1. Introduction
Carnosinase 1 (CN1; EC 3.4.13.20) is a member of the M20 metalloprotease family with a catalytic domain featuring a dinuclear Zn
2+ [
1,
2]. In aqueous solution CN1 is present as homodimer with narrow substrate specificity for the naturally occurring dipeptides carnosine (β-alanyl-
L-histidine), and its derivatives anserine (β-alanyl-
N-methylhistidine) and homocarnosine (γ-amino-butyryl-
L-histidine) [
2,
3]. Carnosinase 1 (CN1) activity, defined by genetic variants, has been linked to the incidence of nephropathy in female patients with diabetes mellitus [
4,
5]. Carnosine scavenges carbonyls [
6,
7], inhibits glycation [
8] and acts as an ACE inhibitor [
9,
10,
11]. Carnosine and anserine can prevent methylglyoxal (MG)-induced advanced glycation endproducts (AGE)
N-(1-Carboxyethyl)-
L-lysine (CEL) formation, even though cellular uptake of carnosine in vitro is low. High quenching capacity of carnosine has been shown for acrolein and 4-hydroxynonenal (HNE) [
12,
13,
14]. Moreover, carnosine may act as an antioxidant, albeit this action could not unequivocally be reproduced [
15,
16,
17]. Anserine upregulates HSP70 expression under conditions of oxidative and glycating stress in tubular cells and in type 2 diabetic mice, anserine also reduces blood glucose levels and vascular permeability and proteinuria [
18]. In mammals and humans carnosine and anserine are the most abundant dipeptides, with highest concentrations in muscles [
1]. Plasma concentrations of carnosine and anserine are higher in rodents compared to humans, because rodents have no carnosine-degrading serum enzyme. Carnosine actions in humans may therefore predominately be exerted via local, tissue specific carnosine metabolism. In kidneys of mice and men, anserine and carnosine are present at high concentrations, with anserine concentrations exceeding those of carnosine [
19]. Supplementation of carnosine in diabetic rodents consistently improved renal histology and function [
20], vascular permeability [
21], and wound healing [
22], but also glucose homeostasis [
23,
24,
25,
26]. Whether the beneficial effects of carnosine resulted from interference with reactive metabolite-induced molecular pathomechanisms or primarily via improving glucose homeostasis is yet unclear. Initial intervention studies in (pre-)diabetic patients also yielded promising results, even though the half-life of carnosine in the human circulation is short even in subjects with low CN1 activity [
27].
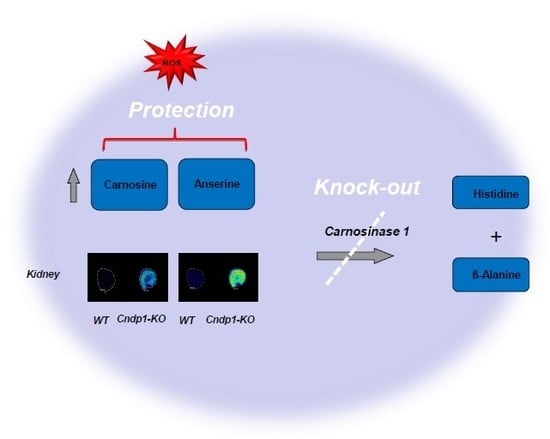
Taken together, interference with the metabolism of carnosine and anserine may be a promising therapeutic approach in patients with diabetes. To date, however, the precise molecular mechanisms of carnosine generation and related metabolic actions are not yet sufficiently understood. Therefore, we characterized a Cndp1-knockout (Cndp1-KO) mouse model with respect to systemic and local tissue carnosine and anserine metabolism and glucose homeostasis.
2. Material and Methods
2.1. Mouse Strains
Animals of the strain B6;129S5-
Cndp1tm1Lex/Mmucd (#032215; MMRRC UC Davis, Davis, UT, USA), harboring a targeted mutation in exons 8 and 9 of the
Cndp1 gene, generated by homologous recombination and verified by Southernblot [
28], were used. These mice were cross-bred with C57BL/6J animals (Charles River, Sulzfeld, Gemany) for 10 generations and offspring homozygous for the
Cndp1 knockout, verified by PCR (
https://mmrrc.ucdavis.edu/protocols/032215Geno_Protocol.pdf; supplement figure), were used in the experiments. C57BL/6J mice were used as wild type controls. Breeding scheme consisted of crossing of homozygous males with homozygous females. Generations F1 to F3 were used for the experiments. The global Cndp1-KO has been confirmed by gene expression (
https://mmrrc.ucdavis.edu/lexiconphenotype.php?id=PRT333N1). Blood chemistry (including non-fasting blood glucose, cholesterol and triglyceride concentrations) have been reported by MMRRC and were not different for
Cndp1-KO compared to WT mice (
https://mmrrc.ucdavis.edu/lexiconphenotype.php?id=PRT333N1). Animals were housed at Interfaculty Biomedical Facility (IBF) at Heidelberg University with a daily cycle of 12 h light and 12 h darkness at 23 °C. Food and water were supplied ad libitum. Experiments were performed with 11- and 55-week-old
Cndp1-KO and wild type mice. The studies were approved by the respective authorities (Regierungspräsidium Karlsruhe, Germany, 35-9165 81/G-209/16).
2.2. Carnosinase Activity
50 mg frozen tissue (either brain, liver, muscle, heart, lungs or kidney) was homogenized in 500 uL buffer (20 mmol/L HEPES (pH 7.2), 210 mmol/L mannitol, 70 mmol/L sucrose). Samples were centrifuged for 10 min at 10,000 g and 37.5 uL of carnosine substrate solution (1 mg/mL) was added to 150 uL sample supernatant. After incubation for 0, 10, 20 and 40 min at 30 °C with 40 uL sample per time point, the reaction was terminated and liberated L-histidine was derivatized by adding 110 uL o-phthaldialdehyde, trichloroacetic acid and Tris buffer (50 mmol/L; pH 7.5) at a 1:1:1 ratio. Fluorescence was then measured using a microplate reader (infinite M200, Tecan, Männedorf, Switzerland) with λEx 360 nm and λEm 460 nm. To inhibit residual renal carnosine degrading activity in
Cndp1-KO mice this assay was repeated with an additionally 0.1 mmol/L bestatin added to the carnosine substrate solution [
21].
2.3. Carnosine and Anserine Concentrations
Carnosine concentrations were measured fluorometrically using HPLC. 50 mg of frozen tissue (either brain, liver, muscle, heart, lungs or kidney) was homogenized in 500 uL buffer (20 mmol/L HEPES; pH 7.2, 210 mmol/L mannitol, 70 mmol/L sucrose) and centrifuged for 10 min at 10,000×
g. 200 uL supernatant was incubated with 50 uL of sulfosalicylic acid (or 100 uL serum with 25 uL sulfosalicylic acid) for 30 min on ice and centrifuged for 5 min at 10,000×
g. Supernatants were then diluted 1:3 with borate buffer (400 mmol/L; pH 9.5) and 180 uL of each sample was derivatized with 180 uL carbazole-9-carbonyl chloride (0.567 mg/mL in acetone) for 90 s and 108 uL of 150 mmol/L hydroxylammonium chloride with 68 mmol/L NaOH and 2% methylthioethanol (
v/
v) for 3 min, before the reaction was terminated with 252 uL of acetonitrile in acetic acid in 8:2 ratio (
v/
v). Samples were applied on a HPLC column (Jupiter column C18, 300 Å, 5 µm particle size, 250 × 4.6 mm, Phenomenex, Aschaffenburg, Germany) and carnosine and anserine were measured by fluorescence detection (RF-20A, Shimadzu, Kyoto, Japan) at λex 287 nm/λem 340 nm. The mobile phase consisted of 82% solution A (50 mmol/L acetate buffer in distilled water; pH 4.37) and 18% solution B (a mixture of acetonitrile, methanol and tetrahydrofuran 70:25:5 (
v/
v/
v)). Solutions were degassed prior to use. All samples were measured at least twice, and one sample was spiked with standards to definitively identify each analyte [
19].
2.4. Blood Glucose
Blood glucose levels of mice were determined after a fasting period of 5 h with a glucometer (Roche, Basel, Switzerland). Experiments and daily treatment were performed during “rest” period at comparable timeslots during the day. Blood was drawn by puncture of the tail vein.
2.5. Hematological Analysis
Hemoglobin (Hb) concentration, red blood cell (RBC) count and erythrocyte indexes (MCV, MCH) were measured on a IDEXX Procyte dx automated blood cell analyzer (Idexx Laboratories).
2.6. Food Intake
Food intake of mice was determined by measuring decrease in food cup weight over a time-span of 3 to 4 days. The difference was divided by the number of mice per cage (1–2 mice per cage).
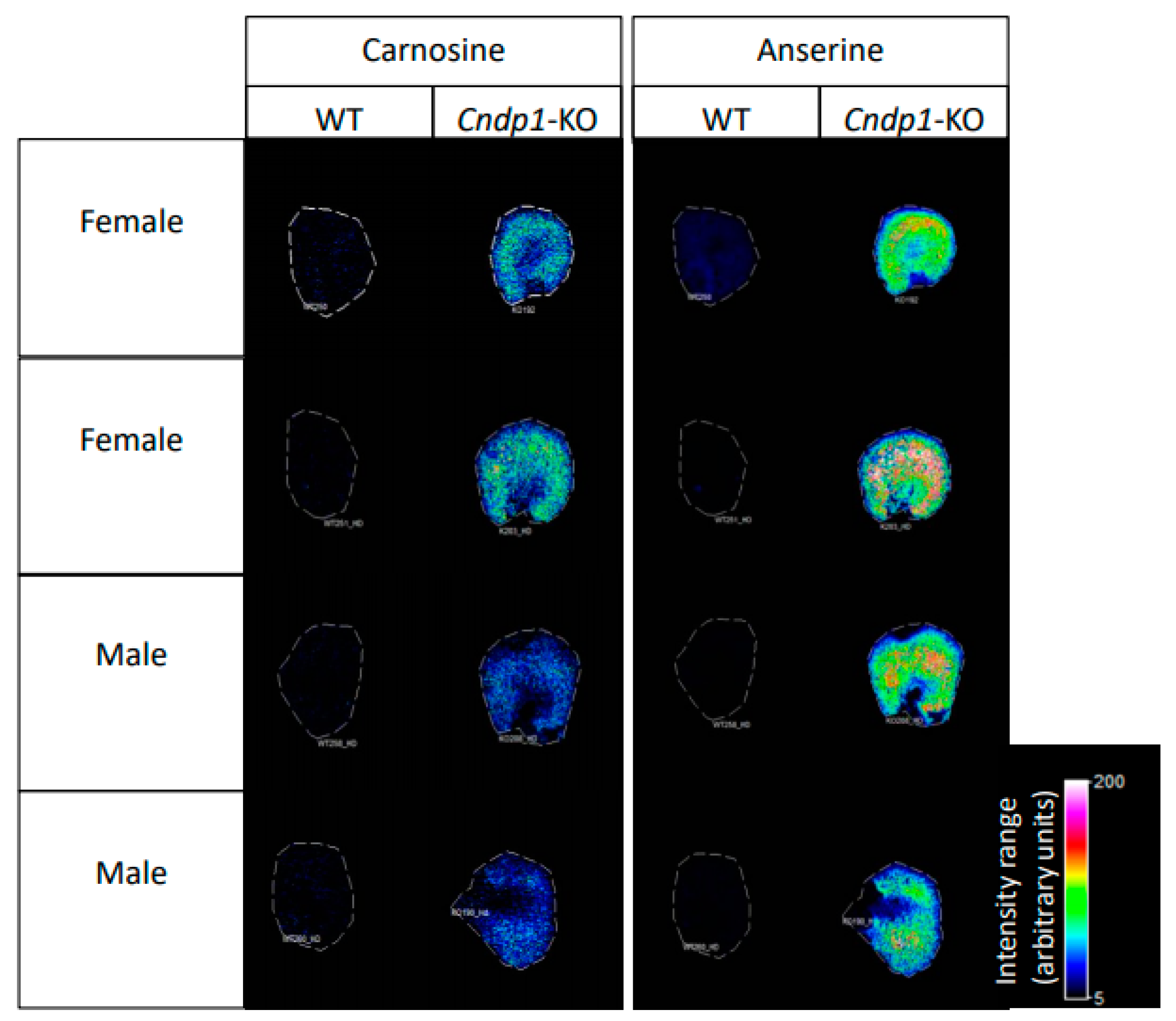
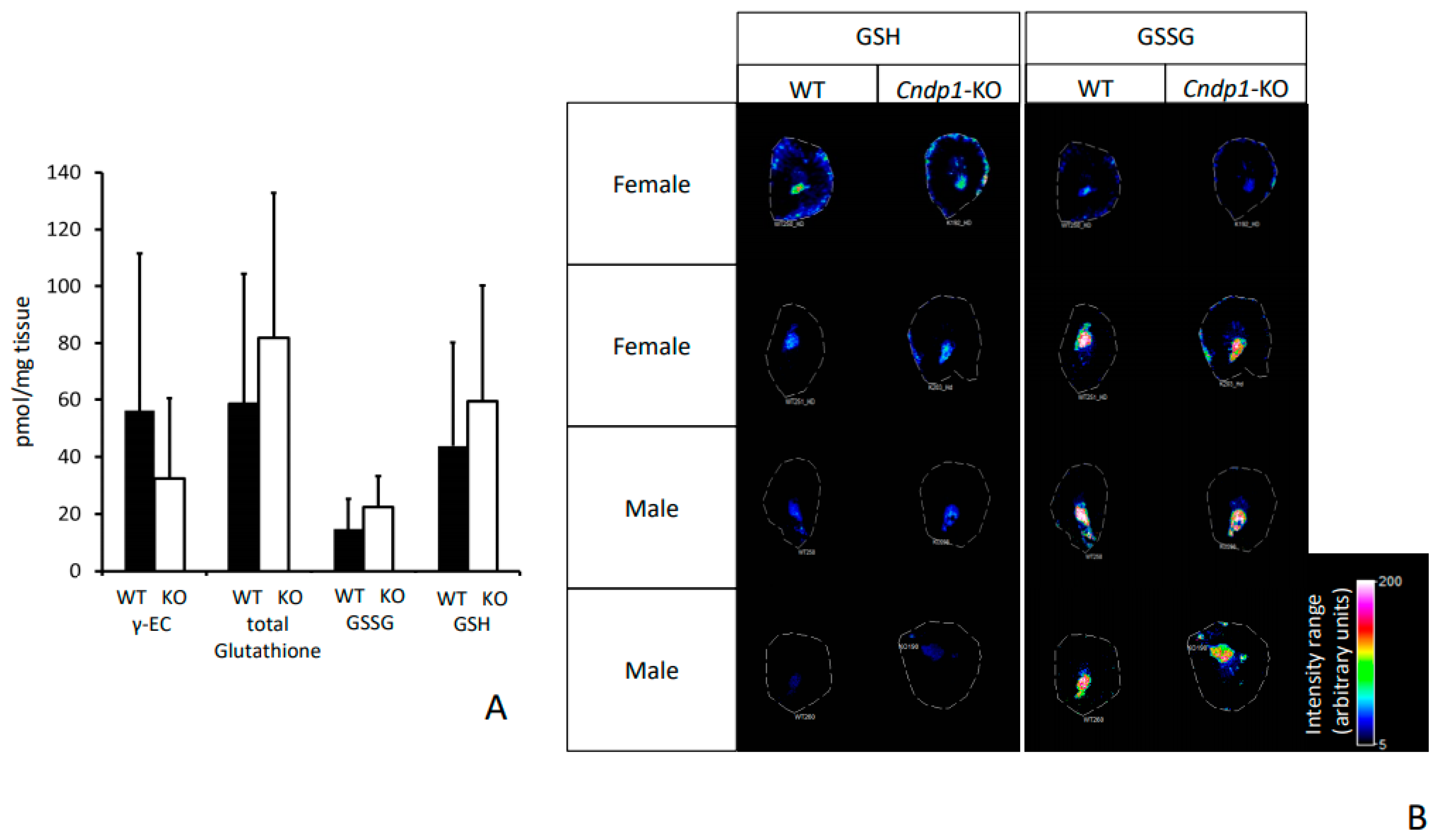
2.7. Localization of Carnosine, Anserine, GSH and GSSG
Localization of carnosine, anserine, glutathione (GSH), and glutathione disulfide (GSSG) was conducted in kidney tissue sections of 55-week-old
Cndp1-KO and wildtype mice by Mass Spectrometry Imaging (MSI). Serial sections of the investigated kidney tissue were collected on a cryomicrotome at 14 µm thickness and thaw-mounted onto indium tin oxide-coated (ITO) glass slides. The ITO slides were scanned using a flatbed scanner and dried in a desiccator until analysis. Kidney tissue sections for measurement in positive ion mode were then sprayed with DHB matrix (20 mg/mL, 60% methanol, 0.1% trifluoroacetic acid) using a TM-Sprayer (HTX Technologies, Chapel Hill, NC, USA). For measurements in negative ion mode 9-aminoacridine (9-AA) matrix (10 mg/mL in 70% ethanol) was used instead. MSI analysis was carried out using a solariX XR 7T FT-ICR mass spectrometer (Bruker Daltonik, Billerica, MA, USA). Detection of anserine and carnosine was conducted in positive ion mode using Continuous Accumulation of Selected Ions (CASI) with multiple
m/
z-windows with a width of ±5 from the theoretical
m/
z of the target compound. MALDI-MSI analysis of GSH and GSSG was performed in negative ion mode using full scan mode of the
m/
z range 100–3000.
m/
z-images were generated using flexImaging 5.0 (Bruker Daltonik, Billerica, MA, USA). To alleviate variations in pixel-to-pixel intensity, the spectrum of each pixel was normalized against the root mean square intensity of all its data points. The investigated compounds (anserine, carnosine, GSH, GSSG) were annotated on the basis of accurate mass (<0.002 Da in CASI mode and <0.005 Da in full scan mode) and by matching to a reference standard spotted on the same ITO slide off-tissue [
29].
2.8. Histological Staining
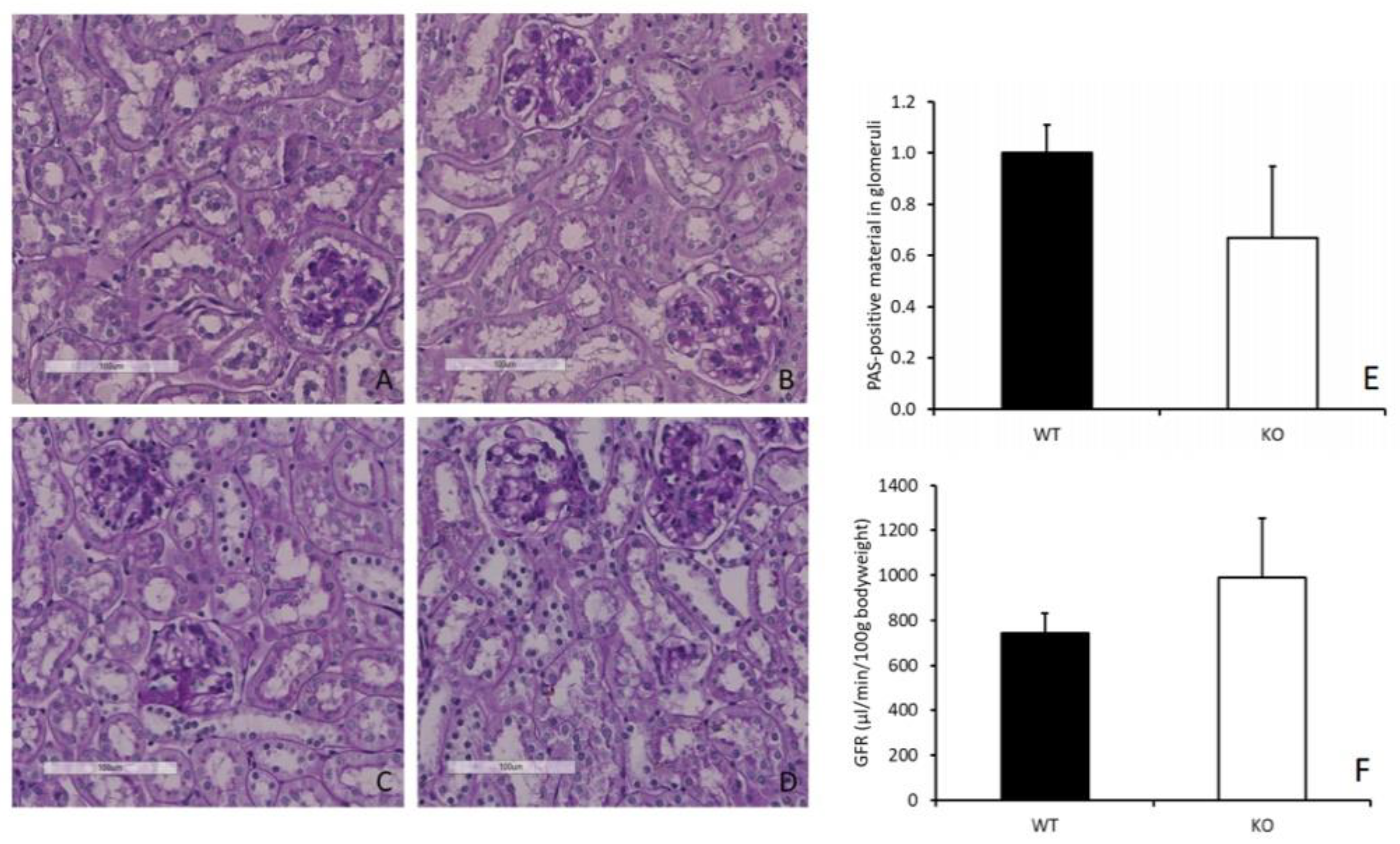
Representative formalin-fixed paraffin-embedded (FFPE) kidney and muscle specimens were cut in 4–6 µm sections using a HM 340E Rotary Microtome (Thermo Scientific Fisher, Waltham, MA, USA) and stained with hematoxylin and eosin (HE) and acid fuchsin orange G (AFOG) according to standard protocols. Additionally, a periodic-acid Schiff reaction (PAS) was performed. Images were acquired at 20-fold magnification using a Hamamatsu NanoZoomer Digital Pathology system (Hamamatsu Photonics, Hamamatsu, Japan).
2.9. Glomerular Filtration Rate
Glomerular filtration rate (GFR) was determined by transcutaneous fluorescence measurement of the clearance of fluorescein isothiocyanate (FITC)-sinistrin. One day before measurement, mice were anesthetized (initial: 5% isoflurane and 600 mL/min O2 for 1 min; following: 2.5% isoflurane and 200 mL/min O2) and the side of the back was shaved and further treated with depilation cream. The next day, mice were again anesthetized and the measuring device with a light emitting diode (λex 480 nm) for excitation of FITC and a photodiode (λem 520 nm) for fluorescence detection was attached to the shaved area. After 2 min for detecting the measurement’s baseline, 60 uL FITC-sinistrin solution (50 mg/mL in sterile saline solution; Fresenius-Kabi, Bad Homburg, Germany) was injected retro-orbitally and mice were placed back in their cages. After 2 h, measuring devices were removed and data were analyzed with MPD-Lab (Mannheim Pharma and Diagnostics, Mannheim, Germany). GFR was calculated out of the half-lifes of the FITC-sinistrin excretion in a semi-empirical approach with a one-compartment model.
2.10. Albumin Creatinine Ratios
Albumin and creatinine were determined from spot urine collections of the mice. Albumin was measured by Mouse Albumin ELISA (Alpco, Salem, MA, USA) and creatinine with Creatinine Colorimetric/Fluorometric Assay Kit (BioVision, Milpitas, CA, USA) according to the manufacturer’s manual.
2.11. Intraperitoneal Insulin Tolerance Test (IPITT) and Intraperitoneal Glucose Tolerance Test (IPGTT)
Prior to both tests, mice were fasted for 5 h and further deprived of food and water for the duration of the experiments. The animals were restrained, and tail veins were punctured. Blood glucose was measured with a glucose meter (Aviva Accu Check, Roche, Mannheim, Germany), representing baseline levels. For the insulin tolerance test, the insulin stock solution (40 U/mL; Insuman Rapid, Sanofi-Aventis, Frankfurt, Germany) was diluted to 1 U/mL in sterile saline solution. Mice were injected intraperitoneal with 1 U/kg body weight. For the glucose tolerance test, mice were injected with 2 g/kg body mass of D-glucose (20% w/v in sterile saline solution). For both experiments blood glucose was measured after 5, 10, 15, 20, 25, 30, 45, 60, 80, 100 and 120 min after injection.
2.12. Thiol- and Amino Acid Concentrations
Free amino acids and thiol compounds were extracted from 30 mg kidney tissue with 0.3 mL of 0.1 mol/L HCl in an ultrasonic ice-bath for 10 min. The resulting homogenates were centrifuged twice for 10 min at 4 °C and 16,400×
g to remove cell debris. Total glutathione was quantified by reducing disulfides with DTT followed by thiol derivatization with the fluorescent dye monobromobimane (Thiolyte, Calbiochem, Darmstadt, Germany). For quantification of GSSG, free thiols were first blocked by NEM followed by DTT reduction and monobromobimane derivatization. GSH equivalents were calculated by subtracting GSSG from total glutathione levels. Derivatization was performed as described in Wirtz et al. [
30]. For ultra performance liquid chromatography-fluorescence detector (UPLC-FLR) analysis an Acquity H-class UPLC system coupled to an Acquity FLR detector (Waters, Milford, CT, USA) was used. Separation was carried out using a binary gradient of buffer A (100 mmol/L potassium acetate, pH 5.3) and solvent B (acetonitrile) with the following gradient: 0 min 2.3% buffer B; 0.99 min 2.3%, 1 min 70%, 1.45 min 70%, and re-equilibration to 2.3% B in 1.05 min at a flow rate of 0.85 mL min
−1. The column (Acquity BEH Shield RP18 column, 50 mm x 2.1 mm, 1.7 µm, Waters, Milford, USA) was maintained at 45 °C and sample temperature was kept constant at 14 °C. Monobromobimane conjugates were detected by fluorescence at 480 nm after excitation at 380 nm. Data acquisition and processing was performed with the Empower3 software suite (Waters, Milford, CT, USA).
Determination of proteinogenic amino acid levels was done as described in Weger et al. [
31]. Beta-Alanine content was analyzed after specific labeling with the fluorescence dye AccQ-Tag™ (Waters, Milford, CT, USA) according to the manufacturer’s protocol using an Acquity I-class UPLC system coupled to a VION Ion Mobility Separation QTof (Waters, Milford, CT, USA). Separation was carried out using a Cortecs C18 column (100 mm × 2.1 mm, 1.6 µm, Waters) at 40 °C. The mobile UPLC phase consisted of binary gradients of ACN with 0.1% formic acid (B) and 0.1% aqueous formic acid (A), flowing at 0.5 mL min
−1. Analytes were initially eluted with 98% A and A was decreased linearly to 35% over 6 min. After this, the column was washed with 85% B for 2 min and re-equilibrated under the initial conditions for 2 min. Measurements were performed with an ESI source operated in positive mode (1.00 kV capillary voltage; source temperature 120 °C, desolvation temperature 500 °C; observed
m/
z 260.103 Da; observed CCS value 161.84). Unifi software (version 1.9.3, Waters, Milford, CT, USA) was used to control the instrument and to acquire and process the MS data.
2.13. Expression Analysis by qPCR
Kidney tissue was homogenized using the Ball mill MM400 from Retsch with the corresponding tissue lyser adaptor sets from QIAGEN. RNeasy Mini kit from QIAGEN was used for RNA purification. Isolated mRNA was transcribed to cDNA (High Capacity cDNA Reverse Transcription Kit, Thermo Fisher Scientific, Waltham, MA, USA). Samples were analyzed using SYBR Green (PowerUP SYBR Green Master mix, Thermo Fisher Scientific) based Quantitative RT-PCR (StepOne Plus RT-PCR System, Applied Biosystems, Foster City, CA, USA). Hypoxanthine guanine phosphoribosyl transferase (Hprt) served as house-keeping gene (
Table 1). Data were analyzed using the ddCT Method.
2.14. Protein Oxidation
To measure introduction of carbonyl groups into proteins, OxyBlot Protein Oxidation Detection Kit (Merck, Darmstadt, Germany) was used. Kidney tissue samples of
Cndp1-KO and WT mice were analyzed according to the manufacturer’s manual [
31].
2.15. Statistical Analysis
Data were obtained from at least three independent experiments and are given as mean with standard deviation (SD). Error of relative differences of means is given as standard error (SE). Relative expression changes are given with a range of fold change. Statistical analysis was performed with R, a language for computational statistics and graphics (
https://www.r-project.org). Means were compared by student’s t-test. Generalized additive mixed models (GAMM) from R package “mgcv” were used to model relative blood glucose levels over time with respect to sex (male/female) and genotype (
Cndp1-KO and wildtype). Analysis of variance (ANOVA) was used to analyze effects of sex (female, male), genotype (
Cndp1-KO and wildtype) and age (11 weeks and 55 weeks of life) on blood glucose levels after fasting. ANOVA post hoc comparisons were computed using estimated marginal means with R package “emmenas”.
p-values of <0.05 were considered significant.
4. Discussion
Carnosine scavenges carbonyls [
6,
7], inhibits glycation [
8], prevents AGE formation and has antioxidant actions. Supplementation of carnosine in diabetic rodents increases renal carnosine concentrations [
32] and consistently preserves renal morphology and function [
20]. In humans carnosinemia, due to CN1 deficiency may be a non-disease; whereas accumulation of carnosine should protect against long-term sequelae of reactive metabolites accumulating, e.g., in diabetes and chronic renal failure [
3,
20] and may prevent cancer development [
33]. Efficacy of therapeutic carnosine supplementation in humans, however, is limited by a highly active serum CN1 largely preventing major increases in serum carnosine concentrations [
27]. The human kidney expresses carnosine-synthetase and CN1 in glomeruli and tubular cells, CN1 activity is increased in diabetic kidneys [
19,
21]. Modulation of kidney carnosine metabolism to increase tissue carnosine and anserine concentrations is a promising therapeutic concept. Before aiming for such therapeutic interventions, the local and systemic metabolic consequences require detailed analyses. We therefore studied a mouse model with high CN1 activity in the WT kidney but none in the muscle, brain, liver, pancreas, lungs, heart or serum. The respective global
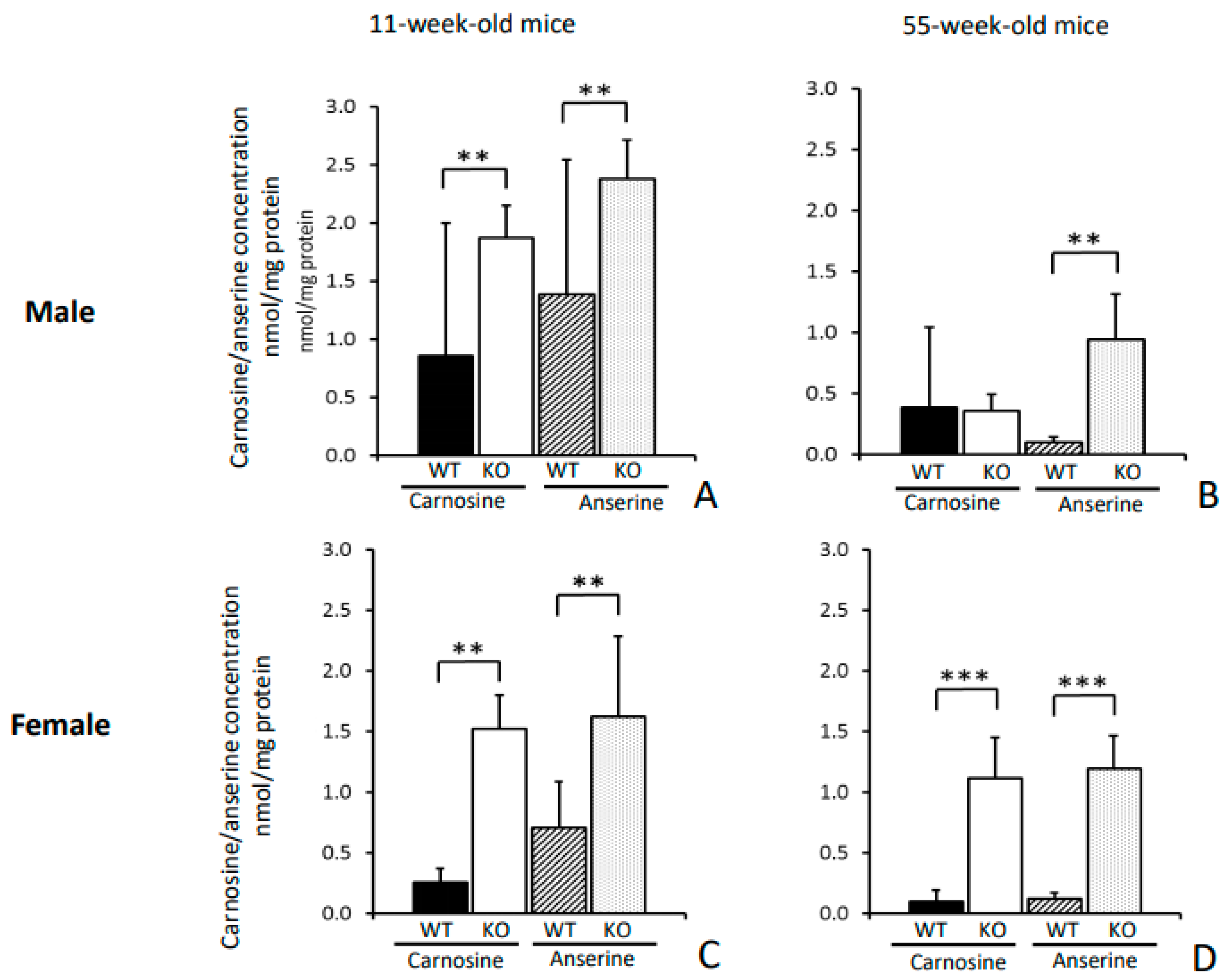
Cndp1-KO resulted in a gender-specific, selective 2- to 9-fold increase in renal carnosine and anserine concentrations, while respective concentrations were unchanged or slightly decreased in all other organs and blood. In these
Cndp1-KO mice, renal carnosine-degrading efficiency was severely reduced, indicating that CN2 hardly compensates for CN1 activity as previously discussed [
1]. Further, we confirmed previous findings [
2] that CN1 activity in this mouse model is only detectable in the kidney. Thus, this mouse model is uniquely suited to study the impact of renal histidine-dipeptides in the development and progression of renal disease.
The persistently high renal concentrations of carnosine and anserine did not result in any renal morphological and/or functional abnormalities in comparison with WT mice. Putative quenching capacity against reactive metabolites, such as acrolein and HNE [
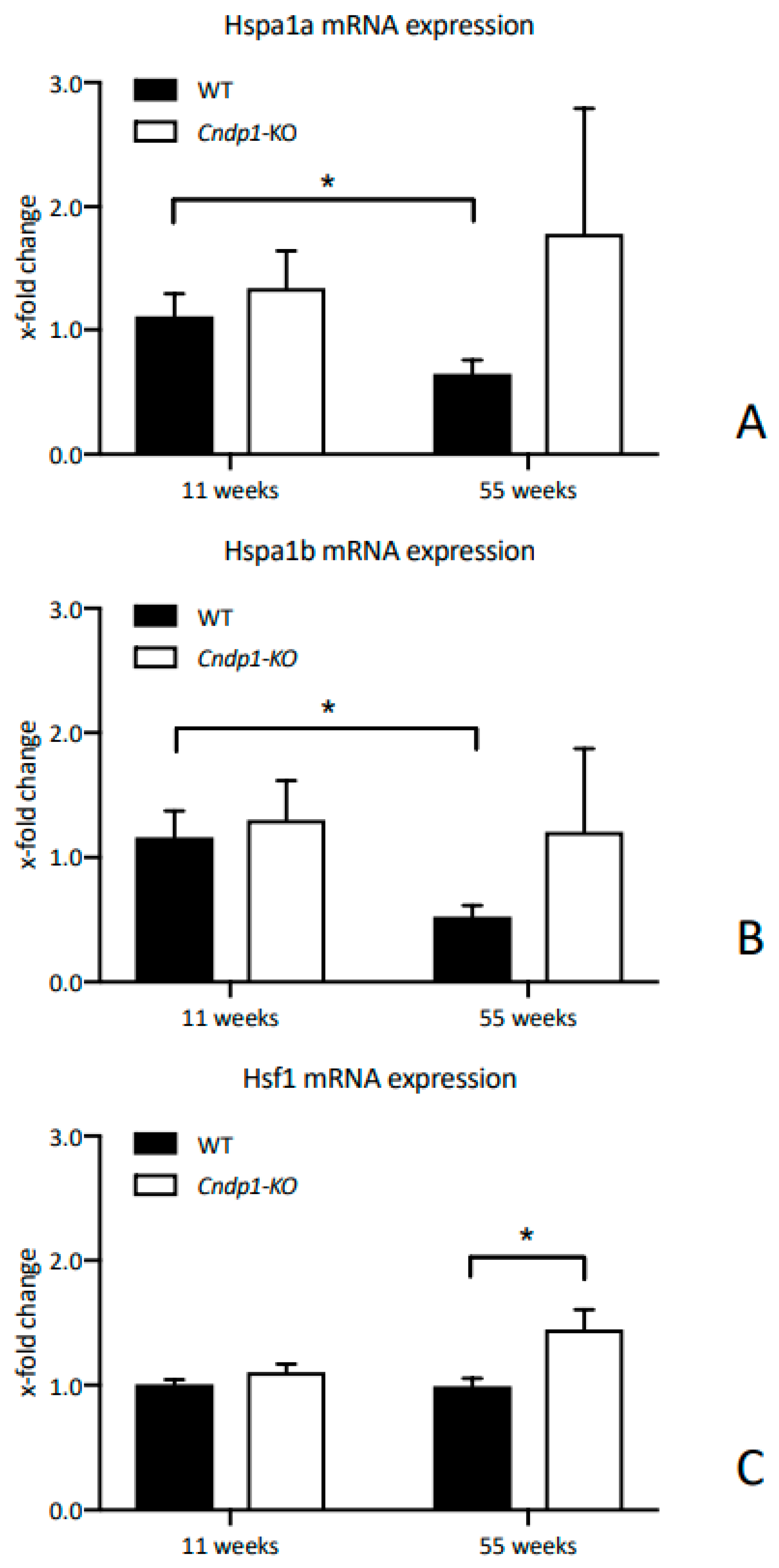
34] could not be demonstrated, the degree of carbonylation was low in all mice. Renal GSSG-to-GSH ratio and the precursor γ-EC were unaltered. Recent studies have shown that heat shock response contributes to establishing a cytoprotective state in a wide variety of human diseases, such as diabetes or cancer. At least part of the protective effect of carnosine might be related to induced expression of members of the heat shock family, such as hsp70, Hsp72 and/or hsp90 [
35,
36,
37]. Age-associated decline of renal HSP70 expression was prevented in 55-week-old
Cndp1-KO mice, Hsf1 increased. HSP70 protects against oxidative stress, participates in disposal of damaged or defective proteins, improves protein integrity and inhibits apoptosis [
38]. Hsf1 is the major transcription factor of HSPs. In human proximal tubular cells HSP70 expression is induced by oxidative and glucose stress in the presence of anserine, but not carnosine [
18]. Whether the
Cndp1-KO related changes in cellular protective mechanisms and the renal accumulation of carnosine and anserine provide a significant protection against diseases such as diabetic nephropathy, remains to be investigated in respective animal models.
In addition to the local renal metabolic changes, the
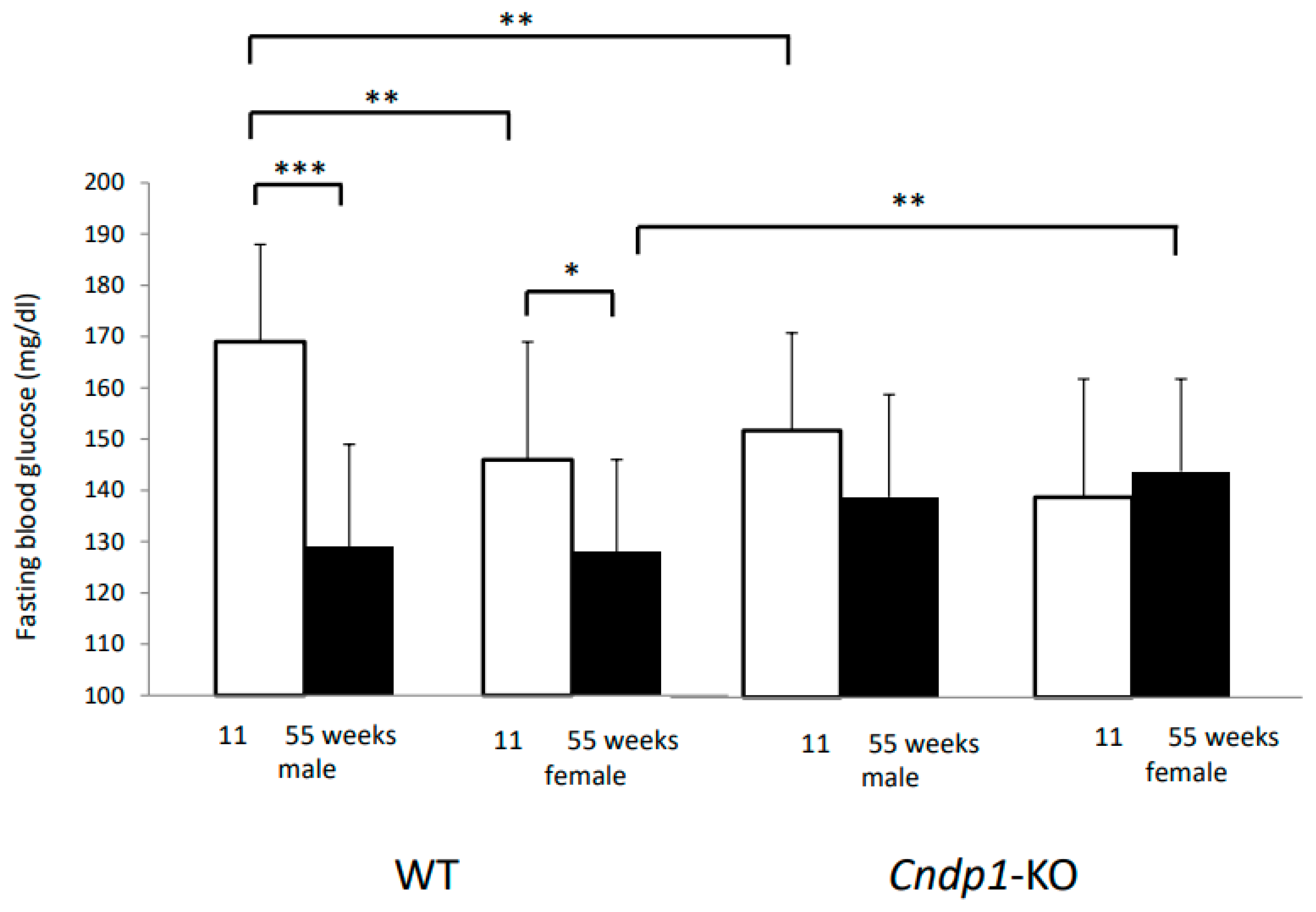
Cndp1-KO, which was functionally confined to the kidneys, resulted in modifications of systemic glucose homeostasis. While there was an age- dependent decline in fasting blood glucose concentrations in the C57BL/6J WT mice as previously described [
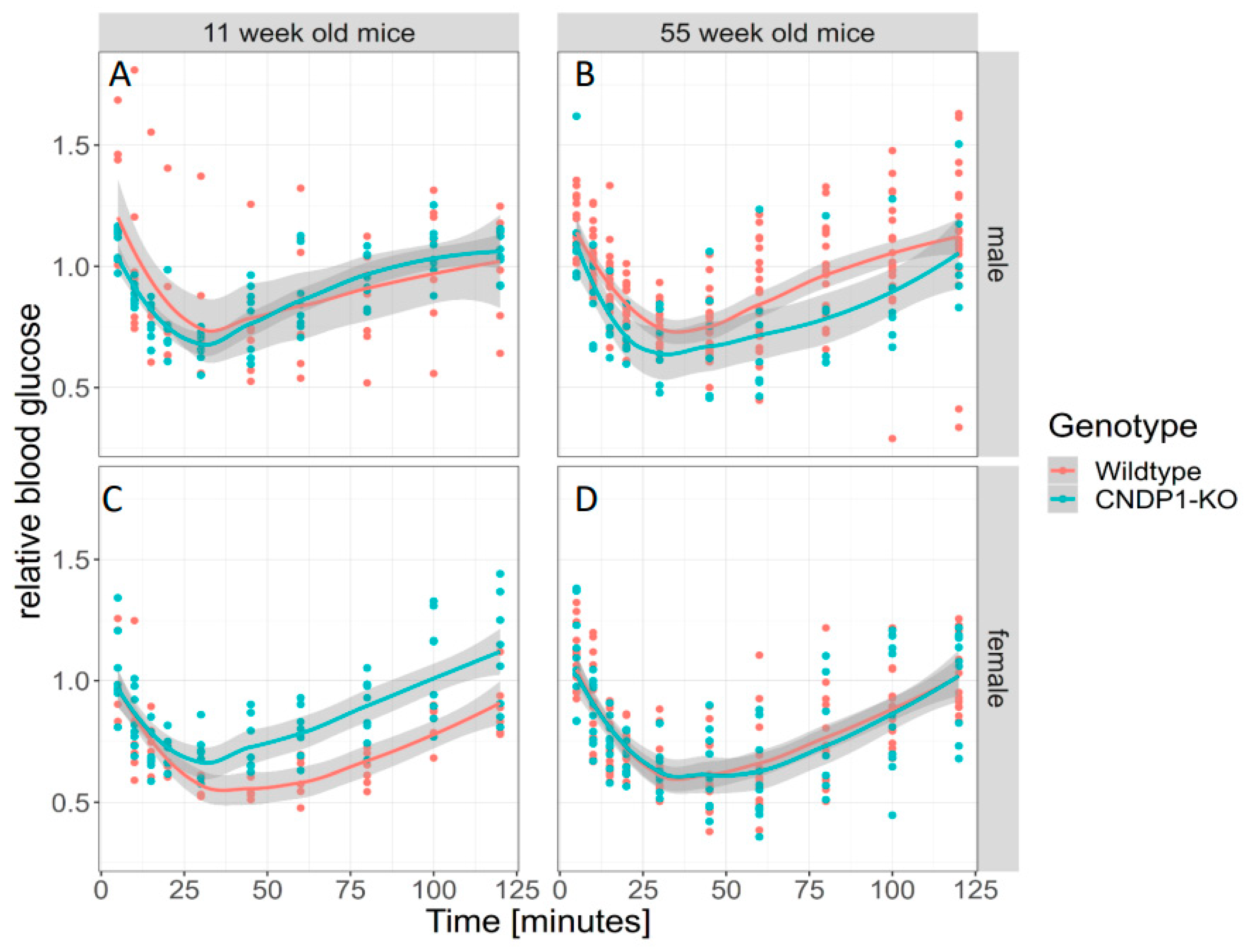
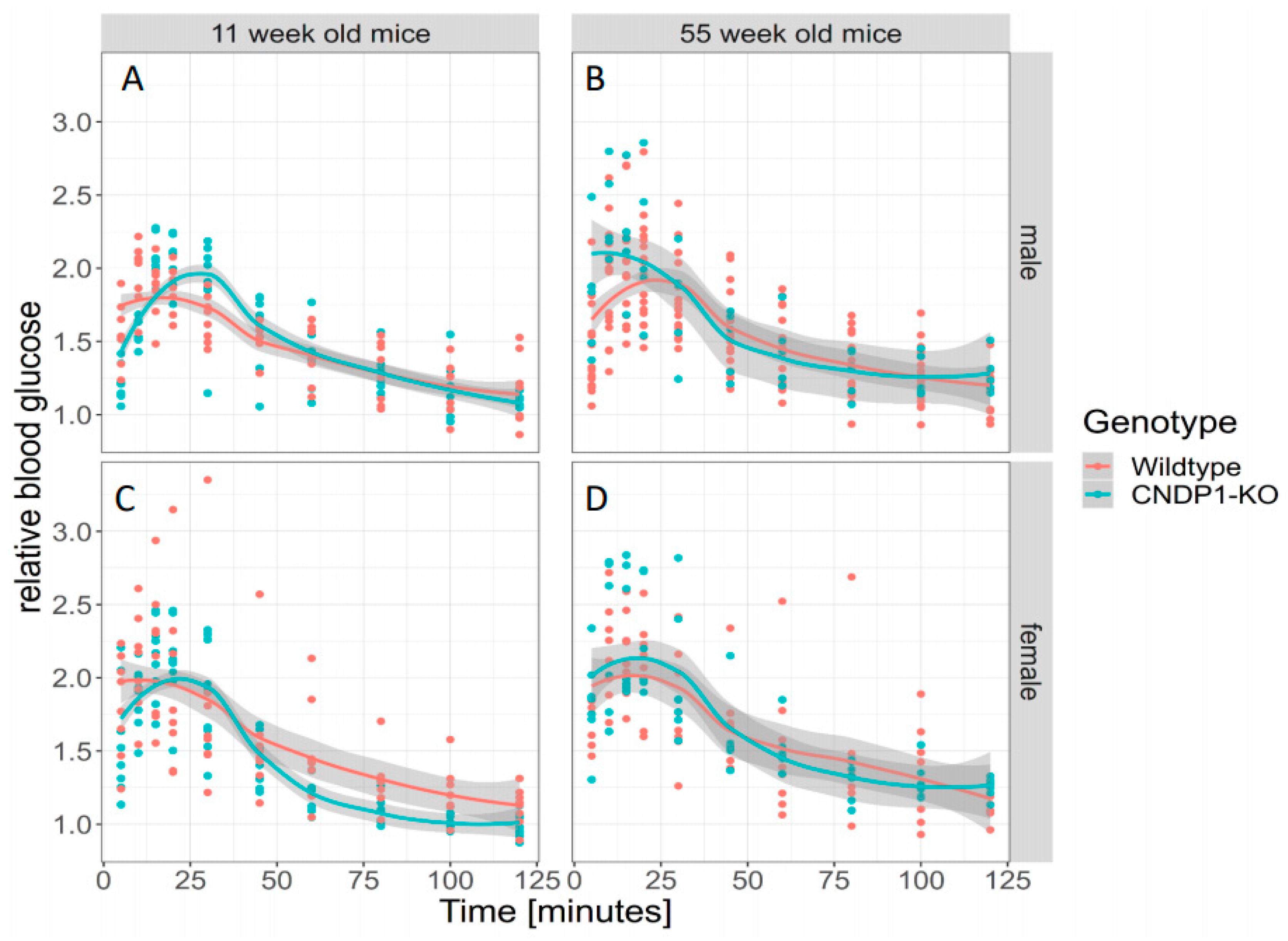
39], blood glucose remained unchanged in the respective KO mice. In line with this, the genotype predicted the response to intraperitoneal insulin injection in 55-week-old mice. However, when gender was taken into account, the observed differences were predicted by gender but not genotype. The response to intraperitoneal glucose was similar in all age and gender groups. Thus, the overall effect of
Cndp1-KO on glucose homeostasis was not pronounced; the molecular mechanisms are uncertain. Carnosine treatment of diabetic mice resulted in improved blood glucose homeostasis. In transgenic diabetic mice overexpressing human CN1 (db/db) [
23], in BTBR ob/ob mice [
24], in nephrectomized rats and in BALB/cA mice with streptozotocin (STZ)-induced diabetes type-1 [
40,
41] blood glucose and HbA1c concentrations were reduced by carnosine supply. Studies in STZ-treated mice indicated that carnosine administration modulates the insulin gene products in the pancreas, preserving insulin gene expression from STZ challenge [
42].
Cndp1 mRNA and CN1 activity could not be detected in the pancreas of our mice. Even though an impact of the proteolytic activity of the pancreatic tissue, cannot be excluded, our findings suggest that the alterations of glucose homeostasis by carnosine may be exerted via different mechanisms in the pancreas or indirectly, e.g., via metabolic effects in the kidney. Renal gluconeogenesis contributes 20–25% to whole-body glucose production [
36]. Renal glutamine concentration, an important substrate for gluconeogenesis in the kidney, were halved in our 11-week-old
Cndp1-KO mice. Thus, the renal
Cndp1-KO may have resulted in altered renal gluconeogenesis rate. As we found HSP70 mRNA levels to be preserved in kidneys of 55-week-old
Cndp1-KO mice, a preserved protein quality control system could also contribute to sustained endocrine pancreas function and insulin secretion.
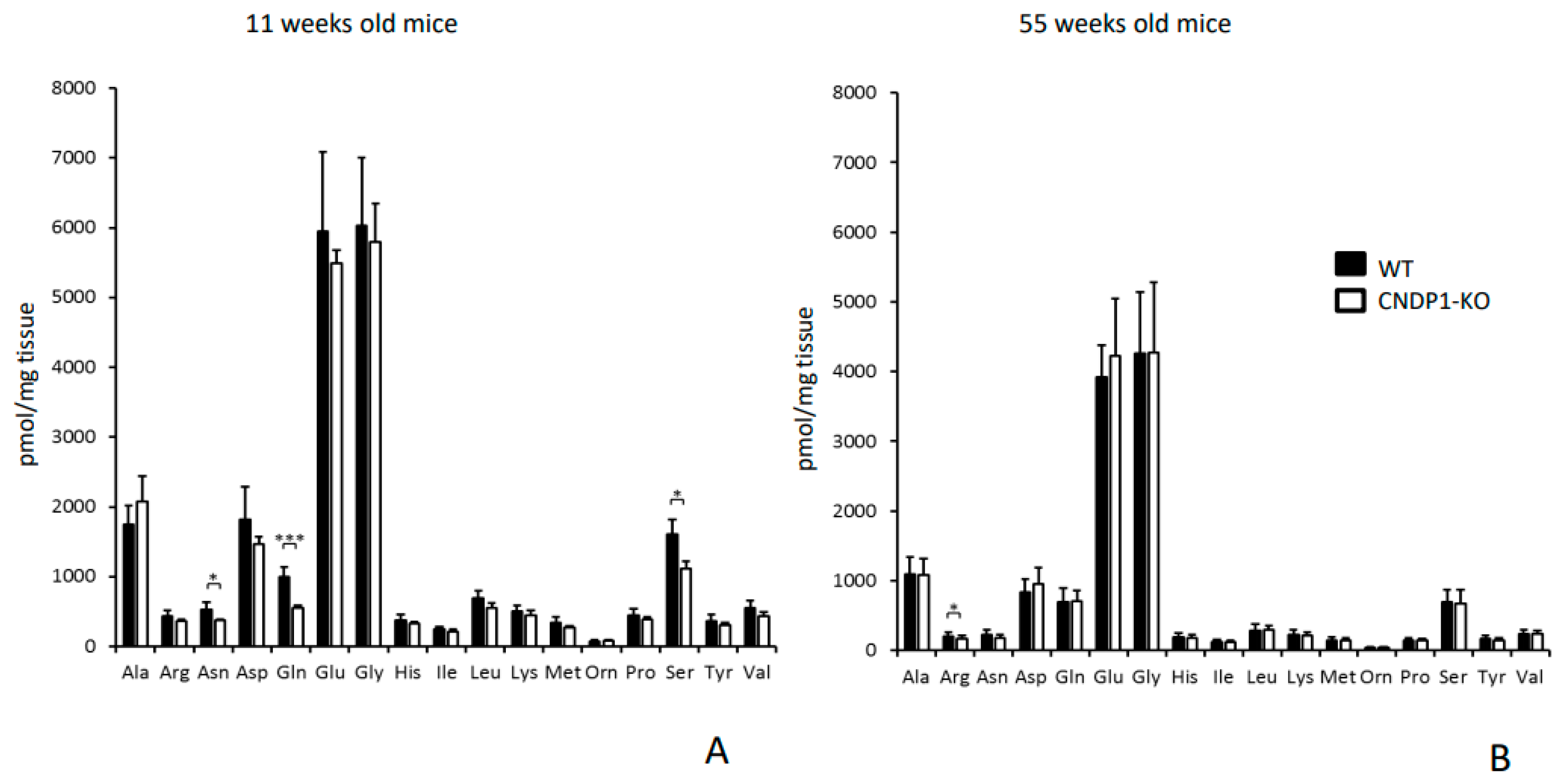
The impact of reduced renal asparagine and serine concentrations in young and of arginine in old
Cndp1-KO mice is uncertain. Asparagine synthetase is present in most mammalian organs, and is required for brain function and the synthesis of ammonia. Serine is important in the biosynthesis of purines and pyrimidines, and is a precursor to several amino acids and of metabolites such as sphingolipids and folate. Renal synthesis of serine only accounts for 5–7% of total body turnover of serine [
43]. Arginine is a conditionally indispensable amino acid, synthesized mainly in the liver and kidney and is a precursor of nitric oxide and of urea synthesis in liver and kidney. It has immunotrophic and strong insulinogenic effects [
43]. Arginine may become limiting under conditions of severe metabolic stress and the kidney may become an important arginine producer. Failure to produce sufficient quantities of arginine was reported to occur in patients with renal failure [
44]. Further studies are required, in particular in mice with renal diseases, to demonstrate whether the
Cndp1-KO associated changes in specific amino acids are of pathophysiological relevance. In the 11-week-old mice amino acid profiling was performed in 4 mice per group only, which increases the risk of a type II error, i.e., of missing differences between the two genotypes.
Higher body weight was reached in both female and male
Cndp1-KO mice until week 55 and was associated with increased absolute weights of all organs except the lung; relative to body weight spleen weight was also increased. The higher body weight gain in
Cndp1-KO mice is probably caused by higher food intake as compared to WT mice. Carnosine supplemented pigs achieve greater body weight gain together with an increased food intake [
45] and carnosine treated STZ mice experience 50% less body weight loss than STZ mice without carnosine supplementation [
42].
Cndp1-KO zebra fish have increased carnosine concentrations, and exhibit less body weight gain with high-fat diet than the respective WT fish [
46] and a combination of genetic variations in CNDP1 and CNDP2 were associated with obesity risk in Japanese men [
47]. A recent meta-analysis of 23 clinical trials suggests reduced waist circumference, HbA1c, and fasting glucose levels with histidine-dipeptide supplementation [
48].
In vitro, carnosine scavenges glucolipotoxins, palmitic and oleic acid and increases myocyte glucose uptake under hyperglycemic conditions [
48]. Thus, there is evidence from different experimental models and from clinical trials that suppression of CN1 activity with consecutive increase in carnosine tissue concentrations and carnosine supplementation, respectively, modifies food intake, glucose homeostasis and body weight gain.
In summary, a global Cndp1-KO in mice results in a selective and marked gender- and age-specific increase in renal carnosine and anserine concentrations, in higher renal Hsp70 expression and an altered renal amino acid profile. Blood glucose homeostasis and the response to intraperitoneal insulin is gender specifically modified, food intake and body weight gain increased. These findings suggest a significant local and systemic metabolic function of renal CN1. Suppression of renal CN1 activity and the subsequent great increase in renal carnosine and anserine concentrations represents a promising therapeutic approach to mitigate or prevent chronic kidney diseases such as diabetic nephropathy.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}