Changes in DNA Methylation and Gene Expression of Insulin and Obesity-Related Gene PIK3R1 after Roux-en-Y Gastric Bypass



 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Anthropometrics Data and Biochemical Profile
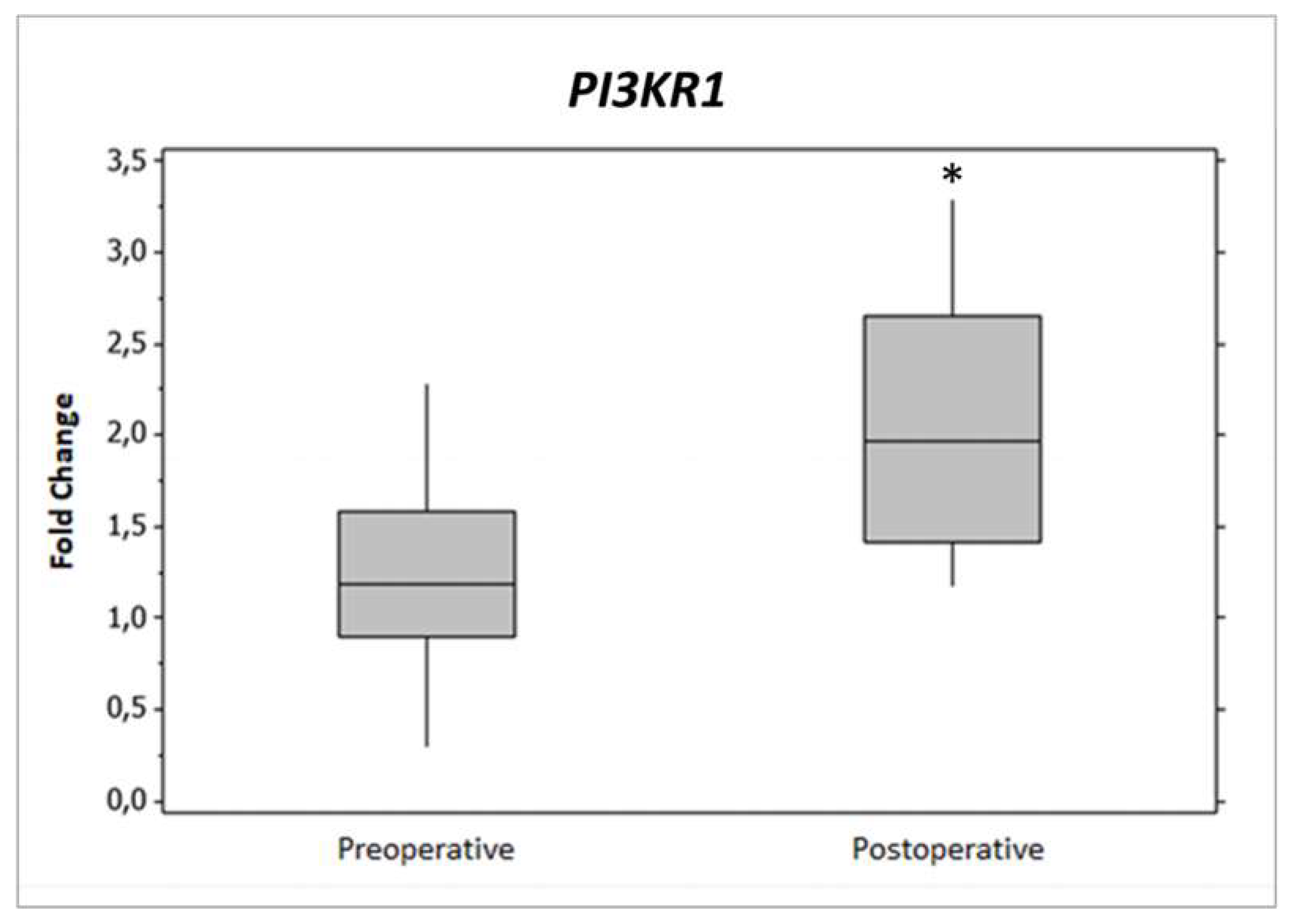
2.2. Gene Expression
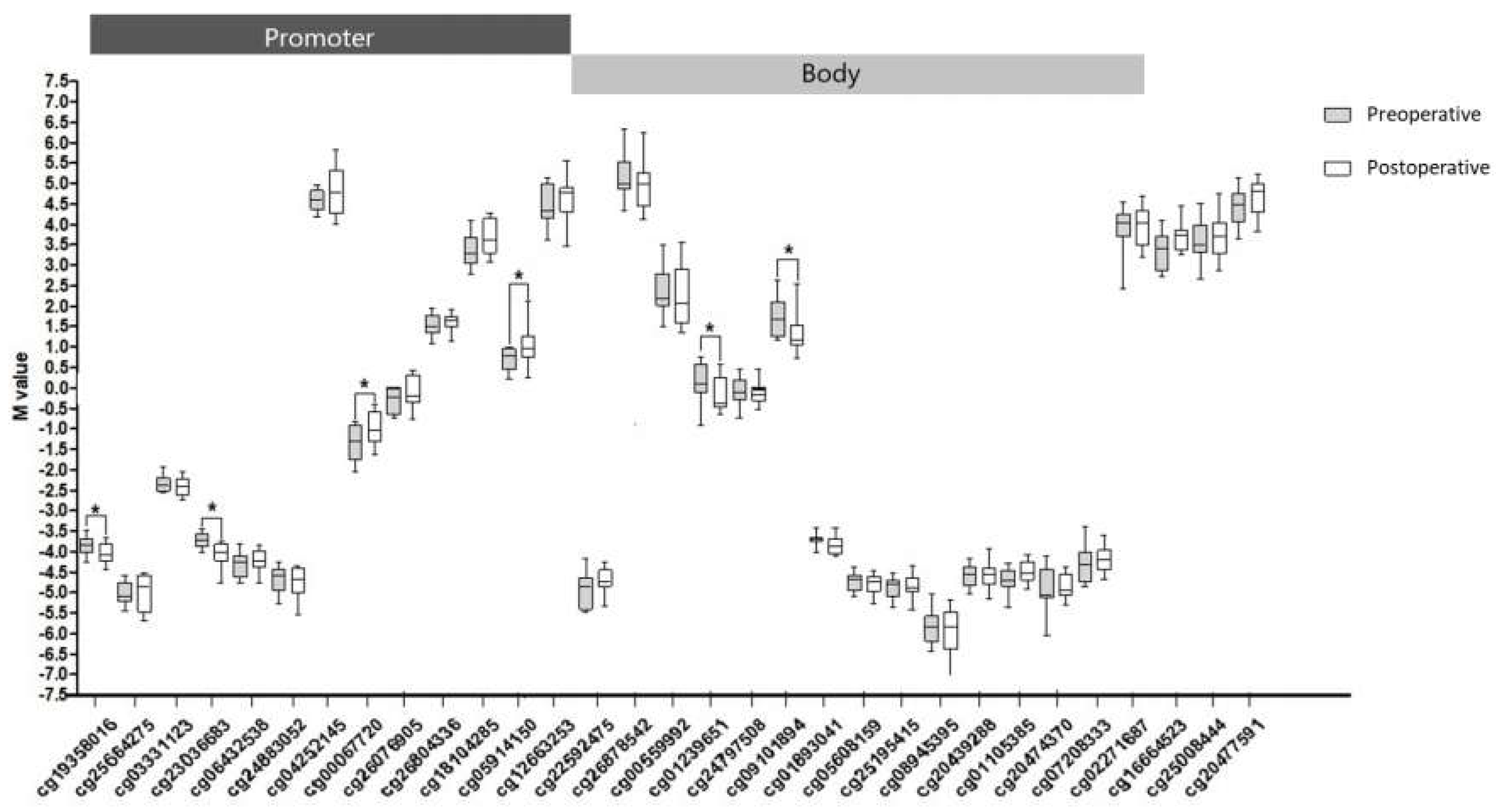
2.3. CpG Methylation of PIK3R1
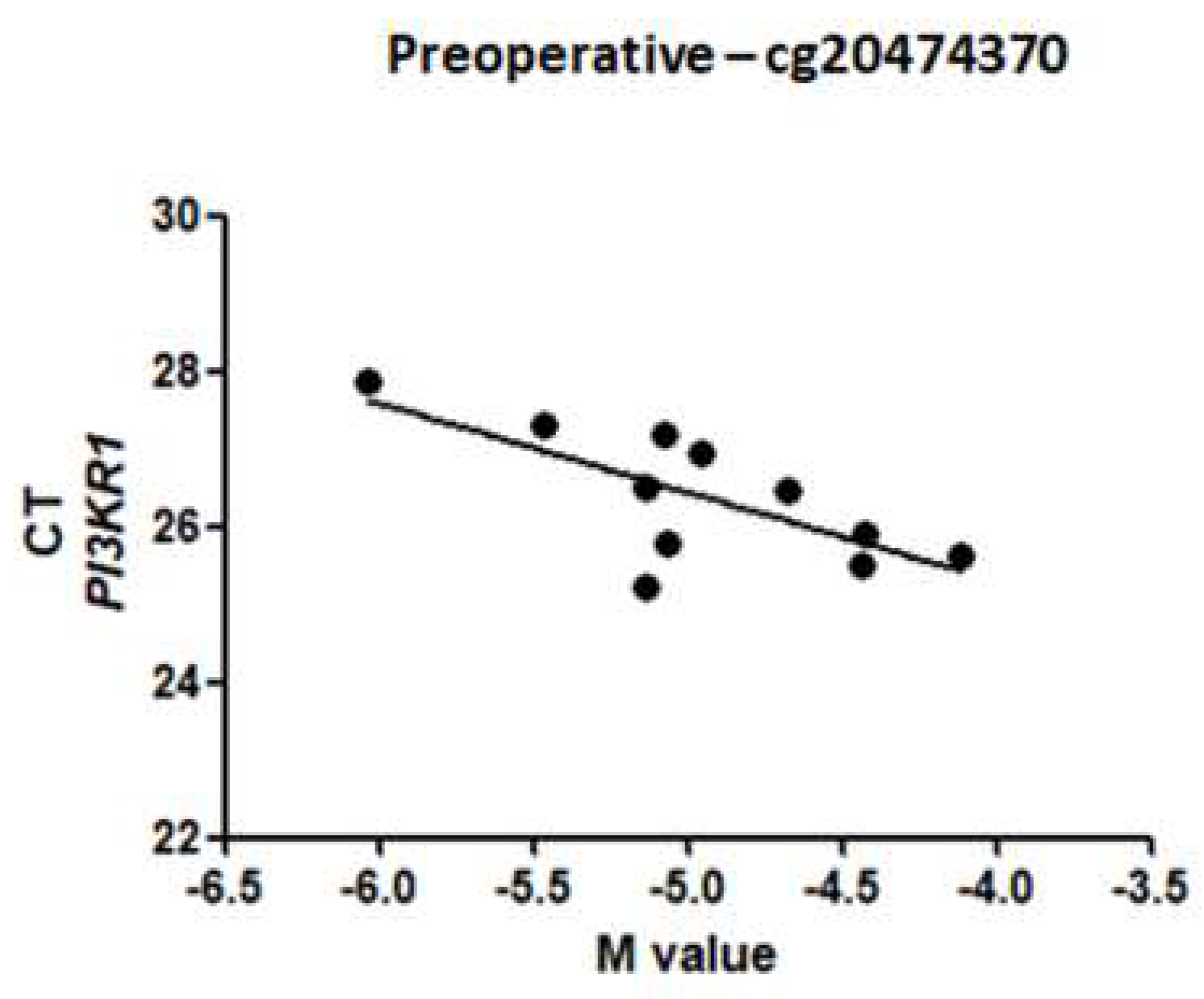
2.4. Relationship between CpG Methylation and PI3KR1 Gene Expression
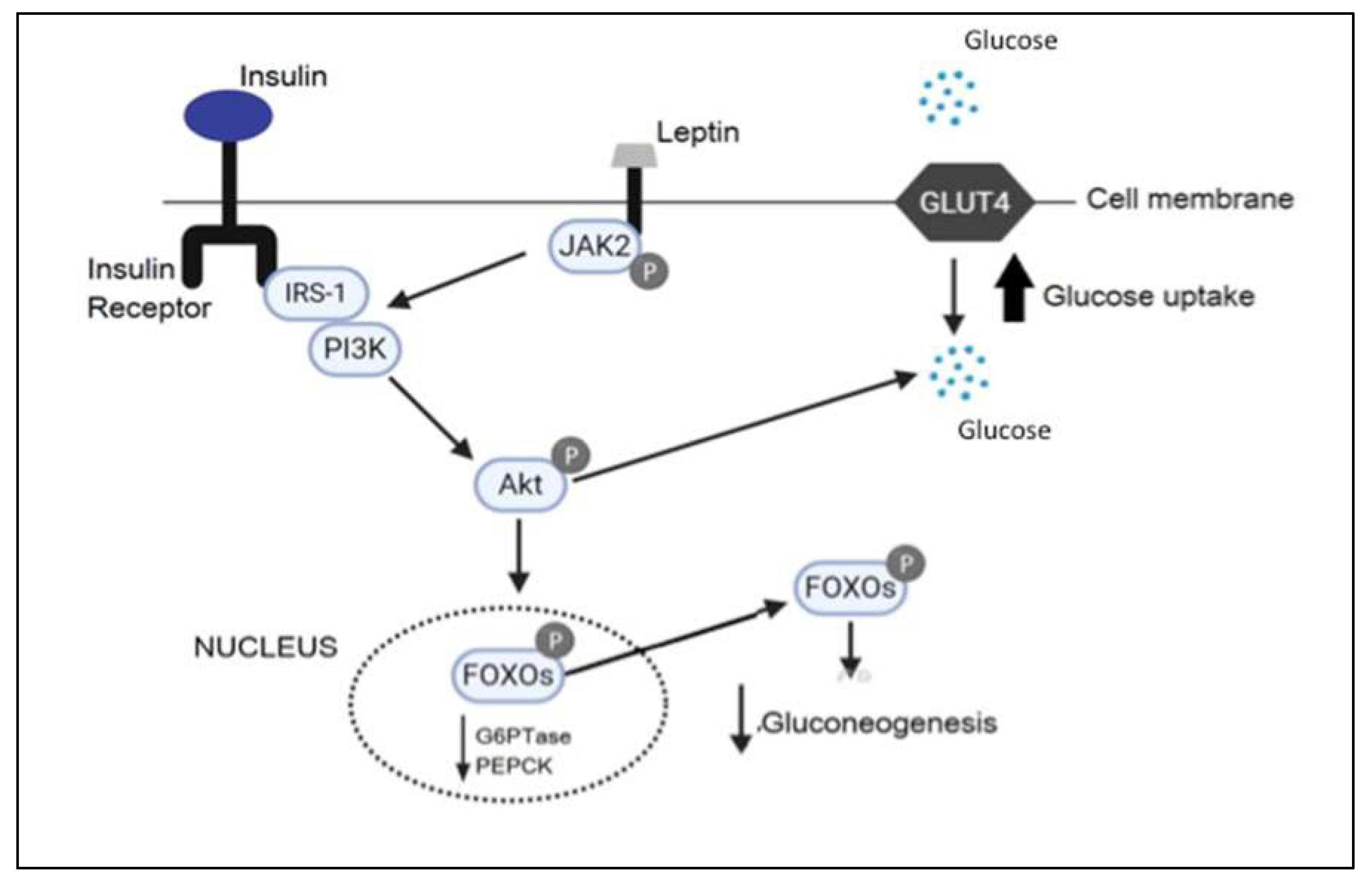
3. Discussion
4. Materials and Methods
4.1. RNA Isolation
4.2. Gene Expression Analysis
4.3. Validation Analysis
4.4. DNA Extraction
4.5. DNA Methylation Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RYGB | Roux-en-Y Gastric Bypass |
| CAAE | Certificate of Presentation for Ethical Consideration |
| BMI | Body Mass Index |
| RIN | RNA Integrity Number |
| FDR | False Discovery Rate |
| GO | Gene Ontology |
References
- Swinburn, B.; Egger, G.; Raza, F. Dissecting obesogenic environments: The development and application of a framework for identifying and prioritizing environmental interventions for obesity. Prev. Med. 1999, 29, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Corbin, L.J.; Timpson, N.J. Body mass index: Has epidemiology started to break down causal contributions to health and disease? Obesity 2016, 24, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llewellyn, C.; Wardle, J. Behavioral susceptibility to obesity: Gene environment interplay in the development of weight. Physiol. Behav. 2015, 152, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.R.; Lal, N.; Giridharan, N.V. Genetic & epigenetic approach to human obesity. Indian J. Med. Res. 2014, 140, 589–603. [Google Scholar]
- Xue, J.; Ideraabdullah, F.Y. An assessment of molecular pathways of obesity susceptible to nutrient, toxicant and genetically induced epigenetic perturbation. J. Nutr. Biochem. 2016, 30, 1–13. [Google Scholar] [CrossRef]
- De Lorenzo, A.; Soldati, L.; Sarlo, F.; Calvani, M.; Di Lorenzo, N.; Di Renzo, L. New obesity classification criteria as a tool for bariatric surgery indication. World J. Gastroenterol. 2016, 14, 681–703. [Google Scholar] [CrossRef]
- Guerron, D.A.; Portenier, D.D. Patient Selection and Surgical Management of High-Risk Patients with Morbid Obesity. Surg. Clin. N. Am. 2016, 96, 743–762. [Google Scholar] [CrossRef]
- Still, C.D.; Wood, G.C.; Chu, X.; Erdman, R.; Manney, C.H.; Benotti, P.N.; Petrick, A.T.; Strodel, W.E.; Mirshahi, U.L.; Mirshahi, T.; et al. High allelic burden of four obesity SNPs is associated with poorer weight loss outcomes following gastric bypass surgery. Obesity 2011, 19, 1676–1683. [Google Scholar] [CrossRef]
- González-Plaza, J.J.; Gutiérrez-Repiso, C.; García-Serrano, S.; Rodriguez-Pacheco, F.; Garrido-Sánchez, L.; Santiago-Fernández, C.; García-Arnés, J.; Moreno-Ruiz, F.J.; Rodríguez-Cañete, A.; García-Fuentes, E. Effect of Roux-en-Y gastric bypass-induced weight loss on the transcriptomic profiling of subcutaneous adipose tissue. Surg. Obes. Relat. Dis. 2016, 12, 257–263. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Nonino, C.B.; de Oliveira, B.A.; de Souza Pinhel, M.A.; Mansego, M.L.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. DNA Methylation and Hydroxymethylation Levels in Relation to Two Weight Loss Strategies: Energy-Restricted Diet or Bariatric Surgery. Obes. Surg. 2016, 26, 603–611. [Google Scholar] [CrossRef]
- Hochberg, Y.; Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Curdy, C.E.M.; Schenk, S.; Holliday, M.J.; Philp, A.; Julie AHouck Patsouris, D.; Majka, S.M. Attenuated Pik3r1 Expression Prevents Insulin Resistance and Adipose Tissue Macrophage Accumulation in Diet-Induced Obese Mice. Diabetes 2012, 61, 2495–2505. [Google Scholar]
- Hubaux, R.; Santos DD, B.; Enfield KS, S.; Rowbotham, D.; Lam, S.; Lam, W.L.; Martinez, V.D. Molecular features in arsenic-induced lung tumors. Rev. Mol. Cancer 2013, 12, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza Pinhel, M.A.; Noronha, N.Y.; Nicoletti, C.F.; de Oliveira, B.A.; Cortes-Oliveira, C.; Pinhanelli, V.C.; Junior, W.S.; Machry, A.J.; da Silva Junior, W.A.; Souza, D.R.; et al. Changes in Global Transcriptional Profiling of Women Following Obesity Surgery Bypass. Obes. Surg. 2018, 28, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Komaki, S.; Shiwa, Y.; Furukawa, R.; Hachiya, T.; Ohmomo, H.; Otomo, R.; Satoh, M.; Hitomi, J.; Sobue, K.; Sasaki, M.; et al. iMETHYL: An integrative database of human DNA methylation, gene expression, and genomic variation. Off. J. Jpn. Soc. Hum. Genet. 2018, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Berisha, S.Z.; Serre, D.; Schauer, P.; Kashyap, S.R.; Smith, J.D. Changes in whole blood gene expression in obese subjects with type 2 diabetes following bariatric surgery: A pilot study. PLoS ONE 2011, 6, e16729. [Google Scholar] [CrossRef] [Green Version]
- Doria, A.; Patti, M.E.; Kahn, C.R. The emerging genetic architecture of type 2 diabetes. Cell Metab. 2008, 8, 186–200. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.; Kim, K.W.; Kim, M.S. Leptin signalling pathways in hypothalamic neurons. Cell Mol. Life Sci. 2016, 73, 1457–1477. [Google Scholar] [CrossRef]
- Hill, J.W.; Xu, Y.; Preitner, F.; Fukuda, M.; Cho, Y.R.; Luo, J.; Balthasar, N.; Coppari, R.; Cantley, L.C.; Kahn, B.B.; et al. Phosphatidyl inositol 3-kinase signaling in hypothalamic proopiomelanocortin neurons contributes to the regulation of glucose homeostasis. Endocrinology 2009, 150, 4874–4882. [Google Scholar] [CrossRef]
- Williams, K.W.; Margatho, L.O.; Lee, C.E.; Choi, M.; Lee, S.; Scott, M.M.; Elias, C.F.; Elmquist, J.K. Segregation of acute leptin and insulin effects in distinct populations of arcuate proopiomelanocortin neurons. J. Neurosci. 2010, 30, 2472–2479. [Google Scholar] [CrossRef] [Green Version]
- Wellen, K.E.; Hotamisligil, G.S. Inflammation, stress, and diabetes. J. Clin. Investig. 2005, 115, 1111–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donato, J., Jr.; Frazão, R.; Elias, C.F. The PI3K signaling pathway mediates the biological effects of leptin. Arq. Bras. Endocrinol. Metabol. 2010, 54, 591–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, E.; Sayols, S.; Moran, S.; Guillaumet, A.A.; Schroeder, S.A.; Royo, R.; Orozco, M. A DNA methylation map of human cancer at single base-pair resolution. Oncogenomics 2017, 36, 5648–5657. [Google Scholar] [CrossRef] [PubMed]
- Nikki, A.; FordKaylyn, L.; Devlin, L.M. Deconvoluting the Obesity and Breast Cancer Link: Secretome, Soil and Seed Interactions. J. Mammary Gland Biol. Neoplasia 2013, 18, 267–275. [Google Scholar]
- Du, P.; Zhang, X.; Huang, C.C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-value and M-value methods for quantifying methylation levels by microarray analysis. BMC Bioinform. 2010, 11, 587. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T. Epigenetic Silencing Mediated Through Activated PI3K/AKT Signaling in Breast Cancer. Cancer Res. 2012, 5, 1752–1762. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.A. The DNA methylation paradox. Trends Genet. 1999, 15, 35–49. [Google Scholar] [CrossRef]
- Guénard, F. Methylation and Expression of Immune and Inflammatory Genes in the Offspring of Bariatric Bypass Surgery Patients. J. Obes. 2013, 2013, 2–9. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Pinhel, M.A.; Diaz-Lagares, A.; Casanueva, F.F.; Jácome, A.; Pinhanelli, V.C.; de Oliveira, B.A.; Crujeiras, A.B.; Nonino, C.B. DNA methylation screening after roux-en Y gastric bypass reveals the epigenetic signature stems from genes related to the surgery per se. BMC Med. Genom. 2019, 12, 72. [Google Scholar] [CrossRef]
- Tian, Y. ChAMP: Updated methylation analysis pipeline for Illumina BeadChips. Bioinformatics 2017, 15, 3982–3984. [Google Scholar] [CrossRef] [Green Version]
- Davé, V.; Yousefi, P.; Huen, K.; Volberg, V.; Holland, N. Relationship between expression and methylation of obesity-related genes in children. Mutagenesis 2015, 30, 411–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Preoperative | Postoperative |
|---|---|---|
| N | 13 | 13 |
| Age (years) | 35.9 ± 9.2 | 36.2 ± 9.3 |
| Weight (kg) | 115.3 ± 19.4 | 85.3 ± 13.8 a |
| BMI (kg/m²) | 42.5 ± 7.0 | 31.8 ± 5.4 a |
| TC (mg/dL) | 177.4 ± 32.7 | 152.5 ± 28.4 a |
| HDL-C (mg/dL) | 41.3 ± 6.0 | 44.3 ± 8.1 |
| LDL-C (mg/dL) | 112.4 ± 27.6 | 88.6 ± 23.0 a |
| TG (mg/dL) | 129.7 ± 42.7 | 75.0 ± 27.0 a |
| Glucose (mg/dL) | 93.3 ± 13.5 | 83.6 ± 28.4 a |
| Symbol | Gene Name | Fold Change * |
|---|---|---|
| Obesity | ||
| EPS8 | Epidermal growth factor receptor pathway substrate 8 | 5.57 × 106 |
| PTPN2 | Protein tyrosine phosphatase, non-receptor type 2 | 2.79 × 106 |
| ACTR3 | ARP3 actin-related protein 3 homolog | 2.32 × 106 |
| RPS6KA3 | Ribosomal protein S6 kinase | 2.45 × 106 |
| MAPK9 | Mitogen-activated protein kinase 9 | 2.26 × 106 |
| RAP1B | RAP1B member of RAS oncogene family | 5.51 × 106 |
| PLA2G4A | Phospholipase A2 | 9.65 × 106 |
| PIK3R1 | Phosphoinositide-3-kinase, regulatory subunit 1 | 6.55 × 106 |
| Insulin Pathway | ||
| EIF4E | Eukaryotic translation initiation factor 4E | 3.76 × 106 |
| IKBKB | Inhibitor of kappa light polypeptide gene enhancer in B-cells kinase beta | 3.90 × 106 |
| PHKB | Phosphorylase kinase beta | 4.13 × 106 |
| MAPK9 | Mitogen-activated protein kinase 9 | 2.26 × 106 |
| PPP1CB | Protein phosphatase 1, catalytic subunit, beta isozyme | 2.19 × 106 |
| SOCS4 | Suppressor of cytokine signaling 4 | 4.13 × 106 |
| PIK3R1 | Phosphoinositide-3-kinase, regulatory subunit 1 alpha | 6.55 × 106 |
| Gene Symbol (Gene ID) | Site | Gene Region | β | Adjusted Multiple r² | p-Value |
|---|---|---|---|---|---|
| PI3KR1 (ENSG00000145675) | cg20474370 | Body | −0.59 | 0.70 | 0.003 |
| Gene | ID (Gene Expression Assay) | Fragment (pb) |
|---|---|---|
| PIK3R1 | Hs00933163 | 82 |
| GAPDH | Hs99999905 | 122 |
| ACTINB | Hs99999903 | 171 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinhel, M.A.S.; Noronha, N.Y.; Nicoletti, C.F.; Pereira, V.A.; de Oliveira, B.A.; Cortes-Oliveira, C.; Salgado, W., Jr.; Barbosa, F., Jr.; Marchini, J.S.; Souza, D.R.; et al. Changes in DNA Methylation and Gene Expression of Insulin and Obesity-Related Gene PIK3R1 after Roux-en-Y Gastric Bypass. Int. J. Mol. Sci. 2020, 21, 4476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124476
Pinhel MAS, Noronha NY, Nicoletti CF, Pereira VA, de Oliveira BA, Cortes-Oliveira C, Salgado W Jr., Barbosa F Jr., Marchini JS, Souza DR, et al. Changes in DNA Methylation and Gene Expression of Insulin and Obesity-Related Gene PIK3R1 after Roux-en-Y Gastric Bypass. International Journal of Molecular Sciences. 2020; 21(12):4476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124476
Chicago/Turabian StylePinhel, Marcela A S, Natália Y Noronha, Carolina F Nicoletti, Vanessa AB Pereira, Bruno AP de Oliveira, Cristiana Cortes-Oliveira, Wilson Salgado, Jr., Fernando Barbosa, Jr., Júlio S Marchini, Doroteia RS Souza, and et al. 2020. "Changes in DNA Methylation and Gene Expression of Insulin and Obesity-Related Gene PIK3R1 after Roux-en-Y Gastric Bypass" International Journal of Molecular Sciences 21, no. 12: 4476. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124476