Lipid Signaling in Ocular Neovascularization


1
Department of Ophthalmology, Graduate School of Medicine, The University of Tokyo, Tokyo 113-0033, Japan
2
Department of Ophthalmology, Nagoya University Graduate School of Medicine, Nagoya 466-8550, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(13), 4758; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134758
Submission received: 17 June 2020
/
Revised: 2 July 2020
/
Accepted: 2 July 2020
/
Published: 4 July 2020
(This article belongs to the Special Issue Recent Advancements in Research and Therapy of Ocular Neovascular Diseases)
Abstract
:Vasculogenesis and angiogenesis play a crucial role in embryonic development. Pathological neovascularization in ocular tissues can lead to vision-threatening vascular diseases, including proliferative diabetic retinopathy, retinal vein occlusion, retinopathy of prematurity, choroidal neovascularization, and corneal neovascularization. Neovascularization involves various cellular processes and signaling pathways and is regulated by angiogenic factors such as vascular endothelial growth factor (VEGF) and hypoxia-inducible factor (HIF). Modulating these circuits may represent a promising strategy to treat ocular neovascular diseases. Lipid mediators derived from membrane lipids are abundantly present in most tissues and exert a wide range of biological functions by regulating various signaling pathways. In particular, glycerophospholipids, sphingolipids, and polyunsaturated fatty acids exert potent pro-angiogenic or anti-angiogenic effects, according to the findings of numerous preclinical and clinical studies. In this review, we summarize the current knowledge regarding the regulation of ocular neovascularization by lipid mediators and their metabolites. A better understanding of the effects of lipid signaling in neovascularization may provide novel therapeutic strategies to treat ocular neovascular diseases and other human disorders.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Living organisms are composed of various organic compounds, including proteins, carbohydrates, nucleic acids, and lipids. Lipids play crucial roles in numerous cellular processes, acting as cellular structural components and biological barriers and regulating numerous signaling pathways [1,2]. The biological membranes of eukaryotes are amphiphilic sheaths consisting of a lipid bilayer, which acts as a cell barrier [3]. The lipid backbones, head groups, chain length, and position and number of carbon double bonds in fatty acyl chains vary immensely, contributing to the diversity of membranes [4]. Glycerophospholipids, sphingolipids, and sterols are major components of membrane lipids [5]. Additionally, bioactive lipid mediators produced from membrane lipids play pivotal roles in various biological processes and have been implicated in numerous disorders [6,7,8,9,10]. Mounting evidence implies that lipid signaling plays a crucial role in angiogenesis [11,12].
Signaling in Ocular Neovascularization
Both vasculogenesis and angiogenesis are essential for the formation of vascular networks. Vasculogenesis is the process of blood vessel development by endothelial cells originating from mesoderm-derived progenitor cells [13]. Angiogenesis is the formation of new vessels in response to angiogenic factors; the new vessels are formed by sprouting of endothelial cells in pre-existing vessels [14]. Angiogenesis is profound during embryogenesis but also occurs in adult tissues under certain circumstances, such as wound healing and tumor development [15]. Both vasculogenesis and angiogenesis are involved in neovascularization [16].
Mature blood vessels usually remain quiescent, and angiogenesis does not occur under physiological conditions. However, several stimuli can trigger neovascularization, often leading to pathogenesis. For instance, chronic hypoxia induces the production of angiogenic factors in human tissues, promoting vascular growth from pre-existing vessels [17]. Angiogenesis occurs in several steps. Angiogenic factors trigger endothelial cell sprouting [18], followed by their proliferation and migration and subsequent vessel elongation [19]. A subset of activated endothelial cells become tip cells extending numerous filopodia; tip cells guide the vessel sprout toward angiogenic factors, such as vascular endothelial growth factor (VEGF) and delta-like ligand 4 (DLL4) [20]. Upon Notch signaling, endothelial cells adjacent to tip cells turn to stalk cells [21], which promote lumen formation. The new vessels maturate after the formation of the basement membrane by pericytes and vascular smooth muscle cells [22].
Although angiogenesis is regulated by numerous factors, including fibroblast growth factors (FGF), neuropilins, platelet-derived growth factors (PDGF), and angiopoietins, VEGF is widely considered a master regulator of the process. VEGF gradients in avascular areas guide tip cell migration and induce stalk cell formation, promoting the elongation of new vessels [23]. Vegf knockout (-/-) in mice is embryonically lethal around midgestation due to vascular defects [24]. VEGF binding to VEGF receptors (VEGFR1-3) regulates angiogenesis, vasculogenesis, and lymphangiogenesis [25]. Among these receptors, VEGFR1 and VEGFR2 are predominantly expressed in endothelial cells. VEGFR2, rather than VEGFR1, is involved in angiogenesis by regulating endothelial cell proliferation, migration, and viability [26]. Cellular responses to VEGF/VEGFR singling facilitate both physiological and pathological angiogenesis by modulating other bioactive molecules, including endothelial nitric oxide synthase (eNOS), mammalian target of rapamycin (mTOR), and Rho family guanosine triphosphatases (GTPases) [27,28]. In addition to VEGF, hypoxia-inducible factor (HIF) is another critical angiogenic factor. Under hypoxic conditions, HIF-1α translocates into the nucleus and dimerizes with HIF-1β binding to hypoxia response elements (HRE); HIF-1α/ HIF-1β dimers promote angiogenesis by inducing the expression of VEGF, glucose transporter 1 (GLUT1), erythropoietin, and inducible nitric oxide synthase (iNOS) [29]. Interleukin (IL)-8 is a pro-angiogenic cytokine; it activates various molecules, including HIF-1, NF-κB, and signal transducers and activator of transcription 3 (STAT3) via CXC chemokine receptor 1 and 2 [30]. Furthermore, IL-8 transactivates VEGFR2 in endothelial cells, enhancing endothelial permeability and promoting angiogenesis [31]. Vascular endothelial (VE)-cadherin is a component of endothelial junctions, which has been shown to drive angiogenesis. VE-cadherin phosphorylation by VEGF induces endothelial junction disruption and internalization, promoting endothelial cell migration [32].
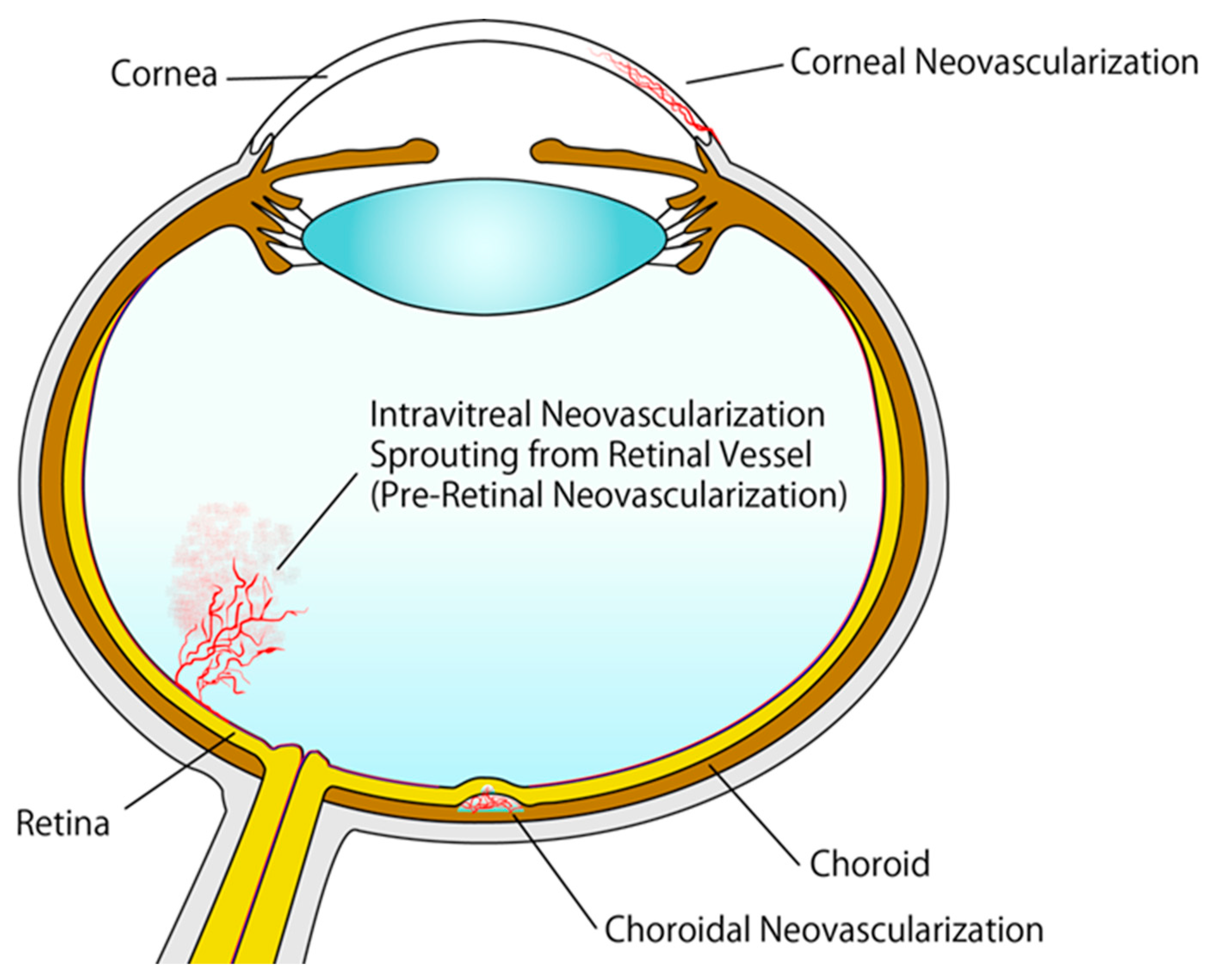
The importance of angiogenesis is reflected in the fact that defects in the process have been implicated in numerous disorders, including cardiovascular diseases and cancer, potentially leading to death [33]. Ocular neovascularization is one of the conditions caused by aberrant angiogenesis. Ocular neovascularization is characterized by pathological neovascularization in the retina, choroid, iris, and cornea (Figure 1) [34]. It is observed in individuals with retinal vein occlusion (RVO), retinopathy of prematurity (ROP), diabetic retinopathy, age-related macular degeneration (AMD), neovascular glaucoma, and corneal neovascularization induced by trauma or inflammation [35].
Retinal neovascularization and choroidal neovascularization (CNV) are the most common vascular diseases of the eye. Retinal neovascularization diseases, such as RVO, ROP, and diabetic retinopathy, are primarily caused by chronic or severe retinal ischemia [36]. Retinal ischemia induces the production of intravitreal angiogenic factors [37], leading to the proliferation of retinal blood vessels. Pre-retinal neovascularization in the vitreous cavity can cause tractional retinal detachment and vitreous hemorrhage due to increased vascular vulnerability (Figure 2). In severe retinopathy cases, pre-retinal neovascularization can lead to vision loss. Considering its importance in angiogenesis, VEGF is a promising therapeutic target for pathologic retinal neovascularization. Hence, intravitreal injection of VEGF-targeting agents is currently a standard treatment option for patients with visual impairments due to pathologic retinal neovascularization [38,39].
AMD is another common condition that can cause visual impairments. It is more prevalent in older people and in developed countries. In AMD, lesions often develop within the macula, the central area of the retina responsible for high-acuity vision. Early-stage AMD manifests as abnormalities in retinal pigment epithelial (RPE) and drusen deposits without impaired visual acuity [40]. Lipoproteins and cholesterol represent major components of soft drusen deposits. Late-stage AMD manifests as exudative AMD or geographic atrophy. CNV is a hallmark of exudative AMD, causing subretinal hemorrhage and vascular leakage (Figure 1). Currently, intravitreal administration of VEGF-targeting agents is the most common treatment for exudative AMD [41,42,43].
The cornea is characterized by complete avascularity under physiological conditions to maintain the clarity required for visual acuity [44]. Under pathological conditions, including corneal infection, traumatic injury, ocular surface inflammation, and limbal stem cell deficiency, corneal neovascularization and sprouting from the pericorneal plexus can occur [45,46,47], potentially leading to vision loss. The new vessels can induce corneal opacification and impair vision. Patients with severe corneal neovascularization may require corneal transplantation. Various corneal neovascularization animal models have been established to investigate disease pathology and test novel therapeutics for artificial general neovascularization as well as corneal neovascularization [48]; in most of these models, corneal neovascularization is induced by suture, chemical treatments, and surgical implantation of hydron pellets with reagents containing angiogenic factors [49].
Although numerous efforts have been made to elucidate the mechanisms underlying ocular neovascularization diseases, the role of bioactive lipids remains elusive. In this review, we summarize current knowledge regarding the role of lipid signaling in ocular neovascularization.
2. Glycerophospholipids in Ocular Neovascularization
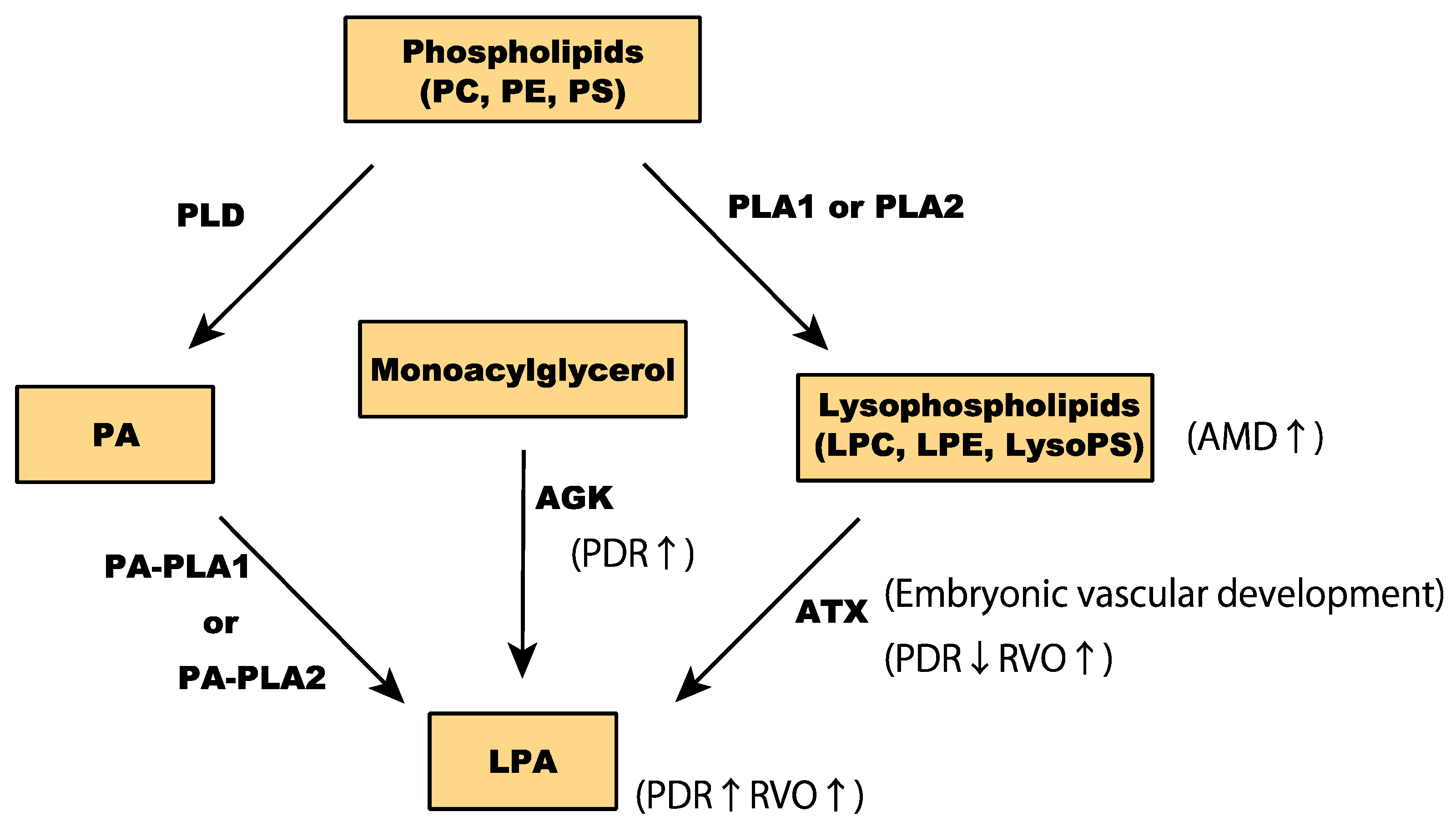
Glycerophospholipids are the most prevalent membrane lipid in mammalian cells. They are composed of a glycerol backbone, two long-chain fatty acids at sn-1 and sn-2 positions of glycerol, and phosphoric acid esterified at sn-3 as a headgroup [50]. The different combinations of headgroups and fatty acids allows for over a thousand glycerophospholipid variants in mammalian cells. Based on their polar headgroups, glycerophospholipids can be classified into phosphatidic acids (PA), phosphatidylcholine (PC), phosphatidylethanolamines (PE), phosphatidylserines (PS), phosphatidylglycerol, and phosphatidylinositol (Figure 3) [51].
Lysophospholipids are synthesized from glycerophospholipids by phospholipase A (PLA). PLA1 or PLA2 enzymatically remove an acyl chain at sn-1 or sn-2 [52]. Among lysophospholipids, lysophosphatidylcholine (LPC) and lysophosphatidylethanolamine (LPE) are the most abundant in mammalian cells [53]. Other lysophospholipids include lysophosphatidic acid (LPA), lysophosphatidylserine (LysoPS), platelet-activating factor, and 2-arachidonoylglycerol, all of which are involved in various cellular responses by acting as signaling molecules [54]. Despite the low intracellular levels of LPA and LysoPS, they regulate important processes, including angiogenesis.
2.1. Lysophosphatidic Acid (LPA)
Mounting evidence implies that LPA plays a crucial role in the pathogenesis of ocular neovascularization. Intracellular LPA is synthesized by glycerophosphate acyltransferase, PA-specific PLA (PA-PLA), and acylglycerol kinase (AGK) [55,56,57], whereas extracellular lysophospholipids are converted to LPA by lysophospholipase D (also known as autotaxin; ATX) [58,59]. LPA is predominantly found in the serum and plasma [60,61].
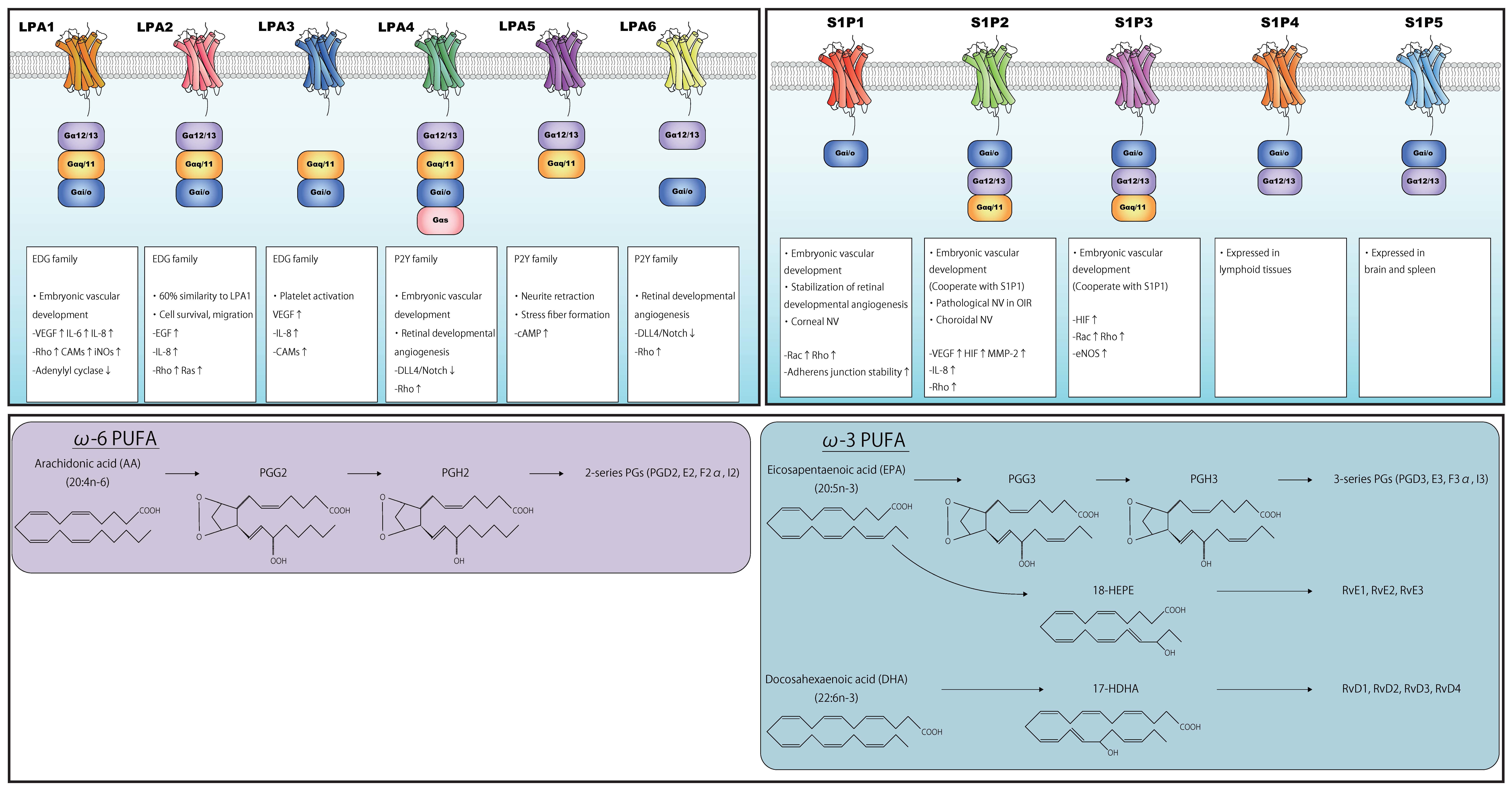
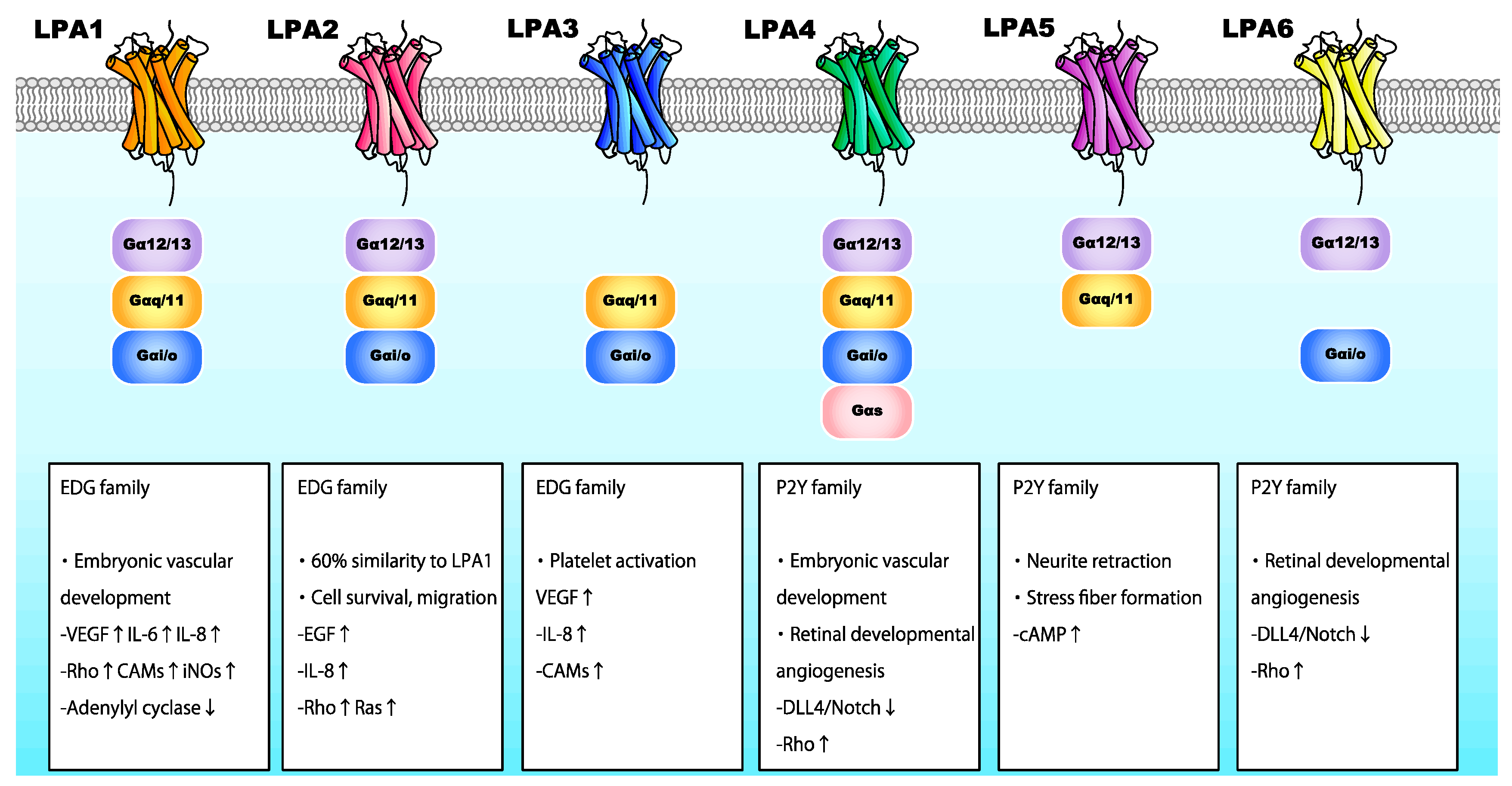
LPA is involved in various processes, including angiogenesis, cell proliferation, neurite retraction, and stress fiber formation via LPA receptors [58,62,63,64]. LPA interacts with specific G-protein-coupled receptors found on the cell surface. Six LPA receptors (LPA1-6) have been identified thus far (Figure 4) [65]. LPA1–3 belong to the endothelial differentiation gene family, whereas LPA4–6 belong to the P2Y receptor family [66]. LPA receptors interact with and activate heterotrimeric Gα proteins, namely Gαq/11, Gα12/13, Gαi/o, and Gαs, thereby regulating various cellular functions (Figure 4). Activated G proteins subsequently regulate the activation status of numerous intracellular signaling molecules, including Rho GTPase, mitogen-activated protein kinases (MAPK), protein kinase B (Akt), and phosphoinositide 3-kinase (PI3K) [67].
Numerous human gene analysis and preclinical studies have investigated the roles of LPA and LPA receptors [68,69]. These studies have shown that LPA is essential for vascularization [70]. Notably, Atx−/− mice were not viable after E10.5 due to severe vascular defects resembling those in Vegf−/− mice [71]. Atx deletion disrupts the development of vascular networks in the embryo and yolk sac. Allantois explants from Atx−/− mice formed vessels, although the lack of ATX and LPA caused endothelial disassembly. These findings imply that the ATX-LPA axis plays a critical role in vascular formation by stabilizing the immature vascular network. Among the LPA receptors, LPA-Gα12/Gα13 is reportedly responsible for this process, as a deficiency thereof results in a similar phenotype to that of Atx−/− mice [72]. A previous study using zebrafish revealed the requirement of ATX for vascular development, and that suppression of LPA-LPA1 and/or LPA-LPA4, both of which interact with Gα12/Gα13, was responsible for these vascular defects [73]. Similarly, in the absence of Rho-associated protein kinases (ROCK-1/2), which act downstream of LPA, mice exhibited impaired vascular remodeling in the yolk sac [74]. These findings highlight the vital role of LPA/LPA1 and LPA/LPA4 in vascular development.
In normal human cells, LPA facilitates angiogenesis by enhancing the production of VEGF, IL-8, MCP-1, and MMP-9 [75,76] in a Gαi/NF-κB-dependent manner, while VEGF upregulation requires the binding of LPA on LPA1 and LPA3 [77]. Moreover, LPA induces HIF-1α expression and nuclear translocation in various cancer cells by activating PI3K/Akt/mTOR and p42/p44 MAPK signaling pathways [78]. Of note, hypoxia or HIF-1α activation enhances LPA-driven cellular responses [79]. Additionally, LPA induces the expression of various cytokines. Notably, LPA promoted IL-6 expression by binding to LPA1 [80], whereas binding to LPA1, LPA2, or LPA3 induced IL-8 expression [81]. The LPA-mediated upregulation of these cytokines was dependent on NF-κB activation by PI3K/Akt and protein kinase C (PKC) signaling pathways. LPA/LPA1 and LPA/LPA3 signaling also upregulated the expression of numerous cell adhesion molecules [82]. LPA binding to LPA1 activated Rho and iNOS in vivo [83,84]. Therefore, it has become evident that LPA regulates angiogenesis in ocular tissues by activating the expression of various angiogenic factors. Nevertheless, the relevance of each LPA receptor in angiogenesis in different tissues remains unclear.
2.2. Lysophosphatidic Acid and Ocular Neovascularization
The biological roles of ATX and LPA in ocular tissues have been partly characterized. ATX is highly expressed in retina [85], and the ATX-encoding gene (ENPP2) has been identified as an RPE signature gene [86]. LPA is required for retinogenesis, promoting retinal growth cone collapse [87,88]. In human RPE cells, LPA was shown to regulate barrier integrity [89]. These findings imply involvement of the ATX/LPA axis in retinal function. A recent study assessing the role of LPA and its receptors in angiogenesis during retinal vascular development demonstrated that endothelial cells of mice deficient in LPA4 and LPA6 exhibited reduced filopodia and vessel sprouting, with a reduced number of blood vessels. LPA4/LPA6 coupling with Gα12/Gα13 was also shown to promote retinal angiogenesis, as well as the global vascularization described above, in mice by activating the Gα12/Gα13-Rho-ROCK axis in endothelial cells [90]. Gα12/Gα13-Rho-ROCK pathway activation inhibited DLL4/Notch signaling by activating Yes-associated protein (YAP) and transcriptional co-activator with PDZ-binding motif (TAZ) in endothelial cells at the vascular front. These findings imply that LPA4 and/or LPA6 facilitate tip cell formation and stalk cell proliferation in the retina, promoting angiogenesis (Figure 4).
Several clinical studies have also investigated the role of LPA in pathological ocular neovascularization. Intravitreal LPA concentrations were significantly elevated in patients with PDR (proliferative diabetic retinopathy) [91]. Intravitreal concentrations of LPA and PA were significantly associated with levels of the inflammatory biomarker vascular cell adhesion molecule-1 (VCAM-1). AGK levels were also higher in PDR patients, implying involvement of the LPA signaling pathway in the pathology of diabetic retinopathy (Figure 3). A subsequent study confirmed the elevated levels of LPA and AGK in the vitreous fluid of PDR patients. On the other hand, ATX levels were significantly lower in PDR patients than in patients without diabetes, implying that AGK-LPA rather than ATX-LPA may be involved in the development and progression of PDR [92]. According to a study investigating LPA expression in RVO, levels of LPA and ATX were significantly higher in vitreous samples from patients with RVO than in samples from patients without RVO. LPA levels were significantly associated with visual acuity impairments and expression levels of MCP-1, VEGF-A, IL-6, and IL-8. Changes in central macular thickness secondary to RVO were also correlated with ATX expression. These findings imply that both LPA and ATX are involved in the pathogenesis of RVO [93]. Nonetheless, the implication of LPA in the pathology of CNV and AMD remains understudied. As LPC metabolism-related markers were higher in the serum of exudative AMD patients [94], it is likely that LPA is also involved in AMD pathology. Future in vivo and in vitro investigations are required to elucidate the biological roles of LPA in pathological ocular neovascularization conditions, in addition to the physiological vascular formation.
3. Sphingolipids in Ocular Neovascularization
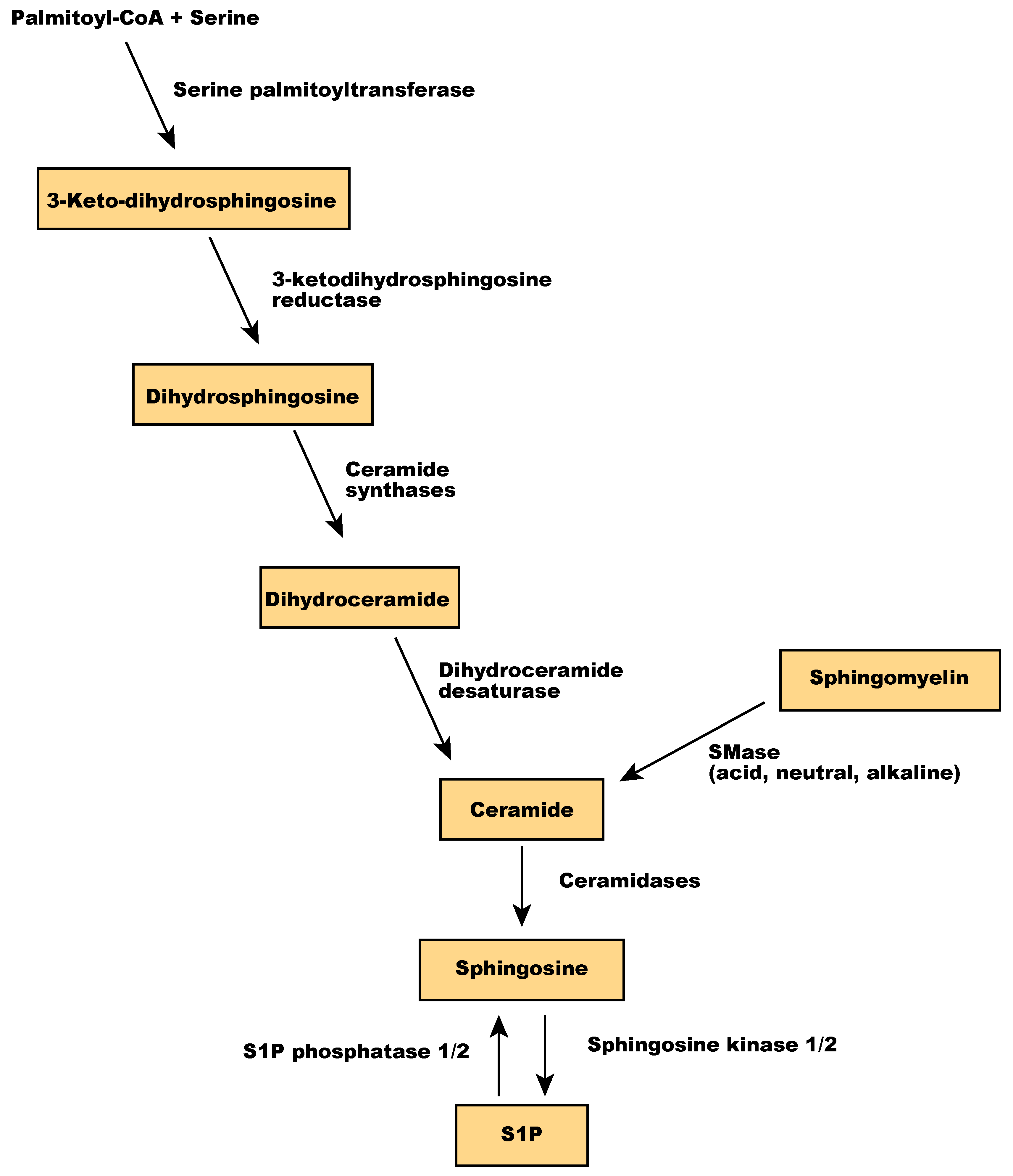
Sphingolipids are one of the main components of the membrane lipid bilayers in eukaryotic cells. They are derived from palmitoyl-CoA and serine and contain a backbone of sphingoid bases, a fatty acid attached to the long-chain sphingoid base via an amide bond, and a headgroup [95]. In mammalian cells, the headgroup of sphingolipids is a phosphocholine or oligosaccharide. Sphingomyelin, a sphingolipid with phosphocholine as a polar headgroup, is a primary component of membrane microdomains called lipid rafts. Sphingomyelin is hydrolyzed by sphingomyelinases (SMase) into ceramides and phosphocholine [96]. Ceramide regulates various cellular processes, including apoptosis and senescence [97], and is a central player in sphingolipid metabolism because sphingosine is synthesized only from ceramide. Ceramidases, including acid ceramidase, neutral ceramidase, alkaline ceramidase 1 (ACER1), ACER2, and ACER3, catalyze the hydrolysis of ceramide to sphingosine [98], which is then phosphorylated by sphingosine kinases to produce sphingosine 1-phosphate (S1P) (Figure 5). Sphingolipids act as signaling molecules regulating inflammation, cell viability, and migration [99,100]. S1P has been implicated in both physiological and pathological neovascularization, as detailed in the next section.
3.1. Sphingosine 1-Phosphate
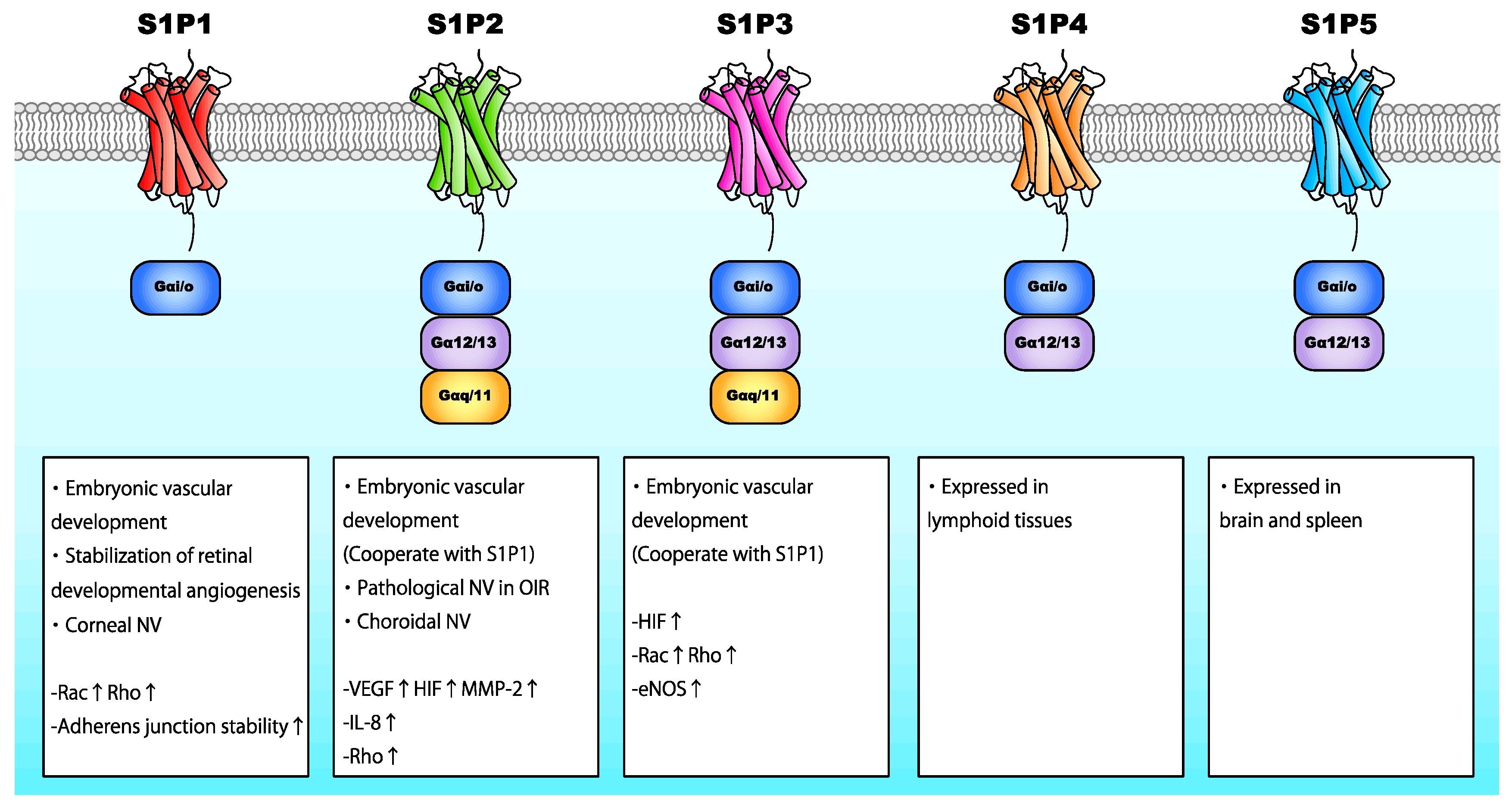
S1P is a bioactive lipid consisting of a long-chain sphingoid base and a phosphate polar headgroup. It is synthesized via the ATP-dependent phosphorylation of the hydroxyl group of sphingosine by sphingosine kinase-1 (SphK1) and SphK2 [101]. SphK1 is predominantly found in the cytosol adjacent to the plasma membrane, whereas SphK2 is located in the endoplasmic reticulum, nucleus, and mitochondria [102,103]. S1P interacts with specific G-protein-coupled receptors localized on the cell surface (S1P1-5), initiating autocrine, paracrine, or endocrine signaling. S1P receptors are coupled to Gαi/o, Gαq/11, and Gα12/13 (Figure 6) [104]. S1P1, S1P2, and S1P3 are ubiquitously expressed, whereas S1P4 is primarily expressed in lymphoid tissues, and S1P5 is expressed in the brain and spleen [105,106]. S1P levels are relatively higher in the blood than intracellularly; the concentration of serum S1P in healthy humans is approximately 1 μM [107]. S1P is stored in endothelial cells, erythrocytes, and thrombocytes [108,109].
Similar to LPA, S1P acts as an angiogenic factor, promoting embryonic vascular development. Sphk1/2−/− mice were devoid of S1P and embryonically lethal due to profound blood vessel defects between E9.5 and E13.5, leading to cranial hemorrhage and implying that S1P is essential for vascular development [110]. Among S1P receptors, S1P1 is considered the most important for the S1P-mediated effects in vascular development. S1pr1-deficient mice exhibited severe vascular smooth muscle defects and hemorrhage, which led to embryonic death [111]. Moreover, the deletion of S1pr2 and/or S1pr3 in S1pr1−/− mice elicited even more severe vascular maturation defects, leading to lethality. S1pr1-3 triple knockout mice had a reduced number of branches and capillary networks, implying that the interplay of the signaling pathways induced by S1P1, S1P2, and S1P3 is also crucial for vessel development [112]. These reports collectively imply that the S1P/S1P1-3 axis is essential for embryonic vascular development.
S1P is believed to promote neovascularization through the activation of angiogenic factors (Figure 6). S1P binding to S1P2 promotes VEGF and MMP-2 activation [113]. S1P has been shown to prevent HIF degradation, promoting HIF-1α signaling independently of hypoxia [114]. Additionally, S1P has been shown to induce HIF-1α expression by activating MAPK and PKCβI in a Gαi/o-dependent manner [115]. Conversely, HIF-2α, a HIF-α subunit predominantly expressed in endothelial cells, cardiomyocytes, and glial cells [116], upregulated the expression of SphK1 by binding to SphK1 promoter [117]. These reports imply that S1P regulates both physiological and pathological angiogenesis by promoting the expression and activation of various angiogenic factors and that hypoxic cellular responses promote S1P expression. Additionally, S1P binding to S1P2 enhanced IL-8 secretion by activating p38 MAPK and extracellular signal-regulated kinase (ERK) 1/2 pathways [118,119]. S1P/S1P1 also modulates endothelial cellular junctions by inducing the translocation of VE-cadherin to adherens junctions and enhancing barrier integrity through Rho and/or Rac activation [120]. On the other hand, the S1P2/Rho/ROCK/PTEN axis as well as VEGF signaling promotes the phosphorylation of VE-cadherin [121]. As different S1P receptors may have opposing biological functions [122], the effects of S1P vary depending on the S1P receptor.
3.2. Sphingosine 1-Phosphate and Ocular Neovascularization
S1P is generated in the retina among other tissues [123], and the essential roles of S1P and S1P receptors in the retinal vascular formation have become apparent. S1pr1 deficiency in endothelial cells increased the number of tip cells and enhanced filopodia formation. Additionally, S1pr1 depletion in endothelial cells led to ectopic endothelial hyper-sprouting and subsequent endothelial hyperplasia without pericyte coverage in the retina, resulting in vessel defects and lack of mural cells. The loss of S1pr1 also impaired VE-cadherin stabilization at endothelial cell–cell junctions, resulting in vascular leakage and lethality [124]. These reports imply an important role for S1P1 in stabilizing sprouting angiogenesis in the retina. Consistently, S1pr1-3 triple knockout mice exhibited a disorganized retinal vascular endothelium with hyper-sprouting and a lack of vascular endothelial barrier and capillary lumens [125]. Moreover, S1pr1-3 deficient mice had insufficient blood perfusion in the retina and severe vascular structure defects due to impaired endothelial cell specialization. S1P has been shown to regulate vascular maturation partly by suppressing the expression of the transcriptional factor JunB in endothelial cells located behind the vascular front. These findings imply that S1P and S1P receptors (mainly S1P1) are required for endothelial cell specialization and vessel maturation in the retina independently of VEGF.
In addition to their role in the physiological vascular development, S1P and S1P receptors have been implicated in pathological retinal neovascularization. A study using an oxygen-induced retinopathy (OIR) mouse model demonstrated that SphK2 overexpression promoted retinal angiogenesis under normoxic conditions [126], reducing the avascular retinal area and exacerbating pathological retinal neovascularization in hypoxia. Conversely, SphK2−/− ameliorated retinal neovascularization and decreased the expression of VEGF and angiopoietin, highlighting the crucial role of SphK2/S1P in normal and pathological retinal angiogenesis. Similar to SphK2−/− mice, S1P2−/− mice lacked intravitreal pathological neovascular tufts, confirming the role of S1P2 in pathological retinal neovascularization [127]. However, in contrast to SphK2 deletion, S1pr2 deficiency significantly decreased the avascular retinal area and restored the formation of retinal vasculatures and capillary plexus, partly due to the negative regulation of eNOS. Considering that S1P1 is the most important S1P receptor regulating retinal vessel formation, these results imply that S1P is required for both physiological and pathological retinal angiogenesis and that S1P/S1P2 may be an essential regulator of pathological retinal neovascularization without affecting the retinal vascular morphology.
Laser-induced CNV and sub-retinal fibrosis induction were alleviated by the intravitreal administration of anti-S1P antibodies [128]. Several humanized anti-S1P monoclonal antibodies repressed CNV in mouse models by inhibiting the IL-8-mediated lymphocyte trafficking [129]. Notably, S1P2 promoted CNV formation by regulating the production of angiogenic factors and inflammatory mediators as well as promoting barrier disruption [130,131]. Although the mechanisms underlying the S1P regulation in pathological neovascularization remain elusive, these findings pinpoint S1P signaling as a promising therapeutic target to suppress pathological retinal and choroidal neovascularization. Unfortunately, a phase II clinical trial found that the intravitreal administration of an anti-S1P antibody could not improve visual impairment in exudative AMD patients [132]. However, there is a possibility that other anti-S1P antibodies or drugs targeting a specific S1P receptor may have the therapeutic effect for exudative AMD.
Yonetsu et al. [133] evaluated the role of S1P and S1P receptors using a corneal neovascularization rabbit model; they found that an S1P1-3 antagonist suppressed corneal neovascularization. Similarly, the S1P receptor modulator FTY720 attenuated corneal neovascularization and vascular leakage induced by VEGF or S1P [134]. Although it remains unclear which S1P receptor was responsible for these effects, these observations imply that S1P and S1P receptors are also involved in the pathogenesis of corneal neovascularization.
4. The Role of Fatty Acids and Their Metabolites in Ocular Neovascularization
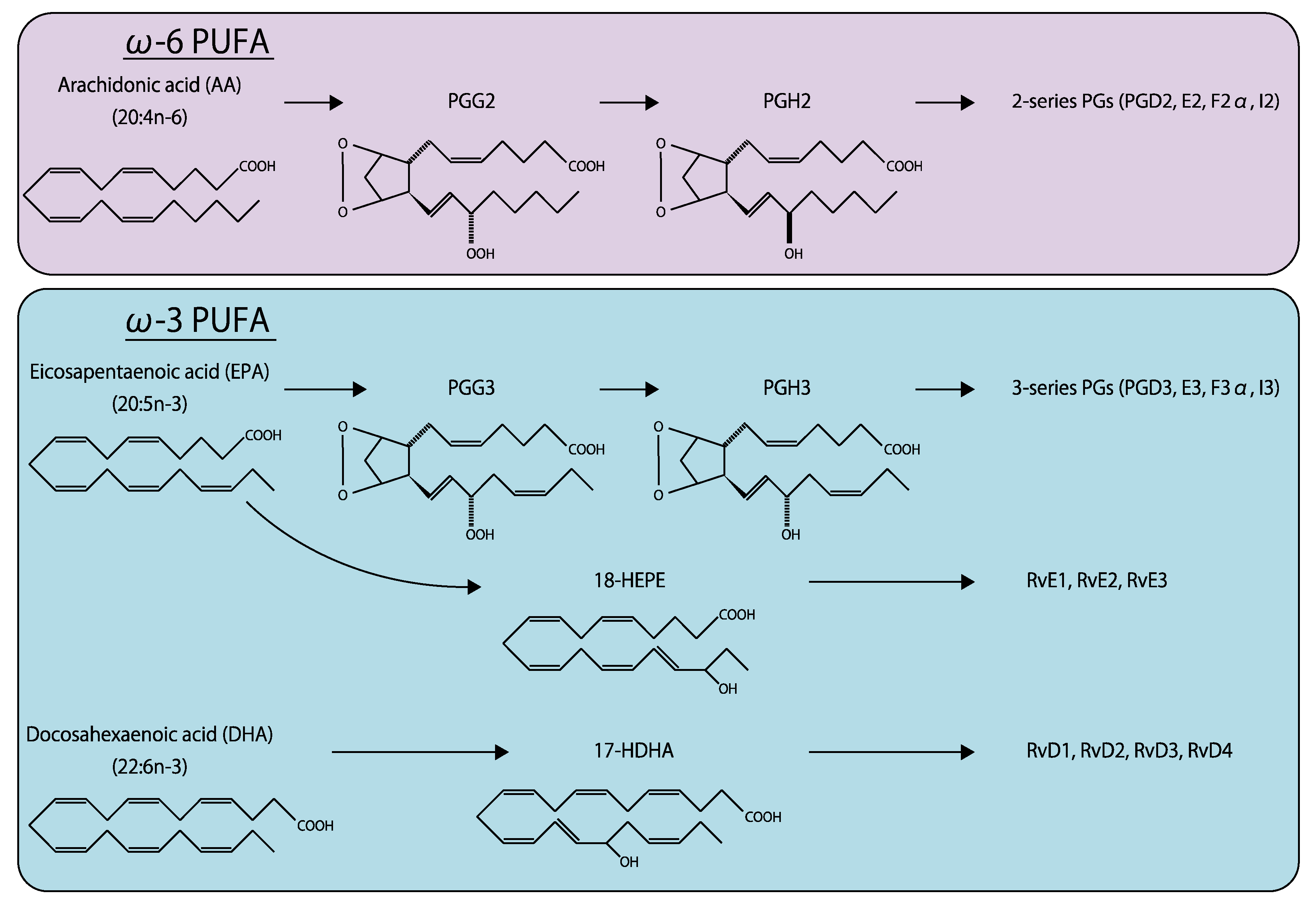
Eicosanoids are fatty acids synthesized from C20 fatty acids, such as arachidonic acid (AA) and eicosapentaenoic acid (EPA), by fatty acid oxygenases (Figure 7) [135]. Docosanoids are derived from C22 fatty acids, including docosahexaenoic acid (DHA). ω-6 fatty acids, such as prostaglandin (PG), thromboxane (TX), leukotriene, and lipoxin, are produced from AA [136]. These AA metabolites are further metabolized by cyclooxygenase (COX)1 and 2 to produce various bioactive lipids, including PGD2, PGE2, PGF2α, prostacyclin (PGI2), and TXA2 (also known as 2-series prostanoids), which are generally considered to have proinflammatory effects. In contrast, ω-3 fatty acids generated from EPA, including PGD3, PGE3, PGF3α, PGI3, and TXA3 (3-series prostanoids), exert anti-inflammatory effects [137]. Compared with lysophospholipids, the effects of eicosanoids and docosanoids in ocular tissues have been more extensively investigated [138].
4.1. ω-6 Polyunsaturated Fatty Acids
ω-6 polyunsaturated fatty acids (PUFAs) are considered a pathological angiogenic factor in ocular tissues. Of note, AA or its derivatives promote retinal vascular degeneration, and the inhibition of multiple 2-series prostanoid receptors repressed retinal and choroidal neovascularization in animal models [11,139,140].
The biological effects of PGE2 have been extensively investigated over many years. It is synthesized by COX from PGH2, which also produces PGD2, PGF2α, PGI2, and TXA2. PGE2 is a potent angiogenic lipid mediator exerting its effects through G protein-coupled receptors (EP1-4) via autocrine and paracrine signaling. EP1 is coupled to Gαq/11, modulating intracellular Ca2+ levels [141]. EP2 and EP4 are coupled with Gαs activating adenylate cyclase, which subsequently produces cyclic adenosine monophosphate (cAMP) [142]. In contrast, EP3 is coupled with Gαi/o suppressing adenylate cyclase and promoting Ca2+ influx [143].
PGE2 promotes angiogenesis by enhancing the production of various cytokines, including tumor necrosis factor (TNF)-α, IL-6, basic fibroblast growth factor (bFGF), and VEGF [144,145]. In several tissues, PGE2 promotes VEGF and CX3CR1 expression [146,147] partially via PI3K/Akt/mTORC1 pathway activation [148,149]. In HEK-293 cells, PGE2 enhanced VEGF and VEGFR1 expression in a Gαi/o-dependent manner [150]. EP4 also promoted VEGF expression [151,152] and has recently emerged as a promising anti-cancer target by suppressing tumor angiogenesis [146,153]. Additionally, PGE2 induced TNF-α expression in macrophages [154], as well as enhanced CXCL12 expression, promoting inflammation and angiogenesis [155].
Hypoxia induced the expression of COX-2 and PGE2 in retinal Müller cells; PGE2 subsequently enhanced VEGF expression, likely via EP2 and/or EP4 [156]. These results imply that PGE2 is an important prostanoid associated with retinal neovascularization. Numerous studies have shown that PGE2 mediated retinal vascularization in a Gαi/o-dependent manner. Mice lacking EP3 exhibited impaired retinal embryonic angiogenesis due to DLL4/Notch signaling inhibition in endothelial cells [157]. The same study also showed that the endothelial-specific silencing of EP3 attenuated the recruitment of tip cells, implying that EP3 modulates developmental angiogenic processes in endothelial cells.
EP4 has also been implicated in pathological angiogenesis. PGE2/EP4 axis inhibition suppressed pathological neovascularization in OIR and laser-CNV mouse models [158]. The importance of PGE2 in corneal neovascularization has also been reported. PGE2 levels were elevated in a corneal suture-injury mouse model; PGE2 exacerbated corneal neovascularization by promoting chronic inflammatory neovascularization [159]. It is worthy to note that COX2 inhibitors suppressed ocular pathological neovascularization in retina, choroid, and cornea [160,161,162], implicating COX2 is also involved in the pathogenesis of ocular neovascularization as well as PGE2.
4.2. ω-3 Polyunsaturated Fatty Acids
In contrast to 2-series prostanoids, 3-series prostanoids have anti-angiogenic and anti-inflammatory effects that protect against several disorders [163]. EPA and DHA found in retinal microvessels are thought to have protective effects against retinal vascular diseases [164]. For instance, ω-3 PUFAs (EPA and DHA) inhibited the expression of TNF-α, IL-1β, VEGF, and cell adhesion molecules in the retina in response to hypoxia or angiogenic factors [165,166]. Similarly, bioactive compounds derived from EPA and DHA, including resolvin E1 (RvE1) and D1 (RvD1), suppressed retinal vascular diseases [167,168].
Oral supplementation with ω-3 PUFAs, including EPA and DHA, suppressed retinal obliteration and pathological neovascularization in an OIR mouse model, by inhibiting proinflammatory cytokine secretion in retinal microglia [169]. RvD1 and RvE1 also inhibited neovascularization, implying that ω-3 PUFA supplementation is a promising prevention strategy for retinal vascular diseases. Additionally, peroxisome proliferator-activated receptor (PPAR), whose ligand is ω-3 PUFAs, reportedly has protective effects against pathological retinal and choroidal neovascularization [166,170,171], implicating that PPAR agonists can be considered as a therapeutic option. Furthermore, dietary intake of ω-3 PUFAs may protect against AMD. The prevalence of exudative AMD presenting with CNV was higher in people who did not consume fish, which are enriched in EPA and DHA [172]. Furthermore, ω-3 PUFA intake lowered the twelve-year incidence of AMD [173]. These findings strongly imply that ω-3 PUFAs have a protective effect against AMD. Consistently, ω-3 PUFA supplementation suppressed laser-induced CNV in mice; it also decreased intravitreal concentrations of VEGF-A and alleviated exudative CNV in humans [174,175]. RvD1 and RvE1 attenuated corneal neovascularization induced by herpes simplex virus (HSV)-1 infection, manual suture, or implantation of pellet secreting angiogenic factors. Their anti-angiogenic effects were attributed to their ability to inhibit the infiltration of inflammatory cells and secretion of cytokines, such as VEGF-A, MMP-9, IL-1β, TNF-α [176,177]. The findings of these studies indicate ω-3 PUFAs to be a promising therapeutic option for patients with ocular pathological neovascularization.
5. Future Perspectives
Numerous studies on membrane lipids and their metabolites demonstrate the crucial role of lipid signaling in ocular neovascularization, among other conditions. Glycerophospholipids, sphingolipids, and fatty acids have strong pro-angiogenic or anti-angiogenic effects by activating complex signaling circuits. Future studies are required to elucidate the mechanisms underlying the effects of lipid signaling in ocular neovascularization and other human disorders.
Author Contributions
Conceptualization, R.T. and H.K.; writing—original draft preparation, R.T.; writing—review and editing, H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Authors appreciate Makoto Aihara, (Professor and Chair of Department of Ophthalmology, Graduate School of Medicine, The University of Tokyo), and Megumi Honjo for their assistance.
Conflicts of Interest
Authors declare no conflict of interest.
Abbreviations
| AA | arachidonic acid |
| ACER | alkaline ceramidase |
| AGK | acylglycerol kinase |
| Akt | protein kinase B |
| AMD | age-related macular degeneration |
| ATX | autotaxin |
| cAMP | cyclic adenosine monophosphate |
| CNV | choroidal neovascularization |
| COX | cyclooxygenase |
| DHA | docosahexaenoic acid |
| DLL4 | delta-like ligand |
| eNOS | endothelial nitric oxide synthase |
| EP | prostaglandin E2 receptor |
| EPA | eicosapentaenoic acid |
| ERK | extracellular signal-regulated kinase |
| FGF | fibroblast growth factor |
| GLUT | glucose transporter |
| GTPases | guanosine triphosphatases |
| HIF | hypoxia inducible factor |
| HSV | herpes simplex virus |
| HRE | hypoxia response elements |
| IL | interleukin |
| iNOS | inducible nitric oxide synthase |
| LPA | lysophosphatidic acid |
| LPC | lysophosphatidylcholine |
| LPE | lysophosphatidylethanolamine |
| LysoPS | lysophosphatidylserine |
| MAPK | mitogen-activated protein kinase |
| mTOR | mammalian target of rapamycin |
| NV | neovascularization |
| OIR | oxygen-induced retinopathy |
| PA | phosphatidic acid |
| PA-PLA | phosphatidic acid specific phospholipase A |
| PC | phosphatidyl choline |
| PDGF | platelet-derived growth factor |
| PDR | proliferative diabetic retinopathy |
| PE | phosphatidylethanolamines |
| PG | prostaglandin |
| PGI2 | prostacyclin |
| PI3K | phosphoinositide 3-kinase |
| PKC | protein kinase C |
| PLA | phospholipase A |
| PLD | phospholipase D |
| PPAR | peroxisome proliferator-activated receptor |
| PS | phosphatidylserine |
| PUFA | polyunsaturated fatty acid |
| ROCK | Rho-associated protein kinase |
| ROP | retinopathy of prematurity |
| RPE | retinal pigment epithelium |
| Rv | resolvin |
| RVO | retinal vein occlusion |
| SMase | sphingomyelinase |
| STAT | signal transducers and activator of transcription |
| SphK | sphingosine kinase |
| S1P | sphingosine 1-phosphate |
| TAZ | transcriptional co-activator with PDZ-binding motif |
| TNF | tumor Necrosis Factor |
| TX | thromboxane |
| VCAM | vascular cell adhesion molecule |
| VE | vascular endothelial |
| VEGF | vascular endothelial growth factor |
| VEGFR | vascular endothelial growth factor receptor |
| YAP | yes-associated protein |
References
- Saliba, A.E.; Vonkova, I.; Gavin, A.C. The systematic analysis of protein-lipid interactions comes of age. Nat. Rev. Mol. Cell Biol. 2015, 16, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.B. Membranes and evolution. Curr. Biol. 2018, 28, R381–R385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane Lipid Composition: Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Escribá, P.V. Membrane-lipid therapy: A new approach in molecular medicine. Trends Mol. Med. 2006, 12, 34–43. [Google Scholar] [CrossRef]
- Hui, D.Y. Intestinal phospholipid and lysophospholipid metabolism in cardiometabolic disease. Curr. Opin. Lipidol. 2016, 27, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Nagahashi, M.; Abe, M.; Sakimura, K.; Takabe, K.; Wakai, T. The role of sphingosine-1-phosphate in inflammation and cancer progression. Cancer Sci. 2018, 109, 3671–3678. [Google Scholar] [CrossRef] [Green Version]
- Saito, J.; Morishige, N.; Chikama, T.; Gu, J.; Sekiguchi, K.; Nishida, T. Differential regulation of focal adhesion kinase and paxillin phosphorylation by the small GTP-binding protein Rho in human corneal epithelial cells. Jpn. J. Ophthalmol. 2004, 48, 199–207. [Google Scholar] [CrossRef]
- Solati, Z.; Ravandi, A. Lipidomics of Bioactive Lipids in Acute Coronary Syndromes. Int. J. Mol. Sci. 2019, 20, 1051. [Google Scholar] [CrossRef] [Green Version]
- Kermorvant-Duchemin, E.; Sennlaub, F.; Sirinyan, M.; Brault, S.; Andelfinger, G.; Kooli, A.; Germain, S.; Ong, H.; d’Orleans-Juste, P.; Gobeil, F., Jr.; et al. Trans-arachidonic acids generated during nitrative stress induce a thrombospondin-1-dependent microvascular degeneration. Nat. Med. 2005, 11, 1339–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierro, I.M.; Kutok, J.L.; Serhan, C.N. Novel lipid mediator regulators of endothelial cell proliferation and migration: Aspirin-triggered-15R-lipoxin A(4) and lipoxin A(4). J. Pharmacol. Exp. Ther. 2002, 300, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Moccia, F.; Negri, S.; Shekha, M.; Faris, P.; Guerra, G. Endothelial Ca(2+) Signaling, Angiogenesis and Vasculogenesis: Just What It Takes to Make a Blood Vessel. Int. J. Mol. Sci. 2019, 20, 3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriya, J.; Minamino, T. Angiogenesis, Cancer, and Vascular Aging. Front. Cardiovasc. Med. 2017, 4, 65. [Google Scholar] [CrossRef] [PubMed]
- Ratajska, A.; Jankowska-Steifer, E.; Czarnowska, E.; Olkowski, R.; Gula, G.; Niderla-Bielińska, J.; Flaht-Zabost, A.; Jasińska, A. Vasculogenesis and Its Cellular Therapeutic Applications. Cells Tissues Organs 2017, 203, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Afzal, A.; Shaw, L.C.; Ljubimov, A.V.; Boulton, M.E.; Segal, M.S.; Grant, M.B. Retinal and choroidal microangiopathies: Therapeutic opportunities. Microvasc. Res. 2007, 74, 131–144. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.Y. Targeting Tumor Adaption to Chronic Hypoxia: Implications for Drug Resistance, and How It Can Be Overcome. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P. Mechanisms of angiogenesis and arteriogenesis. Nat. Med. 2000, 6, 389–395. [Google Scholar] [CrossRef]
- Rajabi, M.; Mousa, S.A. The Role of Angiogenesis in Cancer Treatment. Biomedicines 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Viallard, C.; Larrivée, B. Tumor angiogenesis and vascular normalization: Alternative therapeutic targets. Angiogenesis 2017, 20, 409–426. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duran, C.L.; Howell, D.W.; Dave, J.M.; Smith, R.L.; Torrie, M.E.; Essner, J.J.; Bayless, K.J. Molecular Regulation of Sprouting Angiogenesis. Compr. Physiol. 2017, 8, 153–235. [Google Scholar] [CrossRef] [PubMed]
- De Bock, K.; Georgiadou, M.; Carmeliet, P. Role of endothelial cell metabolism in vessel sprouting. Cell Metab. 2013, 18, 634–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 1996, 380, 435–439. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Karaman, S.; Leppänen, V.M.; Alitalo, K. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145, 151019. [Google Scholar] [CrossRef] [Green Version]
- Estrada, C.C.; Maldonado, A.; Mallipattu, S.K. Therapeutic Inhibition of VEGF Signaling and Associated Nephrotoxicities. J. Am. Soc. Nephrol. 2019, 30, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Fryer, B.H.; Field, J. Rho, Rac, Pak and angiogenesis: Old roles and newly identified responsibilities in endothelial cells. Cancer Lett. 2005, 229, 13–23. [Google Scholar] [CrossRef]
- Bhattarai, D.; Xu, X.; Lee, K. Hypoxia-inducible factor-1 (HIF-1) inhibitors from the last decade (2007 to 2016): A “structure-activity relationship” perspective. Med. Res. Rev. 2018, 38, 1404–1442. [Google Scholar] [CrossRef]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [Green Version]
- Petreaca, M.L.; Yao, M.; Liu, Y.; Defea, K.; Martins-Green, M. Transactivation of vascular endothelial growth factor receptor-2 by interleukin-8 (IL-8/CXCL8) is required for IL-8/CXCL8-induced endothelial permeability. Mol. Biol. Cell 2007, 18, 5014–5023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallez, Y.; Vilgrain, I.; Huber, P. Angiogenesis: The VE-cadherin switch. Trends Cardiovasc. Med. 2006, 16, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, J.W.; Madsen, J.R. VEGF Signaling in Neurological Disorders. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Hou, H.; Liang, H.; Weinreb, R.N.; Wang, H.; Wang, Y. Bone marrow-derived cells in ocular neovascularization: Contribution and mechanisms. Angiogenesis 2016, 19, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.H.; Wang, Z.; Sun, Y.; Chen, J. Animal models of ocular angiogenesis: From development to pathologies. FASEB J. 2017, 31, 4665–4681. [Google Scholar] [CrossRef] [Green Version]
- Campochiaro, P.A. Ocular neovascularization. J. Mol. Med. (Berl) 2013, 91, 311–321. [Google Scholar] [CrossRef]
- Aiello, L.P.; Avery, R.L.; Arrigg, P.G.; Keyt, B.A.; Jampel, H.D.; Shah, S.T.; Pasquale, L.R.; Thieme, H.; Iwamoto, M.A.; Park, J.E.; et al. Vascular endothelial growth factor in ocular fluid of patients with diabetic retinopathy and other retinal disorders. N. Engl. J. Med. 1994, 331, 1480–1487. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Clark, W.L.; Boyer, D.S.; Heier, J.S.; Brown, D.M.; Vitti, R.; Kazmi, H.; Berliner, A.J.; Erickson, K.; Chu, K.W.; et al. Intravitreal aflibercept for macular edema following branch retinal vein occlusion: The 24-week results of the VIBRANT study. Ophthalmology 2015, 122, 538–544. [Google Scholar] [CrossRef]
- Brown, D.M.; Schmidt-Erfurth, U.; Do, D.V.; Holz, F.G.; Boyer, D.S.; Midena, E.; Heier, J.S.; Terasaki, H.; Kaiser, P.K.; Marcus, D.M.; et al. Intravitreal Aflibercept for Diabetic Macular Edema: 100-Week Results From the VISTA and VIVID Studies. Ophthalmology 2015, 122, 2044–2052. [Google Scholar] [CrossRef]
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-related macular degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Brown, D.M.; Kaiser, P.K.; Michels, M.; Soubrane, G.; Heier, J.S.; Kim, R.Y.; Sy, J.P.; Schneider, S. Ranibizumab versus verteporfin for neovascular age-related macular degeneration. N. Engl. J. Med. 2006, 355, 1432–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, P.J.; Brown, D.M.; Heier, J.S.; Boyer, D.S.; Kaiser, P.K.; Chung, C.Y.; Kim, R.Y. Ranibizumab for neovascular age-related macular degeneration. N. Engl. J. Med. 2006, 355, 1419–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heier, J.S.; Brown, D.M.; Chong, V.; Korobelnik, J.F.; Kaiser, P.K.; Nguyen, Q.D.; Kirchhof, B.; Ho, A.; Ogura, Y.; Yancopoulos, G.D.; et al. Intravitreal aflibercept (VEGF trap-eye) in wet age-related macular degeneration. Ophthalmology 2012, 119, 2537–2548. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Illingworth, C. Treatments for corneal neovascularization: A review. Cornea 2011, 30, 927–938. [Google Scholar] [CrossRef]
- Yaylali, V.; Ohta, T.; Kaufman, S.C.; Maitchouk, D.Y.; Beuerman, R.W. In vivo confocal imaging of corneal neovascularization. Cornea 1998, 17, 646–653. [Google Scholar] [CrossRef]
- Scroggs, M.W.; Proia, A.D.; Smith, C.F.; Halperin, E.C.; Klintworth, G.K. The effect of total-body irradiation on corneal neovascularization in the Fischer 344 rat after chemical cauterization. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2105–2111. [Google Scholar]
- Hamill, C.E.; Bozorg, S.; Peggy Chang, H.Y.; Lee, H.; Sayegh, R.R.; Shukla, A.N.; Chodosh, J. Corneal alkali burns: A review of the literature and proposed protocol for evaluation and treatment. Int. Ophthalmol. Clin. 2013, 53, 185–194. [Google Scholar] [CrossRef]
- Safvati, A.; Cole, N.; Hume, E.; Willcox, M. Mediators of neovascularization and the hypoxic cornea. Curr. Eye Res. 2009, 34, 501–514. [Google Scholar] [CrossRef]
- Roshandel, D.; Eslani, M.; Baradaran-Rafii, A.; Cheung, A.Y.; Kurji, K.; Jabbehdari, S.; Maiz, A.; Jalali, S.; Djalilian, A.R.; Holland, E.J. Current and emerging therapies for corneal neovascularization. Ocul. Surf. 2018, 16, 398–414. [Google Scholar] [CrossRef]
- Frisardi, V.; Panza, F.; Seripa, D.; Farooqui, T.; Farooqui, A.A. Glycerophospholipids and glycerophospholipid-derived lipid mediators: A complex meshwork in Alzheimer’s disease pathology. Prog. Lipid Res. 2011, 50, 313–330. [Google Scholar] [CrossRef]
- Ecker, J.; Liebisch, G. Application of stable isotopes to investigate the metabolism of fatty acids, glycerophospholipid and sphingolipid species. Prog. Lipid Res. 2014, 54, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, M.; Hokynar, K.; Somerharju, P. Mechanisms of glycerophospholipid homeostasis in mammalian cells. Prog. Lipid Res. 2011, 50, 240–257. [Google Scholar] [CrossRef] [PubMed]
- Croset, M.; Brossard, N.; Polette, A.; Lagarde, M. Characterization of plasma unsaturated lysophosphatidylcholines in human and rat. Biochem. J. 2000, 345 Pt 1, 61–67. [Google Scholar] [CrossRef]
- D’Arrigo, P.; Servi, S. Synthesis of lysophospholipids. Molecules 2010, 15, 1354–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moolenaar, W.H.; van Meeteren, L.A.; Giepmans, B.N. The ins and outs of lysophosphatidic acid signaling. Bioessays 2004, 26, 870–881. [Google Scholar] [CrossRef]
- Vukotic, M.; Nolte, H.; König, T.; Saita, S.; Ananjew, M.; Krüger, M.; Tatsuta, T.; Langer, T. Acylglycerol Kinase Mutated in Sengers Syndrome Is a Subunit of the TIM22 Protein Translocase in Mitochondria. Mol. Cell 2017, 67, 471–483.e7. [Google Scholar] [CrossRef]
- Zhao, Y.; Natarajan, V. Lysophosphatidic acid signaling in airway epithelium: Role in airway inflammation and remodeling. Cell. Signal. 2009, 21, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Herr, D.R.; Noguchi, K.; Yung, Y.C.; Lee, C.W.; Mutoh, T.; Lin, M.E.; Teo, S.T.; Park, K.E.; Mosley, A.N.; et al. LPA receptors: Subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 157–186. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Su, J.; Chen, J.; Chen, W.; Chen, X.; Peng, C. The role of lysophosphatidic acid in the physiology and pathology of the skin. Life Sci. 2019, 220, 194–200. [Google Scholar] [CrossRef]
- Palis, J.; Robertson, S.; Kennedy, M.; Wall, C.; Keller, G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development 1999, 126, 5073–5084. [Google Scholar]
- Aoki, J. Mechanisms of lysophosphatidic acid production. Semin. Cell Dev. Biol. 2004, 15, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, N.; Ishii, I.; Contos, J.J.; Weiner, J.A.; Chun, J. Lysophospholipid receptors. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 507–534. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.W.; Clair, T.; Kim, Y.S.; McMarlin, A.; Schiffmann, E.; Liotta, L.A.; Stracke, M.L. Autotaxin (NPP-2), a metastasis-enhancing motogen, is an angiogenic factor. Cancer Res. 2001, 61, 6938–6944. [Google Scholar] [PubMed]
- Fukushima, N.; Chun, J. The LPA receptors. Prostaglandins Other Lipid Mediat. 2001, 64, 21–32. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, D.; Oh, Y.S.; Jun, H.S. Lysophosphatidic Acid Signaling in Diabetic Nephropathy. Int. J. Mol. Sci. 2019, 20, 2850. [Google Scholar] [CrossRef] [Green Version]
- Park, F.; Miller, D.D. Role of lysophosphatidic acid and its receptors in the kidney. Physiol. Genomics 2017, 49, 659–666. [Google Scholar] [CrossRef]
- Lin, K.H.; Chiang, J.C.; Ho, Y.H.; Yao, C.L.; Lee, H. Lysophosphatidic Acid and Hematopoiesis: From Microenvironmental Effects to Intracellular Signaling. Int. J. Mol. Sci. 2020, 21, 2015. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.C. Lysophosphatidic Acid and Autotaxin-associated Effects on the Initiation and Progression of Colorectal Cancer. Cancers (Basel) 2019, 11, 958. [Google Scholar] [CrossRef] [Green Version]
- Yung, Y.C.; Stoddard, N.C.; Chun, J. LPA receptor signaling: Pharmacology, physiology, and pathophysiology. J. Lipid Res. 2014, 55, 1192–1214. [Google Scholar] [CrossRef] [Green Version]
- Teo, S.T.; Yung, Y.C.; Herr, D.R.; Chun, J. Lysophosphatidic acid in vascular development and disease. IUBMB Life 2009, 61, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Okudaira, S.; Kishi, Y.; Ohkawa, R.; Iseki, S.; Ota, M.; Noji, S.; Yatomi, Y.; Aoki, J.; Arai, H. Autotaxin stabilizes blood vessels and is required for embryonic vasculature by producing lysophosphatidic acid. J. Biol. Chem. 2006, 281, 25822–25830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offermanns, S.; Mancino, V.; Revel, J.P.; Simon, M.I. Vascular system defects and impaired cell chemokinesis as a result of Galpha13 deficiency. Science 1997, 275, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Yukiura, H.; Hama, K.; Nakanaga, K.; Tanaka, M.; Asaoka, Y.; Okudaira, S.; Arima, N.; Inoue, A.; Hashimoto, T.; Arai, H.; et al. Autotaxin regulates vascular development via multiple lysophosphatidic acid (LPA) receptors in zebrafish. J. Biol. Chem. 2011, 286, 43972–43983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamijo, H.; Matsumura, Y.; Thumkeo, D.; Koike, S.; Masu, M.; Shimizu, Y.; Ishizaki, T.; Narumiya, S. Impaired vascular remodeling in the yolk sac of embryos deficient in ROCK-I and ROCK-II. Genes Cells 2011, 16, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.W.; Chang, W.M.; Chen, K.H.; Hong, C.Z.; Chang, P.J.; Hsu, H.C. Lysophosphatidic acid enhanced the angiogenic capability of human chondrocytes by regulating Gi/NF-kB-dependent angiogenic factor expression. PLoS ONE 2014, 9, e95180. [Google Scholar] [CrossRef]
- Wu, P.Y.; Lin, Y.C.; Lan, S.Y.; Huang, Y.L.; Lee, H. Aromatic hydrocarbon receptor inhibits lysophosphatidic acid-induced vascular endothelial growth factor-A expression in PC-3 prostate cancer cells. Biochem. Biophys. Res. Commun. 2013, 437, 440–445. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Wang, F.; Wang, X.; Yang, J.; Li, Z.; Cong, X.; Chen, X. Lysophosphatidic acid promotes secretion of VEGF by increasing expression of 150-kD Oxygen-regulated protein (ORP150) in mesenchymal stem cells. Biochim. Biophys. Acta 2013, 1831, 1426–1434. [Google Scholar] [CrossRef]
- Lee, J.; Park, S.Y.; Lee, E.K.; Park, C.G.; Chung, H.C.; Rha, S.Y.; Kim, Y.K.; Bae, G.U.; Kim, B.K.; Han, J.W.; et al. Activation of hypoxia-inducible factor-1alpha is necessary for lysophosphatidic acid-induced vascular endothelial growth factor expression. Clin. Cancer Res. 2006, 12, 6351–6358. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S.; Sengupta, S.; Berk, M.; Kwak, Y.G.; Escobar, P.F.; Belinson, J.; Mok, S.C.; Xu, Y. Hypoxia enhances lysophosphatidic acid responsiveness in ovarian cancer cells and lysophosphatidic acid induces ovarian tumor metastasis in vivo. Cancer Res. 2006, 66, 7983–7990. [Google Scholar] [CrossRef] [Green Version]
- Aki, Y.; Kondo, A.; Nakamura, H.; Togari, A. Lysophosphatidic acid-stimulated interleukin-6 and -8 synthesis through LPA1 receptors on human osteoblasts. Arch. Oral. Biol. 2008, 53, 207–213. [Google Scholar] [CrossRef]
- Chen, R.J.; Chen, S.U.; Chou, C.H.; Lin, M.C. Lysophosphatidic acid receptor 2/3-mediated IL-8-dependent angiogenesis in cervical cancer cells. Int. J. Cancer 2012, 131, 789–802. [Google Scholar] [CrossRef]
- Shimada, H.; Rajagopalan, L.E. Rho kinase-2 activation in human endothelial cells drives lysophosphatidic acid-mediated expression of cell adhesion molecules via NF-kappaB p65. J. Biol. Chem. 2010, 285, 12536–12542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brault, S.; Gobeil, F., Jr.; Fortier, A.; Honoré, J.C.; Joyal, J.S.; Sapieha, P.S.; Kooli, A.; Martin, E.; Hardy, P.; Ribeiro-da-Silva, A.; et al. Lysophosphatidic acid induces endothelial cell death by modulating the redox environment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1174–R1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, M.; Rashid, M.H.; Fujita, R.; Contos, J.J.; Chun, J.; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med. 2004, 10, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Strunnikova, N.V.; Maminishkis, A.; Barb, J.J.; Wang, F.; Zhi, C.; Sergeev, Y.; Chen, W.; Edwards, A.O.; Stambolian, D.; Abecasis, G.; et al. Transcriptome analysis and molecular signature of human retinal pigment epithelium. Hum. Mol. Genet. 2010, 19, 2468–2486. [Google Scholar] [CrossRef]
- Zhou, W.L.; Sugioka, M.; Yamashita, M. Lysophosphatidic acid-induced Ca(2+) mobilization in the neural retina of chick embryo. J. Neurobiol. 1999, 41, 495–504. [Google Scholar] [CrossRef]
- Fincher, J.; Whiteneck, C.; Birgbauer, E. G-protein-coupled receptor cell signaling pathways mediating embryonic chick retinal growth cone collapse induced by lysophosphatidic acid and sphingosine-1-phosphate. Dev. Neurosci. 2014, 36, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Lidgerwood, G.E.; Morris, A.J.; Conquest, A.; Daniszewski, M.; Rooney, L.A.; Lim, S.Y.; Hernández, D.; Liang, H.H.; Allen, P.; Connell, P.P.; et al. Role of lysophosphatidic acid in the retinal pigment epithelium and photoreceptors. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2018, 1863, 750–761. [Google Scholar] [CrossRef]
- Yasuda, D.; Kobayashi, D.; Akahoshi, N.; Ohto-Nakanishi, T.; Yoshioka, K.; Takuwa, Y.; Mizuno, S.; Takahashi, S.; Ishii, S. Lysophosphatidic acid-induced YAP/TAZ activation promotes developmental angiogenesis by repressing Notch ligand Dll4. J. Clin. Investig. 2019, 129, 4332–4349. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Nawaz, M.I.; Mohammad, G.; Siddiquei, M.M.; Alam, K.; Mousa, A.; Opdenakker, G. Expression of bioactive lysophospholipids and processing enzymes in the vitreous from patients with proliferative diabetic retinopathy. Lipids Health Dis. 2014, 13, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu El-Asrar, A.M.; Mohammad, G.; Nawaz, M.I.; Siddiquei, M.M.; Kangave, D.; Opdenakker, G. Expression of lysophosphatidic acid, autotaxin and acylglycerol kinase as biomarkers in diabetic retinopathy. Acta Diabetol. 2013, 50, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Dacheva, I.; Ullmer, C.; Ceglowska, K.; Nogoceke, E.; Hartmann, G.; Müller, S.; Rejdak, R.; Nowomiejska, K.; Reich, M.; Nobl, M.; et al. LYSOPHOSPHATIDIC ACIDS AND AUTOTAXIN IN RETINAL VEIN OCCLUSION. Retina 2016, 36, 2311–2318. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Moaddel, R.; Cotch, M.F.; Jonasson, F.; Eiriksdottir, G.; Harris, T.B.; Launer, L.J.; Sun, K.; Klein, R.; Schaumberg, D.A.; et al. Serum lipids in adults with late age-related macular degeneration: A case-control study. Lipids Health Dis. 2019, 18, 7. [Google Scholar] [CrossRef] [Green Version]
- Van Echten-Deckert, G.; Alam, S. Sphingolipid metabolism - an ambiguous regulator of autophagy in the brain. Biol. Chem. 2018, 399, 837–850. [Google Scholar] [CrossRef]
- Bandet, C.L.; Tan-Chen, S.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism: New Insight into Ceramide-Induced Lipotoxicity in Muscle Cells. Int. J. Mol. Sci. 2019, 20, 479. [Google Scholar] [CrossRef] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Coant, N.; Sakamoto, W.; Mao, C.; Hannun, Y.A. Ceramidases, roles in sphingolipid metabolism and in health and disease. Adv. Biol. Regul. 2017, 63, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, T.; Hla, T. Structural and functional characteristics of S1P receptors. J. Cell Biochem. 2004, 92, 913–922. [Google Scholar] [CrossRef]
- Takabe, K.; Paugh, S.W.; Milstien, S.; Spiegel, S. “Inside-out” signaling of sphingosine-1-phosphate: Therapeutic targets. Pharmacol. Rev. 2008, 60, 181–195. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell. Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Siow, D.L.; Anderson, C.D.; Berdyshev, E.V.; Skobeleva, A.; Natarajan, V.; Pitson, S.M.; Wattenberg, B.W. Sphingosine kinase localization in the control of sphingolipid metabolism. Adv. Enzyme Regul. 2011, 51, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Strub, G.M.; Paillard, M.; Liang, J.; Gomez, L.; Allegood, J.C.; Hait, N.C.; Maceyka, M.; Price, M.M.; Chen, Q.; Simpson, D.C.; et al. Sphingosine-1-phosphate produced by sphingosine kinase 2 in mitochondria interacts with prohibitin 2 to regulate complex IV assembly and respiration. FASEB J. 2011, 25, 600–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, A.M.; Del Poeta, M. Sphingosine-1-phosphate receptors and innate immunity. Cell Microbiol. 2018, 20, e12836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gräler, M.H.; Bernhardt, G.; Lipp, M. EDG6, a novel G-protein-coupled receptor related to receptors for bioactive lysophospholipids, is specifically expressed in lymphoid tissue. Genomics 1998, 53, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Im, D.S.; Heise, C.E.; Ancellin, N.; O’Dowd, B.F.; Shei, G.J.; Heavens, R.P.; Rigby, M.R.; Hla, T.; Mandala, S.; McAllister, G.; et al. Characterization of a novel sphingosine 1-phosphate receptor, Edg-8. J. Biol. Chem. 2000, 275, 14281–14286. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Tan, Y.; Wang, L.; Su, X.; Shi, Y. Serum sphingosine-1-phosphate levels and Sphingosine-1-Phosphate gene polymorphisms in acute respiratory distress syndrome: A multicenter prospective study. J. Transl. Med. 2020, 18, 156. [Google Scholar] [CrossRef] [Green Version]
- Poppe, A.; Moritz, E.; Geffken, M.; Schreiber, J.; Greiwe, G.; Amschler, K.; Wruck, M.L.; Schwedhelm, E.; Daum, G.; Kluge, S.; et al. Analyses of sphingosine-1-phosphate in the context of transfusion: How much is in stored blood products and in patient blood? Transfusion 2019, 59, 3071–3076. [Google Scholar] [CrossRef] [Green Version]
- Hänel, P.; Andréani, P.; Gräler, M.H. Erythrocytes store and release sphingosine 1-phosphate in blood. FASEB J. 2007, 21, 1202–1209. [Google Scholar] [CrossRef]
- Mizugishi, K.; Yamashita, T.; Olivera, A.; Miller, G.F.; Spiegel, S.; Proia, R.L. Essential role for sphingosine kinases in neural and vascular development. Mol. Cell Biol. 2005, 25, 11113–11121. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wada, R.; Yamashita, T.; Mi, Y.; Deng, C.X.; Hobson, J.P.; Rosenfeldt, H.M.; Nava, V.E.; Chae, S.S.; Lee, M.J.; et al. Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, is essential for vascular maturation. J. Clin. Investig. 2000, 106, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kono, M.; Mi, Y.; Liu, Y.; Sasaki, T.; Allende, M.L.; Wu, Y.P.; Yamashita, T.; Proia, R.L. The sphingosine-1-phosphate receptors S1P1, S1P2, and S1P3 function coordinately during embryonic angiogenesis. J. Biol. Chem. 2004, 279, 29367–29373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chumanevich, A.; Wedman, P.; Oskeritzian, C.A. Sphingosine-1-Phosphate/Sphingosine-1-Phosphate Receptor 2 Axis Can Promote Mouse and Human Primary Mast Cell Angiogenic Potential through Upregulation of Vascular Endothelial Growth Factor-A and Matrix Metalloproteinase-2. Mediat. Inflamm. 2016, 2016, 1503206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arjamaa, O.; Aaltonen, V.; Piippo, N.; Csont, T.; Petrovski, G.; Kaarniranta, K.; Kauppinen, A. Hypoxia and inflammation in the release of VEGF and interleukins from human retinal pigment epithelial cells. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 1757–1762. [Google Scholar] [CrossRef]
- Kalhori, V.; Kemppainen, K.; Asghar, M.Y.; Bergelin, N.; Jaakkola, P.; Törnquist, K. Sphingosine-1-Phosphate as a Regulator of Hypoxia-Induced Factor-1α in Thyroid Follicular Carcinoma Cells. PLoS ONE 2013, 8, e66189. [Google Scholar] [CrossRef] [Green Version]
- Haase, V.H. Hypoxia-inducible factors in the kidney. Am. J. Physiol. Ren. Physiol. 2006, 291, F271–F281. [Google Scholar] [CrossRef]
- Anelli, V.; Gault, C.R.; Cheng, A.B.; Obeid, L.M. Sphingosine kinase 1 is up-regulated during hypoxia in U87MG glioma cells. Role of hypoxia-inducible factors 1 and 2. J. Biol. Chem. 2008, 283, 3365–3375. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Hu, R.; Wang, Q.; Qi, J.; Yang, Y.; Kijlstra, A.; Yang, P. Sphingosine 1-phosphate elicits proinflammatory responses in ARPE-19 cells. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8200–8207. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, M.J.; Hirota, N.; Martin, J.G. Sphingosine 1-phosphate (S1P) induced interleukin-8 (IL-8) release is mediated by S1P receptor 2 and nuclear factor κB in BEAS-2B cells. PLoS ONE 2014, 9, e95566. [Google Scholar] [CrossRef]
- Lee, M.J.; Thangada, S.; Claffey, K.P.; Ancellin, N.; Liu, C.H.; Kluk, M.; Volpi, M.; Sha’afi, R.I.; Hla, T. Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell 1999, 99, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, T.; Skoura, A.; Wu, M.T.; Casserly, B.; Harrington, E.O.; Hla, T. Induction of vascular permeability by the sphingosine-1-phosphate receptor-2 (S1P2R) and its downstream effectors ROCK and PTEN. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1312–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, T.; Thangada, S.; Wu, M.T.; Kontos, C.D.; Wu, D.; Wu, H.; Hla, T. PTEN as an effector in the signaling of antimigratory G protein-coupled receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 4312–4317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terao, R.; Honjo, M.; Ueta, T.; Obinata, H.; Izumi, T.; Kurano, M.; Yatomi, Y.; Koso, H.; Watanabe, S.; Aihara, M. Light Stress-Induced Increase of Sphingosine 1-Phosphate in Photoreceptors and Its Relevance to Retinal Degeneration. Int. J. Mol. Sci. 2019, 20, 3670. [Google Scholar] [CrossRef] [Green Version]
- Gaengel, K.; Niaudet, C.; Hagikura, K.; Laviña, B.; Muhl, L.; Hofmann, J.J.; Ebarasi, L.; Nyström, S.; Rymo, S.; Chen, L.L.; et al. The sphingosine-1-phosphate receptor S1PR1 restricts sprouting angiogenesis by regulating the interplay between VE-cadherin and VEGFR2. Dev. Cell 2012, 23, 587–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagida, K.; Engelbrecht, E.; Niaudet, C.; Jung, B.; Gaengel, K.; Holton, K.; Swendeman, S.; Liu, C.H.; Levesque, M.V.; Kuo, A.; et al. Sphingosine 1-Phosphate Receptor Signaling Establishes AP-1 Gradients to Allow for Retinal Endothelial Cell Specialization. Dev. Cell 2020, 52, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Eresch, J.; Stumpf, M.; Koch, A.; Vutukuri, R.; Ferreirós, N.; Schreiber, Y.; Schröder, K.; Devraj, K.; Popp, R.; Huwiler, A.; et al. Sphingosine Kinase 2 Modulates Retinal Neovascularization in the Mouse Model of Oxygen-Induced Retinopathy. Investig. Ophthalmol. Vis. Sci. 2018, 59, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoura, A.; Sanchez, T.; Claffey, K.; Mandala, S.M.; Proia, R.L.; Hla, T. Essential role of sphingosine 1-phosphate receptor 2 in pathological angiogenesis of the mouse retina. J. Clin. Investig. 2007, 117, 2506–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, B.; Shen, J.; Dong, A.; Rashid, A.; Stoller, G.; Campochiaro, P.A. Blockade of sphingosine-1-phosphate reduces macrophage influx and retinal and choroidal neovascularization. J. Cell Physiol. 2009, 218, 192–198. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, N.; Jones, S.T.; Williams, D.G.; Cunningham, H.B.; Moreno, K.; Visentin, B.; Gentile, A.; Vekich, J.; Shestowsky, W.; Hiraiwa, M.; et al. Production and characterization of monoclonal anti-sphingosine-1-phosphate antibodies. J. Lipid Res. 2009, 50, 2245–2257. [Google Scholar] [CrossRef] [Green Version]
- Terao, R.; Honjo, M.; Aihara, M. Apolipoprotein M Inhibits Angiogenic and Inflammatory Response by Sphingosine 1-Phosphate on Retinal Pigment Epithelium Cells. Int. J. Mol. Sci. 2017, 19, 112. [Google Scholar] [CrossRef] [Green Version]
- Terao, R.; Honjo, M.; Totsuka, K.; Miwa, Y.; Kurihara, T.; Aihara, M. The role of sphingosine 1-phosphate receptors on retinal pigment epithelial cells barrier function and angiogenic effects. Prostaglandins Other Lipid Mediat. 2019, 145, 106365. [Google Scholar] [CrossRef] [PubMed]
- Efficacy and Safety Study of iSONEP with & without Lucentis/Avastin/Eylea to Treat Wet AMD; Lpath, Inc.: San Diego, CA, USA, 2011; Available online: https://ClinicalTrials.gov/show/NCT01414153.
- Yonesu, K.; Kawase, Y.; Inoue, T.; Takagi, N.; Tsuchida, J.; Takuwa, Y.; Kumakura, S.; Nara, F. Involvement of sphingosine-1-phosphate and S1P1 in angiogenesis: Analyses using a new S1P1 antagonist of non-sphingosine-1-phosphate analog. Biochem. Pharmacol. 2009, 77, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- LaMontagne, K.; Littlewood-Evans, A.; Schnell, C.; O’Reilly, T.; Wyder, L.; Sanchez, T.; Probst, B.; Butler, J.; Wood, A.; Liau, G.; et al. Antagonism of sphingosine-1-phosphate receptors by FTY720 inhibits angiogenesis and tumor vascularization. Cancer Res. 2006, 66, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Ahmed, S.; Stanley, D.; An, C. Eicosanoid-mediated immunity in insects. Dev. Comp. Immunol. 2018, 83, 130–143. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fatty Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, H.; Johnson, D.; Gensler, C.; Decker, E.; Zhang, G. Chemistry and biology of ω-3 PUFA peroxidation-derived compounds. Prostaglandins Other Lipid Mediat. 2017, 132, 84–91. [Google Scholar] [CrossRef]
- Hardy, P.; Beauchamp, M.; Sennlaub, F.; Gobeil, F., Jr.; Tremblay, L.; Mwaikambo, B.; Lachapelle, P.; Chemtob, S. New insights into the retinal circulation: Inflammatory lipid mediators in ischemic retinopathy. Prostaglandins Leukot. Essent. Fatty Acids 2005, 72, 301–325. [Google Scholar] [CrossRef]
- Yanni, S.E.; Clark, M.L.; Yang, R.; Bingaman, D.P.; Penn, J.S. The effects of nepafenac and amfenac on retinal angiogenesis. Brain Res. Bull. 2010, 81, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Woodward, D.F.; Wang, J.W.; Ni, M.; Bauer, A.; Martos, J.L.; Carling, R.W.; Poloso, N.J. In vivo studies validating multitargeting of prostanoid receptors for achieving superior anti-inflammatory effects. FASEB J. 2017, 31, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Wang, X.; Jeschke, U.; von Schönfeldt, V. COX-2-PGE(2)-EPs in gynecological cancers. Arch. Gynecol. Obstet. 2020, 301, 1365–1375. [Google Scholar] [CrossRef]
- Nakao, A.; Allen, M.L.; Sonnenburg, W.K.; Smith, W.L. Regulation of cAMP metabolism by PGE2 in cortical and medullary thick ascending limb of Henle’s loop. Am. J. Physiol. 1989, 256, C652–C657. [Google Scholar] [CrossRef] [PubMed]
- Namba, T.; Sugimoto, Y.; Negishi, M.; Irie, A.; Ushikubi, F.; Kakizuka, A.; Ito, S.; Ichikawa, A.; Narumiya, S. Alternative splicing of C-terminal tail of prostaglandin E receptor subtype EP3 determines G-protein specificity. Nature 1993, 365, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Bos, C.L.; Richel, D.J.; Ritsema, T.; Peppelenbosch, M.P.; Versteeg, H.H. Prostanoids and prostanoid receptors in signal transduction. Int. J. Biochem. Cell Biol. 2004, 36, 1187–1205. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K. Regulation by prostaglandin E2 and histamine of angiogenesis in inflammatory granulation tissue. Yakugaku Zasshi 2003, 123, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, M.; Nandi, P.; Omar, A.; Ugwuagbo, K.C.; Lala, P.K. EP4 as a Therapeutic Target for Aggressive Human Breast Cancer. Int. J. Mol. Sci. 2018, 19, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Croix, B.S. Improving VEGF-targeted therapies through inhibition of COX-2/PGE2 signaling. Mol. Cell Oncol. 2014, 1, e969154. [Google Scholar] [CrossRef] [Green Version]
- Dufour, M.; Faes, S.; Dormond-Meuwly, A.; Demartines, N.; Dormond, O. PGE2-induced colon cancer growth is mediated by mTORC1. Biochem. Biophys. Res. Commun. 2014, 451, 587–591. [Google Scholar] [CrossRef]
- Tang, J.; Shen, Y.; Chen, G.; Wan, Q.; Wang, K.; Zhang, J.; Qin, J.; Liu, G.; Zuo, S.; Tao, B.; et al. Activation of E-prostanoid 3 receptor in macrophages facilitates cardiac healing after myocardial infarction. Nat. Commun. 2017, 8, 14656. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, T.; Fujino, H.; Israel, D.D.; Regan, J.W.; Murayama, T. Human EP3(I) prostanoid receptor induces VEGF and VEGF receptor-1 mRNA expression. Biochem. Biophys. Res. Commun. 2008, 377, 1173–1178. [Google Scholar] [CrossRef]
- Woodward, D.F.; Wang, J.W.; Ni, M.; Bauer, A.J.; Poloso, N.J. In Vivo Choroidal Neovascularization and Macrophage Studies Provide Further Evidence for a Broad Role of Prostacyclin in Angiogenesis. J. Ocul. Pharmacol. Ther. 2019, 35, 98–105. [Google Scholar] [CrossRef]
- Takeuchi, K.; Amagase, K. Roles of Cyclooxygenase, Prostaglandin E2 and EP Receptors in Mucosal Protection and Ulcer Healing in the Gastrointestinal Tract. Curr. Pharm. Des. 2018, 24, 2002–2011. [Google Scholar] [CrossRef] [PubMed]
- Duncan, W.C.; Nio-Kobayashi, J. Targeting angiogenesis in the pathological ovary. Reprod. Fertil. Dev. 2013, 25, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Renz, H.; Gong, J.H.; Schmidt, A.; Nain, M.; Gemsa, D. Release of tumor necrosis factor-alpha from macrophages. Enhancement and suppression are dose-dependently regulated by prostaglandin E2 and cyclic nucleotides. J. Immunol. 1988, 141, 2388–2393. [Google Scholar] [PubMed]
- Obermajer, N.; Muthuswamy, R.; Odunsi, K.; Edwards, R.P.; Kalinski, P. PGE(2)-induced CXCL12 production and CXCR4 expression controls the accumulation of human MDSCs in ovarian cancer environment. Cancer Res. 2011, 71, 7463–7470. [Google Scholar] [CrossRef] [Green Version]
- Yanni, S.E.; McCollum, G.W.; Penn, J.S. Genetic deletion of COX-2 diminishes VEGF production in mouse retinal Müller cells. Exp. Eye Res. 2010, 91, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Tang, J.; Wan, Q.; Zhang, J.; Wang, K.; Shen, Y.; Yu, Y. E-Prostanoid 3 Receptor Mediates Sprouting Angiogenesis Through Suppression of the Protein Kinase A/β-Catenin/Notch Pathway. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 856–866. [Google Scholar] [CrossRef] [Green Version]
- Yanni, S.E.; Barnett, J.M.; Clark, M.L.; Penn, J.S. The role of PGE2 receptor EP4 in pathologic ocular angiogenesis. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5479–5486. [Google Scholar] [CrossRef] [Green Version]
- Liclican, E.L.; Nguyen, V.; Sullivan, A.B.; Gronert, K. Selective activation of the prostaglandin E2 circuit in chronic injury-induced pathologic angiogenesis. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6311–6320. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson-Berka, J.L.; Alousis, N.S.; Kelly, D.J.; Gilbert, R.E. COX-2 inhibition and retinal angiogenesis in a mouse model of retinopathy of prematurity. Investig. Ophthalmol. Vis. Sci. 2003, 44, 974–979. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Yanagi, Y.; Tamaki, Y.; Uchida, S.; Muranaka, K. COX-2-selective inhibitor, etodolac, suppresses choroidal neovascularization in a mice model. Biochem. Biophys. Res. Commun. 2004, 325, 461–466. [Google Scholar] [CrossRef]
- Yamada, M.; Kawai, M.; Kawai, Y.; Mashima, Y. The effect of selective cyclooxygenase-2 inhibitor on corneal angiogenesis in the rat. Curr. Eye Res. 1999, 19, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Ottes Vasconcelos, R.; Nascimento Gomes, R.; Calviello, G. Protective Effects of ω-3 PUFA in Anthracycline-Induced Cardiotoxicity: A Critical Review. Int. J. Mol. Sci. 2017, 18, 2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SanGiovanni, J.P.; Chew, E.Y. The role of omega-3 long-chain polyunsaturated fatty acids in health and disease of the retina. Prog. Retin. Eye Res. 2005, 24, 87–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Esselman, W.J.; Jump, D.B.; Busik, J.V. Anti-inflammatory effect of docosahexaenoic acid on cytokine-induced adhesion molecule expression in human retinal vascular endothelial cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4342–4347. [Google Scholar] [CrossRef]
- Stahl, A.; Sapieha, P.; Connor, K.M.; Sangiovanni, J.P.; Chen, J.; Aderman, C.M.; Willett, K.L.; Krah, N.M.; Dennison, R.J.; Seaward, M.R.; et al. Short communication: PPAR gamma mediates a direct antiangiogenic effect of omega 3-PUFAs in proliferative retinopathy. Circ. Res. 2010, 107, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Shen, J.H.; Shen, S.R.; Das, U.N. Bioactive lipids in pathological retinopathy. Crit. Rev. Food Sci. Nutr. 2014, 54, 1–16. [Google Scholar] [CrossRef]
- Suzumura, A.; Kaneko, H.; Funahashi, Y.; Takayama, K.; Nagaya, M.; Ito, S.; Okuno, T.; Hirakata, T.; Nonobe, N.; Kataoka, K.; et al. n-3 Fatty Acid and Its Metabolite 18-HEPE Ameliorate Retinal Neuronal Cell Dysfunction by Enhancing Müller BDNF in Diabetic Retinopathy. Diabetes 2020, 69, 724–735. [Google Scholar] [CrossRef]
- Connor, K.M.; SanGiovanni, J.P.; Lofqvist, C.; Aderman, C.M.; Chen, J.; Higuchi, A.; Hong, S.; Pravda, E.A.; Majchrzak, S.; Carper, D.; et al. Increased dietary intake of omega-3-polyunsaturated fatty acids reduces pathological retinal angiogenesis. Nat. Med. 2007, 13, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Murata, T.; He, S.; Hangai, M.; Ishibashi, T.; Xi, X.P.; Kim, S.; Hsueh, W.A.; Ryan, S.J.; Law, R.E.; Hinton, D.R. Peroxisome proliferator-activated receptor-gamma ligands inhibit choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2309–2317. [Google Scholar]
- SanGiovanni, J.P.; Chen, J.; Sapieha, P.; Aderman, C.M.; Stahl, A.; Clemons, T.E.; Chew, E.Y.; Smith, L.E. DNA sequence variants in PPARGC1A, a gene encoding a coactivator of the ω-3 LCPUFA sensing PPAR-RXR transcription complex, are associated with NV AMD and AMD-associated loci in genes of complement and VEGF signaling pathways. PLoS ONE 2013, 8, e53155. [Google Scholar] [CrossRef] [PubMed]
- Sangiovanni, J.P.; Agrón, E.; Meleth, A.D.; Reed, G.F.; Sperduto, R.D.; Clemons, T.E.; Chew, E.Y. {omega}-3 Long-chain polyunsaturated fatty acid intake and 12-y incidence of neovascular age-related macular degeneration and central geographic atrophy: AREDS report 30, a prospective cohort study from the Age-Related Eye Disease Study. Am. J. Clin. Nutr. 2009, 90, 1601–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augood, C.; Chakravarthy, U.; Young, I.; Vioque, J.; de Jong, P.T.; Bentham, G.; Rahu, M.; Seland, J.; Soubrane, G.; Tomazzoli, L.; et al. Oily fish consumption, dietary docosahexaenoic acid and eicosapentaenoic acid intakes, and associations with neovascular age-related macular degeneration. Am. J. Clin. Nutr. 2008, 88, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Li, J.; Sun, Y.; Fu, Z.; Liu, C.H.; Evans, L.; Tian, K.; Saba, N.; Fredrick, T.; Morss, P.; et al. Optimization of an Image-Guided Laser-Induced Choroidal Neovascularization Model in Mice. PLoS ONE 2015, 10, e0132643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezende, F.A.; Lapalme, E.; Qian, C.X.; Smith, L.E.; SanGiovanni, J.P.; Sapieha, P. Omega-3 supplementation combined with anti-vascular endothelial growth factor lowers vitreal levels of vascular endothelial growth factor in wet age-related macular degeneration. Am. J. Ophthalmol. 2014, 158, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasagi, N.K.; Reddy, P.B.; Suryawanshi, A.; Mulik, S.; Gjorstrup, P.; Rouse, B.T. Controlling herpes simplex virus-induced ocular inflammatory lesions with the lipid-derived mediator resolvin E1. J. Immunol. 2011, 186, 1735–1746. [Google Scholar] [CrossRef]
- Jin, Y.; Arita, M.; Zhang, Q.; Saban, D.R.; Chauhan, S.K.; Chiang, N.; Serhan, C.N.; Dana, R. Anti-angiogenesis effect of the novel anti-inflammatory and pro-resolving lipid mediators. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4743–4752. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of ocular neovascularization.

Figure 2.
Fundus image of proliferative diabetic retinopathy (PDR) presenting with vitreous (arrow) and preretinal hemorrhage (arrowhead).
Figure 2.
Fundus image of proliferative diabetic retinopathy (PDR) presenting with vitreous (arrow) and preretinal hemorrhage (arrowhead).

Figure 3.
Conversion of glycerophospholipids to LPA (lysophosphatidic acid).

Figure 4.
The role of LPA receptors in neovascularization-associated signaling pathways.

Figure 5.
Overview of sphingolipid metabolism.

Figure 6.
Biological effects of S1P receptors in neovascularization through the interaction with G proteins. S1P exerts various cellular functions by activating different S1P receptors.
Figure 6.
Biological effects of S1P receptors in neovascularization through the interaction with G proteins. S1P exerts various cellular functions by activating different S1P receptors.

Figure 7.
Biochemical structure and metabolism of ω-6 and ω-3 fatty acids.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Terao, R.; Kaneko, H. Lipid Signaling in Ocular Neovascularization. Int. J. Mol. Sci. 2020, 21, 4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134758
AMA Style
Terao R, Kaneko H. Lipid Signaling in Ocular Neovascularization. International Journal of Molecular Sciences. 2020; 21(13):4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134758
Chicago/Turabian StyleTerao, Ryo, and Hiroki Kaneko. 2020. "Lipid Signaling in Ocular Neovascularization" International Journal of Molecular Sciences 21, no. 13: 4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134758
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.