Identification of a Novel Gene, Osbht, in Response to High Temperature Tolerance at Booting Stage in Rice



Abstract
:1. Introduction
2. Results
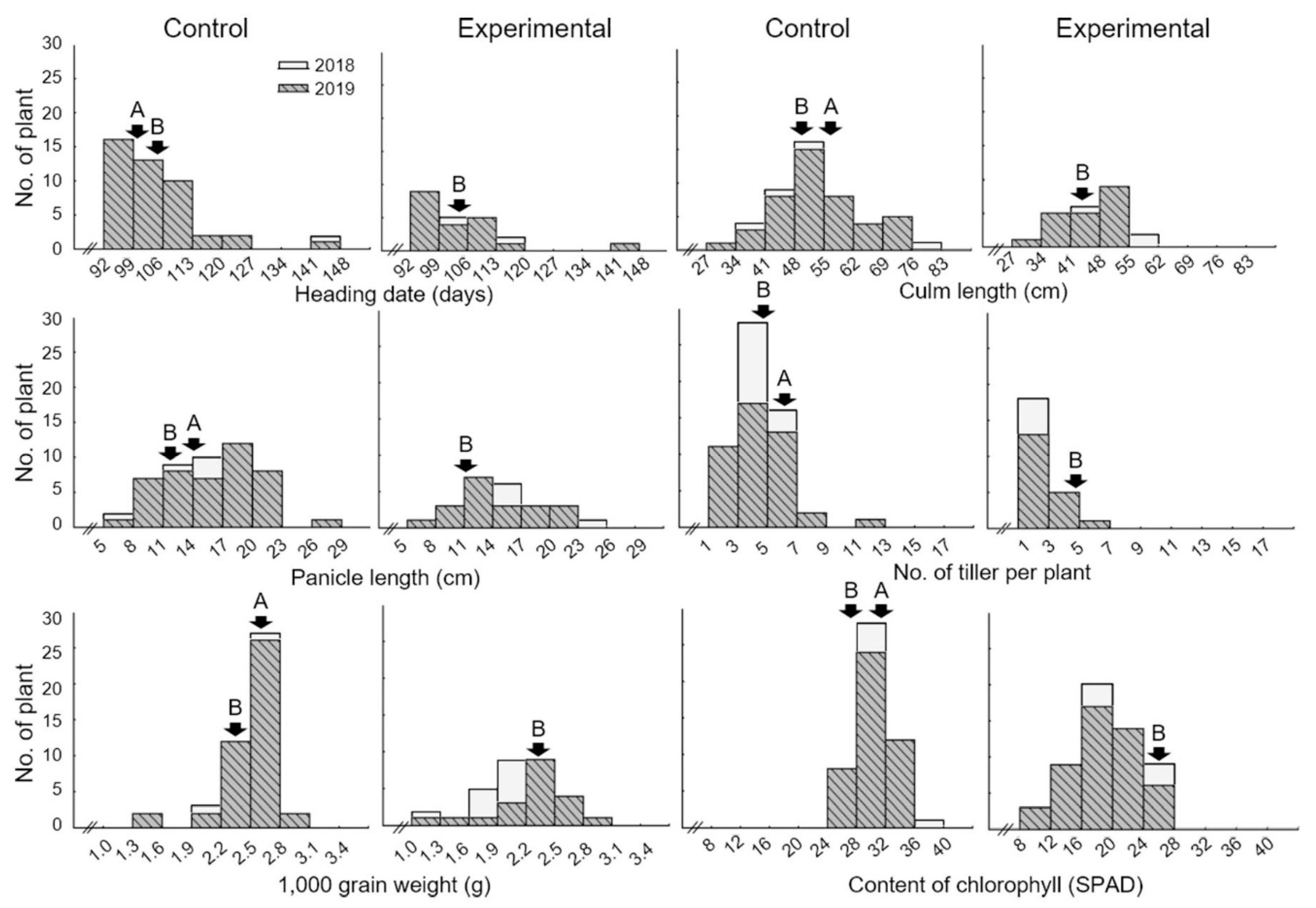
2.1. Phenotypic Evaluation and Comparison with Agricultural Characteristic
2.2. Analysis of QTLs Associated with Heat Tolerance
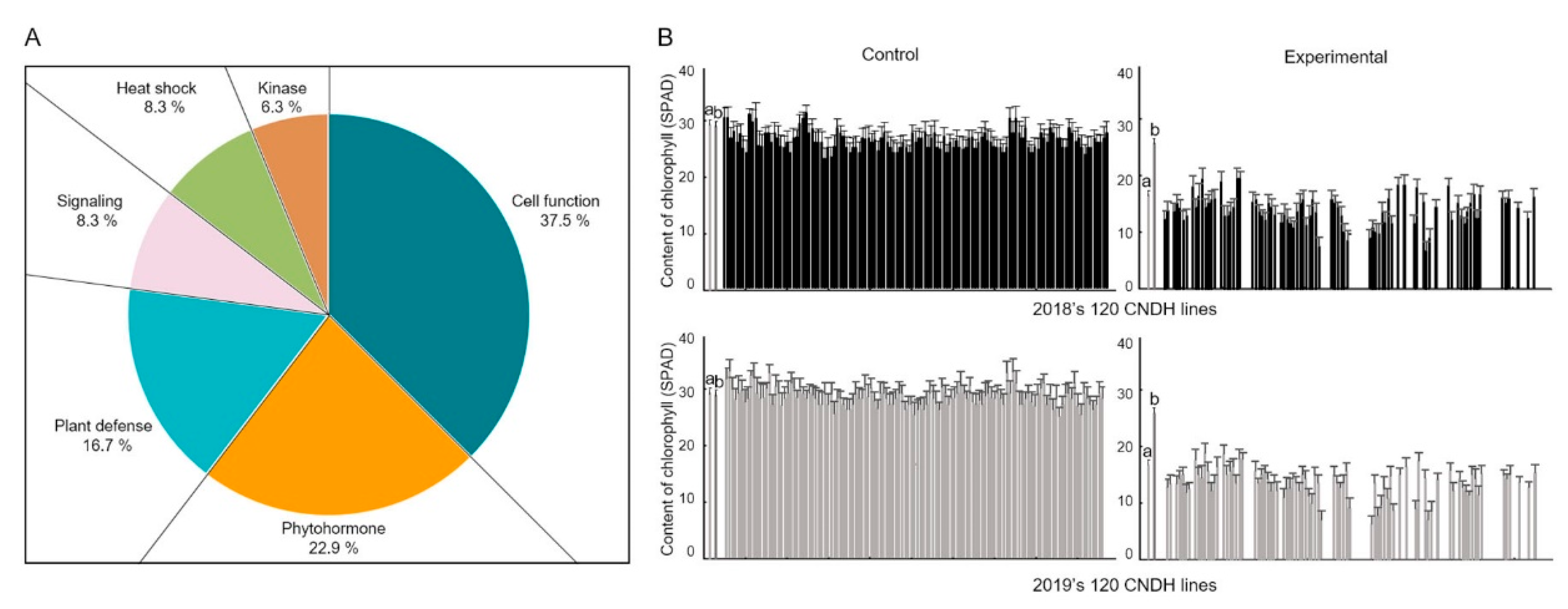
2.3. Search for Candidate Genes Associated with Heat Tolerance Based on QTL Mapping
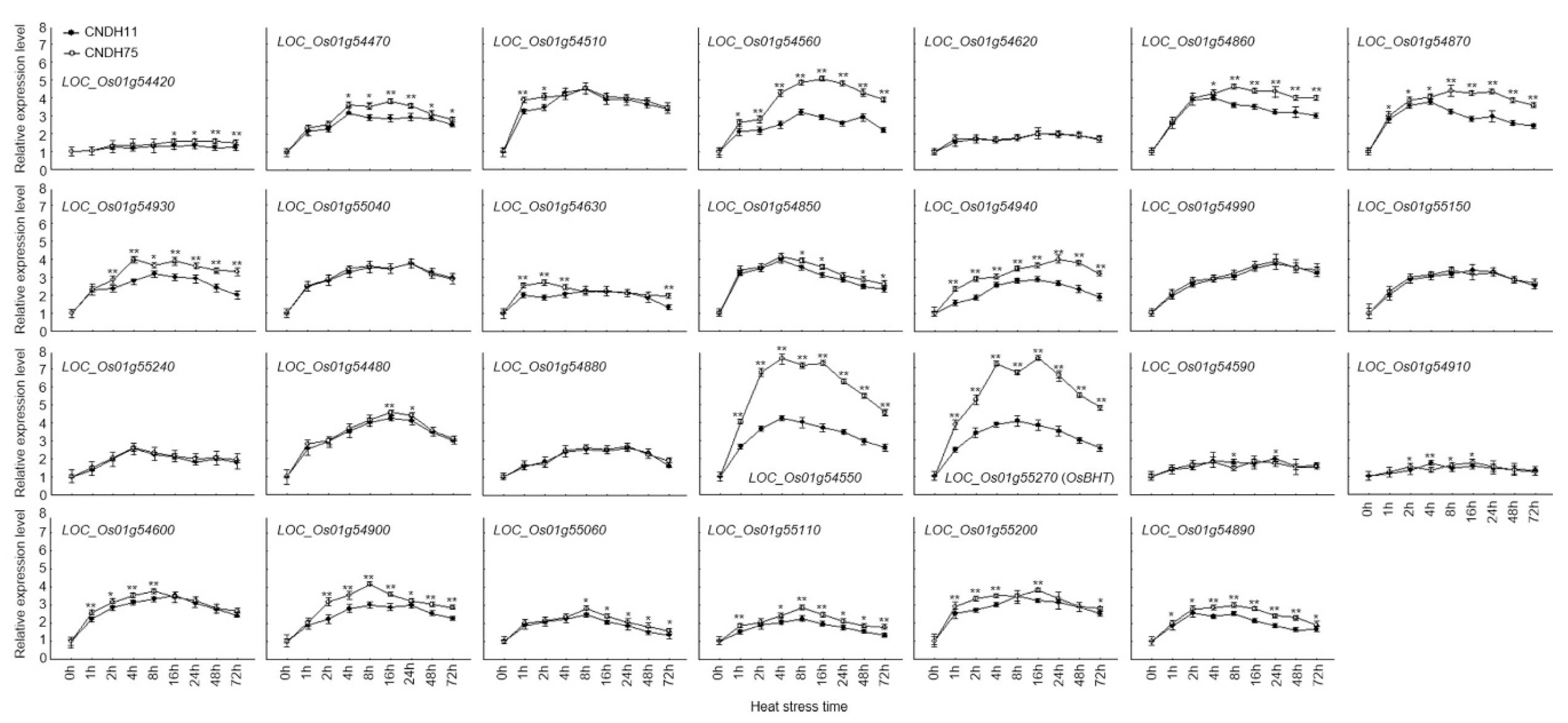
2.4. Candidate Genes Expression Level at Booting Stage with Treatment of High Temperature
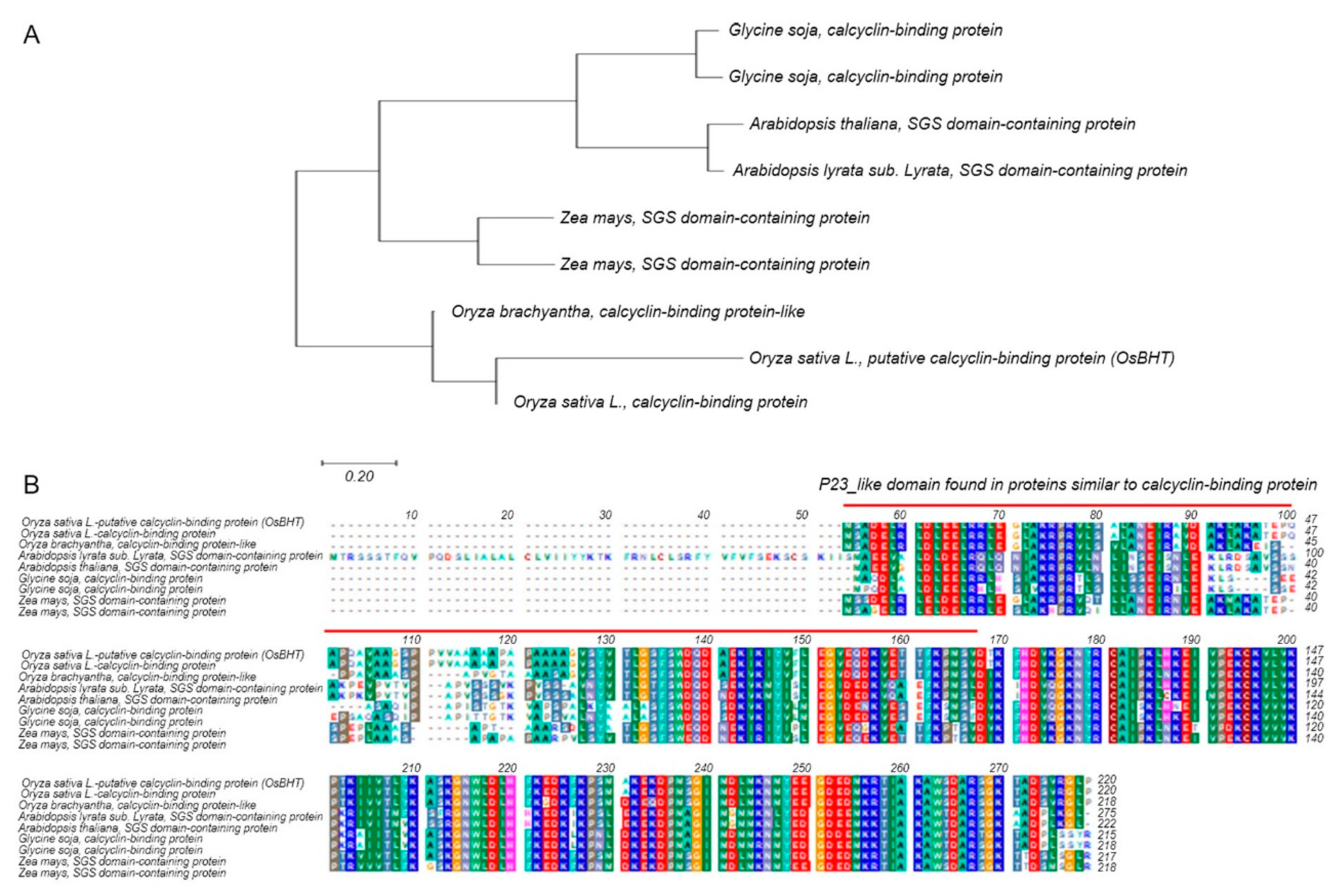
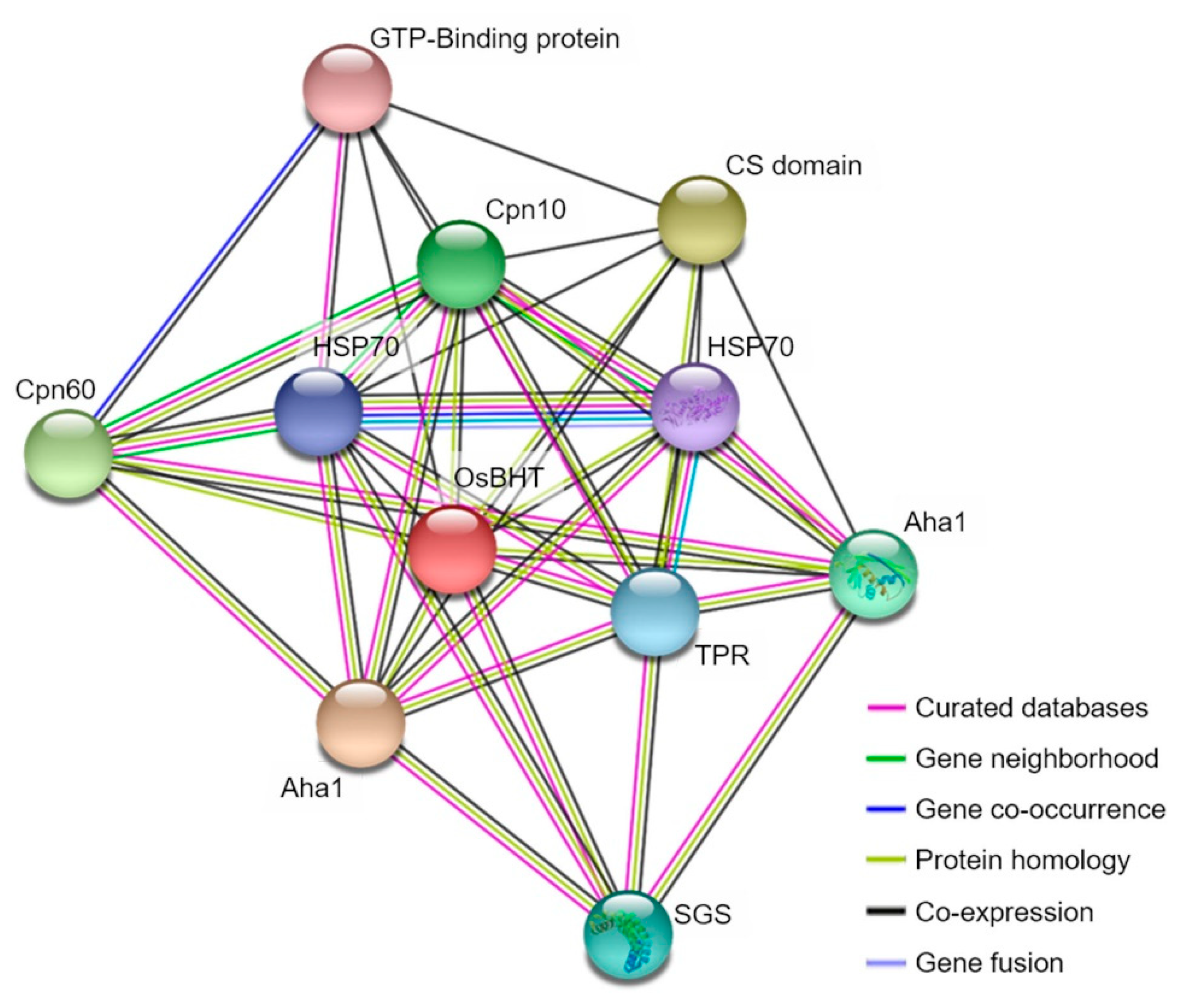
2.5. Analysis of Phylogenetic Tree and Homology Sequence
3. Discussion
4. Materials and Methods
4.1. Plant Material and Field Design
4.2. Phenotypic Evaluation
4.3. Construction of a Genetic Map and QTL Analysis of Heat Tolerance in Rice
4.4. Gene Information Analysis
4.5. Analysis of Candidate Genes Expression Level
4.6. RNA Extraction and PCR Protocol
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. 2017. Available online: http://www.fao.org/faostat/en/#home (accessed on 8 August 2018).
- International Food Policy Research Institute [IFPRI]. 2018 Global Food Policy Report; International Food Policy Research Institute: Washington, DC, USA, 2018. [Google Scholar]
- Suzuki, M. Studies on distinctive patterns of dry matter production in the building process of grain yields in rice plants grown in the warm region in Japan. Bull. Kyushu Agric. Exp. Stn. 1980, 20, 429–494. [Google Scholar]
- Kim, J.; Shon, J.; Lee, C.K.; Yang, W.; Yoon, Y.; Yang, W.H.; Kim, Y.G.; Lee, B.W. Relationship between grain filling duration and leaf senescence of temperature rice under high temperature. Field Crops Res. 2011, 122, 207–213. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Boote, K.J.; Allen, L.H.; Sheehy, J.E.; Thomas, J.M.G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Yoshida, S. Fundamentals of Rice Crop Science; International Rice Research Institute: Los Banos, Philippines, 1981; 269p. [Google Scholar]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [Green Version]
- Kobata, T.; Uemuki, N. High temperatures during the grain-filling period do not reduce the potential grain dry matter increas of rice. Agron. J. 2004, 96, 406–414. [Google Scholar] [CrossRef]
- Tanksley, S.D. Mapping polygenes. Annu. Rev. Genet. 1993, 27, 205–233. [Google Scholar] [CrossRef]
- Delaneau, O.; Ongen, H.; Brown, A.A.; Fort, A.; Panousis, N.I.; Dermitzakis, E.T. A complete tool set for molecular QTL discovery and analysis. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sen, Ś.; Churchill, G.A. A statistical framework for quantitative trait mapping. Genetics 2001, 159, 371–387. [Google Scholar]
- Jagadish, S.V.K.; Cairns, J.; Lafitte, R.; Wheeler, T.R.; Price, A.H.; Craufurd, P.Q. Genetic analysis of heat tolerance at anthesis in rice. Crop Sci. 2010, 50, 1633–1641. [Google Scholar] [CrossRef]
- Qingquan, C.; Sibin, Y.; Chunhai, L. Identification of QTLs for heat tolerance at flowering stage in rice. Sci. Agric. Sin. 2008, 41, 315–321. [Google Scholar]
- Satake, T.; Yosida, S. High temperature-induced sterility in indica rices at flowering. Jpn. J. Crop Sci. 1978, 47, 6–17. [Google Scholar] [CrossRef] [Green Version]
- Farrell, T.C.; Fox, K.M.; Williams, R.L.; Fukai, S. Genotypic variation for cold tolerance during reproductive development in rice: Screening with cold air and cold water. Field Crops Res. 2006, 98, 178–194. [Google Scholar] [CrossRef]
- Yoshida, S. Tropical Climate and its Influence on Rice; IRRI Research Paper Series; IRRI: Los Baños, Philippines, 1978; Volume 20. [Google Scholar]
- Wreford, A.; Adger, W.N. Adaptation in agriculture: Historic effects of heat waves and droughts on UK agriculture. Int. J. Agric. Sustain. 2010, 8, 278–289. [Google Scholar] [CrossRef]
- Maria, P.H.; Johnson, R.R. High temperature stress and pollen viability of maize. Crop Sci. 1980, 20, 796–800. [Google Scholar] [CrossRef]
- Ferris, R.; Wheeler, T.R.; Ellis, R.H.; Hadley, P.; Wollenweber, B.; Porter, J.R.; Schellberg, J. Effects of high temperature extremes on wheat. In Climate Change, Climatic Variability and Agriculture in Europe-an Integrated Assessment; University of Oxford, Environmental Change Institute: Oxford, UK, 2000; pp. 31–57. [Google Scholar]
- Lu, C.F.; Shen, L.S.; Tan, Z.; Xu, Y.; He, P.; Chen, Y.; Zhu, L. Comparative mapping of QTLs for agronomic traits of rice across environments using a doubled haploid population. Theor. Appl. Genet. 1996, 93, 1211–1217. [Google Scholar] [CrossRef]
- Hussain, A.; Mun, B.G.; Imran, Q.M.; Lee, S.U.; Adamu, T.A.; Shahid, M.; Kim, K.M.; Yun, B. Nitric oxide mediated transcriptome profiling reveals activation of multiple regulatory pathways in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 975. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Lee, S.S.; Tanaka, T.; Numa, H.; Kim, J.; Kawahara, Y.; Yamada, Y. Rice Annotation Project Database (RAP-DB): An integrative and interactive database for rice genomics. Plant Cell Physiol. 2013, 54, e6. [Google Scholar] [CrossRef]
- Sato, Y.; Takehisa, H.; Kamatsuki, K.; Minami, H.; Namiki, N.; Ikawa, H.; Nagamura, Y. RiceXPro version 3.0: Expanding the informatics resource for rice transcriptome. Nucleic Acids Res. 2013, 41, D1206–D1213. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.F.; Yang, T.M.; Li, R.Z.; Lei, Y.; Gao, S.H. High temperature induced heat damage and its impacts on early rice yields in Zhejiang province. Chin. J. Agrometeorol. 2009, 30, 628–631. [Google Scholar]
- Morita, S.; Wada, H.; Matsue, Y. Countermeasures for heat damage in rice grain quality under climate change. Plant Prod. Sci. 2016, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Matsuba, S.; Funatsuki, H.; Kawaguchi, K.; Saruyama, H.; Tanida, M.; Sato, Y. Over-expression of a small heat shock protein, sHSP17. 7, confers both heat tolerance and UV-B resistance to rice plants. Mol. Breed. 2004, 13, 165–175. [Google Scholar] [CrossRef]
- Lang, N.T.; Ha, P.T.T.; Tru, P.C.; Toan, T.B.; Buu, B.C.; Cho, Y.C. Breeding for heat tolerance rice based on marker-assisted backcrosing in Vietnam. Plant Breed. Biotechnol. 2015, 3, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Pan, Y.; Luo, L.; Zhang, G.; Deng, H.; Dai, L.; Wang, G.L. Quantitative trait loci associated with seed set under high temperature stress at the flowering stage in rice (Oryza sativa L.). Euphytica 2011, 178, 331–338. [Google Scholar] [CrossRef]
- Ye, C.; Argayoso, M.A.; Redoña, E.D.; Sierra, S.N.; Laza, M.A.; Dilla, C.J.; Diaz, G.Q. Mapping QTL for heat tolerance at flowering stage in rice using SNP markers. Plant Breed. 2012, 131, 33–41. [Google Scholar] [CrossRef]
- Gosavi, G.U.; Jadhav, A.S.; Kale, A.A.; Gadakh, S.R.; Pawar, B.D.; Chimote, V.P. Effect of heat stress on proline, chlorophyll content, heat shock proteins and antioxidant enzyme activity in sorghum (Sorghum bicolor) at seedlings stage. Indian J. Biotechnol. 2014, 13, 356–363. [Google Scholar]
- Guo, L.M.; Li, J.; He, J.; Liu, H.; Zhang, H.M. A class I cytosolic HSP20 of rice enhances heat and salt tolerance in different organisms. Sci. Rep. 2020, 10, 1383. [Google Scholar] [CrossRef]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef]
- Goral, A.; Bieganowski, P.; Prus, W.; Krzemień-Ojak, Ł.; Kądziołka, B.; Fabczak, H.; Filipek, A. Calcyclin binding protein/Siah-1 interacting protein is a Hsp90 binding chaperone. PLoS ONE 2016, 11, e0156507. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ham, B.K.; Paek, K.H.; Park, C.M.; Chua, N.H. An Arabidopsis homologue of human seven-in-absentia-interacting protein is involved in pathogen resistance. Mol. Cells (Springer Sci. Bus. Media BV) 2006, 21, 389–394. [Google Scholar]
- Tosoni, K.; Costa, A.; Sarno, S.; D’Alessandro, S.; Sparla, F.; Pinna, L.A.; Ruzzene, M. The p23 co-chaperone protein is a novel substrate of CK2 in Arabidopsis. Mol. Cell. Biochem. 2011, 356, 245–254. [Google Scholar] [CrossRef]
- Venkatakrishnan, A.J.; Deupi, X.; Lebon, G.; Tate, C.G.; Schertler, G.F.; Babu, M.M. Molecular signatures of G-protein-coupled receptors. Nature 2013, 494, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Errington, N.; Rowe, A.J.; Harding, S.E. Hydrolysable ATP is a requirement for the correct interaction of molecular chaperonins cpn60 and cpn10. Biochem. J. 2002, 364, 849–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.D.; Caron, A.W.; Bourget, L.; Meriin, A.B.; Sherman, M.Y.; Morimoto, R.I.; Massie, B. The chaperone function of hsp70 is required for protection against stress-induced apoptosis. Mol. Cell. Biol. 2000, 20, 7146–7159. [Google Scholar] [CrossRef] [Green Version]
- Wegele, H.; Müller, L.; Buchner, J. Hsp70 and Hsp90—A relay team for protein folding. In Reviews of Physiology, Biochemistry and Pharmacology; Springer: Berlin/Heidelberg, Germany, 2004; Volume 151, pp. 1–44. [Google Scholar] [CrossRef]
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 2008, 283, 18473–18477. [Google Scholar] [CrossRef] [Green Version]
- Sharp, F.R.; Massa, S.M.; Swanson, R.A. Heat-shock protein protection. Trends Neurosci. 1999, 22, 97–99. [Google Scholar] [CrossRef]
- Siligardi, G.; Hu, B.; Panaretou, B.; Piper, P.W.; Pearl, L.H.; Prodromou, C. Co-chaperone regulation of conformational switching in the Hsp90 ATPase cycle. J. Biol. Chem. 2004, 279, 51989–51998. [Google Scholar] [CrossRef] [Green Version]
- Panaretou, B.; Siligardi, G.; Meyer, P.; Maloney, A.; Sullivan, J.K.; Singh, S.; Cramer, R. Activation of the ATPase activity of hsp90 by the stress-regulated cochaperone aha1. Mol. Cell 2002, 10, 1307–1318. [Google Scholar] [CrossRef] [Green Version]
- Wagner, R.N.; Proell, M.; Kufer, T.A.; Schwarzenbacher, R. Evaluation of Nod-like receptor (NLR) effector domain interactions. PLoS ONE 2009, 4, e4931. [Google Scholar] [CrossRef] [Green Version]
- Shirasu, K. The HSP90-SGT1 chaperone complex for NLR immune sensors. Annu. Rev. Plant Biol. 2009, 60, 139–164. [Google Scholar] [CrossRef] [Green Version]
- Kadota, Y.; Shirasu, K. The HSP90 complex of plants. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 689–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Ai, C.R.; Jing, S.J.; Yu, D.Q. Research progress on functional analysis of rice WRKY genes. Rice Sci. 2010, 17, 60–72. [Google Scholar] [CrossRef]
- Li, Q.; Yu, H.M.; Meng, X.F.; Lin, J.S.; Li, Y.J.; Hou, B.K. Ectopic expression of glycosyltransferase UGT 76E11 increases flavonoid accumulation and enhances abiotic stress tolerance in Arabidopsis. Plant Biol. 2018, 20, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K.; Sasaki, K.; Tsurumaru, Y. Prenylation of aromatic compounds, a key diversification of plant secondary metabolites. Phytochemistry 2009, 70, 1739–1745. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Mian, A.; Maathuis, F.J. Overexpression of the rice AKT1 potassium channel affects potassium nutrition and rice drought tolerance. J. Exp. Bot. 2016, 67, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Sewelam, N.; Kazan, K.; Thomas-Hall, S.R.; Kidd, B.N.; Manners, J.M.; Schenk, P.M. Ethylene response factor 6 is a regulator of reactive oxygen species signaling in Arabidopsis. PLoS ONE 2013, 8, e70289. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.; Kim, S.K.; Cho, S.K.; Kang, B.G.; Kim, W.T. Overexpression of OsRDCP1, a rice RING domain-containing E3 ubiquitin ligase, increased tolerance to drought stress in rice (Oryza sativa L.). Plant Sci. 2011, 180, 775–782. [Google Scholar] [CrossRef]
- Rensink, W.A.; Iobst, S.; Hart, A.; Stegalkina, S.; Liu, J.; Buell, C.R. Gene expression profiling of potato responses to cold, heat, and salt stress. Funct. Integr. Genomics 2005, 5, 201–207. [Google Scholar] [CrossRef]
- Yuan, K.; Rashotte, A.M.; Wysocka-Diller, J.W. ABA and GA signaling pathways interact and regulate seed germination and seedling development under salt stress. Acta Physiol. Plant. 2011, 33, 261–271. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Houston, K.; Tucker, M.R.; Chowdhury, J.; Shirley, N.; Little, A. The plant cell wall: A complex and dynamic structure as revealed by the responses of genes under stress conditions. Front. Plant Sci. 2016, 7, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, K.; Izui, K. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 2000, 23, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Hayasaka, H.; Chiba, B.; Tanaka, I.; Shimano, T.; Yamagishi, M.; Yano, M. Mapping quantitative trait loci controlling cool-temperature tolerance at booting stage in temperate japonica rice. Breed. Sci. 2001, 51, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Lei, J.; Huang, Y.; Zhu, S.; Chen, H.; Huang, R.; Yan, S. Mapping quantitative trait loci for heat tolerance at anthesis in rice using chromosomal segment substitution lines. Breed. Sci. 2016, 66, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Li, J.; Yuan, L.; Iwata, N. Identification of QTLs affecting traits of agronomic in a recombinant inbred population derived from a subspecific rice cross. Theor. Appl. Genet. 1996, 92, 230–244. [Google Scholar] [CrossRef]
- Pallant, J.; Manual, S.S. A step by step guide to data analysis using SPSS for windows. In SPSS Survival Manual; Open University Press: London, UK, 2007. [Google Scholar]
- McCouch, S.R. Gene nomenclature system for rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequence (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2005, 33, D501–D504. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit Version 7.0.0. Distributed by the Author. 2004. Available online: www.mbio.ncsu.edu/BioEdit/bioedit.html (accessed on 15 August 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Traits | Year | Parents | DH Population | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cheongcheong | Nagdong | |||||||||
| Control | Experimental | p-Value | Control | Experimental | p-Value | Control | Experimental | p-Value | ||
| Heading date (Days) | 2018 | 101.7 ± 1.2 z | - | - | 104.5 ± 1.5 | 105.2 ± 1.3 | 0.054 | 103.7 ± 10.1 | 104.7 ± 6.7 | 0.854 |
| 2019 | 102.6 ± 0.8 | - | - | 105.3 ± 1.1 | 106.3 ± 1.2 | 0.059 | 104.0 ± 8.3 | 105.4 ± 6.3 | 0.633 | |
| Culm length (cm) | 2018 | 55.8 ± 1.6 | - | - | 53.7 ± 1.4 | 47.8 ± 2.4 | 0.094 | 53.9 ± 10.2 | 46.7 ± 7.6 | 0.007 ** |
| 2019 | 53.7 ± 2.4 | - | - | 52.8 ± 1.8 | 48.1 ± 2.2 | 0.005 | 54.4 ±10.2 | 46.4 ± 7.2 | 0.003 ** | |
| Panicle length (cm) | 2018 | 14.7 ± 2.8 | - | - | 12.6 ± 3.4 | 11.2 ± 2.5 | 0.080 | 15.6 ± 4.5 | 15.6 ± 4.1 | 0.947 |
| 2019 | 12.6 ± 1.7 | - | - | 11.9 ± 1.2 | 10.6 ± 2.5 | 0.150 | 16.0 ±4.7 | 14.9 ± 4.2 | 0.339 | |
| Number of tillers | 2018 | 6.7 ± 1.5 | - | - | 5.4 ± 1.7 | 5.0 ± 1.1 | 0.388 | 5.0 ± 1.1 | 2.5 ± 1.1 | <0.001 *** |
| 2019 | 6.2 ± 1.2 | - | - | 5.1 ± 0.7 | 4.9 ± 0.9 | 0.588 | 4.7 ± 2.1 | 2.4 ± 1.5 | <0.001 *** | |
| 1000 grain weight (g) | 2018 | 2.6 ± 0.1 | - | - | 2.4 ± 0.3 | 2.3 ± 0.2 | 0.151 | 2.6 ± 0.3 | 1.9 ± 0.3 | 0.031 * |
| 2019 | 2.5 ± 1.3 | - | - | 2.3 ± 0.1 | 2.2 ± 0.1 | 0.003 ** | 2.6 ±0.3 | 1.8 ± 0.2 | 0.017 * | |
| Contents of chlorophyll (SPAD) | 2018 | 29.5 ± 2.6 | 18.7 ± 2.7 | <0.001 *** | 28.9 ± 1.2 | 26.7 ± 0.9 | <0.001 *** | 30.3 ± 2.8 | 19.6 ± 4.5 | <0.001 *** |
| 2019 | 28.8 ± 1.7 | 17.5 ± 2.8 | <0.001 *** | 27.8 ± 0.9 | 25.7 ± 1.2 | 0.002 ** | 30.2 ±2.6 | 18.6 ± 4.3 | <0.001 *** | |
| Characteristics | Year | QTL | Chromosome | Interval Markers z | LOD | Additive effect y | r2 x | Increasing Effects w |
|---|---|---|---|---|---|---|---|---|
| Heading date | 2018 | qHdd1 | 1 | RM212–RM1297 | 4.15 | −15.0 | 0.2 | Nagdong |
| qHdd1-1 | 1 | RM11605–RM3530 | 2.70 | −16.1 | 0.2 | Nagdong | ||
| 2019 | qHdd1-2 | 1 | RM212–RM1297 | 4.20 | −11.2 | 0.2 | Nagdong | |
| Culm length | 2018 | qCl1 | 1 | RM212–RM3411 | 4.58 | −7.2 | 0.2 | Nagdong |
| qCl1-1 | 1 | RM11605–RM3530 | 2.70 | −5.7 | 0.2 | Nagdong | ||
| qCl112 | 12 | RM8216–RM28816 | 2.95 | −6.3 | 0.2 | Nagdong | ||
| 2019 | qCl1-2 | 1 | RM212–RM1297 | 4.39 | −7.7 | 0.2 | Nagdong | |
| Panicle length | 2018 | qPl1 | 1 | RM212–RM3411 | 4.24 | −2.2 | 0.3 | Nagdong |
| qPl1-1 | 1 | RM1297–RM3530 | 2.70 | −2.1 | 0.3 | Nagdong | ||
| 2019 | qPl1-2 | 1 | RM212–RM1297 | 3.97 | −2.2 | 0.2 | Nagdong | |
| qPl112 | 12 | RM8216–RM1159 | 3.12 | −2.1 | 0.2 | Nagdong | ||
| Number of tillers | 2018 | qNt1 | 1 | RM212–RM1297 | 3.45 | −0.5 | 0.2 | Nagdong |
| 1000 grain weight | 2018 | qTgw1 | 1 | RM212–RM11694 | 3.80 | −0.3 | 0.2 | Nagdong |
| qTgw 12 | 12 | RM8216–RM1159 | 3.23 | −1.2 | 0.1 | Nagdong | ||
| 2019 | qTgw1-1 | 1 | RM212–RM11669 | 3.78 | −0.3 | 0.2 | Nagdong | |
| Content of chlorophyll | 2018 | qCc1 | 1 | RM3709–RM11669 | 3.09 | −3.3 | 0.2 | Nagdong |
| qCc1-1 | 1 | RM11605–RM3530 | 2.98 | −4.6 | 0.3 | Nagdong | ||
| 2019 | qCc1-2 | 1 | RM3709-RM11669 | 2.98 | −3.4 | 0.2 | Nagdong | |
| qCc1-3 | 1 | RM11605–RM3530 | 3.13 | −4.4 | 0.3 | Nagdong |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-R.; Yang, W.-T.; Kim, D.-H.; Kim, K.-M. Identification of a Novel Gene, Osbht, in Response to High Temperature Tolerance at Booting Stage in Rice. Int. J. Mol. Sci. 2020, 21, 5862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165862
Park J-R, Yang W-T, Kim D-H, Kim K-M. Identification of a Novel Gene, Osbht, in Response to High Temperature Tolerance at Booting Stage in Rice. International Journal of Molecular Sciences. 2020; 21(16):5862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165862
Chicago/Turabian StylePark, Jae-Ryoung, Won-Tae Yang, Doh-Hoon Kim, and Kyung-Min Kim. 2020. "Identification of a Novel Gene, Osbht, in Response to High Temperature Tolerance at Booting Stage in Rice" International Journal of Molecular Sciences 21, no. 16: 5862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21165862