1. Introduction
Viruses infecting bacteria have different strategies for the releasing of the produced progeny phage from the host cell. Most dsDNA and dsRNA bacteriophages use a holin-endolysin system to lyse their bacterial hosts and release new phage progeny [
1]. Among the most widely studied are bacteriophages infecting
Escherichia coli such as lambda (λ), T7, or phage 21 [
1,
2,
3,
4,
5]. Currently known lysis systems are encoded by two to five genes, organized in a so-called lytic cassette. Two of the genes code for the holin and the endolysin, that are directly involved in induction of cell lysis. The others gene products play an accessory role [
1].
The canonical lytic system of lambdoidal phages, such as λ phage, is composed of 5 proteins. The holin gene of λ phage contains a dual-start motif that allows the expression of two products of different lengths. One acts as an effector (holin), and the other as an inhibitor (antiholin) of the holin protein. Such dual start motif was also found for some other holin genes [
1,
3,
6,
7,
8]. Holins, controlled by anti-holin proteins, accumulate in the cytoplasmatic membrane without damaging the host cell. At a genetically determined time, the holins disrupt the inner membrane by forming one large hole, thereby allowing endolysins to reach and digest the peptidoglycan [
1]. In fact, holins act as regulators of the timing of host cell lysis. The role of endolysins is to destroy the cell wall. Two additional proteins, called spanins, participate in the destruction of the outer cell membrane [
9]. Lysis induced by such system is generally very rapid, and is completed within a few seconds [
1].
Phage endolysins are generally small globular proteins which act as transglycosidases, lysozymes, amidases, or endopeptidases [
1,
4,
10], but multi-functional endolysins have also been described [
11]. Most endolysins do not contain a secretory domain, accumulate in the cytoplasm, and depend on holins to pass the inner membrane and reach the peptidoglycan. In the last decade, a group of phage-encoded endolysins that contain a signal-arrest-release (SAR) domain were described [
1,
2,
12]. The SAR domain initially acts as an export signal domain, which directs the endolysin to the periplasm. In export of the SAR-endolysins is involved the host secretion (
sec) system [
1,
2,
13]. However, usually SAR-endolysins are not processed by signal proteases and not released in the periplasm but accumulate in inactive, membrane-tethered form. Their activation is correlated with the release of these enzymes from the membrane and the refolding to an active form. The release may be linked to the membrane depolarization under action of the associated holin, called pinholin (for review see [
1]). As opposed to large-hole-forming holins, pinholins form small holes that depolarize the inner membrane. Recently, the structure of the lethal pinhole of phage 21 was resolved and holes were <2 nm in diameter [
6].
Only few systems encoding SAR-endolysin and pinholin were described. The most studied is the lytic cassette encoded by lambdoid phage 21 specific for
E. coli, composed of pinholin, antiholin, and SAR-endolysin [
2]. In addition, other studies described the existence of lytic cassettes, composed of pinholin and SAR-endolysin, encoded by different phages as ΦKMV, that infects
Pseudomonas aeruginosa [
13], Xfas53 phage infecting
Xylella fastidiosa [
14], coliphage P1 [
12] or KBNP1315 phage that infects Avian Pathogenic
E. coli (APEC) [
15]. The regulation of SAR-endolysin release seems to be complex but associated with pinholin. Other possibilities of lytic mechanism certainly exist: for example, LydA pinholin is not essential for P1 phage release [
12] and the oenococcal phage fOg44 encodes a SAR-endolysin with a cleavable signal sequence [
16].
Seven biologically active dsDNA phages have been described as infecting
Haemophilus influenzae, which is an obligate commensal of the upper respiratory tract in humans but may also be responsible for upper respiratory tract infections and meningitis [
17]. Among them, there is the HP1 phage, that belongs to the
Myoviridae family. HP1 phage genome (NC_001697.1) has been sequenced and is 32 kbp long, encoding 41 potential proteins in five plausible transcriptional units [
18]. The proposed lytic module includes the potential
lys and
hol genes, encoding potential HP1 endolysin and holing [
18].
In the present study, we characterized empirically the proteins encoded by the potential lytic cassette of H. influenzae HP1 bacteriophage. We demonstrated that the HP1 lytic system contains a SAR-endolysin and a pinholin with unusual topology, when activated in the inner membrane. HP1 pinholin seems to participate in control of SAR-endolysin and is necessary to lysis of host cells in the native host.
2. Results
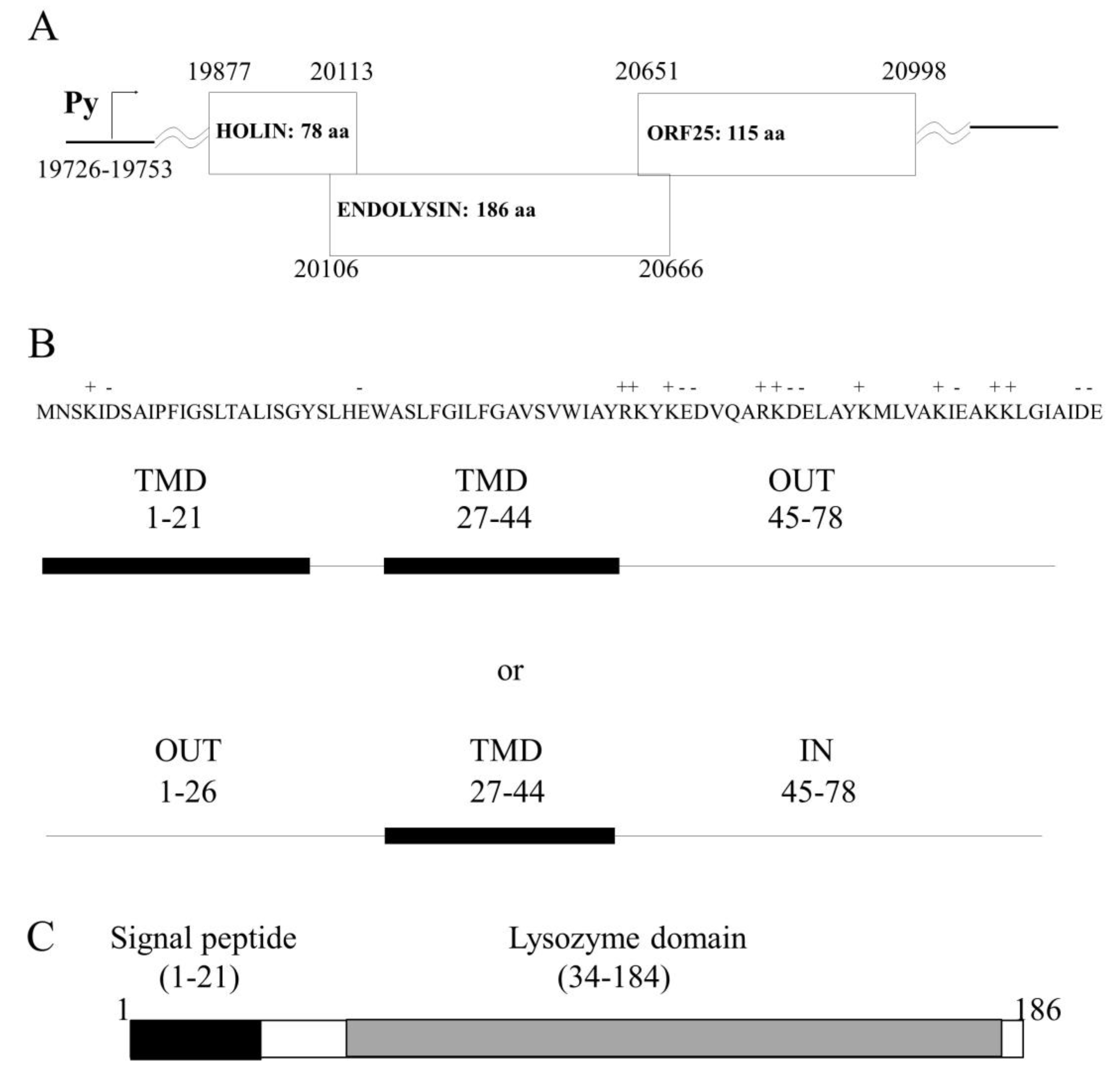
2.1. In silico Analysis of the HP1 Phage Lysis Cassette Showed Two Putative Lysis Genes
The complete nucleotide sequence of the temperate phage HP1 infecting
H. influenzae was previously determined and the host cell lysis system would be encoded by two open reading frames (ORF), which are transcribed as a late phage transcript [
18] (
Figure 1A). The
hol and
lys genes are located between HP1p29 (ORF24), which encodes for protein showing 100% identity to the tail tube protein of HP2 phage and HP1p32 (ORF25), coding for a protein which has not yet been characterized [
18].
Our analysis demonstrates that HP1p32 protein (121 amino acids) shows similarity to LysB protein (Sequence ID: KGQ31227.1) from
Gallibacterium genomospecies (33% identities, 55% positive). HMMER [
19] and TMHMM algorithm analysis revealed the existence of a signal peptide sequence (residues 1 to 25) and a coiled coil region (residues 41 to 93) but no known domain was detected in this protein. HP1p32 cellular localization could not be predicted using in silico algorithms (PSORTb, [
20]).
The
hol gene is the first gene in the lytic cassette and it sequence overlaps by 7 nucleotides the sequence of the gene
lys (
Figure 1A). The analysis of the
hol gene sequence indicated that the
hol gene does not present dual start-motif and HP1 phage seems not to encode an antiholin protein. The predicted, encoded protein is 78 aa long. Blast comparison of predicted HP1 holin showed significant homology to a holin-like protein from
Haemophilus quentini WP_005642949 (96% identities, 98% positive), protein encoded by
Haemophilus haemolyticus WP_11878507 (95% identities, 96% positive), and holin from
Rodentibacter pneumotropicus WP_136125977 (74% identities, 87% positive) and also others putative holins, but none of these proteins was characterized empirically in vitro or in vivo. Holins are transmembrane proteins and our analysis with the PSORTb algorithm tool [
20] predicted cytoplasmic membrane localization also for HP1 holin. Further, we performed several in silico analysis to predict the potential HP1 holin topology in the bacterial inner membrane. Using the SOSUI algorithm predicting the transmembrane helices [
21], we found two transmembrane domains (TMD) for HP1 holin (residues 1 to 21 and 27 to 44) (
Figure 1B). The existence of N-terminal TMD was also confirmed by DeepSig [
22]. In addition, analysis performed with Phyre2 indicated two TMD [
23]. The analysis with other algorithms predicting transmembrane helices (HMMTOP, TMHMM, TMpred, Phobius) indicated that the HP1 holin would have only one TMD (residues 27–44) and that the N-terminal, domain, would be located in the periplasm and the C-terminal domain in the cytoplasm (
Figure 1B). Such topology was already proposed by Reddy et al. [
24], which predicted that HP1 holin (ID 1.E.7.1.2) is a single-pass membrane protein (TMD formed by 27–45 residues) and belongs to the type II holin superfamily (Transporter Classification Database (TCDB);
www.tcdb.org [
24]).
The HP1 endolysin was predicted to be encoded by the
lys gene. The enzyme consists of 186 amino acid residues, and is highly homologous to the lysozymes, encoded by different bacteria and among them to lysozymes encoded by phages (96% identity with
Haemophilus phage HP2 Lys protein NP_536831.1; 49% identity with
Enterobacteria phage P1 Lyz protein 1XJU_A, 48% with
Pasteurella phage F108 Lys protein AAZ93664.1, and 37% with endolysin of bacteriophage K139 AAL47527.1, which infects
Vibrio cholerae). Only one conserved domain of endolysin-autolysin (phage-related lysozyme and muraminidase-COG3772) was found from residue 34 to 184 (
Figure 1C). The SOSUI algorithm predicted a soluble protein. HMMTOP and TMHMM algorithms predicted an N terminal alpha-helix (6–22 residues). The analysis on the N-terminal sequence revealed a stretch of 16 uncharged residues. Such amino terminal extensions typically function as unprocessed transmembrane helices or as cleaved signal sequences for interaction with the secretion machinery [
12,
25]. As suggested by the SignalP [
26] algorithm, this α-helix would be a part of the signal peptide domain (residues 1–22) and its presence would enhance the insertion of the signal peptide into the phospholipid double layer (
Figure 1C). These predictions were also confirmed by iPSORT (residues 1 to 30) and by HMMER (signal peptide residues 1 to 24 and lysozyme domain: residues 60 to 178). All analyses suggested that HP1 endolysin is a SAR (signal-arrest-release) endolysin and has lysozyme activity.
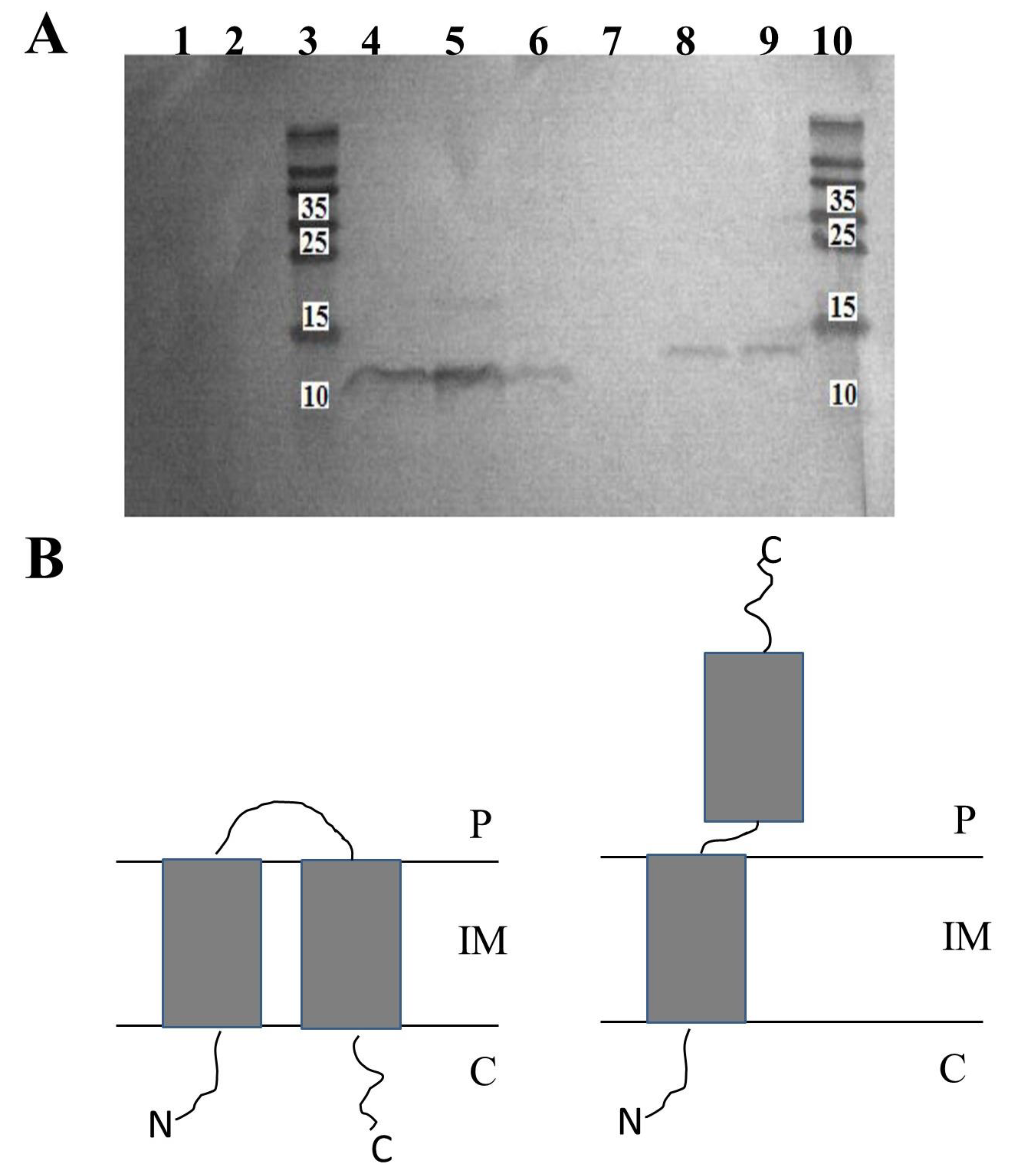
2.2. Overexpressed HP1 Holin Has C-out, N-in Topology in Bacterial Inner Cell Membrane
Since the in silico analysis was not conclusive, we decided to resolve experimentally the issue of the topology of HP1 holin in the bacterial inner membrane.
For this task, we constructed two plasmids on the basis of pET28a(+) vector with His-tag fused to the N- (pNHisHol) or to the C-terminus (pCHisHol) of HP1 holin.
From E. coli BL21(DE3) cultures carrying pNHisHol or pCHisHol constructs, we prepared inverted membrane vesicles (IMV, exposure of intracellular terminus outside) or spheroplasts (exposure of extracellular terminus outside). C- or N-terminus present on the external surfaces of these structures were proteolyzed by adding proteinase K. Then, membrane-associated peptides were isolated and separated by SDS-PAGE. Next, Western blotting, with specific antibodies directed to His-tag, was performed to determine which variant of holin, with His-tagged C- or N-terminus, remained in proteinase-treated IMV or spheroplasts.
As shown in
Figure 2, the detection of His-tagged termini was successful only in case of induction of N-His-tagged or C-His-tagged holin expression by IPTG (
Figure 2A, lines 6 and 9, respectively). In extracts prepared from non-induced cells (negative control), His-tagged holins were not detected (
Figure 2A, lines 1 and 2). Going further, we have detected C-terminal His-tag of HP1 holin in protein extracts from IMV, even after the treatment with proteinase K (
Figure 2A, lines 4 and 5). As the IMV surface represents the cytoplasmatic surface of the cell membrane, the observation that C-terminus is protected from the proteinase K activity indicated that C-terminus of HP1 holin is oriented to the periplasm. On contrary, the His-tagged N-termini were not detected in the samples of membrane proteins isolated from IMV treated with proteinase K (
Figure 2A, line 7), whereas such detection was possible in the case of IMV untreated with proteinase K (
Figure 2A, line 8) or membrane proteins from whole cell lysates (
Figure 2A line 9). These results indicate that N-terminus of the HP1 holin is located on the cytoplasmic side of the cell membrane (sensible to proteinase K treatment in IMV).
Spheroplasts are structures formed only by inner membrane. Preparation of spheroplasts from E. coli cells expressing His-tagged holins was unsuccessful. We hypothesize that the inner membrane was in this condition too damaged to allow the preparation of spheroplasts and detection of His-tagged holins.
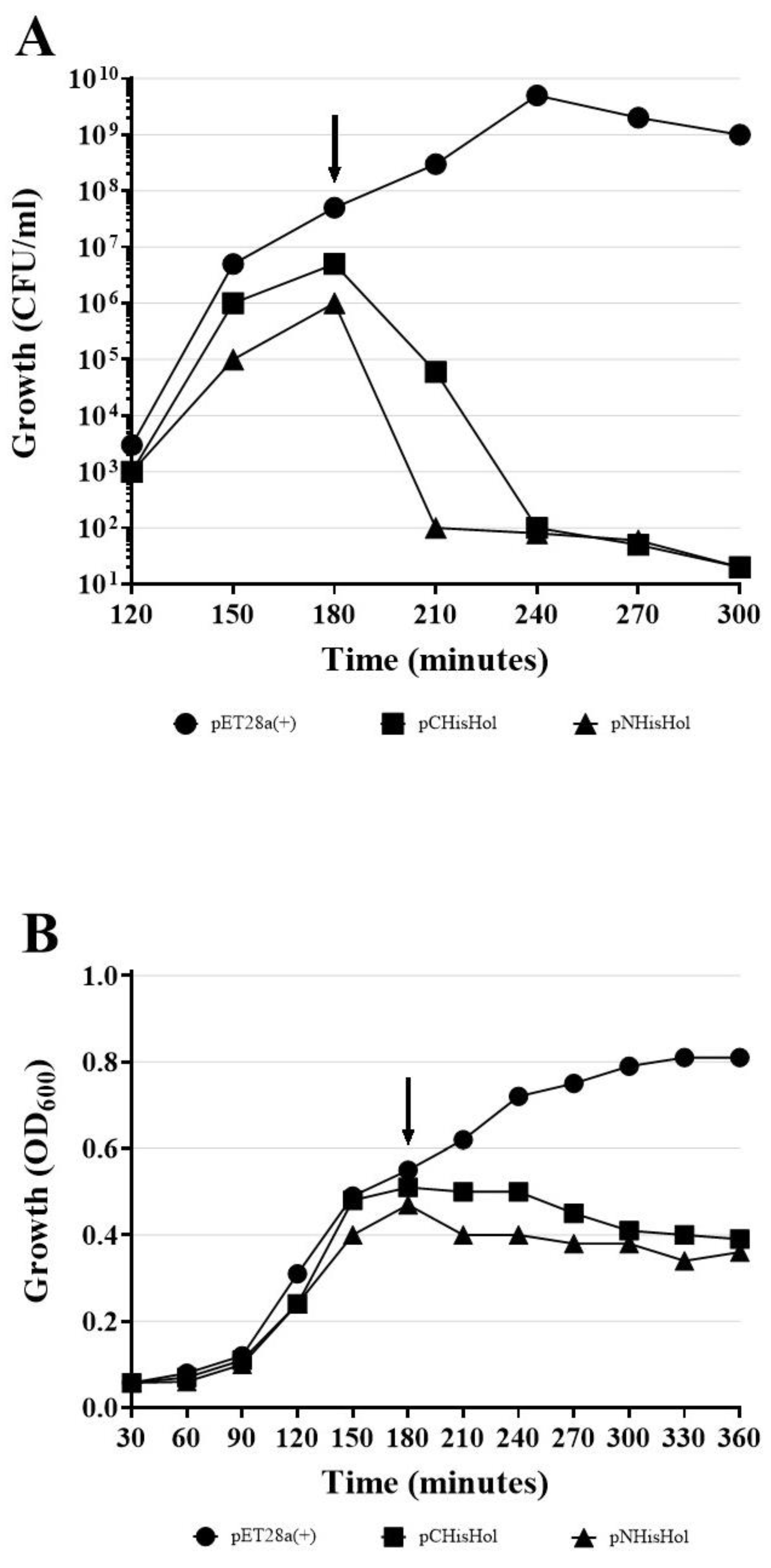
We also verified, that the addition of His-tag did not affect the HP1 holin conformation, since His-tagged HP1 holins induced cell death, as observed by colony-forming units (CFU/ ml) decrease after induction of expression without cell lysis (OD
600 of induced cultures remained constant), when overexpressed in
E. coli (
Figure A1) on the same way as wt HP1 holin did (see
Section 2.6).
All these data together demonstrated that HP1 holin is a transmembrane protein with C-terminus directed to the periplasm and N-terminus placed on the cytoplasmic side of the
E. coli cell membrane when overexpressed/activated (
Figure 2B).
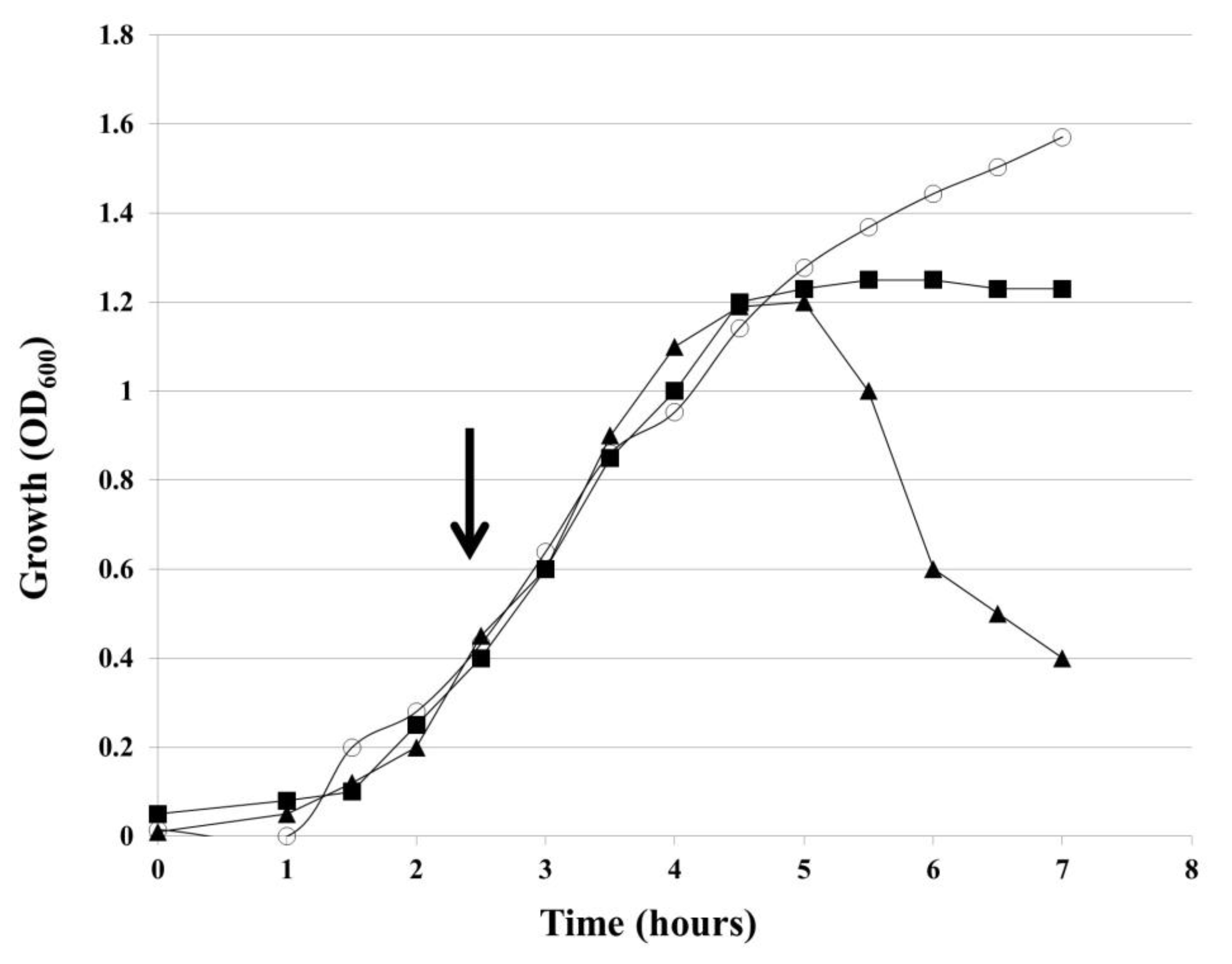
2.3. Inactivation of hol Gene Blocks the Lysis of H. influenzae Cells by HP1 Phage and Prevents the Release of New Phage Particles
H. influenzae Rd30 cells, lysogenized with wild-type HP1 (wt HP1) or
hol-deficient HP1 phage (HP1Δ
hol), were grown in liquid culture and OD
600 of the culture was measured every 30 min. After OD
600 of lysogenic bacteria reached ~0.4, mitomycin C was added in order to induce the prophage excision and to start the lytic cycle. As shown in
Figure 3,
H. influenzae (wt HP1) cells began to lyse 2.5 h after induction of prophages by mitomycin C,
H. influenzae (wt HP1). After reaching the peak of 1.2, OD
600 dropped to 0.4 within 5 h after induction. In contrast, HP1Δ
hol-lysogenized cells continued growing and reached a plateau 2.5 h after the addition of mitomycin C. Such an observation suggests that the product of the
hol gene is necessary for the release of new HP1 phage progeny from host bacterial cells.
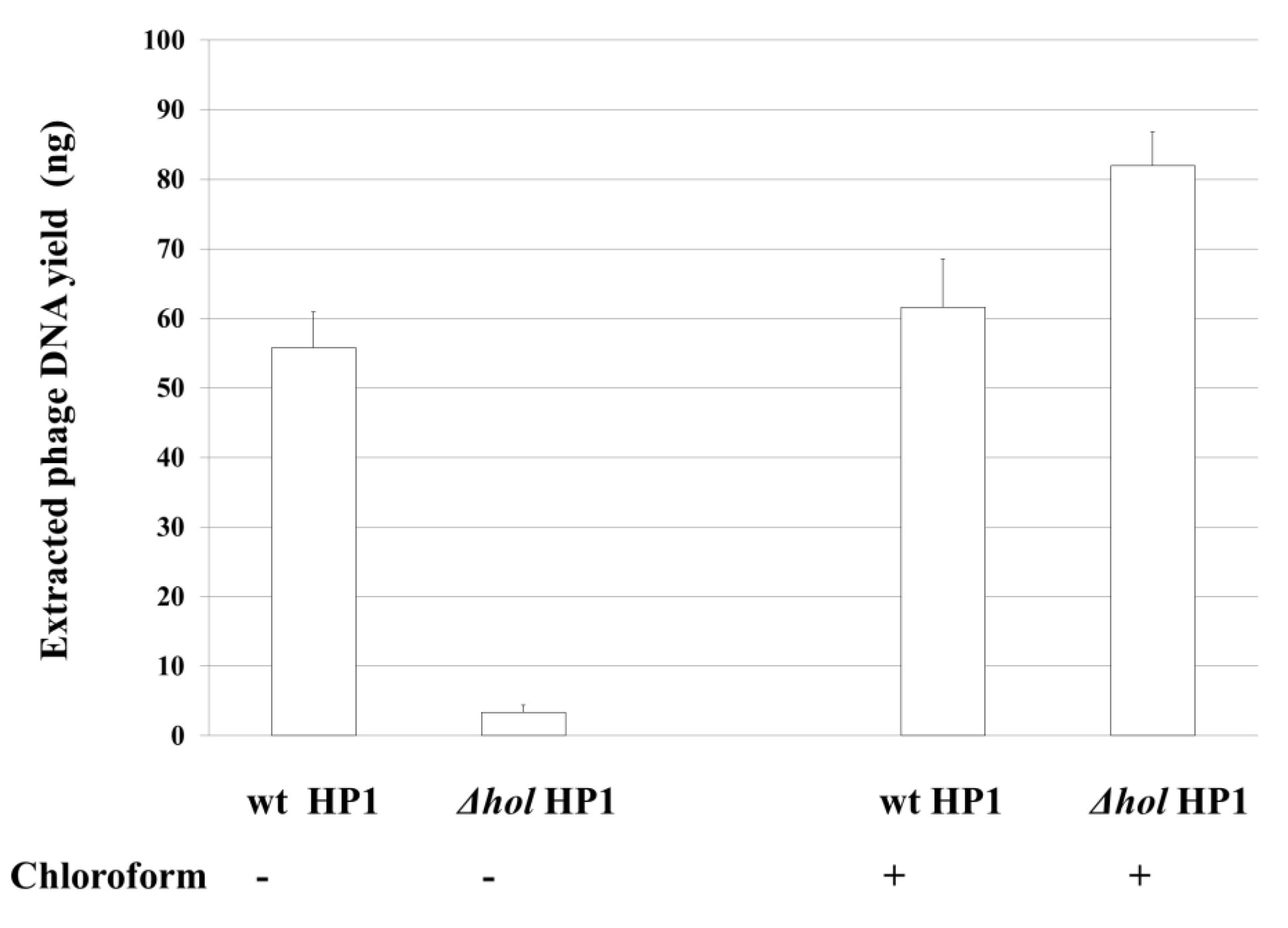
Next, we isolated phage particles from culture supernatants and after removing of bacterial DNA and RNA, we performed phage DNA extraction from bacteriophage capsids. As shown in
Figure 4, 55.75 (±5.20) ng of phage DNA was isolated from 50 mL of initial culture of induced
H. influenzae Rd30 (wt HP1) cells, while only 3.4 (±1.00) ng of phage DNA was isolated in the case of cells lysogenized with
hol-deficient HP1 bacteriophage. As chloroform induces bacterial lysis without affecting the bacteriophage structure, we also extract phage DNA from both cultures treated after chloroform treatment. As shown in
Figure 4, the extracted phage DNA from HP1Δ
hol-lysogenized cells reached in this case 82 (±4.8) ng, while from wt HP1 we have isolated 61.6 (±6.9) ng. So, the absence of lysis of cells lysogenized by HP1Δ
hol phage (
Figure 3) is associated with extremely low phage release into culture supernatants as demonstrated by phage DNA extraction from liberated phages. HP1Δ
hol phages may be liberated by host lysis with chloroform.
Thus, the deletion of the hol gene and absence of HP1 holin prevented the decrease of culture turbidity and the effective release of new bacteriophage particles from mitomycin C-induced H. influenzae (HP1Δhol). HP1 holin is indispensable in liberation of HP1 bacteriophage from H. influenzae cells.
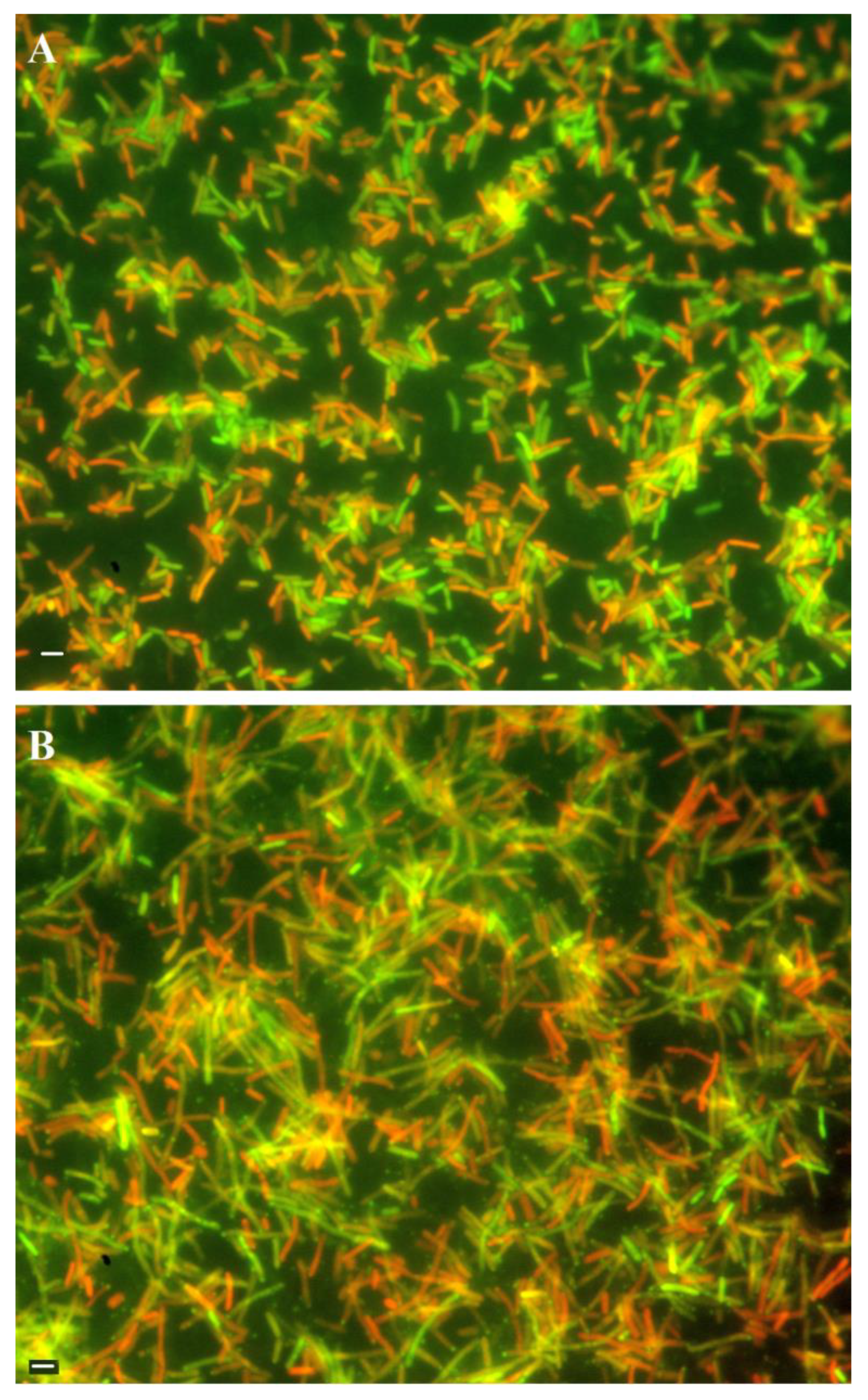
2.4. HP1 Holin Expression during Lytic Cycle of HP1 Phage Causes Reduction in H. influenzae Viability
Bacteriophage lytic cycle was induced with mitomycin C for 2.5 h (
Figure 3) in wt HP1 or HP1Δ
hol-lysogenized
H. influenzae. At this time, the difference in OD
600 between wt HP1 and HP1Δ
hol-lysogenized
H. influenzae strains is 0.2 (
Figure 3). Cell viability was assessed using the LIVE/DEAD bacterial viability kit and the same number of cells was taken for the staining. As shown in
Figure 5A, in case of cells with induced wt HP1 replication, the number of dead red-stained cells was lower to this of alive green-stained cells (ratio ~0.8:1) as estimated by ImageJ [
27]. In case of cells with induced HP1Δ
hol phage replication cycle, cells were almost twice more elongated than induced
H. influenzae cells lysogenized by wt HP1 phage and despite the same number of cells taken for staining, this culture seemed more dense. HP1Δ
hol phage-containing bacterial cells were not uniformly stained: at the edge of viable bacteria a point with accumulated green stain was observed. In case of HP1Δ
hol induction, the estimated ratio between dead and alive cells seems to be lower (~0.46:1) than in case of wt HP1 phage, indicating that less cells died in case of replication of HP1Δ
hol phage than in case of wt HP1.
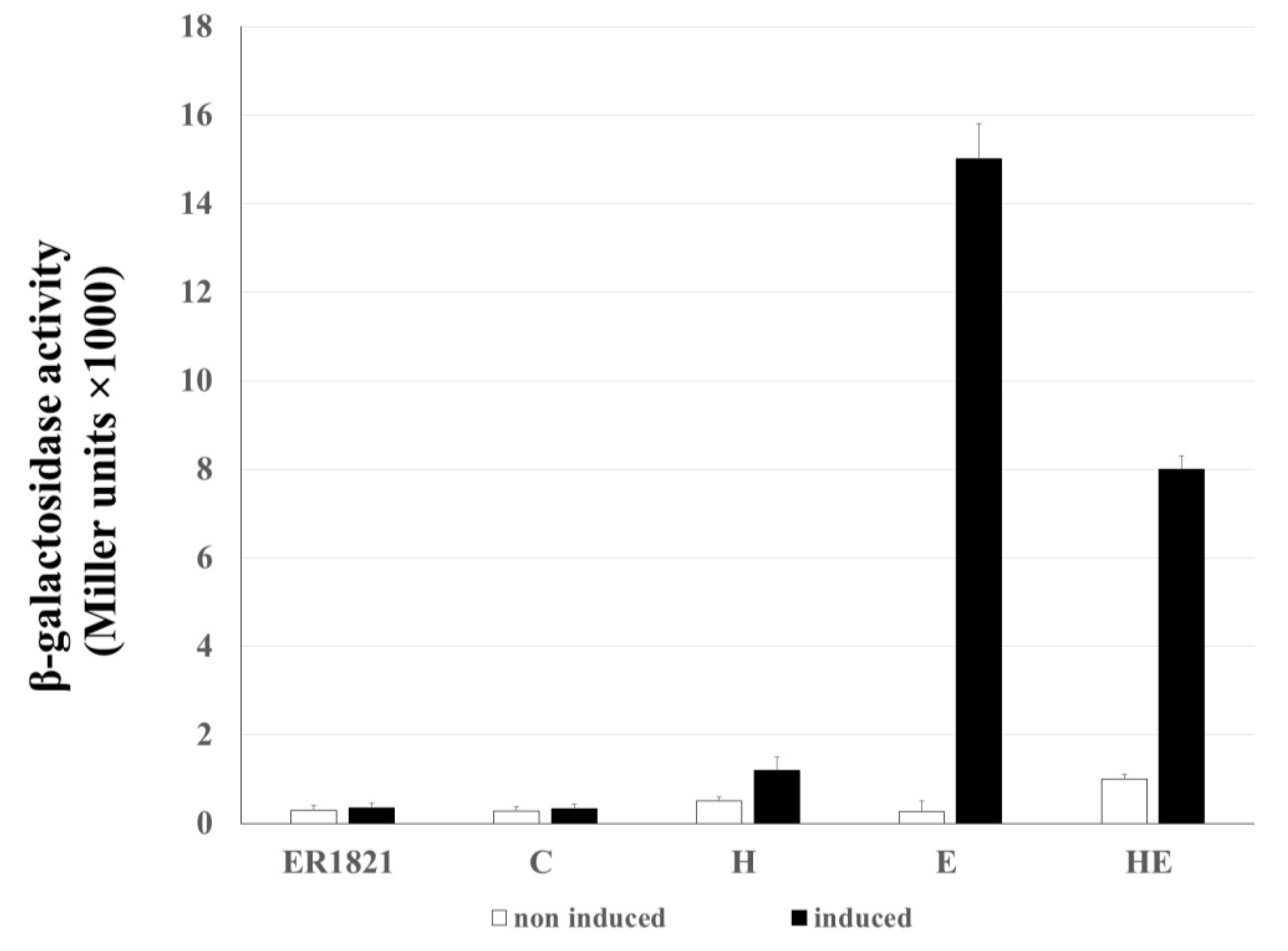
2.5. Membrane Lesions Induced by HP1 Holin Does not Allow the β-Galactosidase Leakage from E. coli Cells
The damage of the cytoplasmic membrane by lytic proteins was described to be sufficient to allow the leakage of cytoplasmic content, including large proteins like β-galactosidase [
28]. The ability of HP1 lytic proteins, encoded by genes
hol and
lys, in induction of lesion in host bacterial envelope was investigated by leakage of β-galactosidase from
E. coli cells (
Figure 6). For this, three constructs designated as H (pMPMT4::
hol), E (pMPMT4::
lys), and HE (pMPMT4::
hollys)
, respectively were introduced to
E. coli ER1821 cells, naturally expressing β-galactosidase. As control (C),
E. coli ER1821 carrying empty pMPMT4Ω vector were used, as well as cells without any vector.
Our results show that the increase in extracellular enzymatic activity of β-galactosidase was not observed when hol gene expression was induced: the lesions to the membrane made by holin alone are not large enough to allow the passage of large proteins such as β-galactosidase. The induction of the lys gene alone induced statistically significantly β-galactosidase activity (15,000 ± 800 Miller units, p < 0.01) in the culture supernatant, indicating β-galactosidase leakage through bacterial envelopes. The β-galactosidase activity increased 60 times as compared to uninduced cells coding the lys gene (p < 0.01). Thus, the overexpression of endolysin alone is sufficient to cause cell lysis even in absence of the HP1 holin.
When
hol and
lys genes were co-expressed, the amount of extracellularly released β-galactosidase was 1.875 lower (8000 ± 300 Miller units), than in the case of cells with the
lys gene expressed alone (
p < 0.01) (
Figure 6).
2.6. Expression of hol and lys Genes of HP1 Phage is Lethal to E. coli Cells
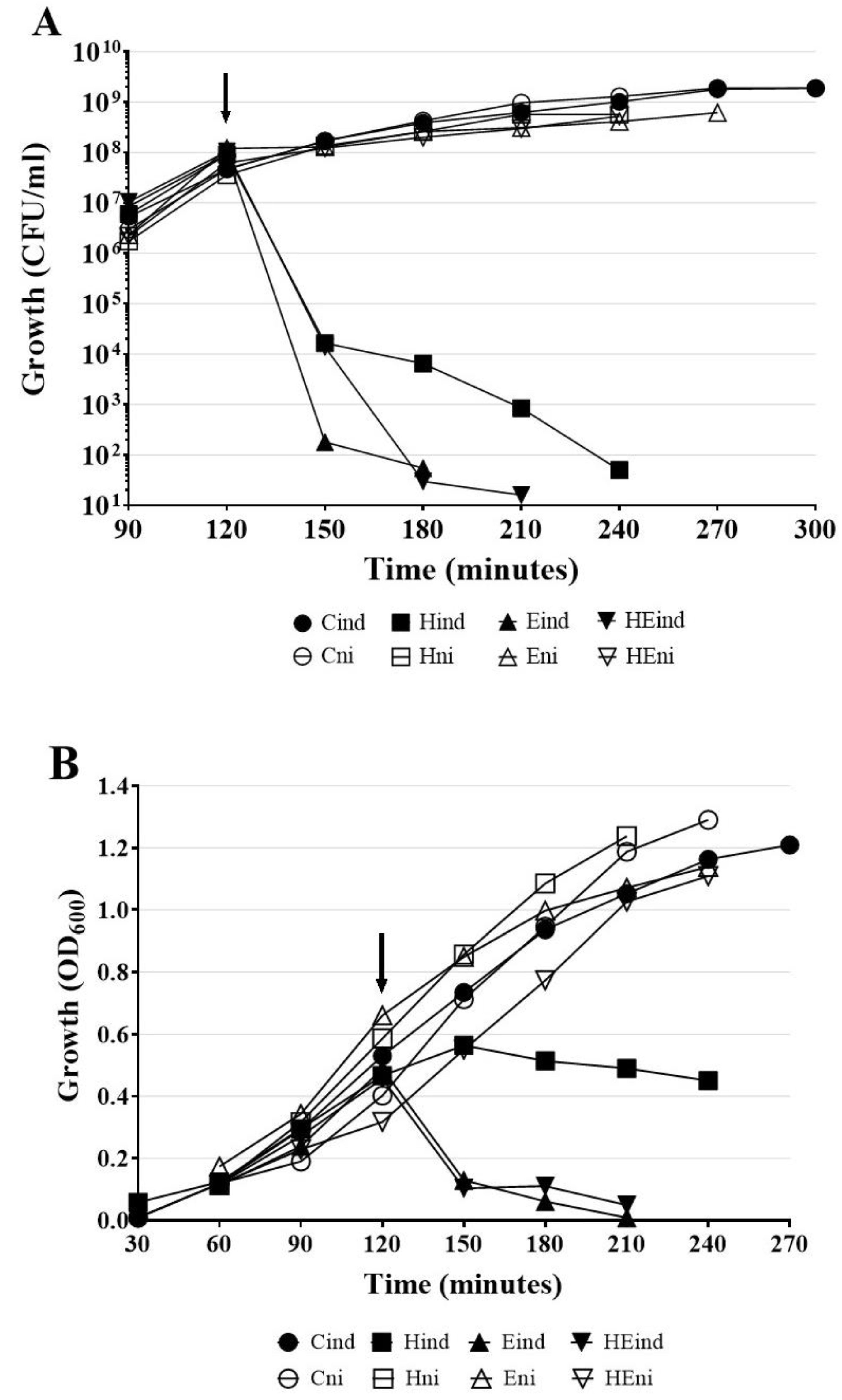
To test whether the expression of hol or lys genes by itself induces bacterial cells lysis, three constructs previously mentioned constructs: H (pMPMT4::hol), E (pMPMT4::lys), and HE (pMPMT4::hollys) were introduced to E. coli Top10 cells, which allow precise control of pBad promotor. As control (C), E. coli Top10 carrying empty pMPMT4Ω vector were used. We evaluated E. coli cell lysis by the measurement of OD600 and overall cell death by counting CFU/mL after induction of lytic genes by arabinose addition.
The lethal effect of the induced expression of HP1 lytic genes was already observed 30 min after the addition of arabinose (
Figure 7A,B, curves Hin, Ein, and HEin). In contrary, the non-induced cells remained viable for at least 3 h. Indeed, viable cells counts showed that after induction of
hol gene expression, the number of viable HP1 holin-expressing cells dramatically decreased from 9.1 × 10
7 CFU/mL at induction moment to 6.5 × 10
3 CFU/mL after 1 h and then to 5 × 10
1 CFU/mL after 2 h from induction (
Figure 7A). The measurement of liquid culture turbidity of
E. coli Top10 cells expressing the
hol gen product showed that OD
600 was constant at 0.450 within 2 h, after the induction of HP1 holin expression (
Figure 7B). This suggests that, in absence of other phage proteins, HP1 holin induced bacterial cell death but without causing cell lysis. When synthesis of the
lys gen product was induced in
E. coli Top10 cells, the number of viable cells in culture began to decrease even prior to induction. In 30 min from induction, the number of viable cells decreased from 9.15 × 10
7 to 1.8 × 10
2 CFU/mL and 1 h after induction the cell number dropped to 5.5 × 10
1 CFU/mL (
Figure 7A). During this time, the turbidity of the culture expressing the
lys gene also dramatically dropped from 0.48 (induction with arabinose time point) to 0.006, 1 h after induction (
Figure 7B). The induction of the
lys gene expression and synthesis of endolysin cause both, cell death (decrease in CFU/mL) and lysis (decrease of turbidity of the culture) in
E. coli cells. The above observations are consistent with the results obtained from the study of β-galactosidase leakage after induction of
lys gene expression (
Figure 6).
E. coli cells with induced expression of both
hol and
lys genes behaved similarly to cells carrying only the
lys gene (
Figure 7A,B). Within 60 min after induction, the number of viable HE plasmid-containing cells decreased from 1.04 × 10
8 to 3.0 × 10
1 CFU/mL. (
Figure 7A). The OD
600 also dropped from 0.456 to 0.11 during this time (
Figure 7B), while the OD
600 of control uninduced cells (HEni) raised from 0.340 to 0.77 during the same laps of time.
All control cultures carrying cloned genes, which expression was not induced by arabinose, had a similar growth profile as measured by viable cells counting (CFU/mL) or spectrophotometry (
Figure 7A,B, curves Hni, Eni, HEni). Control cells (C) containing only pMPMT4Ω vector maintained the growth rate characteristic for
E. coli (
Figure 7A,B). The number of cells increased exponentially and reached 1.88 × 10
9 CFU/mL (OD
600 = 1.3) 3 h after arabinose administration.
E. coli Top 10 cells carrying three constructs HP1 with lysis genes were also grown in the presence of 1% glucose to stop the expression from the pBad promoter. In this case, no decrease in turbidity or in CFU was observed: the growth was similar to control uninduced cultures (presented on
Figure 7A,B), indicating that no lysis occurred when the expression of cloned genes was blocked.
2.7. HP1-Induced Cell Lysis and New Phage Release Involve H. influenzae Sec System
Endolysins, which contain an SAR sequence, are exported in a membrane-bound state by the host
sec system, and subsequently are released in a soluble state into the periplasm. Thus, the release of HP1 phage particles from bacterial cells is presumed to involve the transport of the phage endolysin to the periplasm by the
H. influenzae sec system [
4]. Sodium azide (NaN
3) is the well-known inhibitor of the SecA ATPase activity, necessary for the translocation of exported protein across the membrane [
13,
29]. As we mentioned before, our predictions indicated that HP1 endolysin contains a SAR sequence. To study the potential involvement of the
sec system in HP1-induced lysis of the bacterial host, we used 1 mM NaN
3 to inhibit the HP1 phage progeny release from bacterial cells. Such concentration of sodium azide was not toxic to
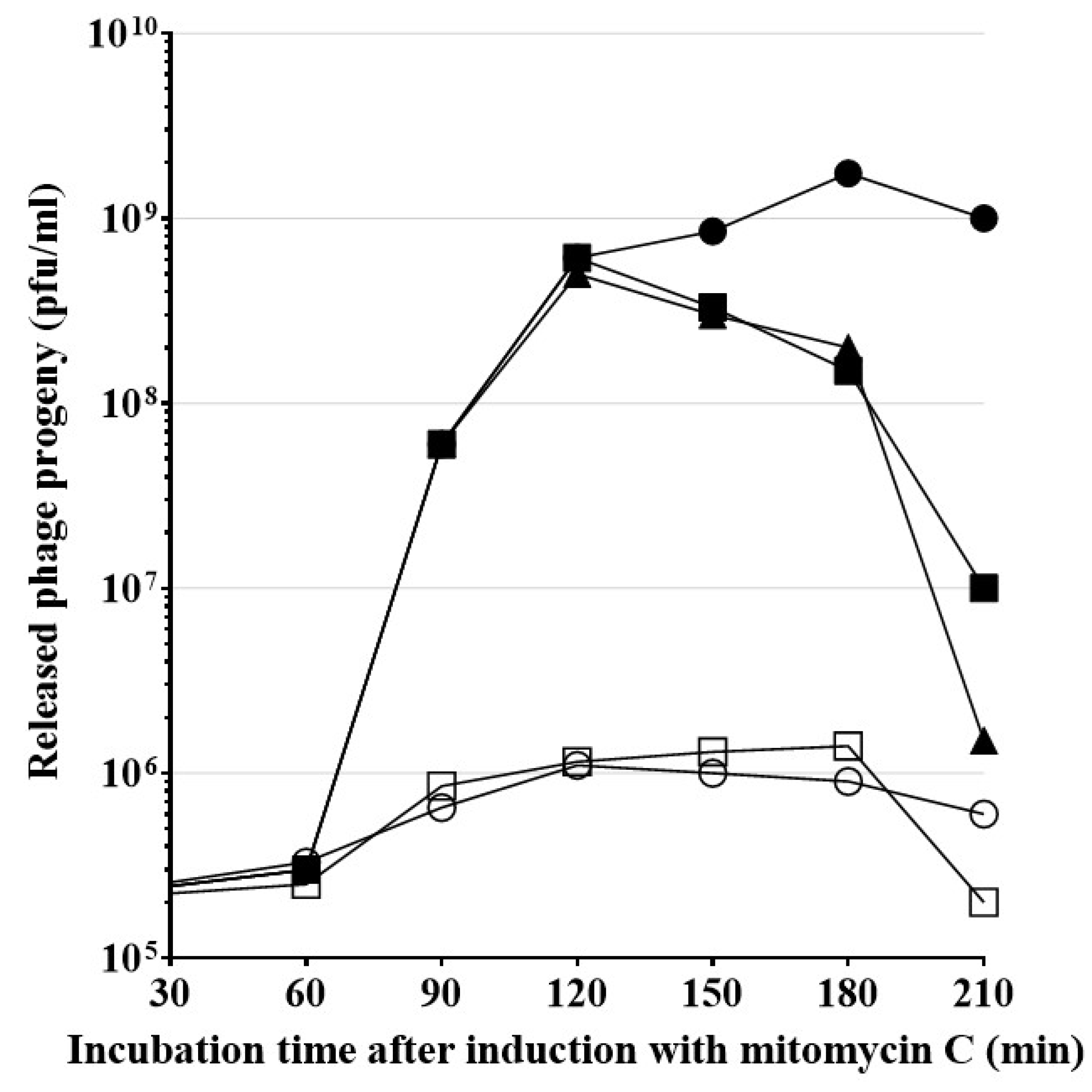
H. influenzae cells, when added to cell cultures (
Figure A2). Lysogenic
H. influenzae Rd30 (wt HP1) culture was treated with mitomycin C at OD
600 = 0.4, to induce the lytic cycle of the virus. As shown in
Figure 8, approximatively 60 min after induction, phage particles were released and counted as plaque-forming units (PFU/mL) in plaque assay. The peak of new phages release occurred 3–4 h after induction with mitomycin C and reached the level of 1.75 × 10
9 PFU/mL. The inhibition of the
sec system with 1 mM NaN
3 blocked the release of phage particles from mitomycin C-induced lysogenic
H. influenzae Rd30 (wt HP1) cells. As shown in
Figure 8, simultaneous addition of NaN
3 and mitomycin C or adding NaN
3, 60 min after induction, dramatically stopped the HP1 progeny release: the maximum of release was only about 2 × 10
5 and 6 × 10
5 PFU/mL, respectively. Addition of NaN
3 90 or 120 min after induction, first slowed the release of new phages, then their number declined rapidly. In this case, the endolysin, exported to periplasm by the
sec system before NaN
3 addition, ensured the release of new phage particles, but after addition of the
sec system inhibitor, no new endolysin is exported and released phage number was stopped.
Since inhibition of the sec system decreased the yield of release of new phage particles, our results strongly suggest that the sec system is involved in the HP1 phage release from the host H. influenzae cells, probably by participating in the transport of SAR-endolysin from the inner membrane to periplasm.
3. Discussion
Bacteriophage-induced lysis may be the source of released genetic information accessible for naturally competent bacteria, which are able to uptake foreign DNA from environment. Thus, phage-induced lysis participates in the horizontal gene transfer. Moreover, the current crisis of antibiotic-based treatments calls for alternative strategies for fighting bacterial infections and phage lytic proteins seem to be potential new antibacterial agents. In this context, it is not surprising that the mechanisms by which bacteriophages cause cell death and lysis are extensively investigated.
In this paper, we described the lytic cassette encoded by
H. influenzae-specific bacteriophage HP1. In addition to
hol and
lys genes, our in silico analysis of the whole HP1 genome reveals a third potential gene that may compose the HP1 lytic cassette: ORF25. Indeed, ORF25-encoded protein displays some similarity to LysB from
G. genomospecies. LysB is annotated as phage lysis regulatory protein and may be a spanin. However, our analysis of ORF25 protein did not reveal any structure similar to known spanins [
30].
Our topological predictions demonstrated that the
lys product is an SAR-endolysin. Overexpression of HP1 endolysin alone in
E. coli cells caused both, cell death and lysis, as demonstrated by the dropping number of CFU/mL and turbidity of cell culture. Moreover, we demonstrated that the release of HP1 phage particles from induced
H. influenzae cells (natural host) can be restrained by inhibiting the host
sec system with sodium azide, the SecA-inhibitor. Such an observation was also made for SAR-endolysins, encoded by different phages: SAR-endolysins are able to induce bacterial lysis without holin presence. For example, introduction of fOg44 lysin, Lys44, gene into
E. coli BL21(DE3) (pLysS) cells results in very unstable clones even in the absence of IPTG. This instability was due to lethal consequences of Lys44 lysin expression [
16]. It was also demonstrated that in the case of phage P1, a certain quantity of P1 SAR-endolysin spontaneously passed into periplasm and was active, without participation of holins [
31,
32]. In addition, the overexpression of SAR-endolysin from phage KNBP1315 induced the decline in the turbidity of
E. coli BL21(DE3) [
15]. Our data support that HP1 endolysin, like other SAR-endolysins, may access the peptidoglycan in the absence of holin and induce cell death and lysis. However, our results indicated that the lysis induced by SAR-endolysin alone occurred only in the case of overexpression of the
lys gene in
E. coli. In the native host of HP1 phage, i.e., during
H. influenzae infection, the participation of wild type HP1 holin protein is indispensable for lysis induction and phage progeny liberation took place only in the presence of HP1 holin. These in vivo studies suggested that holin may regulate the endolysin activity, since the interruption of the
hol gene is sufficient to inhibit the cell lysis and phage release in
H. influenzae cells. Moreover, the release of β-galactosidase was 1.875 times weaker in holin and endolysin expressing
E. coli cells than in the case of expression of endolysin alone. This last result may be due to different expression levels of the
lys gene alone in comparison to two overlapping
hol and
lys genes, but also may be due to a regulating role of HP1 holin in the endolysin-induced lysis process. Such role of pinholin in control of lysis was also described for ΦKMV phage [
13].
We demonstrated that the holes formed by HP1 holin are too small to permit the leakage of large proteins like β-galactosidase (520 kDa) from
E. coli cells. Similar observations were obtained in the case of S
2168 pinholin of phage 21 using fluorescent periplasmic marker TorA-GFP-SsrA [
2]. In contrast, lambda phage canonical holin supported the β-galactosidase flow from
E. coli [
2,
33]. β-galactosidase leakage also took place in the case of the expression of canonical holin from
Actinomyces naeslundii phage Av-1 in
E. coli [
28]. Membrane damage caused by HP1 holin seems to be insufficient to liberate the cell content, however sufficient to make impossible the preparation of spheroplasts. Observations under fluorescence microscope indicated that expression of the
hol gene in
H. influenzae Rd30 (induced by mitomycin C) was accompanied with increased mortality as compared to cells infected with mutant HP1 phage not expressing the holin. This phenomenon was already observed at the early stage of lysis of cells infected with wt HP1. In absence of
hol expression, the infected
H. influenzae cell shape was affected, probably by accumulation of new bacteriophages inside the cell (absence of cell lysis, confirmed by growing culture turbidity). Indeed, in
Figure 5, we can see green points indicating a large amount of DNA, probably phage DNA in HP1Δ
hol-lysogenized and induced
H. influenzae cells. The addition of chloroform to
H. influenzae infected with phage not expressing HP1 holin caused the membrane break down and liberated the progeny phage allowing the phage DNA extraction. This indicates that phage progeny was formed even in the absence of HP1 holin. Taking all these data together, HP1 holin rather does not cause lysis of the bacterial cell by itself (also confirmed by β-galactosidase assays and LIVE/DEAD staining), but seems to be indispensable to the lysis occurrence in vivo, which suggest its role in the control of the lytic process in native conditions. In
E. coli cells, the overproduction of HP1 holin caused a dramatical decrease in cell viability, but not in turbidity of liquid culture. All the data together indicate that the product of HP1
hol gene could be a pinholin associated with a SAR-endolysin.
In silico topological studies are not conclusive, indicating that HP1 holin may be a 2-TMD protein, with C-out topology or 1-TMD protein with N-out and C-in topology, as demonstrated in
Figure 1. The well-studied S
2168 pinholin has two TMD and reveals C-in, N-in topology [
5]. In activated S
2168, the first TMD is located in the cell membrane and the second TMD dimerizes to form holes: the activated S
2168 is C-in, N-out [
5]. The topology of other known pinholins is only predicted in silico and nothing is known about their active state topology. In our study, we prepared IMV from
E. coli cells overexpressing the HP1 holin. With this strategy, we demonstrated that overexpressed HP1 pinholin has C-out and N-in topology. The preparation of spheroplasts, the structures deprived of the outer membrane and peptidoglycan, was impossible. We cannot exclude that overexpressed pinholin, embedded in the inner membrane in the active state, form holes that disrupt the inner membrane which, in turn, shows too many discontinuities to create a stable structure. Indeed, we demonstrated that the overexpression of HP1 pinholin in
E. coli was insufficient to cause cell lysis (constant culture turbidity), but the cell membrane perturbation was sufficient to cause cell death (abrupt reduction of CFU/mL).
The C terminal domain of HP1 holin is much more hydrophilic than the rest of the protein, contains almost 50% charged amino acids, and is slightly positively charged (
Figure 1). Usually, the positive-inside rule determines the topology of inner membrane proteins [
34]. However, T4 phage holin, with long and highly hydrophilic C-terminus, has a C-out, N-in orientation [
35]. The unusual periplasmic domain of T4 holin is involved in the control of lysis timing [
36]. The T4 antiholin protein, analogous to the S107 antiholin of phage λ, blocks the lysis by binding the C-terminus of T4 holin. T4 holin has only one TMD and by tis is different of HP1 holin [
7]. We hypothesize that the demonstrated unusual C-out and N-in topology of overexpressed HP1 pinholin reflects the active-lethal form of this protein i.e., the topology that forms holes after depolarization of the membrane (
Figure 1). The nonactive HP1 holin would have the same topology that S
2168 pinholin from phage 21, i.e., two TMD and C-in and N-in topology. Recently performed topological and phylogenetic analyses of phage holins and putative holins based on their average sizes, predicted topologies and organism source allowed to distinguish seven superfamilies comprising 52 families [
24]. HP1 holin topology is characteristic for class II holins. However, the activation of S
2168 consists in the exposure of N-terminal domain to the cytoplasm. In contrast, in the case of HP1 holin, the TMD2 and C-terminus would be externalized to the periplasm after its activation (
Figure 1.). It is not clear what is the signal triggering the activation of HP1 holin. In contrary to the gene encoding S
2168 pinholin from phage 21, the studied
hol gene from HP1 phage does not possess a dual start motif. This excludes the existence of a sequence in HP1 lytic cassette encoding a protein comparable to S
2171 antiholin. As we mentioned above, antiholin also regulates the activation of T4 phage holin with periplasmic C-terminus [
36]. We screened all other genes encoded by HP1 phage for their similarity with the known protein, but none was similar to an antiholin. Possibly, the activation of lysis is the effect of the membrane depolarization as consequence of HP1 increased concentration in the inner membrane. Evaluation of the effect of HP1 lytic genes on β-galactosidase leakage suggested the control of lysis by holin (the leakage is inhibited in case of holin and endolysin co-expression in comparison to endolysin alone, but the mechanism remains unknown). Further studies should be performed to explain the exact mechanism of activation of HP1 holin and subsequent depolarization of the inner membrane.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}