An Overview of Different Strategies to Recreate the Physiological Environment in Experimental Erythropoiesis

 and
and 
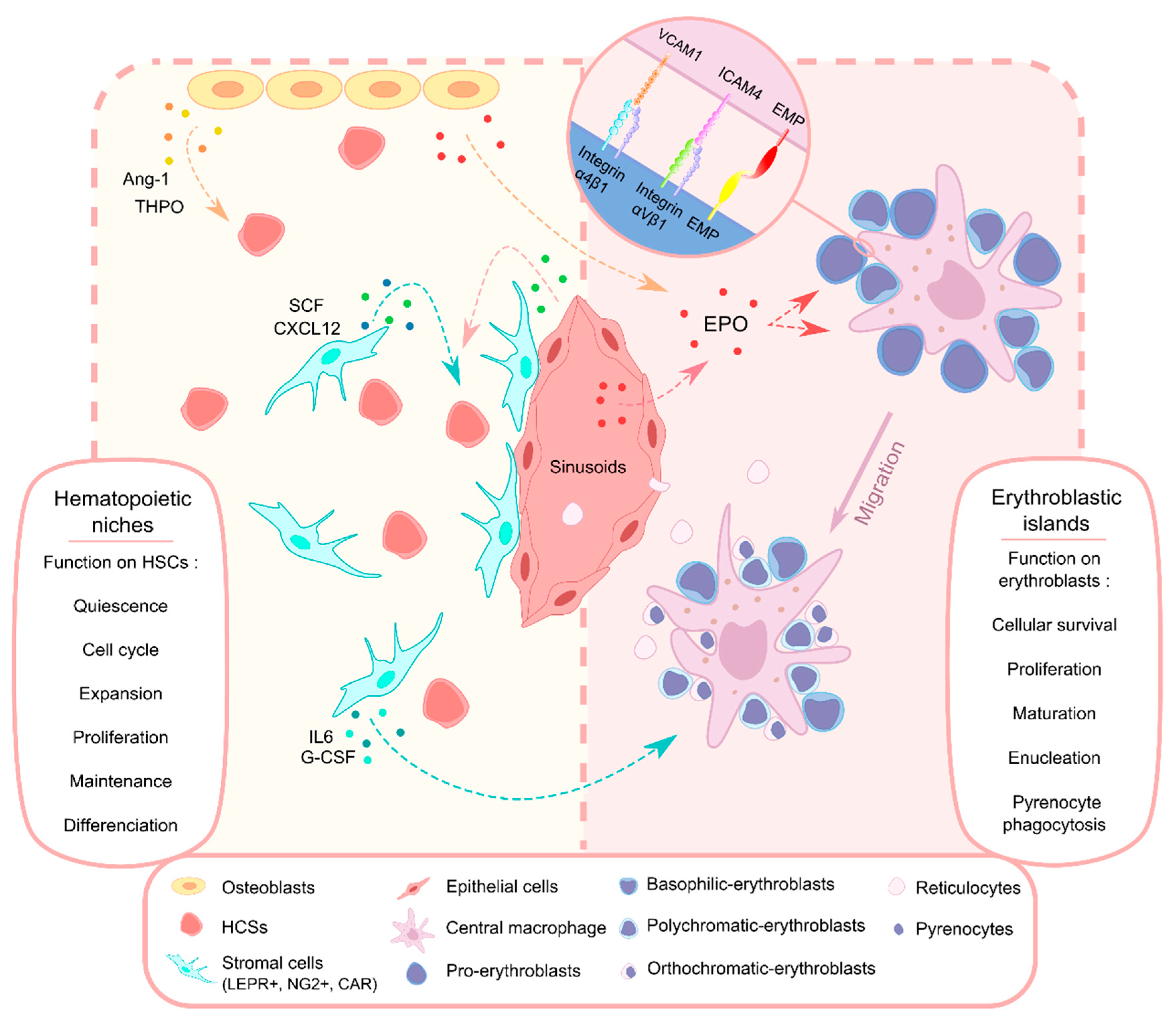{kind=link}
Abstract
:1. Introduction
2. Evolution of Experimental Approaches
2.1. Hematopoietic Stem Cells
2.2. Inducible Pluripotent Stem Cell (iPSC)
2.3. Mice Models
3. Bone Marrow and Microenvironments
3.1. HSCs Localization and Mesenchymal Stromal Cells
3.2. Osteoblasts
3.3. Extracellular Matrix
4. Erythroblastic Islands
4.1. Localization and Characterization
4.2. Function
4.3. Co-Culture
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ang-1 | Antiopoietin-1 |
| Baso-E | Basophilic erythroblast |
| BFU-E | Burst forming unit erythroid |
| BM | Bone marrow |
| BMSC | Bone marrow stromal cell |
| CAR | Contractile actin ring |
| CB | Coord blood |
| CFU-E | Colony forming unit erythroid |
| EBI | Erythroblastic island |
| ECM | Extracellular matrix |
| EMP | Erythroblast macrophage protein |
| EPO | Erythropoietin |
| ESC | Embryonic stem cell |
| GMP | Good manufacturing practice |
| GPA | Glycophorin A |
| HbF | Fetal hemoglobin |
| hESC | Human embryonic stem cell |
| HSC | Hematopoietic stem cell |
| iPSC | Inducible pluripotent stem cell |
| KLF1 | Erythroid Kruppel-like factor 1 |
| MAEA | Macrophage erythroblast attacher |
| MDS | Myelodysplastic Syndrome |
| mFLSC | Mouse fetal liver-delivered stromal cells |
| MSC | Mesenchymal stromal cell |
| Ortho-E | Orthochromatic erytroblast |
| PB | Peripheral blood |
| PBMC | Peripheral blood mononuclear cell |
| Poly-E | Polychromatic erythroblast |
| RBCs | Red blood cells |
| SCF | Stem cell factor |
| THPO | Thrombopoietin |
References
- Dzierzak, E.; Philipsen, S. Erythropoiesis: Development and differentiation. Cold Spring Harb. Perspect. Med. 2013, 3, a011601. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, S.K.; Ulirsch, J.C.; Sankaran, V.G. Advances in understanding erythropoiesis: Evolving perspectives. Br. J. Haematol. 2016, 173, 206–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, S.; Bala, S.; Gwynn, B.; Sahr, K.E.; Peters, L.L.; Hanspal, M. Absence of erythroblast macrophage protein (Emp) leads to failure of erythroblast nuclear extrusion. J. Biol. Chem. 2006, 281, 20181–20189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Huggins, M.; Ahmed, J.; Hashimoto, D.; Lucas, D.; Kunisaki, Y.; Pinho, S.; Leboeuf, M.; Noizat, C.; van Rooijen, N.; et al. CD169(+) macrophages provide a niche promoting erythropoiesis under homeostasis and stress. Nat. Med. 2013, 19, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Palis, J. Primitive and definitive erythropoiesis in mammals. Front. Physiol. 2014, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ji, P. New insights into the mechanisms of mammalian erythroid chromatin condensation and enucleation. Int. Rev. Cell Mol. Biol. 2015, 316, 159–182. [Google Scholar] [CrossRef]
- Bejar, R.; Levine, R.; Ebert, B.L. Unraveling the molecular pathophysiology of myelodysplastic syndromes. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 504–515. [Google Scholar] [CrossRef]
- Kato, G.J.; Steinberg, M.H.; Gladwin, M.T. Intravascular hemolysis and the pathophysiology of sickle cell disease. J. Clin. Investig. 2017, 127, 750–760. [Google Scholar] [CrossRef]
- Taher, A.T.; Weatherall, D.J.; Cappellini, M.D. Thalassaemia. Lancet 2018, 391, 155–167. [Google Scholar] [CrossRef]
- Gaudio, A.; Xourafa, A.; Rapisarda, R.; Zanoli, L.; Signorelli, S.S.; Castellino, P. Hematological Diseases and Osteoporosis. Int. J. Mol. Sci. 2020, 21, 3538. [Google Scholar] [CrossRef]
- Nilsson, L.; Astrand-Grundström, I.; Anderson, K.; Arvidsson, I.; Hokland, P.; Bryder, D.; Kjeldsen, L.; Johansson, B.; Hellström-Lindberg, E.; Hast, R.; et al. Involvement and functional impairment of the CD34(+)CD38(-)Thy-1(+) hematopoietic stem cell pool in myelodysplastic syndromes with trisomy 8. Blood 2002, 100, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Yang, L.; Guo, L.; Cui, N.; Zhang, G.; Liu, C.; Xing, L.; Shao, Z.; Wang, H. Impaired Mitophagy of Nucleated Erythroid Cells Leads to Anemia in Patients with Myelodysplastic Syndromes. Oxid. Med. Cell. Longev. 2018, 2018, 6328051. [Google Scholar] [CrossRef] [PubMed]
- Taher, A.T.; Cappellini, M.D. How I manage medical complications of beta-thalassemia in adults. Blood 2018, 132, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Peng, Y.; Liu, J. Research advances in erythrocyte regeneration sources and methods in vitro. Cell Regen. 2018, 7, 45–49. [Google Scholar] [CrossRef]
- Fibach, E.; Manor, D.; Oppenheim, A.; Rachmilewitz, E.A. Proliferation and maturation of human erythroid progenitors in liquid culture. Blood 1989, 73, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, G.; Di Pietro, R.; di Giacomo, V.; Di Baldassarre, A.; Migliaccio, A.R.; Maccioni, L.; Galanello, R.; Papayannopoulou, T. In vitro mass production of human erythroid cells from the blood of normal donors and of thalassemic patients. Blood Cells Mol. Dis. 2002, 28, 169–180. [Google Scholar] [CrossRef]
- Tanavde, V.M.; Malehorn, M.T.; Lumkul, R.; Gao, Z.; Wingard, J.; Garrett, E.S.; Civin, C.I. Human stem-progenitor cells from neonatal cord blood have greater hematopoietic expansion capacity than those from mobilized adult blood. Exp. Hematol. 2002, 30, 816–823. [Google Scholar] [CrossRef]
- Giarratana, M.-C.; Kobari, L.; Lapillonne, H.; Chalmers, D.; Kiger, L.; Cynober, T.; Marden, M.C.; Wajcman, H.; Douay, L. Ex vivo generation of fully mature human red blood cells from hematopoietic stem cells. Nat. Biotechnol. 2005, 23, 69–74. [Google Scholar] [CrossRef]
- Wilson, M.C.; Trakarnsanga, K.; Heesom, K.J.; Cogan, N.; Green, C.; Toye, A.M.; Parsons, S.F.; Anstee, D.J.; Frayne, J. Comparison of the Proteome of Adult and Cord Erythroid Cells, and Changes in the Proteome Following Reticulocyte Maturation. Mol. Cell. Proteom. 2016, 15, 1938–1946. [Google Scholar] [CrossRef] [Green Version]
- Heshusius, S.; Heideveld, E.; Burger, P.; Thiel-Valkhof, M.; Sellink, E.; Varga, E.; Ovchynnikova, E.; Visser, A.; Martens, J.H.A.; von Lindern, M.; et al. Large-scale in vitro production of red blood cells from human peripheral blood mononuclear cells. Blood Adv. 2019, 3, 3337–3350. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, G.; Sanchez, M.; Masiello, F.; Tirelli, V.; Varricchio, L.; Whitsett, C.; Migliaccio, A.R. Humanized culture medium for clinical expansion of human erythroblasts. Cell Transplant. 2010, 19, 453–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, M.H. Concise Review: Early Embryonic Erythropoiesis: Not so Primitive After All. Stem Cells 2013, 31, 849–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.S.; Zhu, Y.; Deng, Q.; Chin, C.J.; He, C.B.; Grieco, A.J.; Dravid, G.G.; Parekh, C.; Hollis, R.P.; Lane, T.F.; et al. Erythropoiesis from human embryonic stem cells through erythropoietin-independent AKT signaling. Stem Cells 2014, 32, 1503–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Lapillonne, H.; Kobari, L.; Mazurier, C.; Tropel, P.; Giarratana, M.-C.; Zanella-Cleon, I.; Kiger, L.; Wattenhofer-Donze, M.; Puccio, H.; Hebert, N.; et al. Red blood cell generation from human induced pluripotent stem cells: Perspectives for transfusion medicine. Haematologica 2010, 95, 1651–1659. [Google Scholar] [CrossRef] [Green Version]
- Kobari, L.; Yates, F.; Oudrhiri, N.; Francina, A.; Kiger, L.; Mazurier, C.; Rouzbeh, S.; El-Nemer, W.; Hebert, N.; Giarratana, M.-C.; et al. Human induced pluripotent stem cells can reach complete terminal maturation: In vivo and in vitro evidence in the erythropoietic differentiation model. Haematologica 2012, 97, 1795–1803. [Google Scholar] [CrossRef]
- Huang, X.; Shah, S.; Wang, J.; Ye, Z.; Dowey, S.N.; Tsang, K.M.; Mendelsohn, L.G.; Kato, G.J.; Kickler, T.S.; Cheng, L. Extensive ex vivo expansion of functional human erythroid precursors established from umbilical cord blood cells by defined factors. Mol. Ther. 2014, 22, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.B.; Sommer, A.G.; Leung, A.; Park, S.; Chui, D.H.K.; Steinberg, M.H.; Mostoslavsky, G.; Murphy, G.J. Multisystem, Induced Pluripotent Stem Cell (iPSC) Modeling Reveals a Role for Growth Differentiation Factors (GDFs) in the Etiology of β Thalassemia and Ineffective Erythropoiesis. Blood 2014, 124, 4039. [Google Scholar] [CrossRef]
- Hansen, M.; Varga, E.; Aarts, C.; Wust, T.; Kuijpers, T. E ffi cient production of erythroid, megakaryocytic and myeloid cells, using single cell-derived iPSC colony di ff erentiation. Stem Cell Res. 2018, 29, 232–244. [Google Scholar] [CrossRef]
- Fujita, A.; Uchida, N.; Haro-Mora, J.J.; Winkler, T.; Tisdale, J. β-Globin-Expressing Definitive Erythroid Progenitor Cells Generated from Embryonic and Induced Pluripotent Stem Cell-Derived Sacs. Stem Cells 2016, 34, 1541–1552. [Google Scholar] [CrossRef] [Green Version]
- Uchida, N.; Haro-Mora, J.J.; Fujita, A.; Lee, D.-Y.; Winkler, T.; Hsieh, M.M.; Tisdale, J.F. Efficient Generation of beta-Globin-Expressing Erythroid Cells Using Stromal Cell-Derived Induced Pluripotent Stem Cells from Patients with Sickle Cell Disease. Stem Cells 2017, 35, 586–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-T.; Ma, R.; Axton, R.A.; Jackson, M.; Taylor, A.H.; Fidanza, A.; Marenah, L.; Frayne, J.; Mountford, J.C.; Forrester, L.M. Activation of KLF1 Enhances the Differentiation and Maturation of Red Blood Cells from Human Pluripotent Stem Cells. Stem Cells 2017, 35, 886–897. [Google Scholar] [CrossRef] [Green Version]
- Barbarani, G.; Fugazza, C.; Strouboulis, J.; Ronchi, A.E. The Pleiotropic Effects of GATA1 and KLF1 in Physiological Erythropoiesis and in Dyserythropoietic Disorders. Front. Physiol. 2019, 10, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pishesha, N.; Thiru, P.; Shi, J.; Eng, J.C.; Sankaran, V.G.; Lodish, H.F. Transcriptional divergence and conservation of human and mouse erythropoiesis. Proc. Natl. Acad. Sci. USA 2014, 111, 4103–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, X.; Schulz, V.P.; Mohandas, N.; Gallagher, P.G. Human and murine erythropoiesis. Curr. Opin. Hematol. 2015, 22, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Neildez-Nguyen, T.M.A.; Wajcman, H.; Marden, M.C.; Bensidhoum, M.; Moncollin, V.; Giarratana, M.-C.; Kobari, L.; Thierry, D.; Douay, L. Human erythroid cells produced ex vivo at large scale differentiate into red blood cells in vivo. Nat. Biotechnol. 2002, 20, 467–472. [Google Scholar] [CrossRef]
- Hu, Z.; Van Rooijen, N.; Yang, Y.-G. Macrophages prevent human red blood cell reconstitution in immunodeficient mice. Blood 2011, 118, 5938–5946. [Google Scholar] [CrossRef] [Green Version]
- Rahmig, S.; Kronstein-Wiedemann, R.; Fohgrub, J.; Kronstein, N.; Nevmerzhitskaya, A.; Bornhäuser, M.; Gassmann, M.; Platz, A.; Ordemann, R.; Tonn, T.; et al. Improved Human Erythropoiesis and Platelet Formation in Humanized NSGW41 Mice. Stem Cell Rep. 2016, 7, 591–601. [Google Scholar] [CrossRef] [Green Version]
- Sharma, Y.; Astle, C.M.; Harrison, D.E. Heterozygous kit mutants with little or no apparent anemia exhibit large defects in overall hematopoietic stem cell function. Exp. Hematol. 2007, 35, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Cosgun, K.N.; Rahmig, S.; Mende, N.; Reinke, S.; Hauber, I.; Schafer, C.; Petzold, A.; Weisbach, H.; Heidkamp, G.; Purbojo, A.; et al. Kit regulates HSC engraftment across the human-mouse species barrier. Cell Stem Cell 2014, 15, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, C.; Abdulhay, N.J.; McFarland, S.K.; Munschauer, M.; Ulirsch, J.C.; Chiarle, R.; Sankaran, V.G. Developmentally-faithful and effective human erythropoiesis in immunodeficient and Kit mutant mice. Am. J. Hematol. 2017, 92, E513–E519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, A.; Yapundich, M.; Nassehi, T.; Gamer, J.; Drysdale, C.M.; Haro-mora, J.J.; Demirci, S.; Hsieh, M.M.; Uchida, N.; Tisdale, J.F. Low-Dose Busulfan Reduces Human CD34 + Cell Doses Required for Engraftment in c-kit Mutant Immunodeficient Mice. Mol. Ther. Methods Clin. Dev. 2019, 15, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, G.M.; Jeffery, E.; Morrison, S.J. Adult haematopoietic stem cell niches. Nat. Publ. Gr. 2017, 17, 573–590. [Google Scholar] [CrossRef] [PubMed]
- Kobolak, J.; Dinnyes, A.; Memic, A.; Khademhosseini, A.; Mobasheri, A. Mesenchymal stem cells: Identification, phenotypic characterization, biological properties and potential for regenerative medicine through biomaterial micro-engineering of their niche. Methods 2016, 99, 62–68. [Google Scholar] [CrossRef]
- Gomes, A.C.; Gomes, M.S. Hematopoietic niches, erythropoiesis and anemia of chronic infection. Exp. Hematol. 2016, 44, 85–91. [Google Scholar] [CrossRef]
- Kiel, M.J.; Yilmaz, O.H.; Iwashita, T.; Yilmaz, O.H.; Terhorst, C.; Morrison, S.J. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell 2005, 121, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Nombela-Arrieta, C.; Pivarnik, G.; Winkel, B.; Canty, K.J.; Harley, B.; Mahoney, J.E.; Park, S.-Y.; Lu, J.; Protopopov, A.; Silberstein, L.E. Quantitative imaging of haematopoietic stem and progenitor cell localization and hypoxic status in the bone marrow microenvironment. Nat. Cell Biol. 2013, 15, 533–543. [Google Scholar] [CrossRef]
- Itkin, T.; Gur-Cohen, S.; Spencer, J.A.; Schajnovitz, A.; Ramasamy, S.K.; Kusumbe, A.P.; Ledergor, G.; Jung, Y.; Milo, I.; Poulos, M.G.; et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 2016, 532, 323–328. [Google Scholar] [CrossRef]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Ema, H.; Karlsson, G.; Yamaguchi, T.; Miyoshi, H.; Shioda, S.; Taketo, M.M.; Karlsson, S.; Iwama, A.; Nakauchi, H. Nonmyelinating Schwann cells maintain hematopoietic stem cell hibernation in the bone marrow niche. Cell 2011, 147, 1146–1158. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Saunders, T.L.; Enikolopov, G.; Morrison, S.J. Endothelial and perivascular cells maintain haematopoietic stem cells. Nature 2012, 481, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenbaum, A.; Hsu, Y.-M.S.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Nagasawa, T.; Link, D.C. CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 2013, 495, 227–230. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Morrison, S.J. Haematopoietic stem cells and early lymphoid progenitors occupy distinct bone marrow niches. Nature 2013, 495, 231–235. [Google Scholar] [CrossRef]
- Asada, N.; Kunisaki, Y.; Pierce, H.; Wang, Z.; Fernandez, N.F.; Birbrair, A.; Ma’ayan, A.; Frenette, P.S. Differential cytokine contributions of perivascular haematopoietic stem cell niches. Nat. Cell Biol. 2017, 19, 214–223. [Google Scholar] [CrossRef] [Green Version]
- Jing, D.; Fonseca, A.-V.; Alakel, N.; Fierro, F.A.; Muller, K.; Bornhauser, M.; Ehninger, G.; Corbeil, D.; Ordemann, R. Hematopoietic stem cells in co-culture with mesenchymal stromal cells--modeling the niche compartments in vitro. Haematologica 2010, 95, 542–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, D.S.; Hanson, E.T.; Lewis, R.L.; Auerbach, R.; Thomson, J.A. Hematopoietic colony-forming cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 10716–10721. [Google Scholar] [CrossRef] [Green Version]
- Vodyanik, M.A.; Bork, J.A.; Thomson, J.A.; Slukvin, I.I. Human embryonic stem cell-derived CD34+ cells: Efficient production in the coculture with OP9 stromal cells and analysis of lymphohematopoietic potential. Blood 2005, 105, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Ebihara, Y.; Umeda, K.; Sakai, H.; Hanada, S.; Zhang, H.; Zaike, Y.; Tsuchida, E.; Nakahata, T.; Nakauchi, H.; et al. Generation of functional erythrocytes from human embryonic stem cell-derived definitive hematopoiesis. Proc. Natl. Acad. Sci. USA 2008, 105, 13087–13092. [Google Scholar] [CrossRef] [Green Version]
- Lazar-Karsten, P.; Dorn, I.; Meyer, G.; Lindner, U.; Driller, B.; Schlenke, P. The influence of extracellular matrix proteins and mesenchymal stem cells on erythropoietic cell maturation. Vox Sang. 2011, 101, 65–76. [Google Scholar] [CrossRef]
- Chen, T.-L.; Chiang, Y.-W.; Lin, G.-L.; Chang, H.-H.; Lien, T.-S.; Sheh, M.-H.; Sun, D.-S. Different effects of granulocyte colony-stimulating factor and erythropoietin on erythropoiesis. Stem Cell Res. Ther. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, J.; Gumenyuk, M.; Kang, H.; Vodyanik, M.; Yu, J.; Thomson, J.A.; Slukvin, I.I. Generation of red blood cells from human induced pluripotent stem cells. Stem Cells Dev. 2011, 20, 1639–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-T.; French, A.; Goh, P.A.; Pagnamenta, A.; Mettananda, S.; Taylor, J.; Knight, S.; Nathwani, A.; Roberts, D.J.; Watt, S.M.; et al. Human induced pluripotent stem cell derived erythroblasts can undergo definitive erythropoiesis and co-express gamma and beta globins. Br. J. Haematol. 2014, 166, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omatsu, Y.; Sugiyama, T.; Kohara, H.; Kondoh, G.; Fujii, N.; Kohno, K. Article The Essential Functions of Adipo-osteogenic Progenitors as the Hematopoietic Stem and Progenitor Cell Niche. Immunity 2010, 33, 387–399. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; von der Mark, K.; Henry, S.; Norton, W.; Adams, H.; de Crombrugghe, B. Chondrocytes transdifferentiate into osteoblasts in endochondral bone during development, postnatal growth and fracture healing in mice. PLoS Genet. 2014, 10, e1004820. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.K.F.; Chen, C.-C.; Luppen, C.A.; Kim, J.-B.; DeBoer, A.T.; Wei, K.; Helms, J.A.; Kuo, C.J.; Kraft, D.L.; Weissman, I.L. Endochondral ossification is required for haematopoietic stem-cell niche formation. Nature 2009, 457, 490–494. [Google Scholar] [CrossRef]
- Seike, M.; Omatsu, Y.; Watanabe, H.; Kondoh, G.; Nagasawa, T. Stem cell niche-specific Ebf3 maintains the bone marrow cavity. Genes Dev. 2018, 32, 359–372. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, B.; Wang, X.; Xiao, R.; Wang, C. Three-dimensional co-culture of mesenchymal stromal cells and differentiated osteoblasts on human bio-derived bone scaffolds supports active multi-lineage hematopoiesis in vitro: Functional implication of the biomimetic HSC niche. Int. J. Mol. Med. 2016, 38, 1141–1151. [Google Scholar] [CrossRef] [Green Version]
- Arai, F.; Hirao, A.; Ohmura, M.; Sato, H.; Matsuoka, S.; Takubo, K.; Ito, K.; Koh, G.Y.; Suda, T. Tie2/Angiopoietin-1 Signaling Regulates Hematopoietic Stem Cell Quiescence in the Bone Marrow Niche. Cell 2004, 118, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Kasama, T.; Isozaki, T.; Odai, T.; Matsunawa, M.; Wakabayashi, K.; Takeuchi, H.T.; Matsukura, S.; Adachi, M.; Tezuka, M.; Kobayashi, K. Expression of angiopoietin-1 in osteoblasts and its inhibition by tumor necrosis factor-alpha and interferon-gamma. Transl. Res. 2007, 149, 265–273. [Google Scholar] [CrossRef]
- Qian, H.; Buza-Vidas, N.; Hyland, C.D.; Jensen, C.T.; Antonchuk, J.; Månsson, R.; Thoren, L.A.; Ekblom, M.; Alexander, W.S.; Jacobsen, S.E.W. Critical Role of Thrombopoietin in Maintaining Adult Quiescent Hematopoietic Stem Cells. Cell Stem Cell 2007, 1, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, H.; Arai, F.; Hosokawa, K.; Hagiwara, T.; Takubo, K.; Nakamura, Y.; Gomei, Y.; Iwasaki, H.; Matsuoka, S.; Miyamoto, K.; et al. Thrombopoietin/MPL signaling regulates hematopoietic stem cell quiescence and interaction with the osteoblastic niche. Cell Stem Cell 2007, 1, 685–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visnjic, D.; Kalajzic, Z.; Rowe, D.W.; Katavic, V.; Lorenzo, J.; Aguila, H.L. Hematopoiesis is severely altered in mice with an induced osteoblast deficiency. Blood 2004, 103, 3258–3264. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Purton, L.E.; Rodda, S.J.; Chen, M.; Weinstein, L.S.; McMahon, A.P.; Scadden, D.T.; Kronenberg, H.M. Osteoblastic regulation of B lymphopoiesis is mediated by Gs{alpha}-dependent signaling pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 16976–16981. [Google Scholar] [CrossRef] [Green Version]
- Rankin, E.B.; Wu, C.; Khatri, R.; Wilson, T.L.S.; Andersen, R.; Araldi, E.; Rankin, A.L.; Yuan, J.; Kuo, C.J.; Schipani, E.; et al. The HIF Signaling Pathway in Osteoblasts Directly Modulates Erythropoiesis through the Production of EPO. Cell 2012, 149, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Salati, S.; Lisignoli, G.; Manferdini, C.; Pennucci, V.; Zini, R.; Bianchi, E.; Norfo, R.; Facchini, A.; Ferrari, S.; Manfredini, R. Co-culture of hematopoietic stem/progenitor cells with human osteblasts favours mono/macrophage differentiation at the expense of the erythroid lineage. PLoS ONE 2013, 8, e53496. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Giaccia, A.J.; Rankin, E.B. Osteoblasts: A novel source of erythropoietin. Curr. Osteoporos. Rep. 2014, 12, 428–432. [Google Scholar] [CrossRef] [Green Version]
- Holst, J.; Watson, S.; Lord, M.S.; Eamegdool, S.S.; Bax, D.V.; Nivison-Smith, L.B.; Kondyurin, A.; Ma, L.; Oberhauser, A.F.; Weiss, A.S.; et al. Substrate elasticity provides mechanical signals for the expansion of hemopoietic stem and progenitor cells. Nat. Biotechnol. 2010, 28, 1123–1128. [Google Scholar] [CrossRef]
- Prewitz, M.C.; Stißel, A.; Friedrichs, J.; Träber, N.; Vogler, S.; Bornhäuser, M.; Werner, C. Extracellular matrix deposition of bone marrow stroma enhanced by macromolecular crowding. Biomaterials 2015, 73, 60–69. [Google Scholar] [CrossRef]
- Goncharova, V.; Serobyan, N.; Iizuka, S.; Schraufstatter, I.; de Ridder, A.; Povaliy, T.; Wacker, V.; Itano, N.; Kimata, K.; Orlovskaja, I.A.; et al. Hyaluronan expressed by the hematopoietic microenvironment is required for bone marrow hematopoiesis. J. Biol. Chem. 2012, 287, 25419–25433. [Google Scholar] [CrossRef] [Green Version]
- Bessis, M. Erythroblastic island, functional unity of bone marrow. Rev. Hematol. 1958, 13, 8–11. [Google Scholar] [PubMed]
- Mohandas, N.; Prenant, M. Three-dimensional model of bone marrow. Blood 1978, 51, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, T.; Etoh, T.; Kitagawa, H.; Tsukahara, S.; Kannan, Y. Migration of erythroblastic islands toward the sinusoid as erythroid maturation proceeds in rat bone marrow. J. Vet. Med. Sci. 2003, 65, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Lucas, D.; Hidalgo, A.; Méndez-Ferrer, S.; Hashimoto, D.; Scheiermann, C.; Battista, M.; Leboeuf, M.; Prophete, C.; van Rooijen, N.; et al. Bone marrow CD169+ macrophages promote the retention of hematopoietic stem and progenitor cells in the mesenchymal stem cell niche. J. Exp. Med. 2011, 208, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, R.N.; Forristal, C.E.; Raggatt, L.J.; Nowlan, B.; Barbier, V.; Kaur, S.; Van Rooijen, N.; Winkler, I.G.; Pettit, A.R. Mobilization with granulocyte colony-stimulating factor blocks medullar erythroid island macrophages in the mouse. Exp. Hematol. 2014, 42, 547–561.e4. [Google Scholar] [CrossRef] [Green Version]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; van Rooijen, N.; Alexander, K.A.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, Y.; Zhao, H.; Zhang, H.; Xu, Y.; Wang, S.; Guo, X.; Huang, Y.; Zhang, S.; Han, Y.; et al. Identification and transcriptome analysis of erythroblastic island macrophages. Blood 2019, 134, 480–491. [Google Scholar] [CrossRef]
- Seu, K.G.; Papoin, J.; Fessler, R.; Hom, J.; Huang, G.; Mohandas, N.; Blanc, L.; Kalfa, T.A. Unraveling Macrophage Heterogeneity in Erythroblastic Islands. Front. Immunol. 2017, 8, 1140. [Google Scholar] [CrossRef] [Green Version]
- Heideveld, E.; Akker, E. Van Den Immunobiology Digesting the role of bone marrow macrophages on hematopoiesis. Immunobiology 2017, 222, 814–822. [Google Scholar] [CrossRef]
- Hanspal, M.; Hanspal, J.S. The association of erythroblasts with macrophages promotes erythroid proliferation and maturation: A 30-kD heparin-binding protein is involved in this contact. Blood 1994, 84, 3494–3504. [Google Scholar] [CrossRef] [Green Version]
- Gifford, S.C.; Derganc, J.; Shevkoplyas, S.S.; Yoshida, T.; Bitensky, M.W. A detailed study of time-dependent changes in human red blood cells: From reticulocyte maturation to erythrocyte senescence. Br. J. Haematol. 2006, 135, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Sadahira, Y.; Yoshino, T.; Monobe, Y. Very late activation antigen 4-vascular cell adhesion molecule 1 interaction is involved in the formation of erythroblastic islands. J. Exp. Med. 1995, 181, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Lo, A.; Short, S.A.; Mankelow, T.J.; Spring, F.; Parsons, S.F.; Yazdanbakhsh, K.; Mohandas, N.; Anstee, D.J.; Chasis, J.A. Targeted gene deletion demonstrates that the cell adhesion molecule ICAM-4 is critical for erythroblastic island formation. Blood 2006, 108, 2064–2071. [Google Scholar] [CrossRef] [Green Version]
- Toda, S.; Segawa, K.; Nagata, S. MerTK-mediated engulfment of pyrenocytes by central macrophages in erythroblastic islands. Blood 2014, 123, 3963–3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, R.N.; Perkins, A.C.; Levesque, J.-P. Macrophages and regulation of erythropoiesis. Curr. Opin. Hematol. 2015, 22, 212–219. [Google Scholar] [CrossRef]
- Javan, G.T.; Can, I.; Yeboah, F.; Lee, Y.; Soni, S. Blood Cells, Molecules and Diseases Novel interactions between erythroblast macrophage protein and cell migration. Blood Cells Mol. Dis. 2016, 60, 24–27. [Google Scholar] [CrossRef] [Green Version]
- Javan, G.T.; Salhotra, A.; Finley, S.J.; Soni, S. Erythroblast macrophage protein (Emp): Past, present, and future. Eur. J. Haematol. 2018, 100, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Hampton-O’Neil, L.A.; Severn, C.E.; Cross, S.J.; Gurung, S.; Nobes, C.D.; Toye, A.M. Ephrin/Eph receptor interaction facilitates macrophage recognition of differentiating human erythroblasts. Haematologica 2020, 105, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, M.M.; Kopsombut, P.; Bondurant, M.C.; Price, J.O.; Koury, M.J. Adherence to macrophages in erythroblastic islands enhances erythroblast proliferation and increases erythrocyte production by a different mechanism than erythropoietin. Blood 2008, 111, 1700–1708. [Google Scholar] [CrossRef] [Green Version]
- Falchi, M.; Varricchio, L.; Martelli, F.; Masiello, F.; Federici, G.; Zingariello, M.; Girelli, G.; Whitsett, C.; Petricoin, E.F., 3rd; Moestrup, S.K.; et al. Dexamethasone targeted directly to macrophages induces macrophage niches that promote erythroid expansion. Haematologica 2015, 100, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Ramos, P.; Casu, C.; Gardenghi, S.; Breda, L.; Crielaard, B.J.; Guy, E.; Marongiu, M.F.; Gupta, R.; Levine, R.L.; Abdel-Wahab, O.; et al. Macrophages support pathological erythropoiesis in polycythemia vera and β-thalassemia. Nat. Med. 2013, 19, 437–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chasis, J.A.; Mohandas, N. Erythroblastic islands: Niches for erythropoiesis. Blood 2008, 112, 470–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manwani, D.; Bieker, J.J. The erythroblastic island. Curr. Top. Dev. Biol. 2008, 82, 23–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, E.Y.; Krauss, S.W.; Short, S.A.; Lee, G.; Villalobos, J.; Etzell, J.; Koury, M.J.; Ney, P.A.; Chasis, J.A.; Grigoryev, S.A. Chromatin condensation in terminally differentiating mouse erythroblasts does not involve special architectural proteins but depends on histone deacetylation. Chromosome Res. 2009, 17, 47–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, P.; Yeh, V.; Ramirez, T.; Murata-Hori, M.; Lodish, H.F. Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts. Haematologica 2010, 95, 2013–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Kawane, K.; Koike, M.; Mori, Y.; Uchiyama, Y.; Nagata, S. Phosphatidylserine-dependent engulfment by macrophages of nuclei from erythroid precursor cells. Nature 2005, 437, 754–758. [Google Scholar] [CrossRef]
- Bessis, M.; Breton-Gorius, J. Various aspects of iron in the organism. I. Ferritin and ferruginous micelles. J. Biophys. Biochem. Cytol. 1959, 6, 231–236. [Google Scholar] [CrossRef]
- Yeo, J.H.; Colonne, C.K.; Tasneem, N.; Cosgriff, M.P.; Fraser, S.T. The iron islands: Erythroblastic islands and iron metabolism. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 466–471. [Google Scholar] [CrossRef]
- Heideveld, E.; Masiello, F.; Marra, M.; Esteghamat, F.; Yağcı, N.; von Lindern, M.; Migliaccio, A.R.F.; van den Akker, E. CD14+ cells from peripheral blood positively regulate hematopoietic stem and progenitor cell survival resulting in increased erythroid yield. Haematologica 2015, 100, 1396–1406. [Google Scholar] [CrossRef] [Green Version]
- Belay, E.; Hayes, B.J.; Blau, C.A.; Torok-Storb, B. Human Cord Blood and Bone Marrow CD34+ Cells Generate Macrophages That Support Erythroid Islands. PLoS ONE 2017, 12, e0171096. [Google Scholar] [CrossRef]
- Quentmeier, H.; Zaborski, M.; Drexler, H.G. The human bladder carcinoma cell line 5637 constitutively secretes functional cytokines. Leuk. Res. 1997, 21, 343–350. [Google Scholar] [CrossRef]
- Zarif, J.C.; Hernandez, J.R.; Verdone, J.E.; Campbell, S.P.; Drake, C.G.; Pienta, K.J. A phased strategy to differentiate human CD14+monocytes into classically and alternatively activated macrophages and dendritic cells. Biotechniques 2016, 61, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Yrigoyen, M.; Yang, C.T.; Fidanza, A.; Cassetta, L.; Taylor, A.H.; McCahill, A.; Sellink, E.; von Lindern, M.; van den Akker, E.; Mountford, J.C.; et al. Genetic programming of macrophages generates an in vitro model for the human erythroid island niche. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elvarsdóttir, E.M.; Mortera-Blanco, T.; Dimitriou, M.; Bouderlique, T.; Jansson, M.; Hofman, I.J.F.; Conte, S.; Karimi, M.; Sander, B.; Douagi, I.; et al. A three-dimensional in vitro model of erythropoiesis recapitulates erythroid failure in myelodysplastic syndromes. Leukemia 2020, 34, 271–282. [Google Scholar] [CrossRef] [Green Version]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deleschaux, C.; Moras, M.; Lefevre, S.D.; Ostuni, M.A. An Overview of Different Strategies to Recreate the Physiological Environment in Experimental Erythropoiesis. Int. J. Mol. Sci. 2020, 21, 5263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155263
Deleschaux C, Moras M, Lefevre SD, Ostuni MA. An Overview of Different Strategies to Recreate the Physiological Environment in Experimental Erythropoiesis. International Journal of Molecular Sciences. 2020; 21(15):5263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155263
Chicago/Turabian StyleDeleschaux, Cécile, Martina Moras, Sophie D. Lefevre, and Mariano A. Ostuni. 2020. "An Overview of Different Strategies to Recreate the Physiological Environment in Experimental Erythropoiesis" International Journal of Molecular Sciences 21, no. 15: 5263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155263