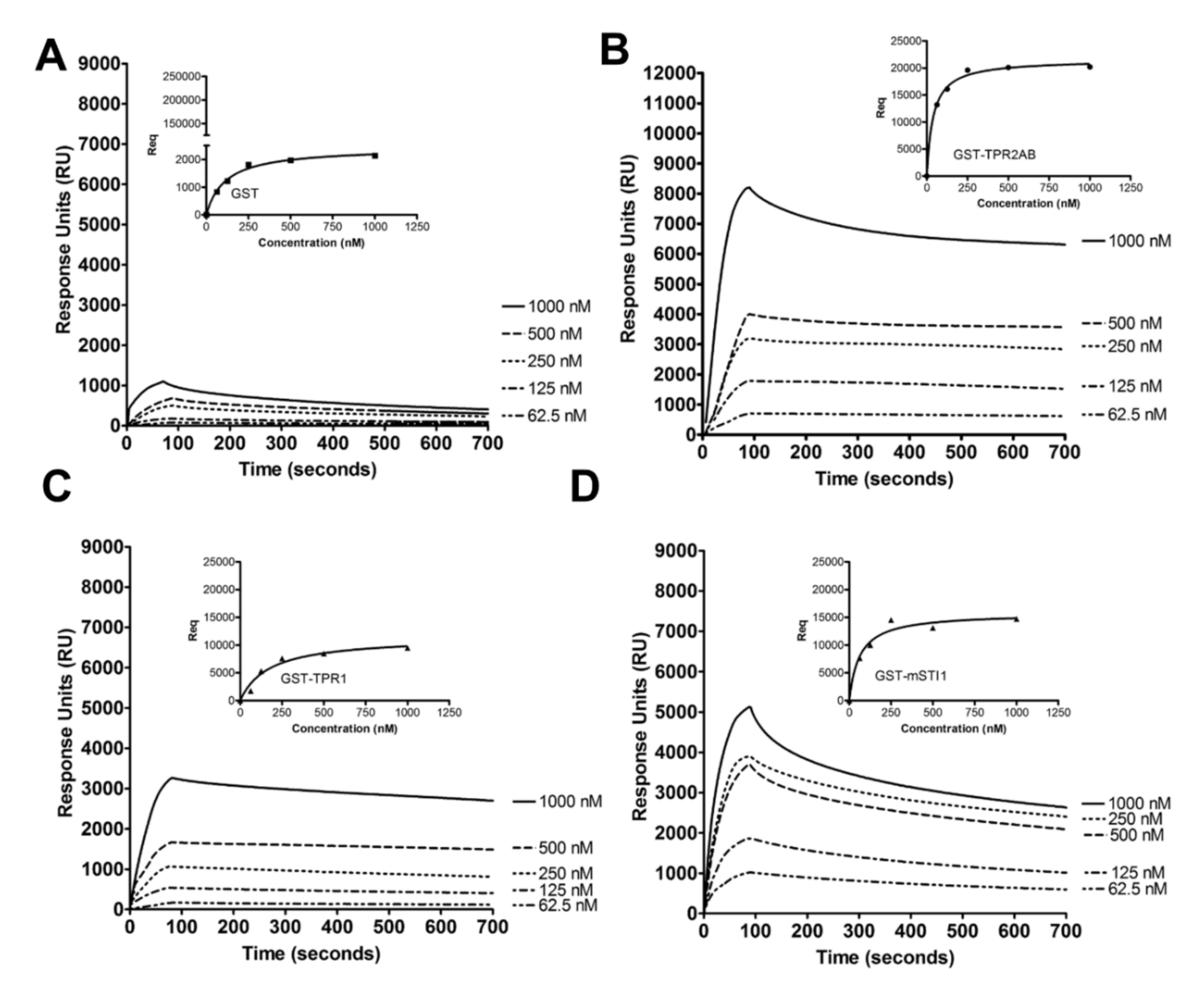
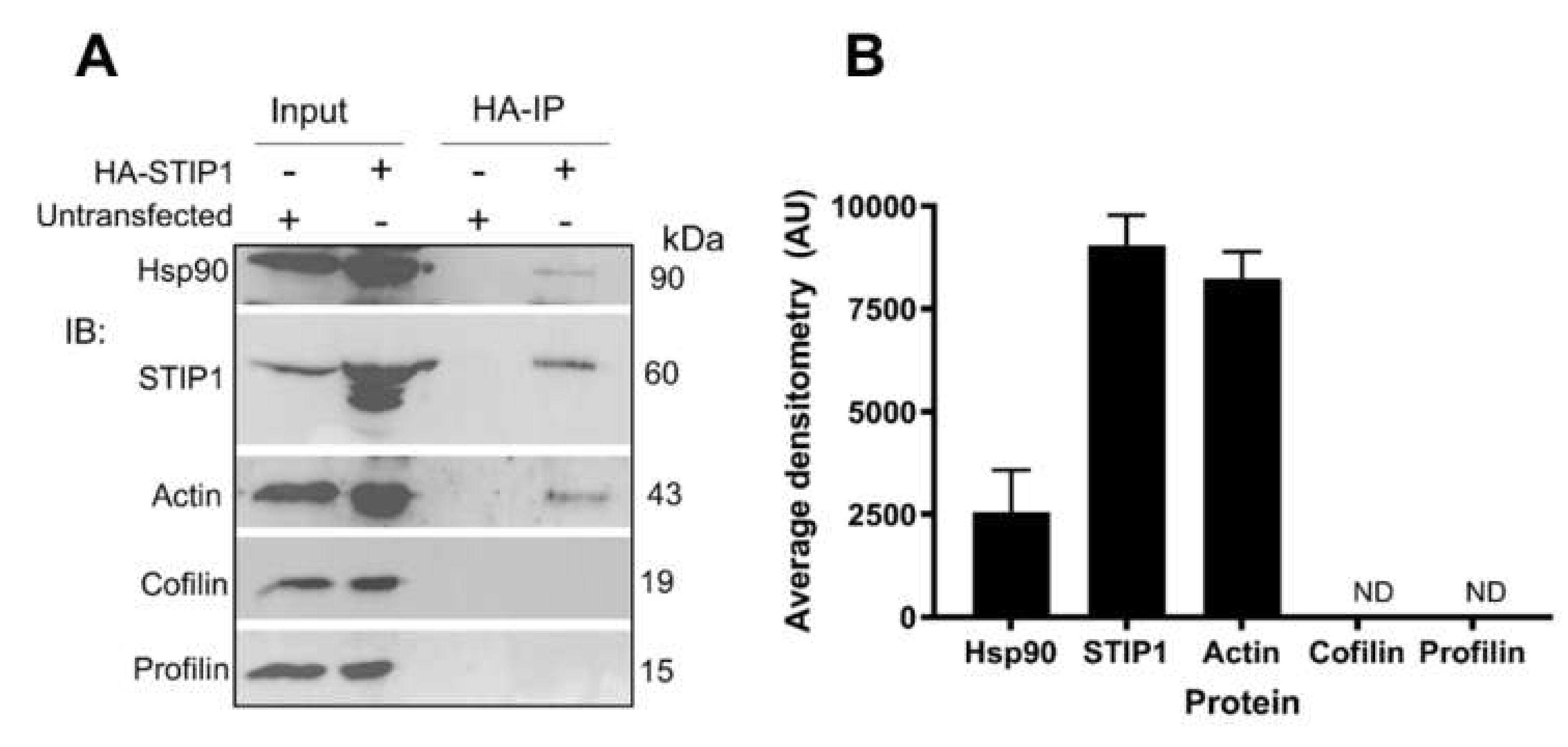
2.1. mSTI1 Interacts with F-actin via the TPR2AB Domain
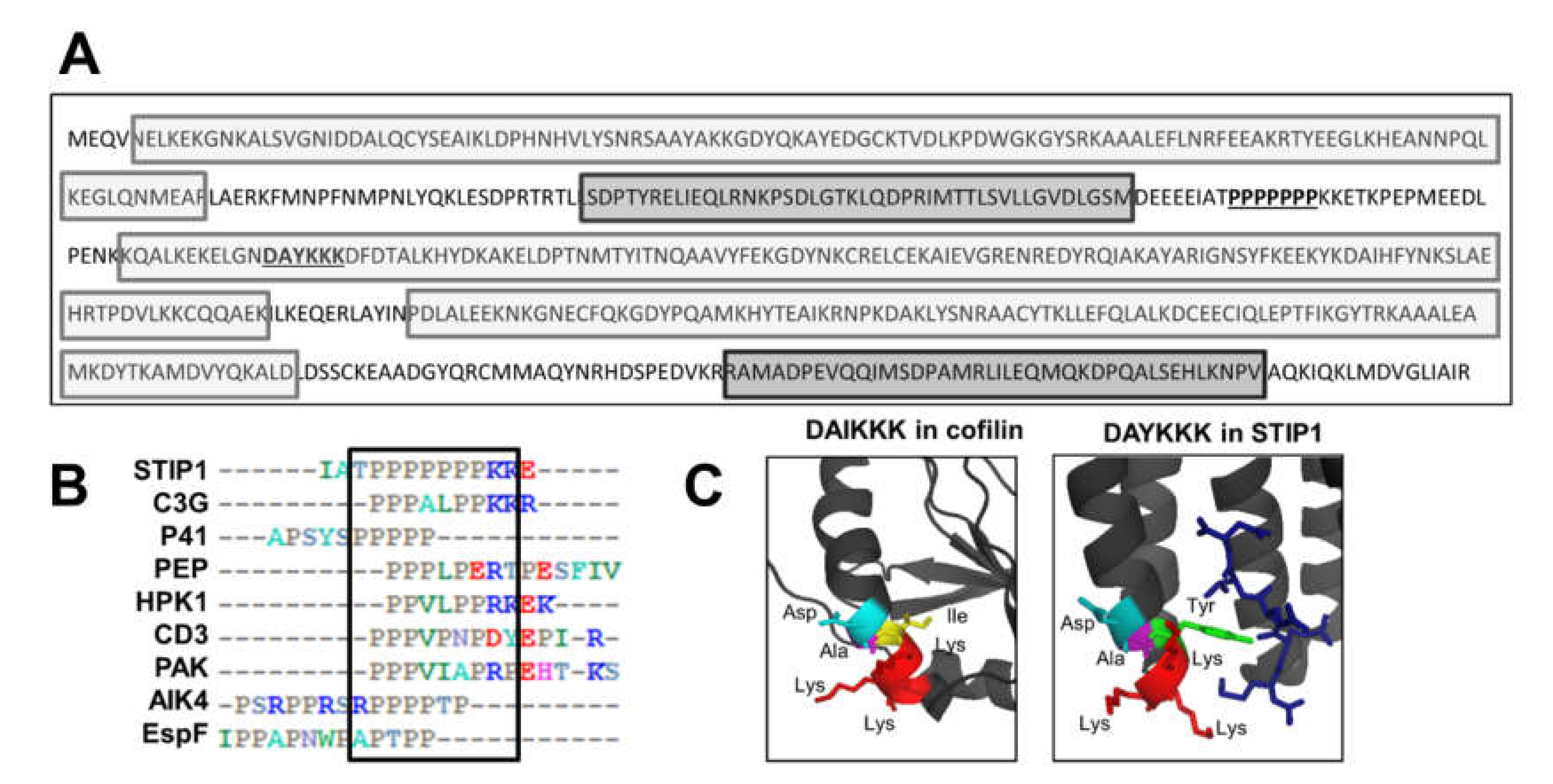
2.2. Putative Actin-Binding Sites in the STIP1 Sequence
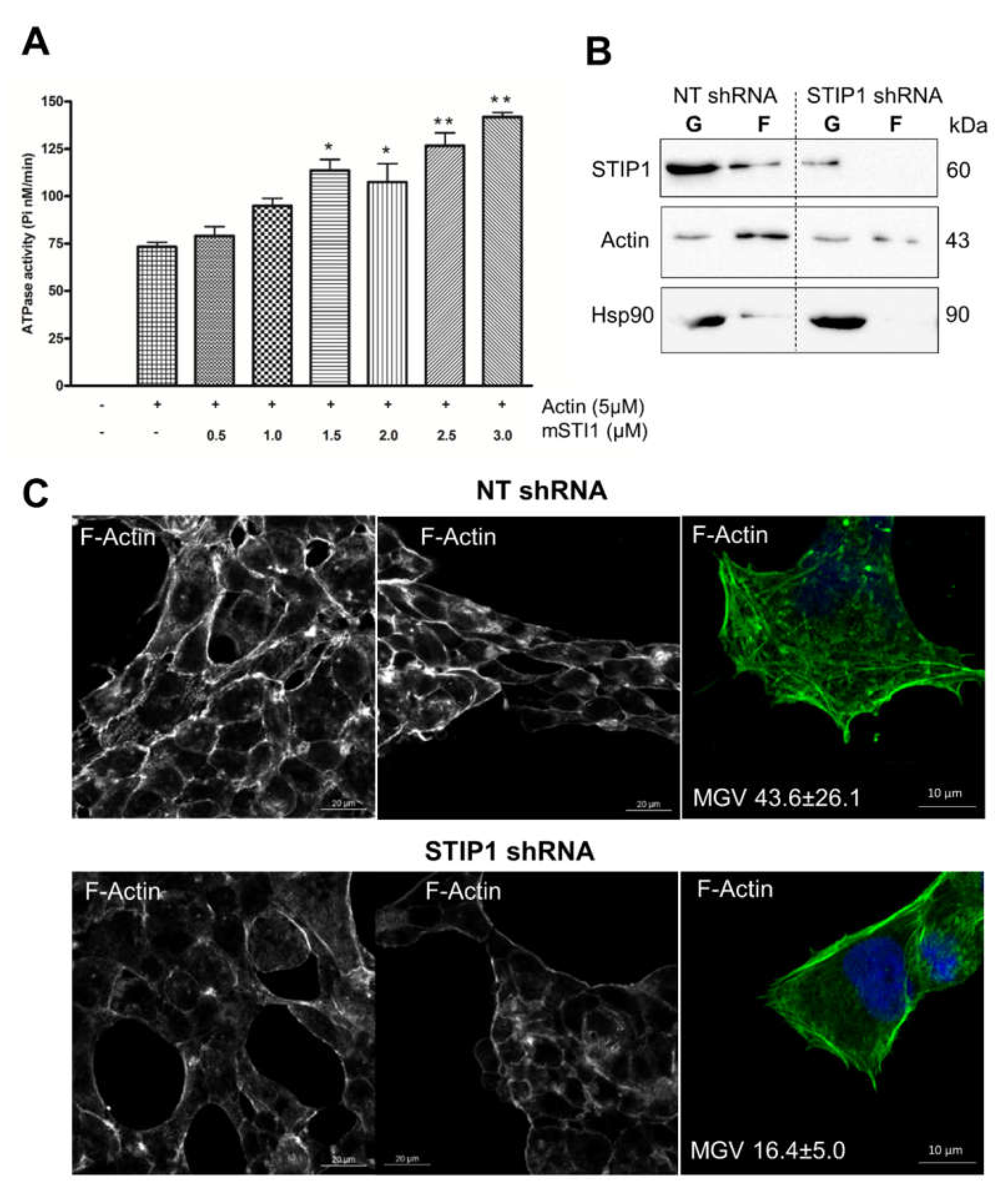
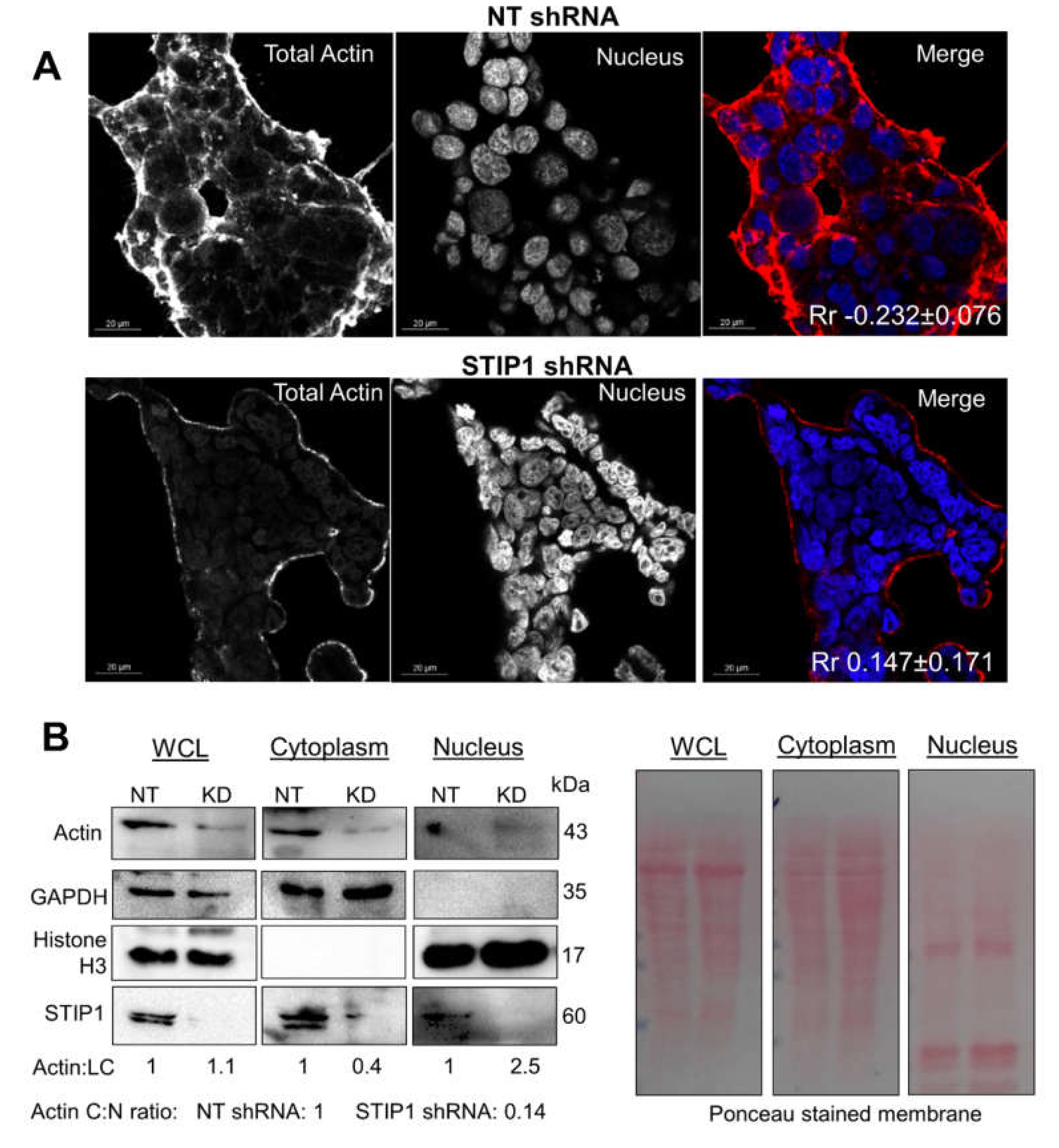
2.3. STIP1 Increases Actin ATPase In Vitro but STIP1 Depletion in Cells Reduces F-Actin
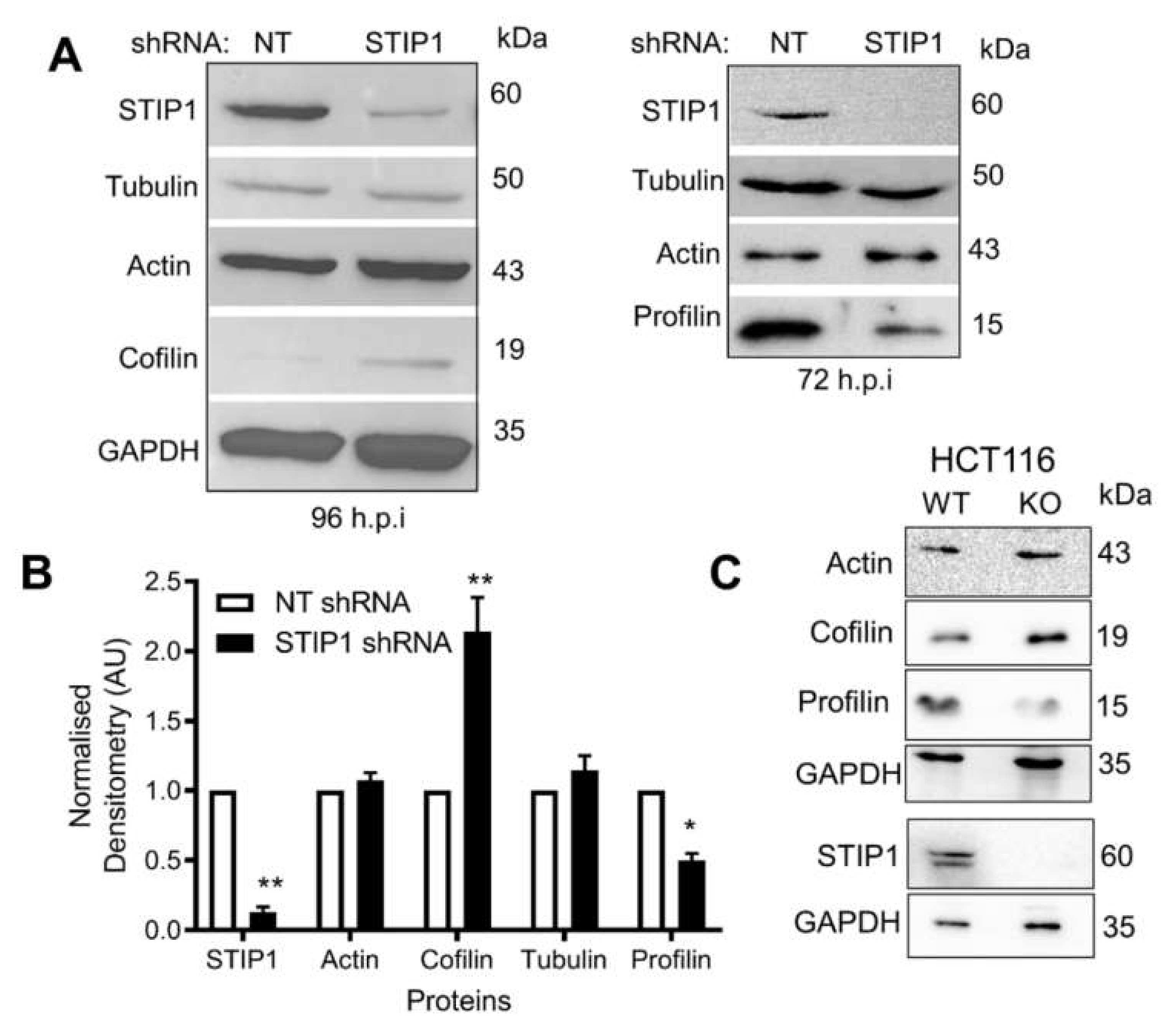
2.4. STIP1 Depletion Alters the Ratio of Actin-Binding Proteins Cofilin and Profilin
3. Materials and Methods
3.1. Expression and Purification of GST-Tagged mSTI1 Proteins
3.2. SDS-PAGE and Western Blotting
3.3. F-Actin Formation, Co-Sedimentation and Bundling Assay
3.4. ATPase Assay
3.5. Surface Plasmon Resonance Spectroscopy
3.6. Maintenance of Cell Lines
3.7. Immunofluorescence and Confocal Microscopy
3.8. Transfection and HA-Immunoprecipitation
3.9. Fractionation of G- and F-Actin from Cell Lysates
3.10. Cytoplasmic and Nuclear Fractionation
3.11. Statistical Analysis and Reproducibility
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yamaguchi, H.; Condeelis, J. Regulation of the actin cytoskeleton in cancer cell migration and invasion. Biochim. Biophys. Acta 2007, 1773, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaffer, C.L.; Weinberg, R. A Perspective on Cancer Cell Metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Woodham, E.F.; Machesky, L.M. Polarised cell migration: Intrinsic and extrinsic drivers. Curr. Opin. Cell Biol. 2014, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.D.; Gerlach, B.D. The roles and regulation of the actin cytoskeleton, intermediate filaments and microtubules in smooth muscle cell migration. Respir. Res. 2017, 18, 54. [Google Scholar] [CrossRef] [Green Version]
- Guasch, R.M.; Scambler, P.; Jones, G.E.; Ridley, A.J. RhoE Regulates Actin Cytoskeleton Organization and Cell Migration. Mol. Cell. Biol. 1998, 18, 4761–4771. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Rotty, J.D.; Bear, J.E. Competition and collaboration between different actin assembly pathways allows for homeostatic control of the actin cytoskeleton. Bioarchitecture 2015, 5, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, J.-M.; Wanger, M.; Keiser, T.; Wegner, A. Treadmilling of actin. J. Muscle Res. Cell Motil. 1983, 4, 507–527. [Google Scholar] [CrossRef]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Narita, A. Minimum requirements for the actin-like treadmilling motor system. Bioarchitecture 2011, 1, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, I.; Vavylonis, D.; Pollard, T.D. Polymerization kinetics of ADP-and ADP-Pi-actin determined by fluorescence microscopy. Proc. Nat. Acad. Sci. 2007, 104, 8827–8832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paavilainen, V.O.; Bertling, E.; Falck, S.; Lappalainen, P. Regulation of cytoskeletal dynamics by actin-monomer-binding proteins. Trends Cell Biol. 2004, 14, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Ressad, F.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D.; Carlier, M.F. Kinetic analysis of the interaction of actin-depolymerizing factor (ADF)/cofilin with G- and F-actins. Comparison of plant and human ADFs and effect of phosphorylation. J. Biol. Chem. 1998, 273, 20894–20902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ampe, C.; Markey, F.; Lindberg, U.; Vandekerckhove, J. The primary structure of human platelet profilin: Reinvestigation of the calf spleen profilin sequence. FEBS Lett. 1988, 228, 17–21. [Google Scholar] [CrossRef]
- Craig, S.W.; Pollard, T.D. Actin-binding proteins. Trends Biochem. Sci. 1982, 7, 88–92. [Google Scholar] [CrossRef]
- Griffith, L.M.; Pollard, T.D. The interaction of actin filaments with microtubules and microtubule-associated proteins. J. Biol. Chem. 1982, 257, 9143–9151. [Google Scholar] [PubMed]
- Dominguez, R. Actin-binding proteins—A unifying hypothesis. Trends Biochem. Sci. 2004, 29, 572–578. [Google Scholar] [CrossRef]
- Dopie, J.; Skarp, K.-P.; Rajakylä, E.K.; Tanhuanpää, K.; Vartiainen, M.K. Active maintenance of nuclear actin by importin 9 supports transcription. Proc. Nat. Acad. Sci. 2012, 109, E544–E552. [Google Scholar] [CrossRef] [Green Version]
- Shekhar, S.; Carlier, M.-F. Enhanced depolymerization of actin filaments by ADF/cofilin and monomer funneling by capping protein cooperate to accelerate barbed-end growth. Curr. Biol. 2017, 27, 1990–1998. [Google Scholar] [CrossRef] [Green Version]
- Pantaloni, D.; Carlier, M.-F. How profilin promotes actin filament assembly in the presence of thymosin β4. Cell 1993, 75, 1007–1014. [Google Scholar] [CrossRef]
- Andrianantoandro, E.; Pollard, T.D. Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/Cofilin. Mol. Cell 2006, 24, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, D.; Muhlrad, A.; Cooper, J.; Wear, M.; Reisler, E. Actin filament severing by cofilin. J. Mol. Biol. 2007, 365, 1350–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Eddy, R.; Condeelis, J. The cofilin pathway in breast cancer invasion and metastasis. Nat. Rev. Cancer 2007, 7, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Janke, J.; Schlüter, K.; Jandrig, B.; Theile, M.; Kölble, K.; Arnold, W.; Grinstein, E.; Schwartz, A.; Estevéz-Schwarz, L.; Schlag, P.M.; et al. Suppression of tumorigenicity in breast cancer cells by the microfilament protein profilin 1. J. Exp. Med. 2002, 191, 1675–1686. [Google Scholar] [CrossRef] [Green Version]
- Roy, P.; Jacobson, K. Overexpression of profilin reduces the migration of invasive breast cancer cells. Cell Motil. Cytoskelet. 2004, 57, 84–95. [Google Scholar] [CrossRef]
- Ding, Z.; Lambrechts, A.; Parepally, M.; Roy, P. Silencing profilin-1 inhibits endothelial cell proliferation, migration and cord morphogenesis. J. Cell Sci. 2006, 119, 4366. [Google Scholar] [CrossRef] [Green Version]
- Frantzi, M.; Klimou, Z.; Makridakis, M.; Zoidakis, J.; Latosinska, A.; Borràs, D.M.; Janssen, B.; Giannopoulou, I.; Lygirou, V.; Lazaris, A.C.; et al. Silencing of profilin-1 suppresses cell adhesion and tumor growth via predicted alterations in integrin and Ca2+ signaling in T24M-based bladder cancer models. Oncotarget 2016, 7, 70750–70768. [Google Scholar] [CrossRef] [Green Version]
- Vartiainen, M.K.; Guettler, S.; Larijani, B.; Treisman, R. Nuclear actin regulates dynamic subcellular localization and activity of the SRF cofactor MAL. Science 2007, 316, 1749–1752. [Google Scholar] [CrossRef] [Green Version]
- Farrants, A.K.Ö. Chromatin remodelling and actin organisation. FEBS Lett. 2008, 582, 2041–2050. [Google Scholar] [CrossRef] [Green Version]
- Rando, O.J.; Zhao, K.; Janmey, P.; Crabtree, G.R. Phosphatidylinositol-dependent actin filament binding by the SWI/SNF-like BAF chromatin remodeling complex. Proc. Nat. Acad. Sci. 2002, 99, 2824–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philimonenko, V.V.; Zhao, J.; Iben, S.; Dingová, H.; Kyselá, K.; Kahle, M.; Zentgraf, H.; Hofmann, W.A.; de Lanerolle, P.; Hozák, P.; et al. Nuclear actin and myosin I are required for RNA polymerase I transcription. Nat. Cell Biol. 2004, 6, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Fomproix, N.; Percipalle, P. An actin-myosin complex on actively transcribing genes. Exp. Cell Res. 2004, 294, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, W.A.; Stojiljkovic, L.; Fuchsova, B.; Vargas, G.M.; Mavrommatis, E.; Philimonenko, V.; Kysela, K.; Goodrich, J.A.; Lessard, J.L.; Hope, T.J.; et al. Actin is part of pre-initiation complexes and is necessary for transcription by RNA polymerase II. Nat. Cell Biol. 2004, 6, 1094–1101. [Google Scholar] [CrossRef]
- Kukalev, A.; Nord, Y.; Palmberg, C.; Bergman, T.; Percipalle, P. Actin and hnRNP U cooperate for productive transcription by RNA polymerase II. Nat. Struct. Mol. Biol. 2005, 12, 238–244. [Google Scholar] [CrossRef]
- Munsie, L.N.; Desmond, C.R.; Truant, R. Cofilin nuclear–cytoplasmic shuttling affects cofilin–actin rod formation during stress. J. Cell Sci. 2012, 125, 3977–3988. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, A.; Pope, B.; Weeds, A.; Koffer, A. Latrunculin B or ATP depletion induces cofilin-dependent translocation of actin into nuclei of mast cells. J. Biol. Chem. 2003, 278, 14394–14400. [Google Scholar] [CrossRef] [Green Version]
- Iida, K.; Matsumoto, S.; Yahara, I. The KKRKK Sequence is involved in heat shock-induced nuclear translocation of the 18-kDa actin-binding protein, cofilin. Cell Struct. Funct. 2009, 17, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Stüven, T.; Hartmann, E.; Görlich, D. Exportin 6: A novel nuclear export receptor that is specific for profilin-actin complexes. EMBO J. 2003, 22, 5928–5940. [Google Scholar] [CrossRef] [Green Version]
- Vartiainen, M.K. Nuclear actin dynamics—From form to function. FEBS Lett. 2008, 582, 2033–2040. [Google Scholar] [CrossRef] [Green Version]
- Sreedhar, A.S.; Kalmár, E.; Csermely, P.; Shen, Y.-F. Hsp90 isoforms: Functions, expression and clinical importance. FEBS Lett. 2004, 562, 11–15. [Google Scholar] [CrossRef]
- Hartl, F.U.; Hayer-Hartl, M. Molecular chaperones in the cytosol: From nascent chain to folded protein. Science 2002, 295, 1852–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyasu, S.; Nishida, E.; Kadowaki, T.; Matsuzaki, F.; Iida, K.; Harada, F.; Kasuga, M.; Sakai, H.; Yahara, I. Two mammalian heat shock proteins, HSP90 and HSP100, are actin-binding proteins. Proc. Nat. Acad. Sci. 1986, 83, 8054–8058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Suetsugu, S.; Sagara, H.; Takenawa, T. HSP90 cross-links branched actin filaments induced by N-WASP and the Arp2/3 complex. Genes Cells 2007, 12, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W. Changes in cell morphology and actin organization during heat shock in Dictyostelium discoideum: Does HSP70 play a role in acquired thermotolerance? FEMS Microbiol. Lett. 2002, 178, 95–107. [Google Scholar] [CrossRef]
- Mounier, N.; Arrigo, A.P. Actin cytoskeleton and small heat shock proteins: How do they interact? Cell Stress Chaperones 2002, 7, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, M.; Luthold, C.; Guilbert, S.M.; Varlet, A.A.; Lambert, H.; Jetté, A.; Elowe, S.; Landry, J.; Lavoie, J.N. A role for the chaperone complex BAG3-HSPB8 in actin dynamics, spindle orientation and proper chromosome segregation during mitosis. PLoS Genet. 2015, 11, 1–32. [Google Scholar] [CrossRef]
- Caplan, A.J. What is a co-chaperone? Cell Stress Chaperones 2003, 8, 105–107. [Google Scholar] [CrossRef]
- Baindur-Hudson, S.; Edkins, A.L.; Blatch, G.L. Hsp70/Hsp90 Organising Protein (Hop): Beyond Interactions with Chaperones and Prion Proteins. In The Networking of Chaperones by Co-chaperones. Subcellular Biochemistry; Blatch, G.L., Edkins, A.L., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 78, pp. 69–90. [Google Scholar]
- Beraldo, F.H.; Thomas, A.; Kolisnyk, B.; Hirata, P.H.; De Jaeger, X.; Martyn, A.C.; Fan, J.; Goncalves, D.F.; Cowan, M.F.; Masood, T.; et al. Hyperactivity and attention deficits in mice with decreased levels of stress-inducible phosphoprotein 1 (STIP1). Dis. Model. Mech. 2015, 8, 1457–1466. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Shao, J.; Su, F. Prognostic significance of STIP1 expression in human cancer: A meta-analysis. Clin. Chim. Acta 2018, 486, 168–176. [Google Scholar] [CrossRef]
- Luo, X.; Liu, Y.; Ma, S.; Liu, L.; Xie, R.; Li, M.; Shen, P.; Wang, S. STIP1 is over-expressed in hepatocellular carcinoma and promotes the growth and migration of cancer cells. Gene 2018, 662, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; You, H.; Qi, J.; Yang, C.; Ren, Y.; Cheng, H. Autocrine and paracrine STIP1 signaling promote osteolytic bone metastasis in renal cell carcinoma. Oncotarget 2017, 8, 17012–17026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Kim, S.; Shin, H.-Y.; Chung, E.J.; Kitano, H.; Hyon Park, J.; Park, L.; Chung, J.-Y.; Hewitt, S.M.; Kim, J.-H. Expression of stress-induced phosphoprotein1 (STIP1) is associated with tumor progression and poor prognosis in epithelial ovarian cancer. Genes Chromosom. Cancer 2014, 53, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.-H.; Zhou, R.-S.; She, B.; Xu, H.-F.; Wang, J.-Y.; Wei, L.-X. Expression and clinical significance of STIP1 in papillary thyroid carcinoma. Tumor Biol. 2014, 35, 2391–2395. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Lai, C.H.; Tsai, C.L.; Hsueh, S.; Hsueh, C.; Lin, C.Y.; Chou, H.H.; Lin, Y.J.; Chen, H.W.; Chang, T.C.; et al. Tumor stress-induced phosphoprotein1 (STIP1) as a prognostic biomarker in ovarian cancer. PLoS ONE 2013, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Fonseca, A.C.C.; Romão, L.; Amaral, R.F.; Assad Kahn, S.; Lobo, D.; Martins, S.; Marcondes de Souza, J.; Moura-Neto, V.; Lima, F.R.S. Microglial stress inducible protein 1 promotes proliferation and migration in human glioblastoma cells. Neuroscience 2012, 200, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix metalloproteinase-9: Many shades of function in cardiovascular disease. Physiology (Bethesda) 2013, 28, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Willmer, T.; Contu, L.; Blatch, G.L.; Edkins, A.L. Knockdown of Hop downregulates RhoC expression, and decreases pseudopodia formation and migration in cancer cell lines. Cancer Lett. 2013, 328, 252–260. [Google Scholar] [CrossRef]
- De Souza, L.E.R.; Moura Costa, M.D.; Bilek, E.S.; Lopes, M.H.; Martins, V.R.; Püschel, A.W.; Mercadante, A.F.; Nakao, L.S.; Zanata, S.M. STI1 antagonizes cytoskeleton collapse mediated by small GTPase Rnd1 and regulates neurite growth. Exp. Cell Res. 2014, 324, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, J.; Barber, D. Actin Co-Sedimentation Assay; for the Analysis of Protein Binding to F-Actin. J. Vis. Exp. 2008, 13, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Kajander, T.; Sachs, J.N.; Goldman, A.; Regan, L. Electrostatic interactions of Hsp-organizing protein tetratricopeptide domains with Hsp70 and Hsp90: Computational analysis and protein engineering. J. Biol. Chem. 2009, 284, 25364–25374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, A.B.; Lagleder, S.; Gräwert, M.A.; Röhl, A.; Hagn, F.; Wandinger, S.K.; Cox, M.B.; Demmer, O.; Richter, K.; Groll, M.; et al. The architecture of functional modules in the Hsp90 co-chaperone Sti1/Hop. EMBO J. 2012, 31, 1506–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flom, G.; Behal, R.H.; Rosen, L.; Cole, D.G.; Johnson, J.L. Definition of the minimal fragments of Sti1 required for dimerization, interaction with Hsp70 and Hsp90 and in vivo functions. Biochem. J. 2007, 404, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheufler, C.; Brinker, A.; Bourenkov, G.; Pegoraro, S.; Moroder, L.; Bartunik, H.; Hartl, F.U.; Moarefi, I. Structure of TPR domain-peptide complexes: Critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 2000, 101, 199–210. [Google Scholar] [CrossRef]
- Brinker, A.; Scheufler, C.; Von Der Mülbe, F.; Fleckenstein, B.; Herrmann, C.; Jung, G.; Moarefi, I.; Ulrich Hartl, F. Ligand discrimination by TPR domains. Relevance and selectivity of EEVD-recognition in Hsp70·Hop·Hsp90 complexes. J. Biol. Chem. 2002, 277, 19265–19275. [Google Scholar] [CrossRef]
- Li, J.; Sun, X.; Wang, Z.; Chen, L.; Li, D.; Zhou, J.; Liu, M. Regulation of vascular endothelial cell polarization and migration by HsP70/HsP90-organizing protein. PLoS ONE 2012, 7, e36389. [Google Scholar] [CrossRef]
- Coitinho, A.S.; Lopes, M.H.; Hajj, G.N.M.; Rossato, J.I.; Freitas, A.R.; Castro, C.C.; Cammarota, M.; Brentani, R.R.; Izquierdo, I.; Martins, V.R. Short-term memory formation and long-term memory consolidation are enhanced by cellular prion association to stress-inducible protein 1. Neurobiol. Dis. 2007, 26, 282–290. [Google Scholar] [CrossRef]
- Kituyi, S.N.; Edkins, A.L. Hop/STIP1 depletion alters nuclear structure via depletion of nuclear structural protein emerin. Biochem. Biophys. Res. Commun. 2018, 507, 503–509. [Google Scholar] [CrossRef]
- Lässle, M.; Blatch, G.L.; Kundra, V.; Takatori, T.; Zetter, B.R. Stress-inducible, murine protein mSTI1. Characterization of binding domains for heat shock proteins and in vitro phosphorylation by different kinases. J. Biol. Chem. 1997, 272, 1876–1884. [Google Scholar] [CrossRef] [Green Version]
- Odunuga, O.O.; Hornby, J.A.; Bies, C.; Zimmermann, R.; Pugh, D.J.; Blatch, G.L. Tetratricopeptide repeat motif-mediated Hsc70-mSTI1 interaction. Molecular characterization of the critical contacts for successful binding and specificity. J. Biol. Chem. 2003, 278, 6896–6904. [Google Scholar] [CrossRef] [Green Version]
- Obenauer, J.C.; Cantley, L.C.; Yaffe, M.B. Scansite 2.0: Proteome-wide prediction of cell signaling interactions using short sequence motifs. Nucleic Acids Res. 2003, 31, 3635–3641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagni, M.; Ioannidis, V.; Cerutti, L.; Zahn-Zabal, M.; Jongeneel, C.V.; Hau, J.; Martin, O.; Kuznetsov, D.; Falquet, L. MyHits: Improvements to an interactive resource for analyzing protein sequences. Nucleic Acids Res. 2007, 35, W433–W437. [Google Scholar] [CrossRef] [PubMed]
- Alexandropoulos, K.; Cheng, G.; Baltimore, D. Proline-rich sequences that bind to Src homology 3 domains with individual specificities. Proc. Nat. Acad. Sci. 1995, 92, 3110–3114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saksela, K.; Permi, P. SH3 domain ligand binding: What’s the consensus and where’s the specificity? FEBS Lett. 2012, 586, 2609–2614. [Google Scholar] [CrossRef] [Green Version]
- He, H.J.; Wang, X.S.; Pan, R.; Wang, D.L.; Liu, M.N.; He, R.Q. The proline-rich domain of tau plays a role in interactions with actin. BMC Cell Biol. 2009, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Holt, M.R.; Koffer, A. Cell motility: Proline-rich proteins promote protrusions. Trends Cell Biol. 2001, 11, 38–46. [Google Scholar] [CrossRef]
- Kay, B.K.; Williamson, M.P.; Sudol, M. The importance of being proline: The interaction of proline-rich motifs in signaling proteins with their cognate domains. FASEB J. 2000, 14, 231–241. [Google Scholar] [CrossRef]
- Urbanek, A.N.; Smith, A.P.; Allwood, E.G.; Booth, W.I.; Ayscough, K.R. A novel actin-binding motif in Las17/WASP nucleates actin filaments independently of Arp2/3. Curr. Biol. 2013, 23, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, N.; Nishida, E.; Ohba, M.; Seki, M.; Kumagai, H.; Sakai, H. An actin-interacting heptapeptide in the cofilin sequence. Eur. J. Biochem. 1989, 183, 235–238. [Google Scholar] [CrossRef]
- Pantaloni, D.; Hill, T.L.; Carlier, M.F.; Korn, E.D. A model for actin polymerization and the kinetic effects of ATP hydrolysis. Proc. Nat. Acad. Sci. 1985, 82, 7207–7211. [Google Scholar] [CrossRef] [Green Version]
- Wegele, H.; Haslbeck, M.; Reinstein, J.; Buchner, J. Sti1 is a novel activator of the Ssa proteins. J. Biol. Chem. 2003, 278, 25970–25976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bork, P.; Sander, C.; Valencia, A. An ATPase domain common to prokaryotic cell cycle proteins, sugar kinases, actin, and hsp70 heat shock proteins. Proc. Nat. Acad. Sci. 1992, 89, 7290–7294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinchuk, V.; Zinchuk, O.; Okada, T. Quantitative colocalization analysis of multicolor confocal immunofluorescence microscopy images: Pushing pixels to explore biological phenomena. Acta Histochem. Cytochem. 2007, 40, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Lau, A.; Morris, T.J.; Guo, L.; Fordyce, C.B.; Stanley, E.F. A Syntaxin 1, G(alpha)o, and N-Type Calcium Channel Complex at a Presynaptic Nerve Terminal: Analysis by Quantitative Immunocolocalization. J. Neurosci. 2004, 24, 4070–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukui, Y. Dynamics of nuclear actin bundle induction by dimethyl sulfoxide and factors affecting its development. J. Cell Biol. 2004, 84, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Cordero, J.J.; Oser, M.; Chen, X.; Eddy, R.; Hodgson, L.; Condeelis, J. A novel spatiotemporal RhoC activation pathway locally regulates cofilin activity at invadopodia. Curr. Biol. 2011, 21, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Spuy, J.; Cheetham, M.E.; Dirr, H.W.; Blatch, G.L. The cochaperone murine stress-inducible protein 1: Overexpression, purification, and characterization. Protein Expr. Purif. 2001, 21, 462–469. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Nat. Acad. Sci. 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Posern, G.; Sotiropoulos, A.; Treisman, R. Mutant actins demonstrate a role for unpolymerized actin in control of transcription by serum response factor. Mol. Biol. Cell 2002, 13, 4167–4178. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beckley, S.J.; Hunter, M.C.; Kituyi, S.N.; Wingate, I.; Chakraborty, A.; Schwarz, K.; Makhubu, M.P.; Rousseau, R.P.; Ruck, D.K.; de la Mare, J.-A.; et al. STIP1/HOP Regulates the Actin Cytoskeleton through Interactions with Actin and Changes in Actin-Binding Proteins Cofilin and Profilin. Int. J. Mol. Sci. 2020, 21, 3152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093152
Beckley SJ, Hunter MC, Kituyi SN, Wingate I, Chakraborty A, Schwarz K, Makhubu MP, Rousseau RP, Ruck DK, de la Mare J-A, et al. STIP1/HOP Regulates the Actin Cytoskeleton through Interactions with Actin and Changes in Actin-Binding Proteins Cofilin and Profilin. International Journal of Molecular Sciences. 2020; 21(9):3152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093152
Chicago/Turabian StyleBeckley, Samantha Joy, Morgan Campbell Hunter, Sarah Naulikha Kituyi, Ianthe Wingate, Abantika Chakraborty, Kelly Schwarz, Matodzi Portia Makhubu, Robert Pierre Rousseau, Duncan Kyle Ruck, Jo-Anne de la Mare, and et al. 2020. "STIP1/HOP Regulates the Actin Cytoskeleton through Interactions with Actin and Changes in Actin-Binding Proteins Cofilin and Profilin" International Journal of Molecular Sciences 21, no. 9: 3152. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093152