Life in the Current: Anatomy and Morphology of Utricularia neottioides

 ,
,  , and
, and 
Abstract
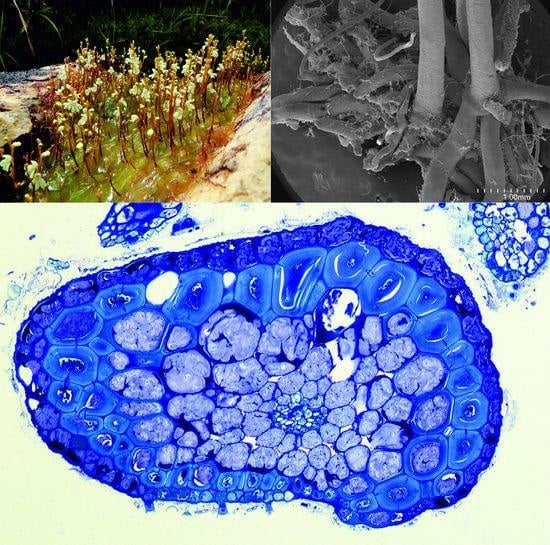
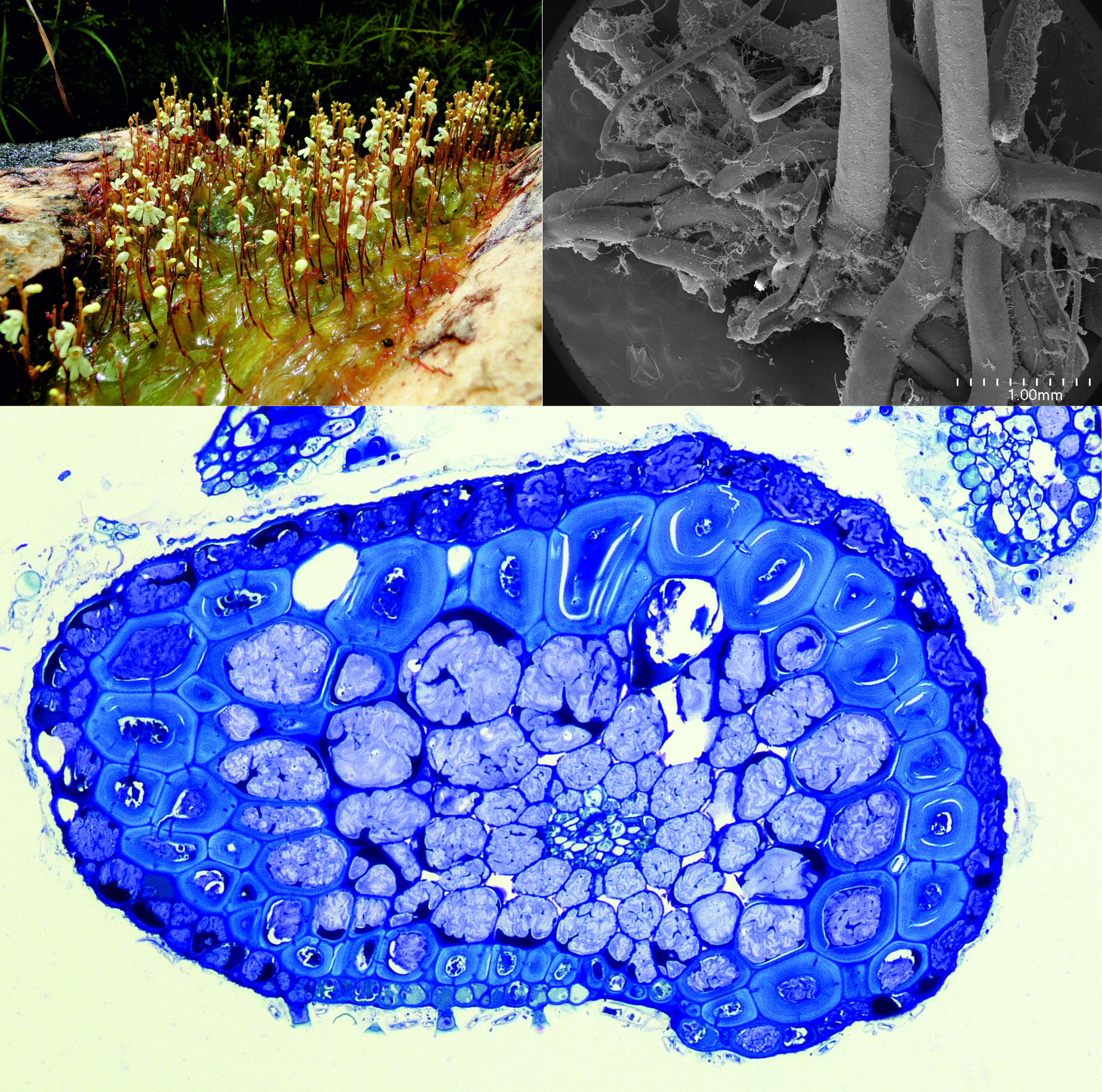
:
1. Introduction
2. Results
2.1. Utricularia neottioides
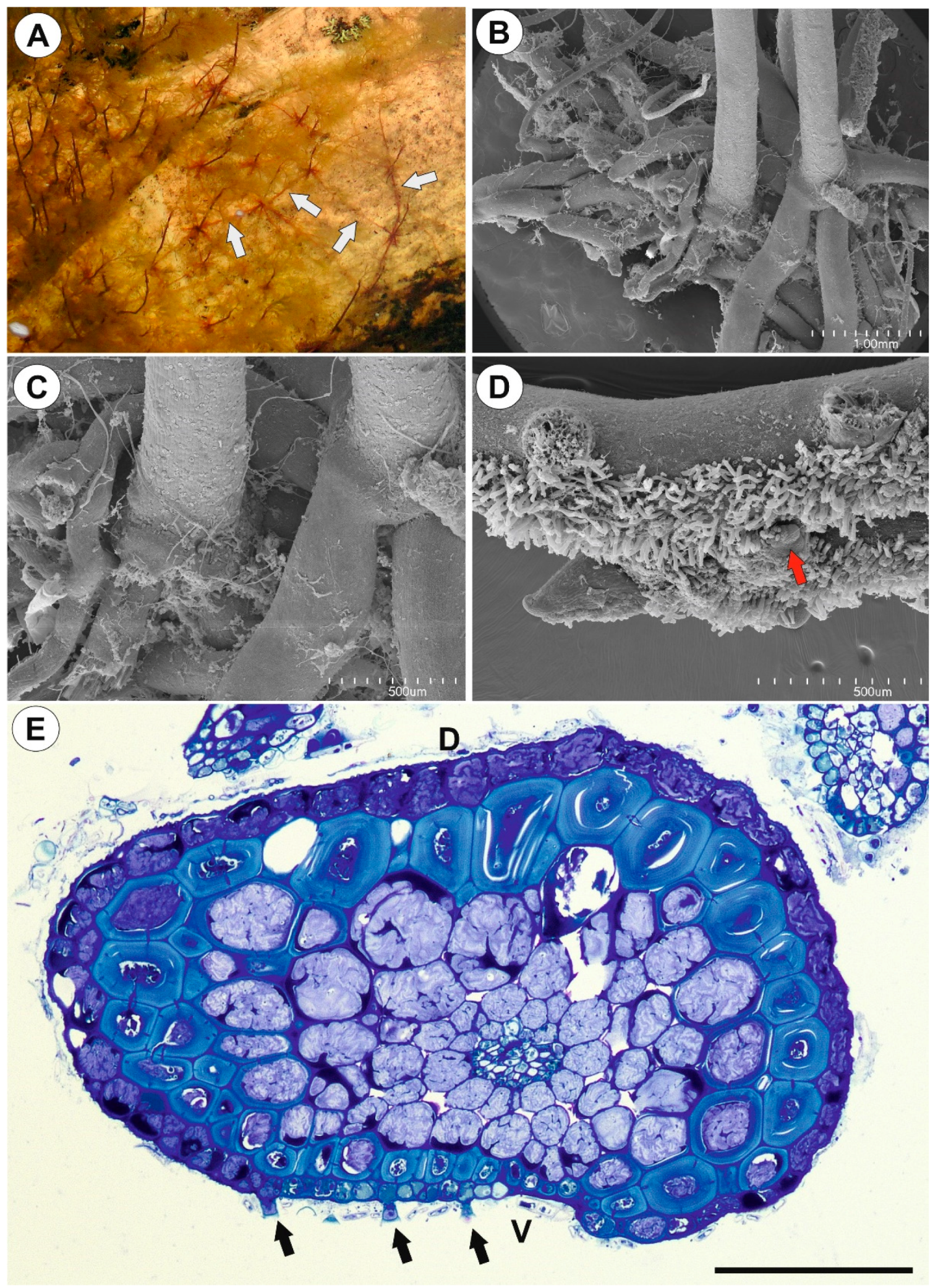
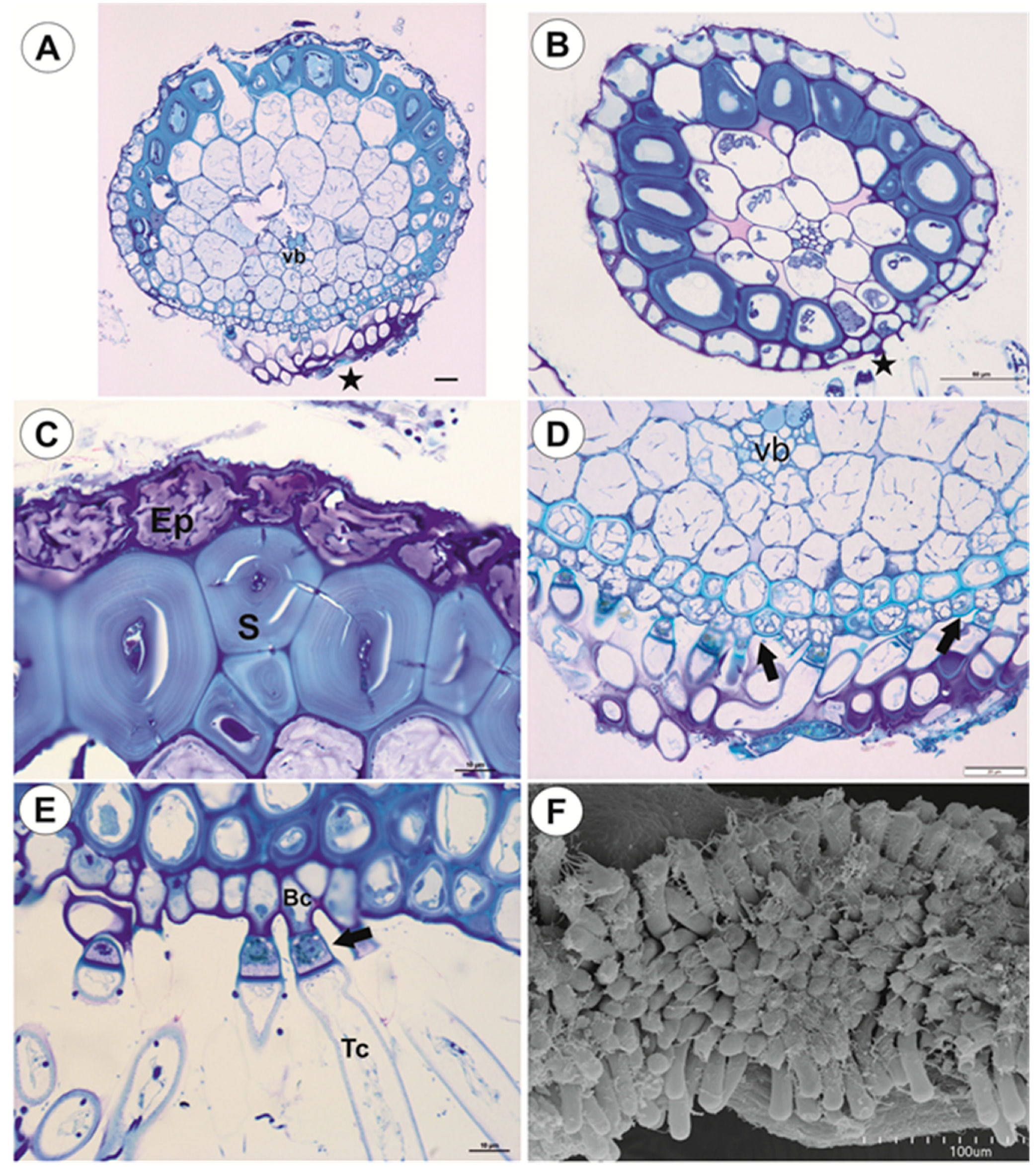
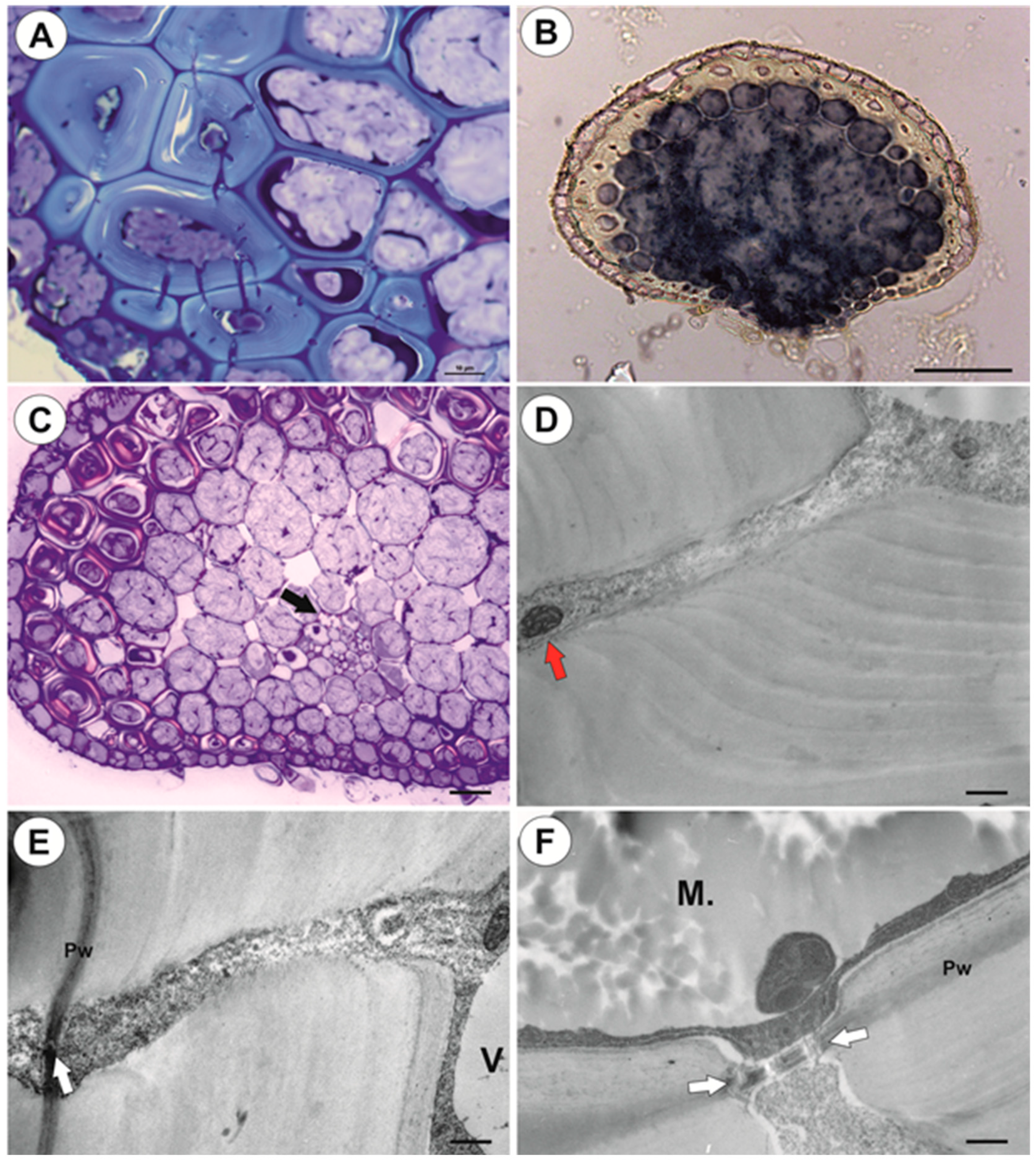
2.1.1. Anchor Stolons (Claw-Like ‘Rhizoids’) Fixing Plant
2.1.2. Running Stolons
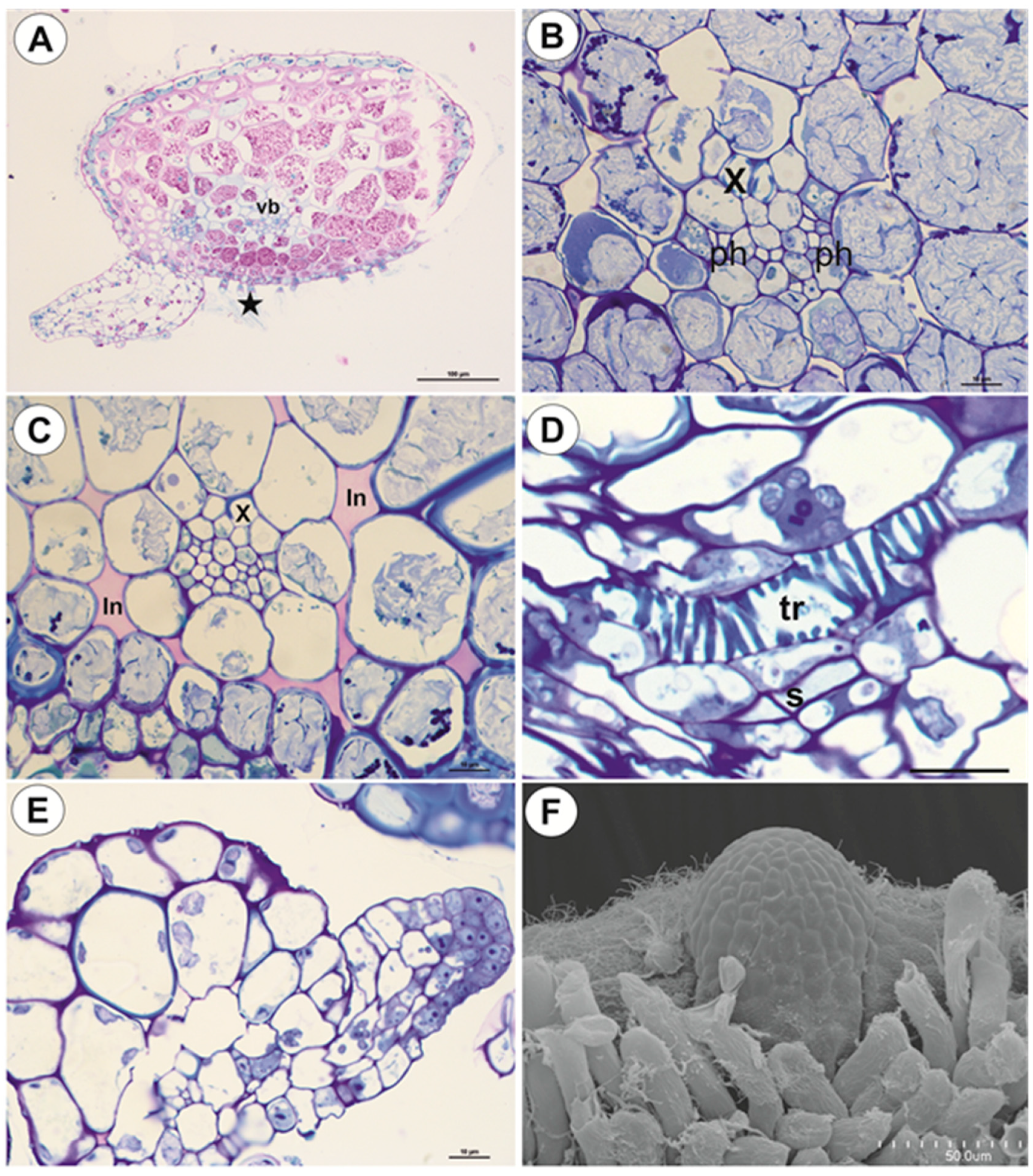
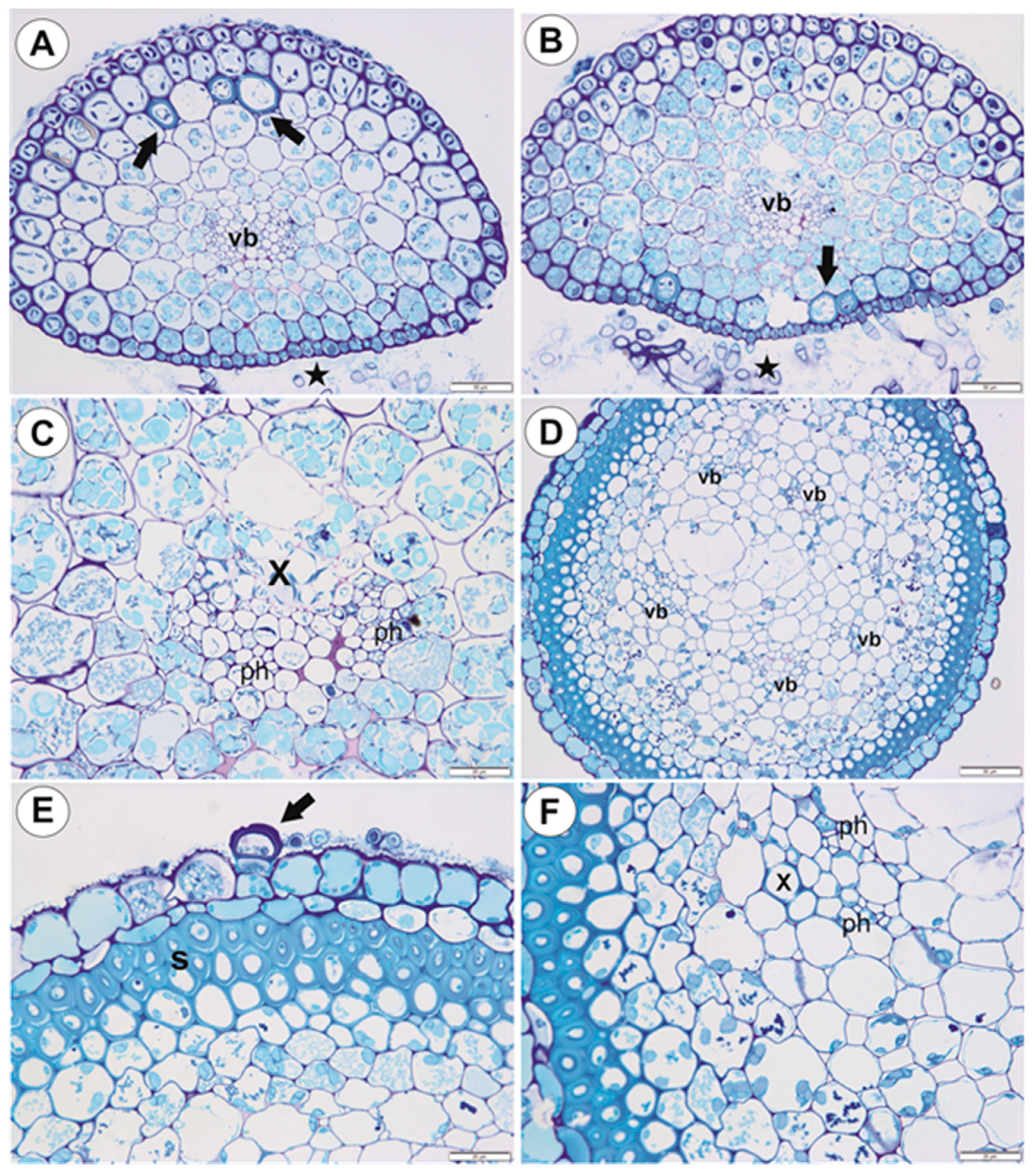
2.1.3. Basal Portion of Inflorescence Stalk-Stolon Bearing Leaf-Like Shoots and Forming Inflorescence
2.1.4. Filamentous Leaf-Like Shoots
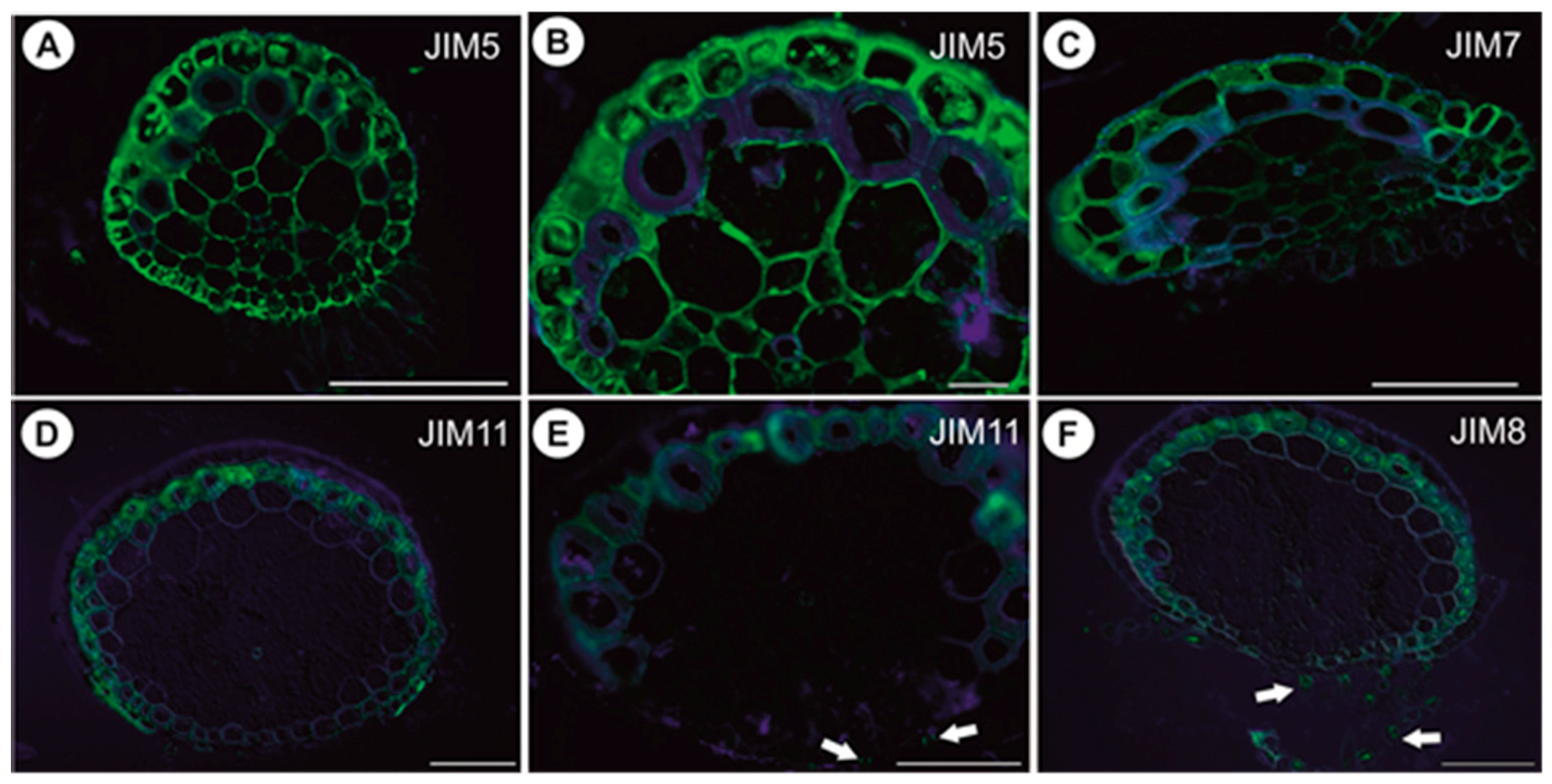
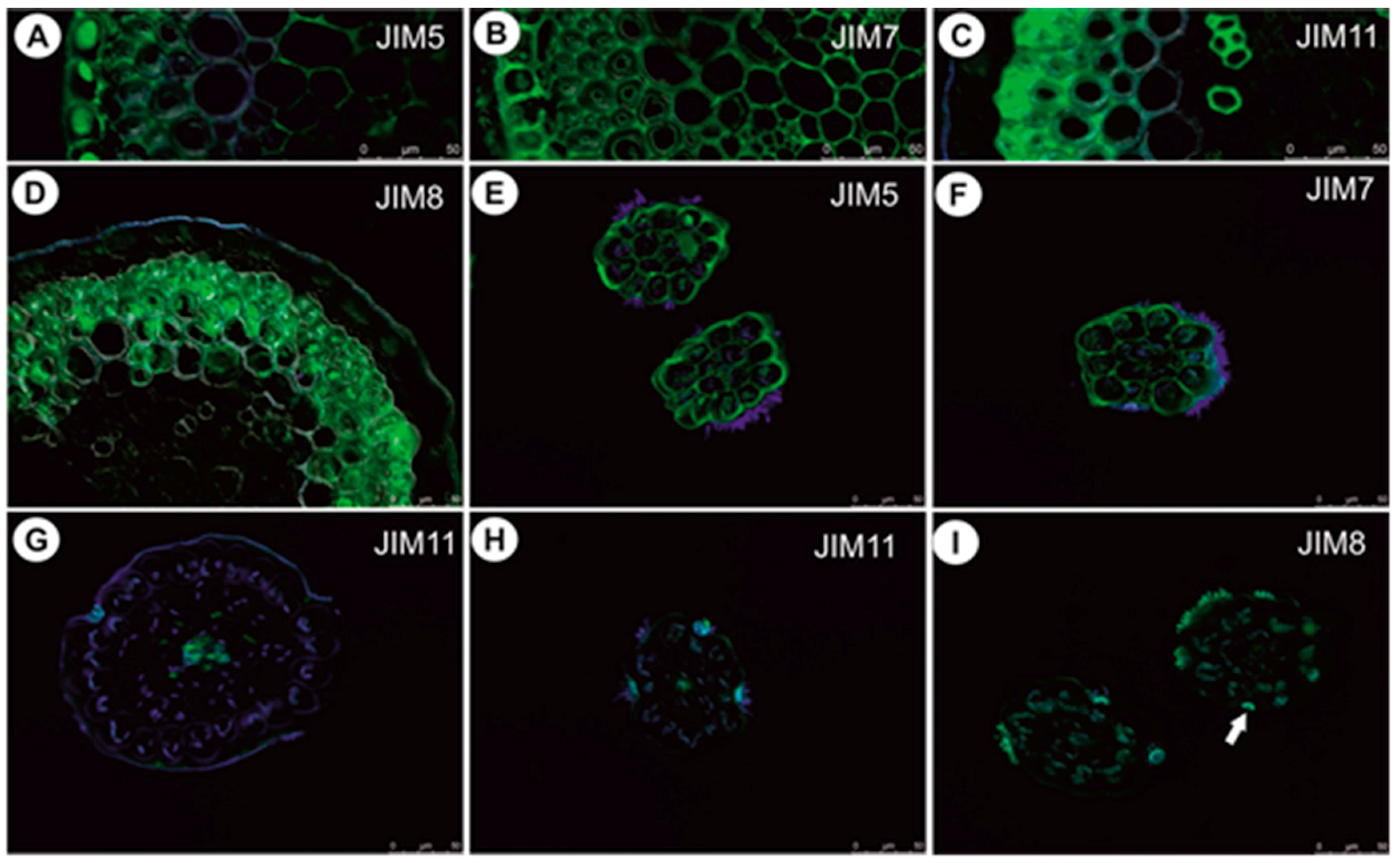
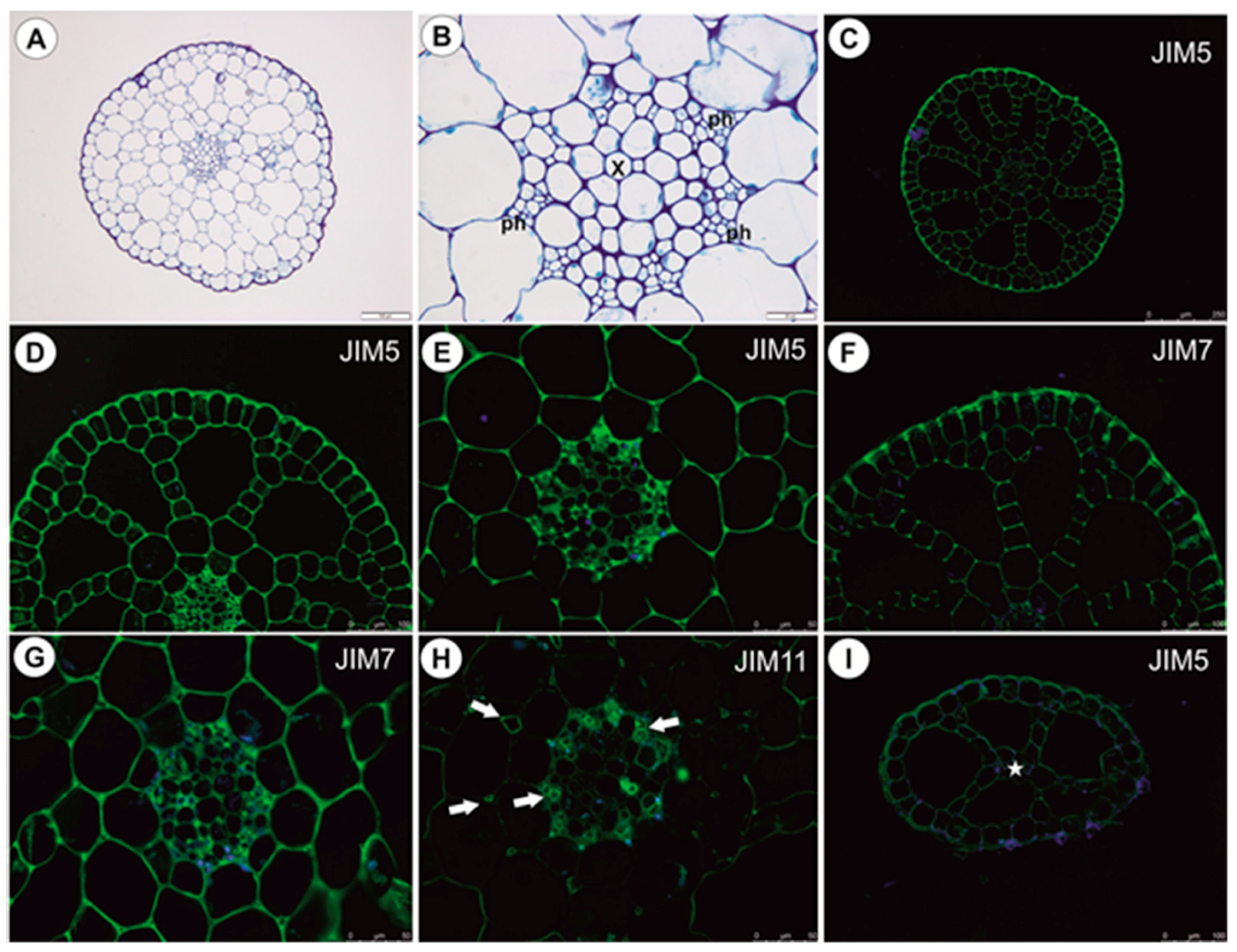
2.1.5. Immunodetection of Cell Wall Components
2.2. Utricularia reflexa
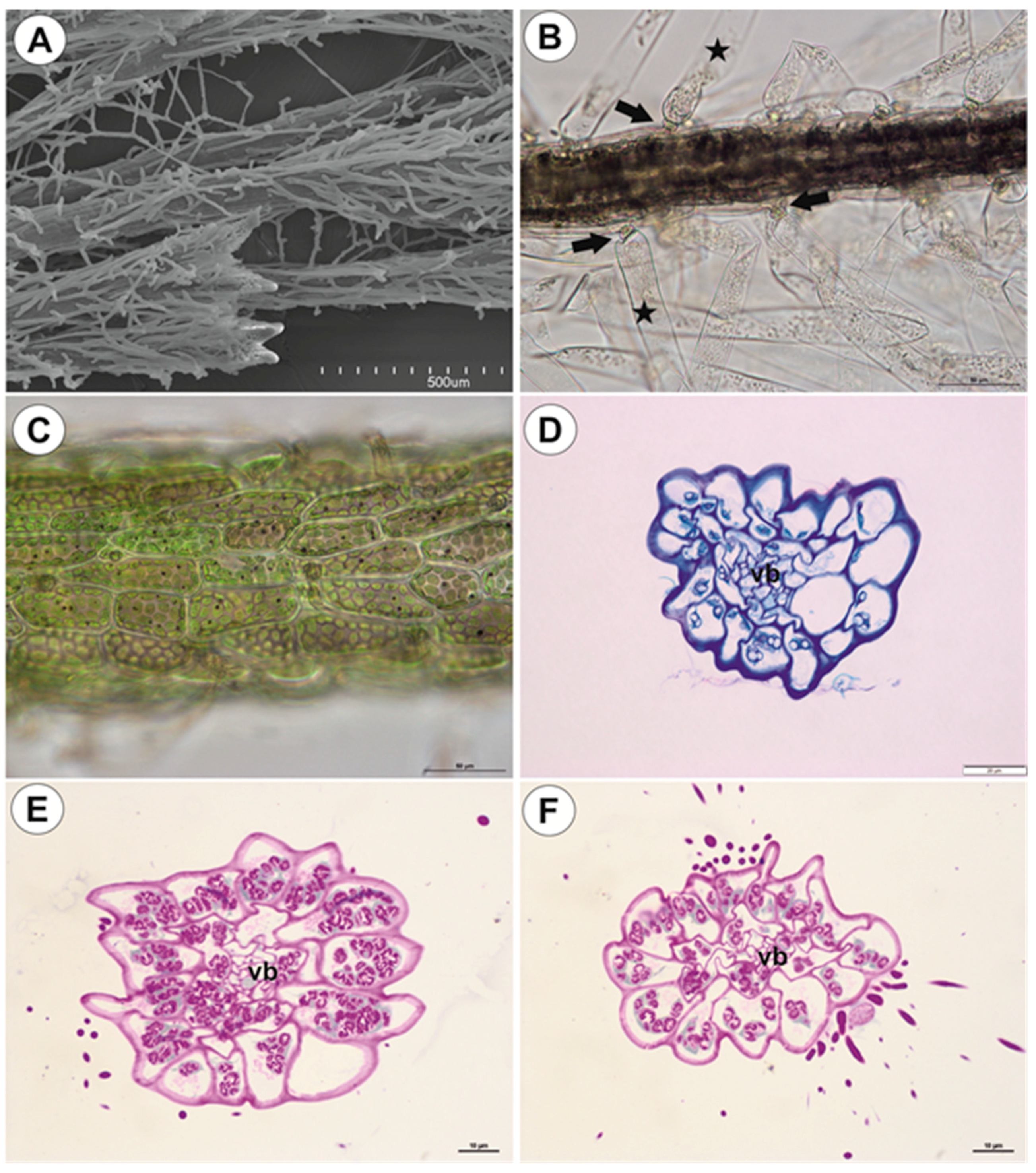
2.2.1. Shoots
2.2.2. Leaf-Like Shoots
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Morphological and Anatomical Studies
4.3. Immunodetection of Cell Wall Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taylor, P. The Genus Utricularia: A Taxonomic Monograph; Kew Botanic Gardens: London, UK, 1989. [Google Scholar]
- Miranda, V.F.O.; Menezes, C.G.; Silva, S.R.; Díaz, Y.C.A.; Rivadavia, F. Lentibulariaceae in Lista de Espécies da Flora do Brasil; Jardim Botânico do Rio de Janeiro: Rio de Janeiro, Brasil, 2015. Available online: http://floradobrasil.jbrj.gov.br/jabot/floradobrasil/FB30235 (accessed on 5 April 2020).
- Flora do Brasil 2020 under Construction. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/ (accessed on 30 April 2020).
- Adamec, L.; Fleischmann, A.; Pásek, K. Biology of the trapless rheophytic Utricularia neottioides: Is it possible to grow this specialized species in cultivation? Carniv. Plant Newslett. (Fullerton) 2015, 44, 104–114. [Google Scholar]
- Jobson, R.W.; Baleeiro, P.C.; Guisande, C. Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 89–104. [Google Scholar]
- Rutishauser, R.; Isler, B. Developmental genetics and morphological evolution of flowering plants, especially bladderworts (Utricularia): Fuzzy Arberian morphology complements classical morphology. Ann. Bot. 2001, 88, 1173–1202. [Google Scholar] [CrossRef] [Green Version]
- Van Steenis, C.G.G.J. Rheophytes of the World. An Account of the Flood-Resistant Flowering Plants and Ferns and the Theory of Autonomous Evolution; with 23 Photographs, 44 Figs.; Sijthoff & Noordhoff: Alphen aan den Rijn, The Netherlands; Rockville, MA, USA, 1981; 408p. [Google Scholar]
- Müller, K.; Borsch, T. Phylogenetics of Utricularia (Lentibulariaceae) and molecular evolution of the trnK intron in a lineage with high substitutional rates. Plant Syst. Evol. 2005, 250, 39–67. [Google Scholar] [CrossRef]
- Silva, S.R.; Gibson, R.; Adamec, L.; Domínguez, Y.; Miranda, V.F.O. Molecular phylogeny of bladderworts: A wide approach of Utricularia (Lentibulariaceae) species relationships based on six plastidial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2018, 118, 244–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satter, R.; Rutishauser, R. Structural and dynamic descriptions of the development of Utricularia foliosa and U. australis. Can. J. Bot. 1990, 68, 1989–2003. [Google Scholar] [CrossRef]
- Adamec, L. Ecophysiology of aquatic carnivorous plants. In Carnivorous Plants: Physiology, Ecology, and Evolution; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 256–269. [Google Scholar]
- Adamec, L. Biological flora of Central Europe: Utricularia intermedia Hayne, U. ochroleuca R.W. Hartm., U. stygia Thor and U. bremii Heer ex Kölliker. Perspect. Plant Ecol. Evol. Syst. 2020, 44, e125520. [Google Scholar] [CrossRef]
- Guisande, C.; Granado-Lorencio, C.; Andrade-Sossa, C.; Duque, S.R. Bladderworts. Funct. Plant Sci. Biotechnol. 2007, 1, 58–68. [Google Scholar]
- Rivadavia, F. Utricularia neottioides. Carniv. Plant Newslett. 1996, 25, 46–47. [Google Scholar]
- Irina, S.; Constantin, T. Morpho-structural adaptations of some aquatic carnivorous plant species (Aldrovanda vesiculosa L. and Utricularia vulgaris L.). Buletinul Grădinii Botanice Iaşi. 2007, 14, 7–12. [Google Scholar]
- Schweingruber, F.; Kučerová, A.; Adamec, L.; Doležal, J. Anatomic Atlas of Aquatic and Wetland Plant Stems; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Reut, M.S.; Świątek, P.; Miranda, V.F.O.; Płachno, B.J. Living between land and water—Structural and functional adaptations in vegetative organs of bladderworts. 2020; submitted. [Google Scholar]
- Von Luetzelburg, P. Beiträge zur Kenntnis der Utricularien. Flora 1910, 100, 145–212. [Google Scholar] [CrossRef]
- Evert, R.F.; Eichhorn, S.E. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; John Wiley &Sons: Hoboken, NJ, USA, 2006; 601p. [Google Scholar]
- Cook, C.D.K.; Rutishauser, R. Podostemaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 2007; Volume 9, pp. 304–344. [Google Scholar]
- Grob, V.; Pfeifer, E.; Rutishauser, R. Morphology, development and regeneration of Thelethylax minutiflora, a Madagascan river-weed (Podostemaceae). Phyton (Horn, Austria) 2007, 47, 205–229. [Google Scholar]
- Lloyd, F.E. The Carnivorous Plants; Chronica Botanica Company: Waltham, MA, USA, 1942. [Google Scholar]
- Rutishauser, R. Evolution of unusual morphologies in Lentibulariaceae (bladderworts and allies) and Podostemaceae (river-weeds): A pictorial report at the interface of developmental biology and morphological diversification. Ann. Bot. 2016, 117, 811–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameka, K.G.; Pfeifer, E.; Rutishauser, R. Developmental morphology of Saxicolella amicorum and S. submersa (Podostemaceae: Podostemoideae) from Ghana. Bot. J. Linn. Soc. 2002, 139, 255–273. [Google Scholar] [CrossRef]
- Jäger-Zürn, I.; Grubert, M. Podostemaceae depend on sticky biofilms with respect to attachment to rocks in waterfalls. Int. J. Plant Sci. 2000, 161, 599–607. [Google Scholar] [CrossRef]
- Heide-Jørgensen, H. Parasitic Flowering Plants; Koninklijke Brill NV: Leiden, The Netherlands, 2008. [Google Scholar]
- Seilacher, A.; MacClintock, C. Crinoid anchoring strategies for soft-bottom dwelling. Palaios 2005, 20, 224–240. [Google Scholar] [CrossRef]
- Plotnick, R.E.; Ebey, C.M.; Zinga, A. A radicle solution: Morphology and biomechanics of the Eucalyptocrinites “root” system. Lethaia 2016, 49, 130–144. [Google Scholar] [CrossRef]
- Ferretti, A.; Ausich, W.; Corradini, C.; Corriga, M.; Schönlaub, H. Stars in the Silurian sky: Echinoderm holdfasts from the Carnic Alps, Austria. Geol. Acta 2016, 14, 335–347. [Google Scholar]
- Slinger, J. The morphology and anatomy of Utricularia transrugosa Stapf. Bothalia 1954, 6, 385–406. [Google Scholar] [CrossRef]
- Rutishauser, R. The developmental plasticity of Utricularia aurea (Lentibulariaceae) and its floats. Aquat. Bot. 1993, 45, 119–143. [Google Scholar] [CrossRef]
- Chormanski, T.A. Morphology and anatomy of three common Everglade Utricularia species; U. gibba, U. cornuta, and U. subulata. Master’s Thesis, Florida International University, Miami, FL, USA, 2007. [Google Scholar]
- Brugger, J.; Rutishauser, R. Bau und Entwicklung landbewohnender Utricularia-Arten. Bot. Helv. 1989, 99, 91–146. [Google Scholar]
- Reut, M.S.; Płachno, B.J. Unusual developmental morphology and anatomy of vegetative organs in Utricularia dichotoma—leaf, shoot and root dynamics. Protoplasma 2020, 257, 371–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Płachno, B.J.; Adamec, L.; Kozieradzka-Kiszkurno, M.; Świątek, P.; Kamińska, I. Cytochemical and ultrastructural aspects of aquatic carnivorous plant turions. Protoplasma 2014, 251, 1449–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischmann, A. Do we have any evidence that any plants have given up carnivory? Carniv. Plant Newslett. 2011, 40, 37. [Google Scholar]
- Lustofin, K.; Świątek, P.; Miranda, V.F.O.; Płachno, B.J. Flower nectar trichome structure of carnivorous plants from the genus butterworts Pinguicula L. (Lentibulariaceae). Protoplasma 2020, 257, 245–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Co.: New York, NY, USA, 1940. [Google Scholar]
- Braune, W.; Leman, A.; Taubert, H. Praktikum Anatomii Roślin; Państwowe Wydawnictwo Naukowe: Warszawa, Polska, 1975; p. 295. [Google Scholar]
- Slazak, B.; Kapusta, M.; Malik SBohdanowicz, J.; Kuta, E.; Malec, P.; Göransson, U. Immunolocalization of cyclotides in plant cells, tissues and organ supports their role in host defense. Planta 2016, 244, 1029–1040. [Google Scholar] [CrossRef]
- Gawecki, R.; Sala, K.; Kurczyńska EU Świątek, P.; Płachno, B.J. Immunodetection of some pectic, arabinogalactan proteins and hemicellulose epitopes in the micropylar transmitting tissue of apomictic dandelions (Taraxacum, Asteraceae, Lactuceae). Protoplasma 2017, 254, 657–668. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH Range (n = 3) | Temp (°C) | Date-Time |
|---|---|---|
| 4.99–5.02 | 29.8 | 07 February 2019–15:15 |
| 6.22–6.36 | 27.1 | 07 February 2019–17:52 |
| 5.66–5.75 | 26.8 | 09 February 2019–14:44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płachno, B.J.; Adamec, L.; Świątek, P.; Kapusta, M.; Miranda, V.F.O. Life in the Current: Anatomy and Morphology of Utricularia neottioides. Int. J. Mol. Sci. 2020, 21, 4474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124474
Płachno BJ, Adamec L, Świątek P, Kapusta M, Miranda VFO. Life in the Current: Anatomy and Morphology of Utricularia neottioides. International Journal of Molecular Sciences. 2020; 21(12):4474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124474
Chicago/Turabian StylePłachno, Bartosz J., Lubomír Adamec, Piotr Świątek, Małgorzata Kapusta, and Vitor F. O. Miranda. 2020. "Life in the Current: Anatomy and Morphology of Utricularia neottioides" International Journal of Molecular Sciences 21, no. 12: 4474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124474