TATA-Like Boxes in RNA Polymerase III Promoters: Requirements for Nucleotide Sequences


Abstract
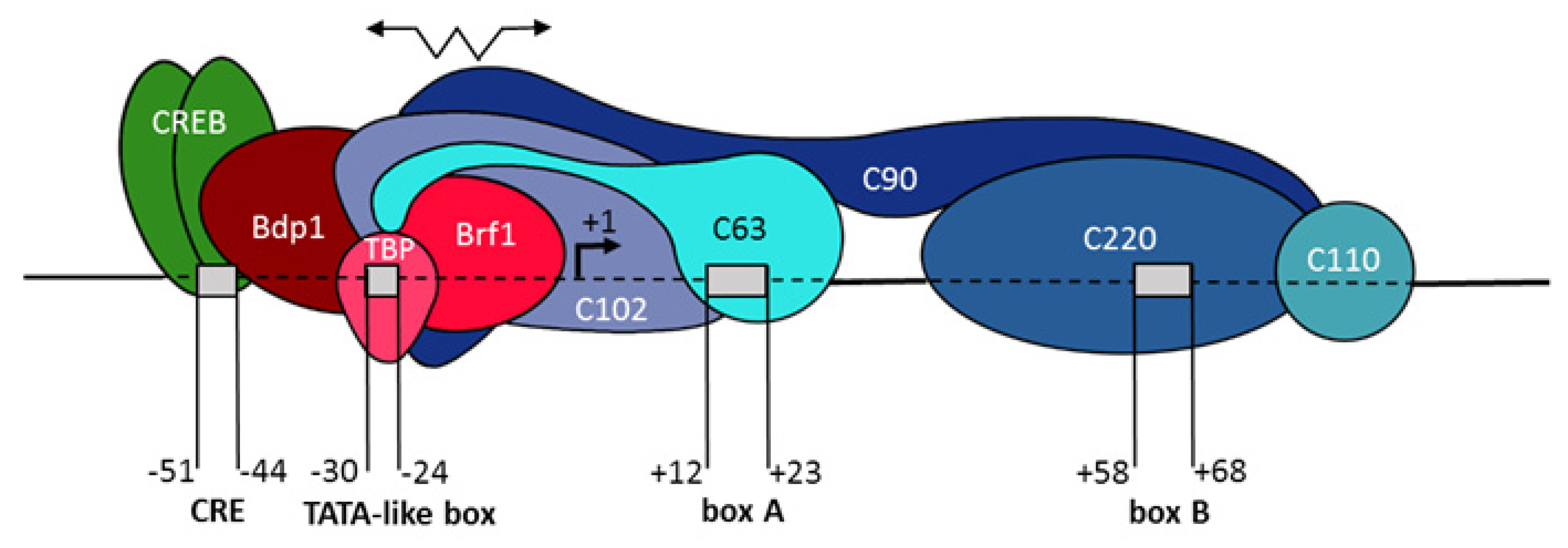
:1. Introduction
2. Results
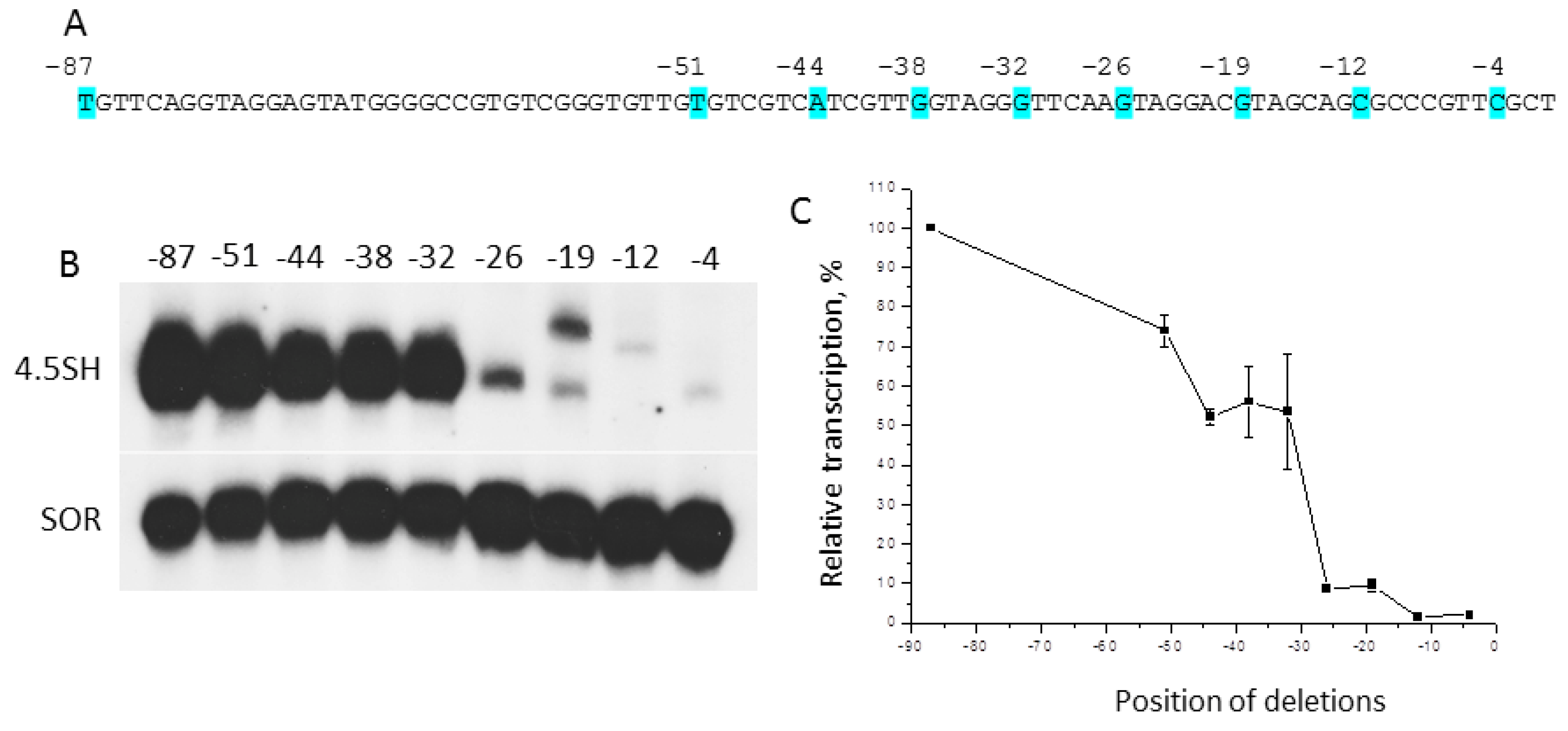
2.1. The Deletion Analysis of 5′-Flanking Sequence of Murine 4.5SH RNA Gene
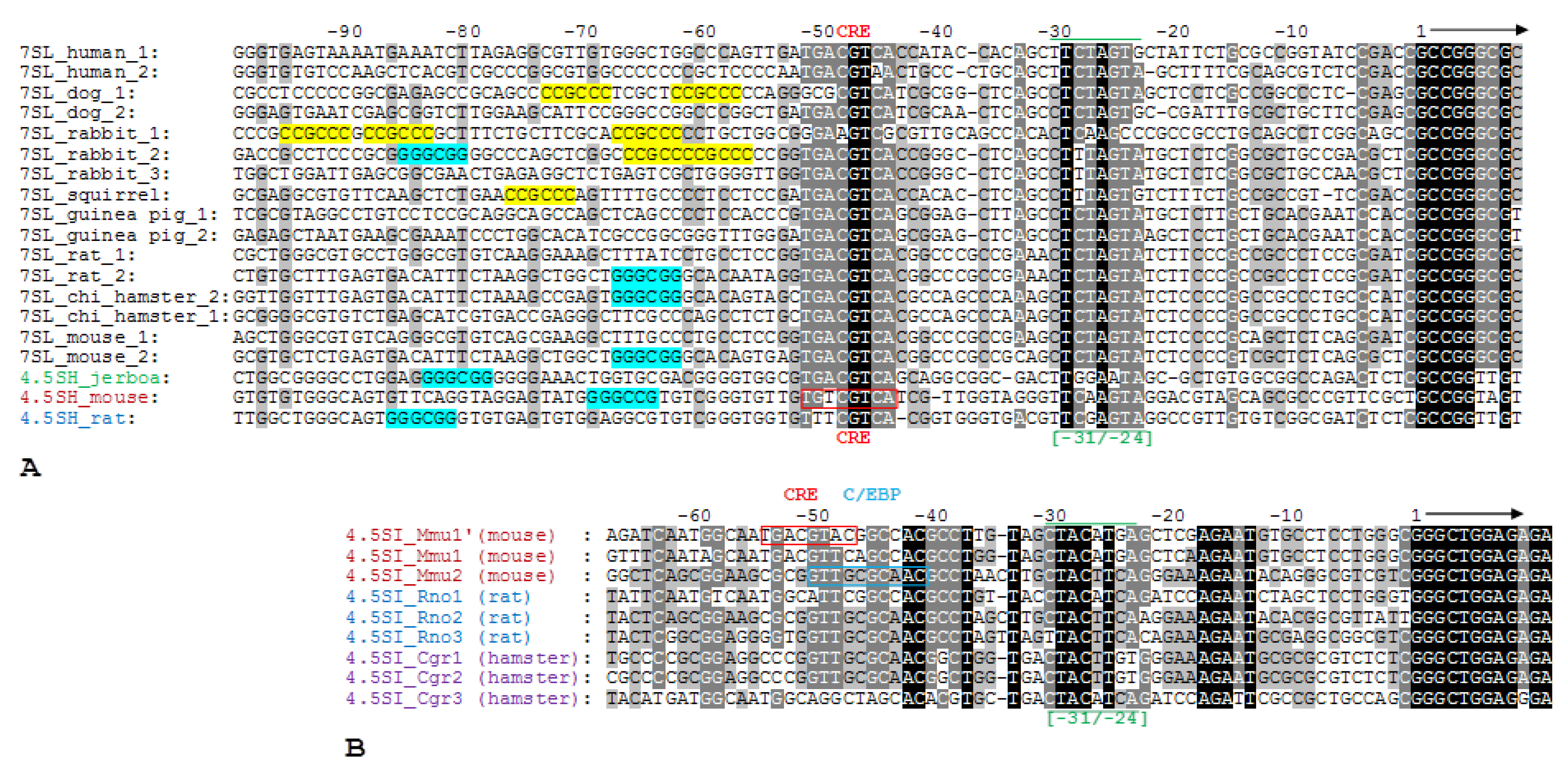
2.2. The Comparison Analysis of 5′-FSs of 4.5SH and 7SL RNA Genes
2.3. Influence of Nucleotide Substitutions in TATA-Like Box of 4.5SH RNA Gene on Its Transcription
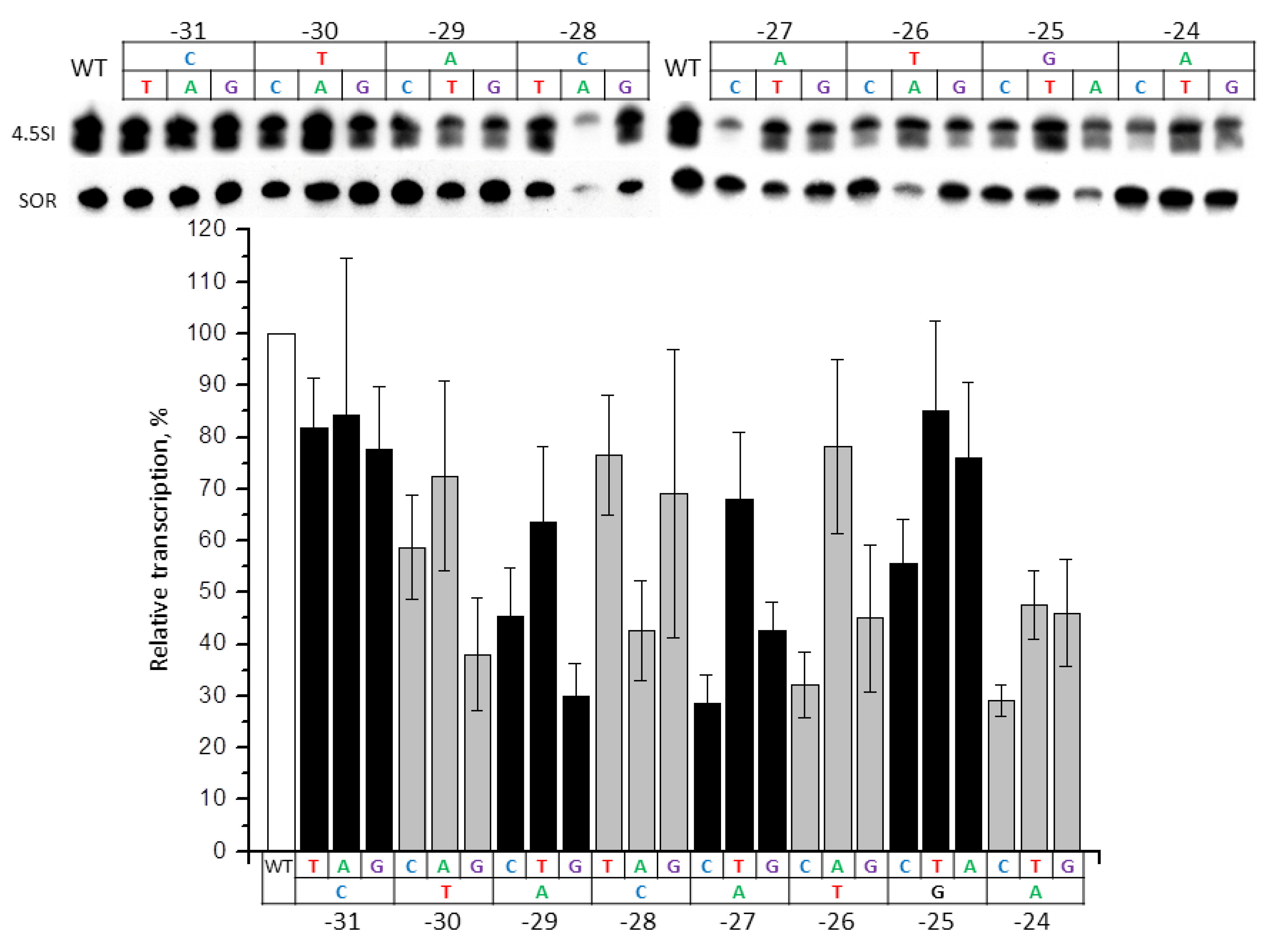
2.4. Influence of Nucleotide Substitutions in TATA-Like Box of 4.5SI RNA Gene on its Transcription
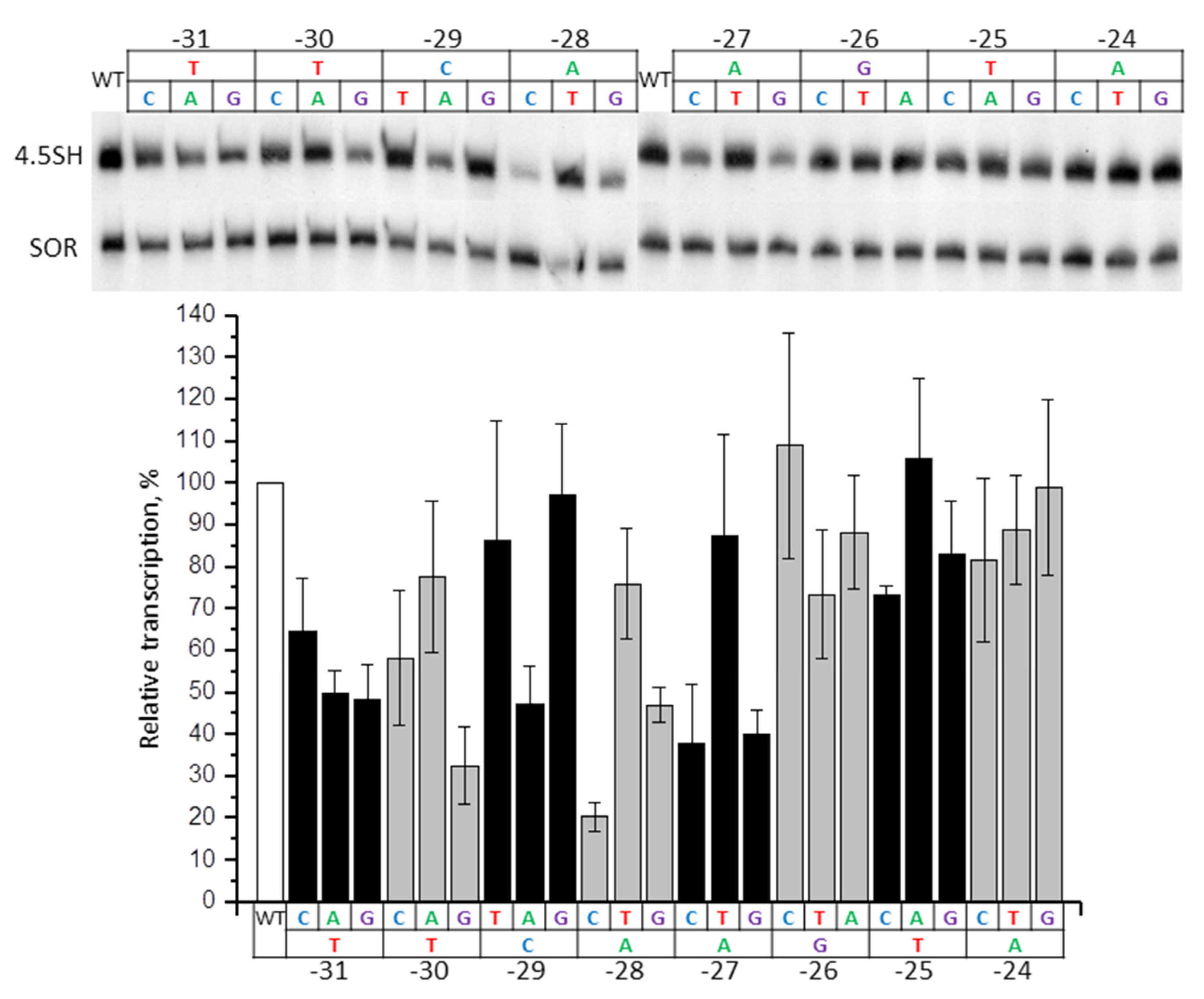
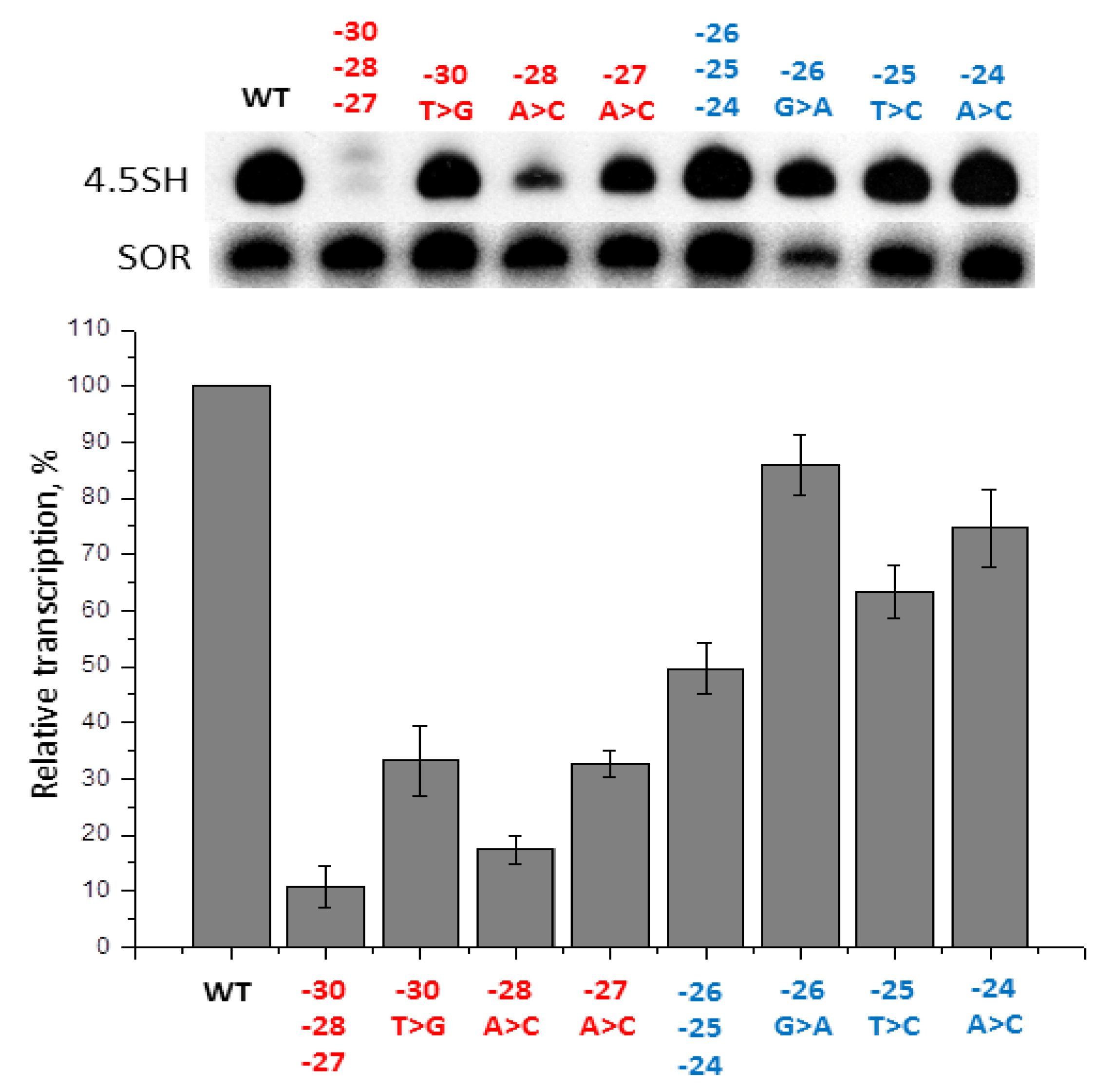
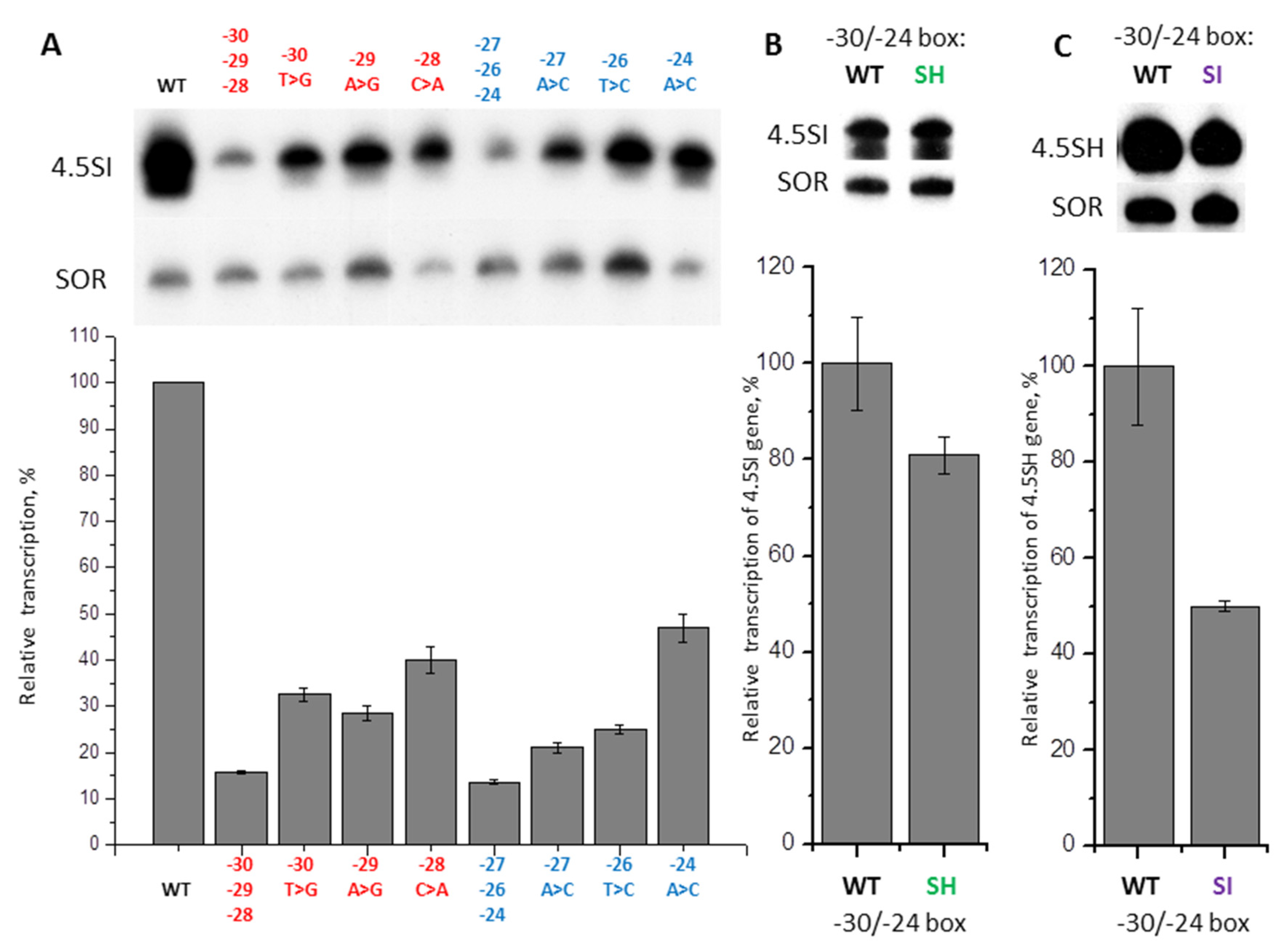
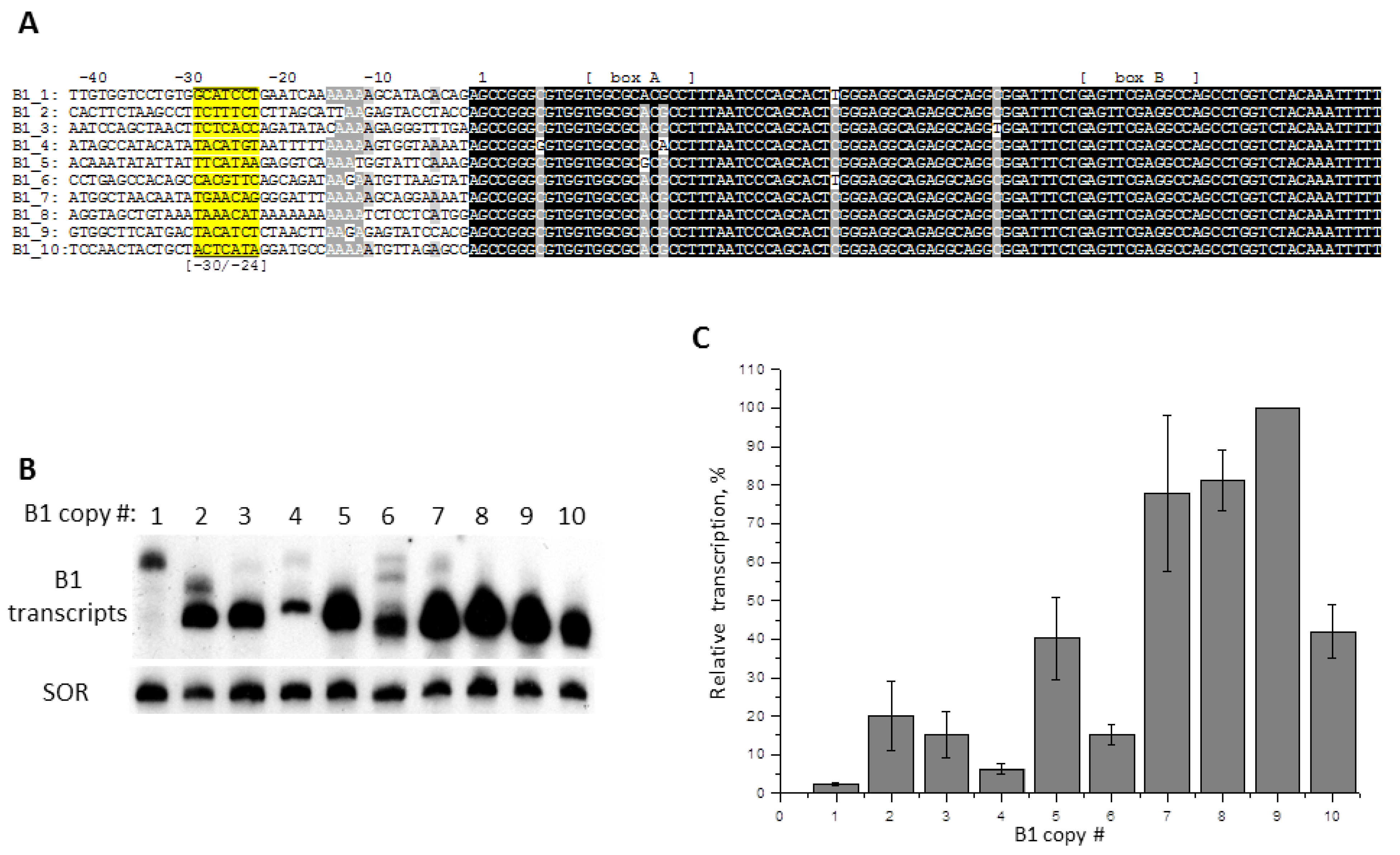
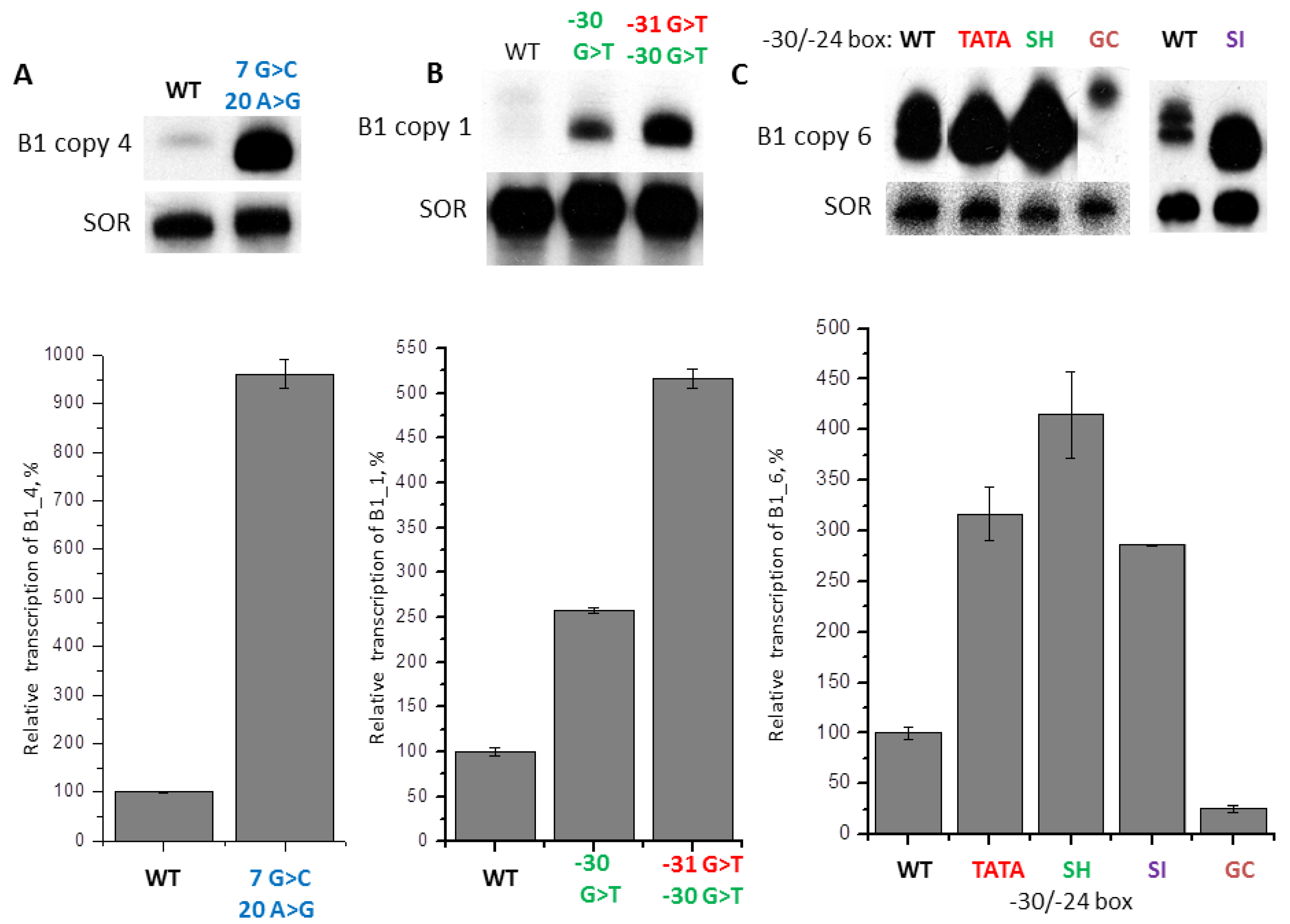
2.5. Contribution of −31/−24 Region to the Efficiency of SINE B1 Transcription
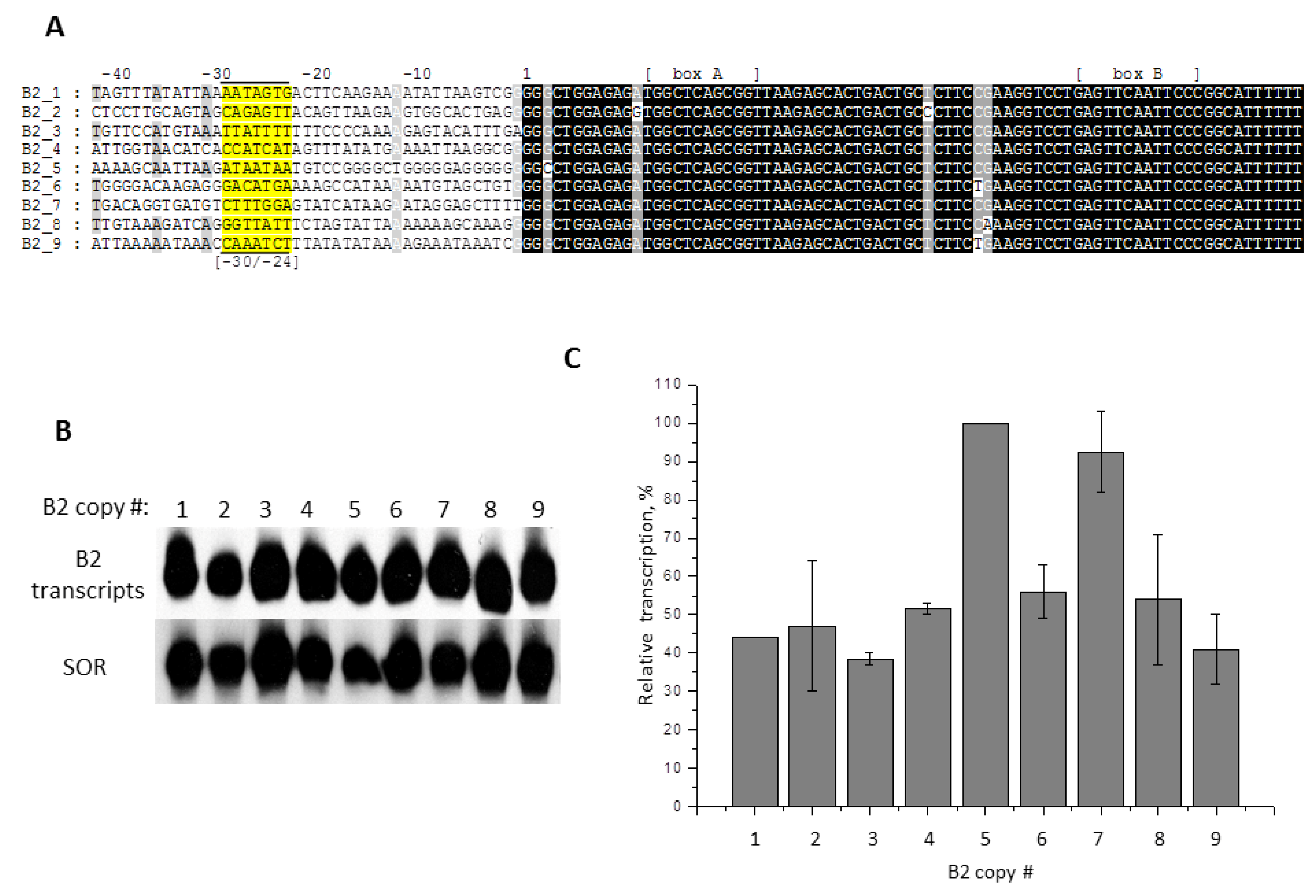
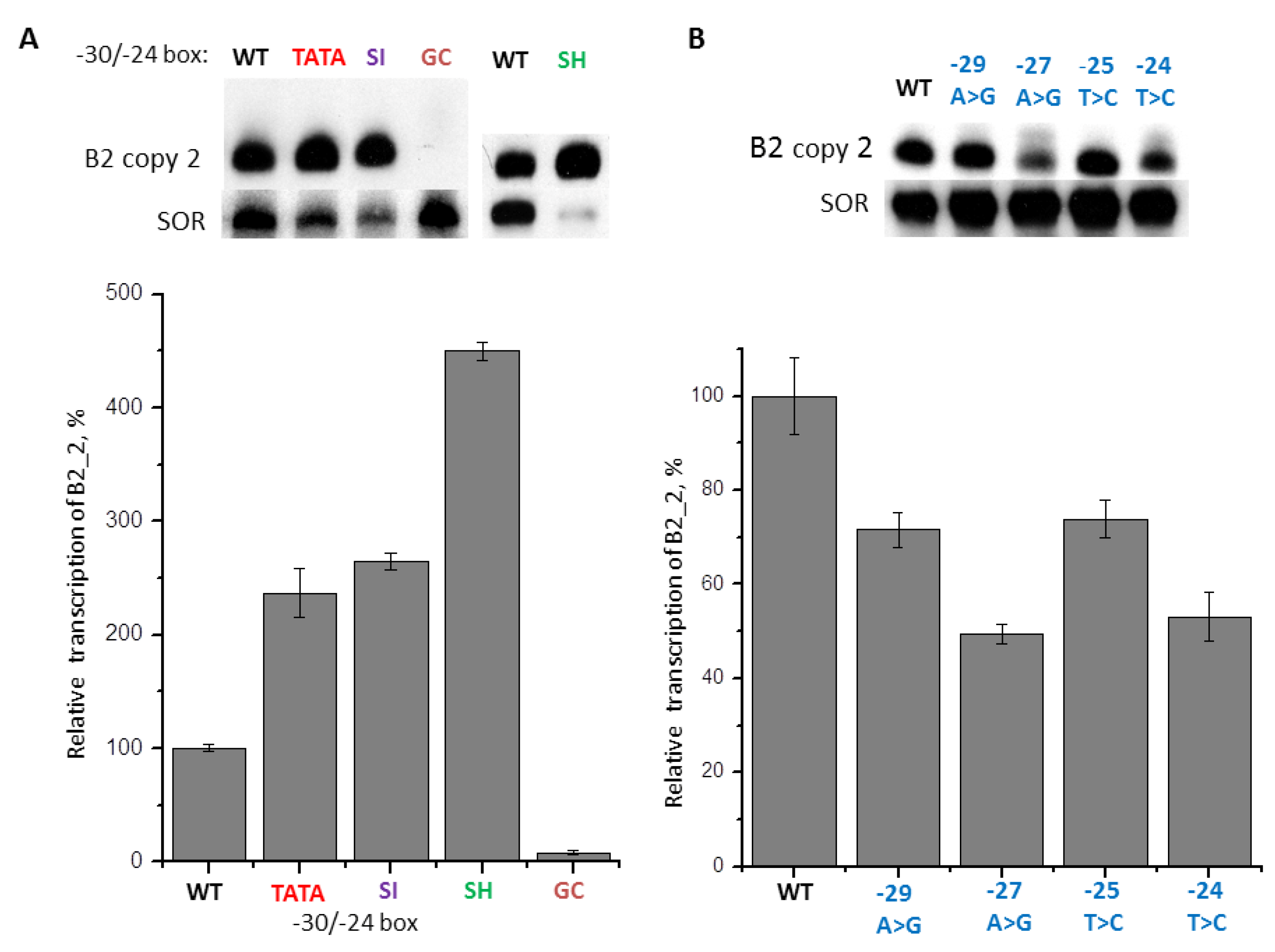
2.6. Contribution of −31/−24 Region to the Efficiency of SINE B2 Transcription
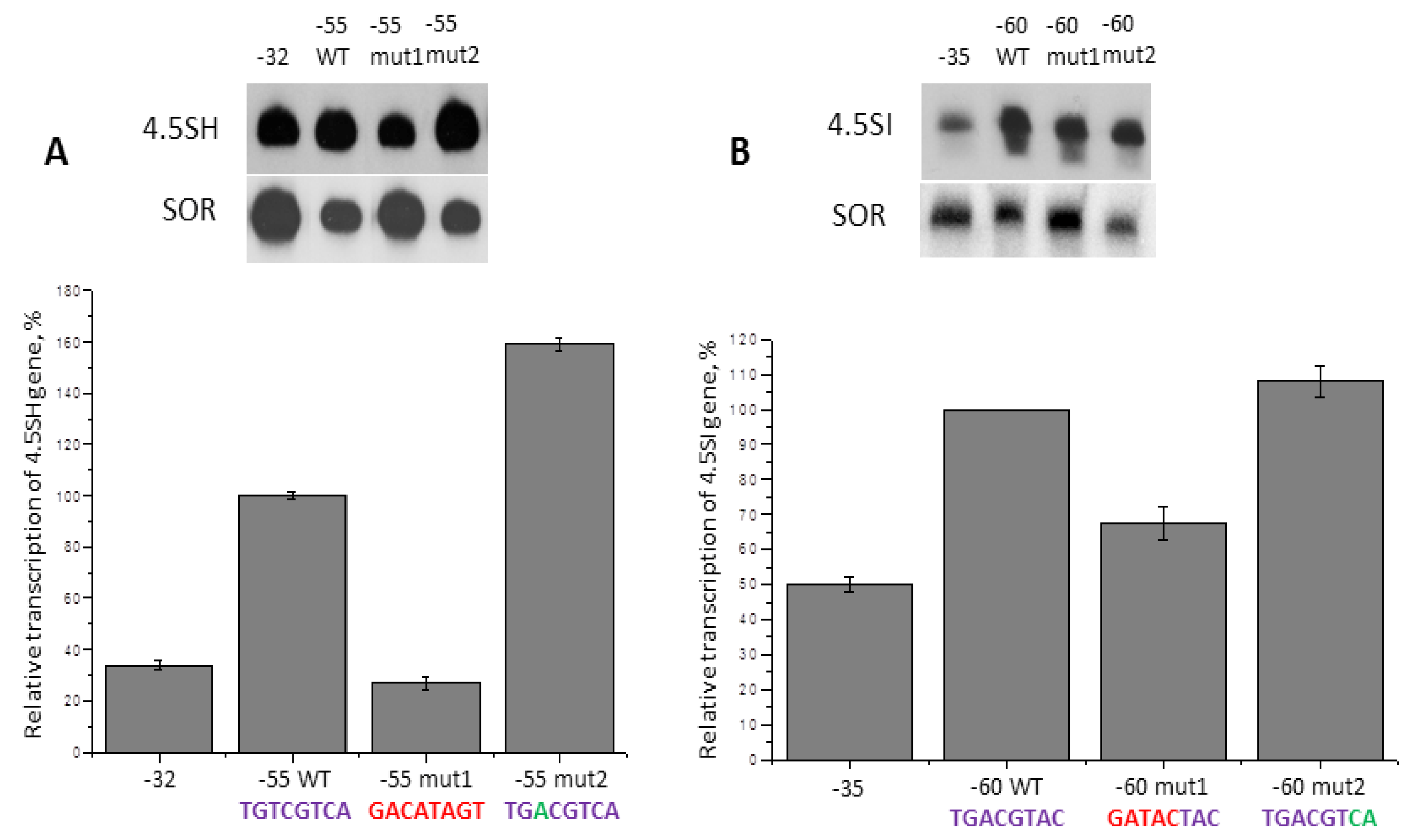
2.7. Additional Regulatory Elements in 5′- Flanking Sequences of 4.5SH Genes
2.8. Additional Regulatory Elements in 5′-Flanking Sequences of 4.5SI RNA Genes
3. Discussion
3.1. TATA-Like Boxes in 4.5SH RNA and 4.5SI RNA Genes
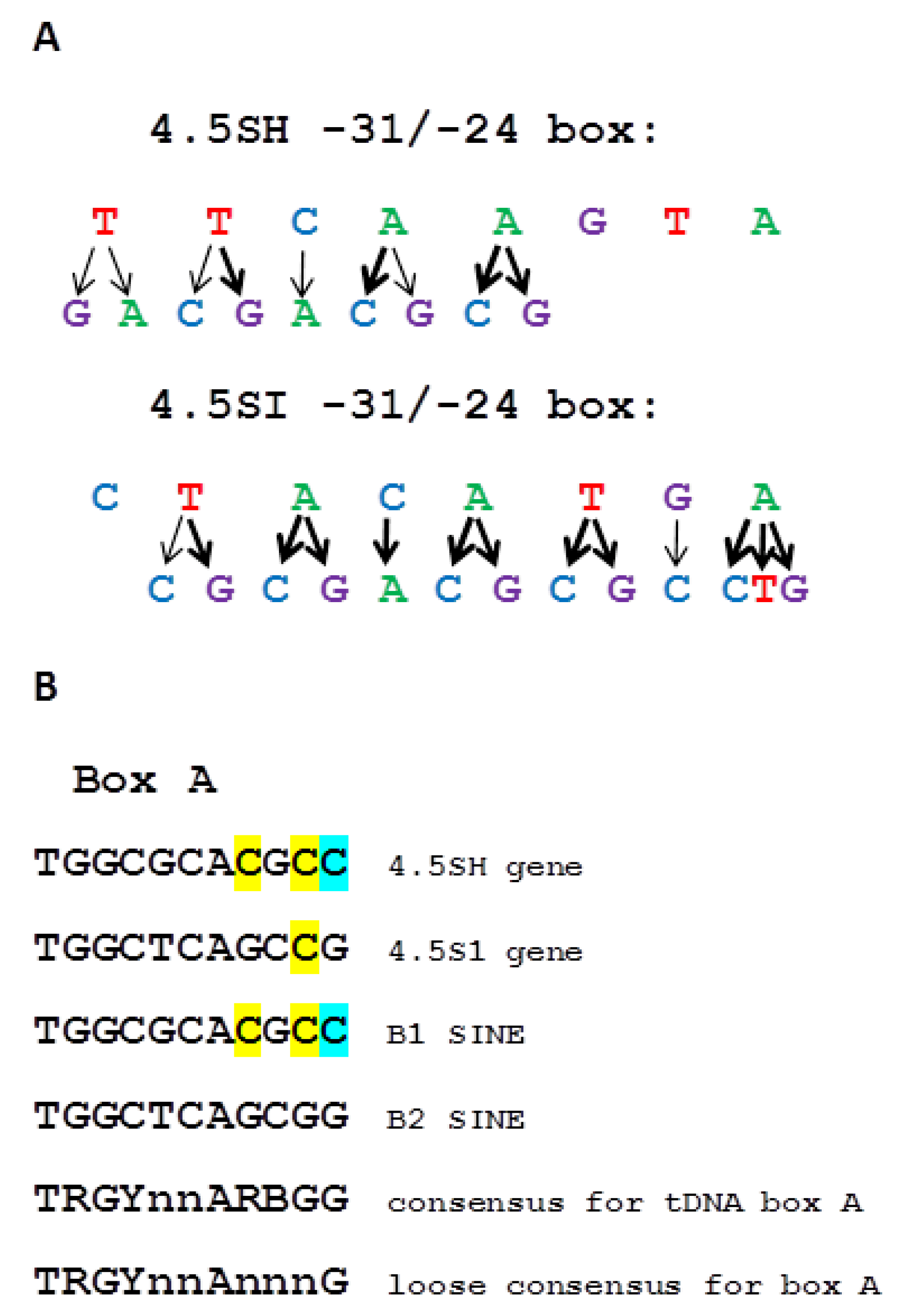
3.2. The Role of −31/−24 Sequence in B1 and B2 SINE Transcription
3.3. Changed −31/−24 Box may Shift the TSS
3.4. Potential Regulatory Elements in 5′-Flanking Sequences of Genes with Type 2 pol III Promoters
4. Materials and Methods
4.1. Plasmid Constructs
4.2. Cell Transfection and Northern-Blot Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 5′-FS | 5′-flanking sequence |
| ATF | activating transcription factor |
| Bdp1 | B double prime 1 factor |
| Brf | TFIIIB-related factor |
| C/EBP | CCAAT-enhancer binding protein |
| CRE | cAMP-response element |
| CREB | cAMP-response element binding protein |
| DSE | distal sequence element |
| EBER | Epstein-Barr virus-encoded RNA |
| pol III | RNA polymerase III |
| PSE | proximal sequence element |
| SINE | short interspersed element |
| TBP | TATA box binding protein |
| TFIII | transcription factor for pol III |
| TSS | transcription start site |
| vtRNA | vault RNA |
References
- Dieci, G.; Fiorino, G.; Castelnuovo, M.; Teichmann, M.; Pagano, A. The expanding RNA polymerase III transcriptome. Trends Genet. 2007, 23, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution-trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [Green Version]
- Kheir, E.; Krude, T. Non-coding Y RNAs associate with early replicating euchromatin in concordance with the origin recognition complex. J. Cell Sci. 2017, 130, 1239–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horos, R.; Buscher, M.; Kleinendorst, R.; Alleaume, A.M.; Tarafder, A.K.; Schwarzl, T.; Dziuba, D.; Tischer, C.; Zielonka, E.M.; Adak, A.; et al. The Small Non-coding Vault RNA1-1 Acts as a Riboregulator of Autophagy. Cell 2019, 176, 1054–1067 e1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Pestova, T.V.; Hellen, C.U.; Tiedge, H. Translational control by a small RNA: Dendritic BC1 RNA targets the eukaryotic initiation factor 4A helicase mechanism. Mol. Cell. Biol. 2008, 28, 3008–3019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA polymerase III transcribes human microRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Pagano, A.; Castelnuovo, M.; Tortelli, F.; Ferrari, R.; Dieci, G.; Cancedda, R. New small nuclear RNA gene-like transcriptional units as sources of regulatory transcripts. PLoS Genet. 2007, 3, e1. [Google Scholar] [CrossRef] [Green Version]
- Gigoni, A.; Costa, D.; Gaetani, M.; Tasso, R.; Villa, F.; Florio, T.; Pagano, A. Down-regulation of 21A Alu RNA as a tool to boost proliferation maintaining the tissue regeneration potential of progenitor cells. Cell Cycle 2016, 15, 2420–2430. [Google Scholar] [CrossRef] [Green Version]
- Schramm, L.; Hernandez, N. Recruitment of RNA polymerase III to its target promoters. Genes Dev. 2002, 16, 2593–2620. [Google Scholar] [CrossRef] [Green Version]
- Orioli, A.; Pascali, C.; Pagano, A.; Teichmann, M.; Dieci, G. RNA polymerase III transcription control elements: Themes and variations. Gene 2012, 493, 185–194. [Google Scholar] [CrossRef]
- Oler, A.J.; Alla, R.K.; Roberts, D.N.; Wong, A.; Hollenhorst, P.C.; Chandler, K.J.; Cassiday, P.A.; Nelson, C.A.; Hagedorn, C.H.; Graves, B.J.; et al. Human RNA polymerase III transcriptomes and relationships to Pol II promoter chromatin and enhancer-binding factors. Nat. Struct. Mol. Biol. 2010, 17, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Vassetzky, N.S.; Kramerov, D.A. SINEBase: A database and tool for SINE analysis. Nucleic Acids Res. 2013, 41, D83–D89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiduschek, E.P.; Kassavetis, G.A. The RNA polymerase III transcription apparatus. J. Mol. Biol. 2001, 310, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Huang, Y.; Lowe, T.M.; Maraia, R.J. Widespread use of TATA elements in the core promoters for RNA polymerases III, II, and I in fission yeast. Mol. Cell. Biol. 2001, 21, 6870–6881. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, C.; Martinez, M.J.; Young, L.S.; Sprague, K.U. TATA-Binding protein-TATA interaction is a key determinant of differential transcription of silkworm constitutive and silk gland-specific tRNA(Ala) genes. Mol. Cell. Biol. 2000, 20, 1329–1343. [Google Scholar] [CrossRef] [Green Version]
- Giuliodori, S.; Percudani, R.; Braglia, P.; Ferrari, R.; Guffanti, E.; Ottonello, S.; Dieci, G. A composite upstream sequence motif potentiates tRNA gene transcription in yeast. J. Mol. Biol. 2003, 333, 1–20. [Google Scholar] [CrossRef]
- Zhang, G.; Lukoszek, R.; Mueller-Roeber, B.; Ignatova, Z. Different sequence signatures in the upstream regions of plant and animal tRNA genes shape distinct modes of regulation. Nucleic Acids Res. 2011, 39, 3331–3339. [Google Scholar] [CrossRef] [Green Version]
- White, R.J.; Jackson, S.P. Mechanism of TATA-binding protein recruitment to a TATA-less class III promoter. Cell 1992, 71, 1041–1053. [Google Scholar] [CrossRef]
- Joazeiro, C.A.; Kassavetis, G.A.; Geiduschek, E.P. Identical components of yeast transcription factor IIIB are required and sufficient for transcription of TATA box-containing and TATA-less genes. Mol. Cell. Biol. 1994, 14, 2798–2808. [Google Scholar] [CrossRef] [Green Version]
- Howe, J.G.; Shu, M.D. Upstream basal promoter element important for exclusive RNA polymerase III transcription of the EBER 2 gene. Mol. Cell. Biol. 1993, 13, 2655–2665. [Google Scholar] [CrossRef] [Green Version]
- Englert, M.; Felis, M.; Junker, V.; Beier, H. Novel upstream and intragenic control elements for the RNA polymerase III-dependent transcription of human 7SL RNA genes. Biochimie 2004, 86, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Kickhoefer, V.A.; Emre, N.; Stephen, A.G.; Poderycki, M.J.; Rome, L.H. Identification of conserved vault RNA expression elements and a non-expressed mouse vault RNA gene. Gene 2003, 309, 65–70. [Google Scholar] [CrossRef]
- Krayev, A.S.; Markusheva, T.V.; Kramerov, D.A.; Ryskov, A.P.; Skryabin, K.G.; Bayev, A.A.; Georgiev, G.P. Ubiquitous transposon-like repeats B1 and B2 of the mouse genome: B2 sequencing. Nucleic Acids Res. 1982, 10, 7461–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, F.; Takeuchi, Y.; Kato, N. Molecular cloning and in vitro transcription of rat 4.5S RNAH genes. Nucleic Acids Res. 1986, 14, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Haynes, S.R.; Jelinek, W.R. Low molecular weight RNAs transcribed in vitro by RNA polymerase III from Alu-type dispersed repeats in Chinese hamster DNA are also found in vivo. Proc. Natl. Acad. Sci. USA 1981, 78, 6130–6134. [Google Scholar] [CrossRef] [Green Version]
- Harada, F.; Kato, N. Nucleotide sequences of 4.5S RNAs associated with poly(A)-containing RNAs of mouse and hamster cells. Nucleic Acids Res. 1980, 8, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Ro-Choi, T.S.; Redy, R.; Henning, D.; Takano, T.; Taylor, C.W.; Busch, H. Nucleotide sequence of 4.5 S ribonucleic acid of Novikoff hepatoma cell nuclei. J. Biol. Chem. 1972, 247, 3205–3222. [Google Scholar]
- Gogolevskaya, I.K.; Koval, A.P.; Kramerov, D.A. Evolutionary history of 4.5SH RNA. Mol. Biol. Evol. 2005, 22, 1546–1554. [Google Scholar] [CrossRef] [Green Version]
- Gogolevskaya, I.K.; Kramerov, D.A. Evolutionary history of 4.5SI RNA and indication that it is functional. J. Mol. Evol. 2002, 54, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Gogolevskaya, I.K.; Kramerov, D.A. 4.5SI RNA genes and the role of their 5′-flanking sequences in the gene transcription. Gene 2010, 451, 32–37. [Google Scholar] [CrossRef]
- Schoeniger, L.O.; Jelinek, W.R. 4.5S RNA is encoded by hundreds of tandemly linked genes, has a short half-life, and is hydrogen bonded in vivo to poly(A)-terminated RNAs in the cytoplasm of cultured mouse cells. Mol. Cell. Biol. 1986, 6, 1508–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatosyan, K.A.; Koval, A.P.; Gogolevskaya, I.K.; Kramerov, D.A. 4.5SI and 4.5SH RNAs: Expression in various rodent organs and abundance and distribution in the cell. Mol. Biol. (Mosk) 2017, 51, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Koval, A.P.; Gogolevskaya, I.K.; Tatosyan, K.A.; Kramerov, D.A. Complementarity of end regions increases the lifetime of small RNAs in mammalian cells. PLoS ONE 2012, 7, e44157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatosyan, K.A.; Kramerov, D.A. Heat shock increases lifetime of a small RNA and induces its accumulation in cells. Gene 2016, 587, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Miyauchi, K.; Kimura, Y.; Mito, M.; Okada, S.; Suzuki, T.; Nakagawa, S. Regulation of gene expression via retrotransposon insertions and the noncoding RNA 4.5S RNAH. Genes Cells 2015, 20, 887–901. [Google Scholar] [CrossRef]
- Kramerov, D.A.; Vassetzky, N.S. SINEs. Wiley Interdiscip. Rev. RNA 2011, 2, 772–786. [Google Scholar] [CrossRef]
- Kajikawa, M.; Okada, N. LINEs mobilize SINEs in the eel through a shared 3′ sequence. Cell 2002, 111, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, K.; Okada, N. SINEs and LINEs: Symbionts of eukaryotic genomes with a common tail. Cytogenet. Genome Res. 2005, 110, 475–490. [Google Scholar] [CrossRef]
- Kramerov, D.A.; Grigoryan, A.A.; Ryskov, A.P.; Georgiev, G.P. Long double-stranded sequences (dsRNA-B) of nuclear pre-mRNA consist of a few highly abundant classes of sequences: Evidence from DNA cloning experiments. Nucleic Acids Res. 1979, 6, 697–713. [Google Scholar] [CrossRef] [Green Version]
- Deininger, P.L.; Jolly, D.J.; Rubin, C.M.; Friedmann, T.; Schmid, C.W. Base sequence studies of 300 nucleotide renatured repeated human DNA clones. J. Mol. Biol. 1981, 151, 17–33. [Google Scholar] [CrossRef]
- Veniaminova, N.A.; Vassetzky, N.S.; Kramerov, D.A. B1 SINEs in different rodent families. Genomics 2007, 89, 678–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serdobova, I.M.; Kramerov, D.A. Short retroposons of the B2 superfamily: Evolution and application for the study of rodent phylogeny. J. Mol. Evol. 1998, 46, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Quentin, Y. Emergence of master sequences in families of retroposons derived from 7sl RNA. Genetica 1994, 93, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Brindle, P.K.; Montminy, M.R. The CREB family of transcription activators. Curr. Opin. Genet. Dev. 1992, 2, 199–204. [Google Scholar] [CrossRef]
- Tsukada, J.; Yoshida, Y.; Kominato, Y.; Auron, P.E. The CCAAT/enhancer (C/EBP) family of basic-leucine zipper (bZIP) transcription factors is a multifaceted highly-regulated system for gene regulation. Cytokine 2011, 54, 6–19. [Google Scholar] [CrossRef]
- Cook, T.; Gebelein, B.; Urrutia, R. Sp1 and its likes: Biochemical and functional predictions for a growing family of zinc finger transcription factors. Ann. N. Y. Acad. Sci. 1999, 880, 94–102. [Google Scholar] [CrossRef]
- Gogolevskaya, I.K.; Stasenko, D.V.; Tatosyan, K.A.; Kramerov, D.A. Influence of 5′-flanking sequence on 4.5SI RNA gene transcription by RNA polymerase III. Genome 2018, 61, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Koval, A.P.; Veniaminova, N.A.; Kramerov, D.A. Additional box B of RNA polymerase III promoter in SINE B1 can be functional. Gene 2011, 487, 113–117. [Google Scholar] [CrossRef]
- Koval, A.P.; Kramerov, D.A. 5′-flanking sequences can dramatically influence 4.5SH RNA gene transcription by RNA-polymerase III. Gene 2009, 446, 75–80. [Google Scholar] [CrossRef]
- Lundin, M.; Nehlin, J.O.; Ronne, H. Importance of a flanking AT-rich region in target site recognition by the GC box-binding zinc finger protein MIG1. Mol. Cell. Biol. 1994, 14, 1979–1985. [Google Scholar] [CrossRef] [Green Version]
- Osada, S.; Yamamoto, H.; Nishihara, T.; Imagawa, M. DNA binding specificity of the CCAAT/enhancer-btableinding protein transcription factor family. J. Biol. Chem. 1996, 271, 3891–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Pascale, F.; Nama, S.; Muhuri, M.; Quah, S.; Ismail, H.M.; Chan, X.H.D.; Sundaram, G.M.; Ramalingam, R.; Burke, B.; Sampath, P. C/EBPbeta mediates RNA polymerase III-driven transcription of oncomiR-138 in malignant gliomas. Nucleic Acids Res. 2018, 46, 336–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiduschek, E.P.; Tocchini-Valentini, G.P. Transcription by RNA polymerase III. Annu. Rev. Biochem. 1988, 57, 873–914. [Google Scholar] [CrossRef] [PubMed]
- Marck, C.; Kachouri-Lafond, R.; Lafontaine, I.; Westhof, E.; Dujon, B.; Grosjean, H. The RNA polymerase III-dependent family of genes in hemiascomycetes: Comparative RNomics, decoding strategies, transcription and evolutionary implications. Nucleic Acids Res. 2006, 34, 1816–1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnokov, I.; Schmid, C.W. Flanking sequences of an Alu source stimulate transcription in vitro by interacting with sequence-specific transcription factors. J. Mol. Evol. 1996, 42, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Carnevali, D.; Bollati, V.; Fustinoni, S.; Pellegrini, M.; Dieci, G. Identification of RNA polymerase III-transcribed Alu loci by computational screening of RNA-Seq data. Nucleic Acids Res. 2015, 43, 817–835. [Google Scholar] [CrossRef] [Green Version]
- Joazeiro, C.A.; Kassavetis, G.A.; Geiduschek, E.P. Alternative outcomes in assembly of promoter complexes: The roles of TBP and a flexible linker in placing TFIIIB on tRNA genes. Genes Dev. 1996, 10, 725–739. [Google Scholar] [CrossRef] [Green Version]
- Niller, H.H.; Salamon, D.; Ilg, K.; Koroknai, A.; Banati, F.; Bauml, G.; Rucker, O.; Schwarzmann, F.; Wolf, H.; Minarovits, J. The in vivo binding site for oncoprotein c-Myc in the promoter for Epstein-Barr virus (EBV) encoding RNA (EBER) 1 suggests a specific role for EBV in lymphomagenesis. Med. Sci. Monit. 2003, 9, HY1–HY9. [Google Scholar]
- Fowlkes, D.M.; Shenk, T. Transcriptional control regions of the adenovirus VAI RNA gene. Cell 1980, 22, 405–413. [Google Scholar] [CrossRef]
- Piras, G.; Dittmer, J.; Radonovich, M.F.; Brady, J.N. Human T-cell leukemia virus type I Tax protein transactivates RNA polymerase III promoter in vitro and in vivo. J. Biol. Chem. 1996, 271, 20501–20506. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, A.; Rozhdestvensky, T.S.; Kuryshev, V.Y.; Schmitz, J.; Brosius, J. An unusual primate locus that attracted two independent Alu insertions and facilitates their transcription. J. Mol. Biol. 2005, 350, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, M.S.; Kramerov, D.A.; Tulchinsky, E.M.; Revasova, E.S.; Lukanidin, E.M. Activation of putative transposition intermediate formation in tumor cells. EMBO J. 1985, 4, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Spearow, J.; Rubin, C.M.; Schmid, C.W. Physiological stresses increase mouse short interspersed element (SINE) RNA expression in vivo. Gene 1999, 239, 367–372. [Google Scholar] [CrossRef]
- Leonova, K.I.; Brodsky, L.; Lipchick, B.; Pal, M.; Novototskaya, L.; Chenchik, A.A.; Sen, G.C.; Komarova, E.A.; Gudkov, A.V. p53 cooperates with DNA methylation and a suicidal interferon response to maintain epigenetic silencing of repeats and noncoding RNAs. Proc. Natl. Acad. Sci. USA 2013, 110, E89–E98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnokov, I.; Chu, W.M.; Botchan, M.R.; Schmid, C.W. p53 inhibits RNA polymerase III-directed transcription in a promoter-dependent manner. Mol. Cell. Biol. 1996, 16, 7084–7088. [Google Scholar] [CrossRef] [Green Version]
- Cairns, C.A.; White, R.J. p53 is a general repressor of RNA polymerase III transcription. EMBO J. 1998, 17, 3112–3123. [Google Scholar] [CrossRef] [Green Version]
- Shumyatsky, G.P.; Tillib, S.V.; Kramerov, D.A. B2 RNA and 7SK RNA, RNA polymerase III transcripts, have a cap-like structure at their 5′ end. Nucleic Acids Res. 1990, 18, 6347–6351. [Google Scholar] [CrossRef] [Green Version]
- Borodulina, O.R.; Kramerov, D.A. Short interspersed elements (SINEs) from insectivores. Two classes of mammalian SINEs distinguished by A-rich tail structure. Mamm. Genome 2001, 12, 779–786. [Google Scholar] [CrossRef]
- Borodulina, O.R.; Golubchikova, J.S.; Ustyantsev, I.G.; Kramerov, D.A. Polyadenylation of RNA transcribed from mammalian SINEs by RNA polymerase III: Complex requirements for nucleotide sequences. Biochim. Biophys. Acta 2016, 1859, 355–365. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NcRNA (Gene) 1 | TATA-like Box: Sequence 2, Position | −50 Elements: Name 3, Position, Sequences 4 | −90/−60 Elements 3 | References |
|---|---|---|---|---|
| Epstein–Barr virus: EBER 1 EBER 2 | ATGTAGAC −28/−22 GTATAGAG −28/−22 | Atf/Cre −51/−44 TGACGTAG TGACGTAG | Sp1 −65/−60 CCGCCC CCCGCC | [20,58] |
| Adenovirus VAI | CTAGACCG −30/−24 | Atf/Cre −38/−31 TGACGCTC | [59,60] | |
| 7SL-1 (Hs) 7SL-3 (Hs) | TTCTAGTA −29/−23 TTCTAGTG −30/−24 | Atf/Cre −50/−43 TGACGTCA TGACGTAA | Sp1 GGGCGG CCCGCC | [21], this study |
| vt | TCAAGAAA −30/−24 | Atf/Cre −53/−46 TGACGTAG | [22] | |
| BC200/G22 | CTATGAAA −32/−26 | about −80/−70 unidentified | [61] | |
| 4.5SH | TTCAAGTA −30/−24 | Atf/Cre −51/−44 TGTCGTCA | Sp1 Region: −87 to −60 GGGCGG 5 | This study |
| 4.5SI Mmu1ʹ 4.5SI Mmu2 | CTACATGA −30/−24 CTACTTCA −30/−24 | Atf/Cre −54/−47 TGACGTAC C/EBP −51/−42 GTTGCGCAAC | This study and [47] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatosyan, K.A.; Stasenko, D.V.; Koval, A.P.; Gogolevskaya, I.K.; Kramerov, D.A. TATA-Like Boxes in RNA Polymerase III Promoters: Requirements for Nucleotide Sequences. Int. J. Mol. Sci. 2020, 21, 3706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103706
Tatosyan KA, Stasenko DV, Koval AP, Gogolevskaya IK, Kramerov DA. TATA-Like Boxes in RNA Polymerase III Promoters: Requirements for Nucleotide Sequences. International Journal of Molecular Sciences. 2020; 21(10):3706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103706
Chicago/Turabian StyleTatosyan, Karina A., Danil V. Stasenko, Anastasia P. Koval, Irina K. Gogolevskaya, and Dmitri A. Kramerov. 2020. "TATA-Like Boxes in RNA Polymerase III Promoters: Requirements for Nucleotide Sequences" International Journal of Molecular Sciences 21, no. 10: 3706. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103706