Impact of Extracellular Vesicle Isolation Methods on Downstream miRNA Analysis in Semen: A Comparative Study


Abstract
:1. Introduction
2. Results
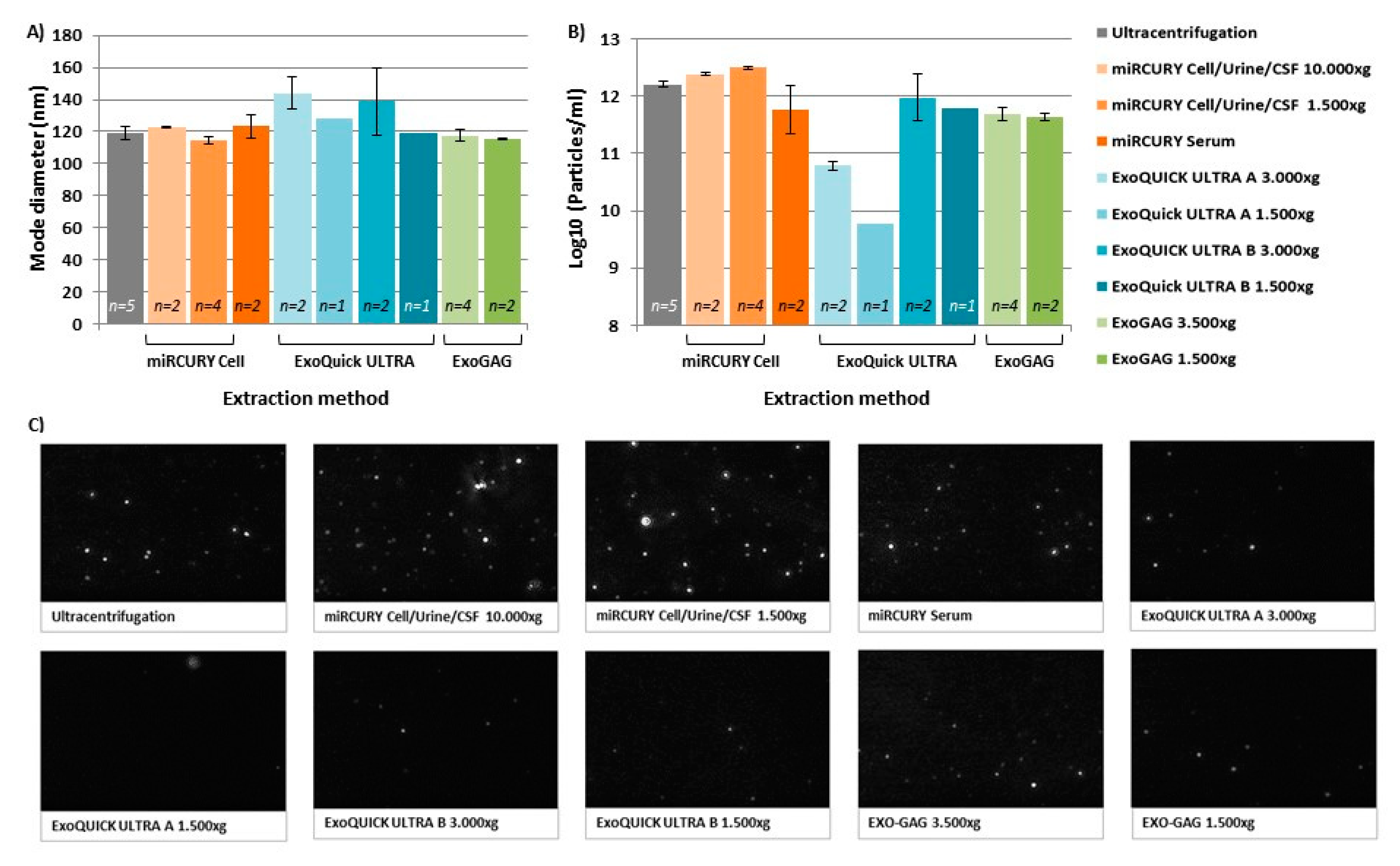
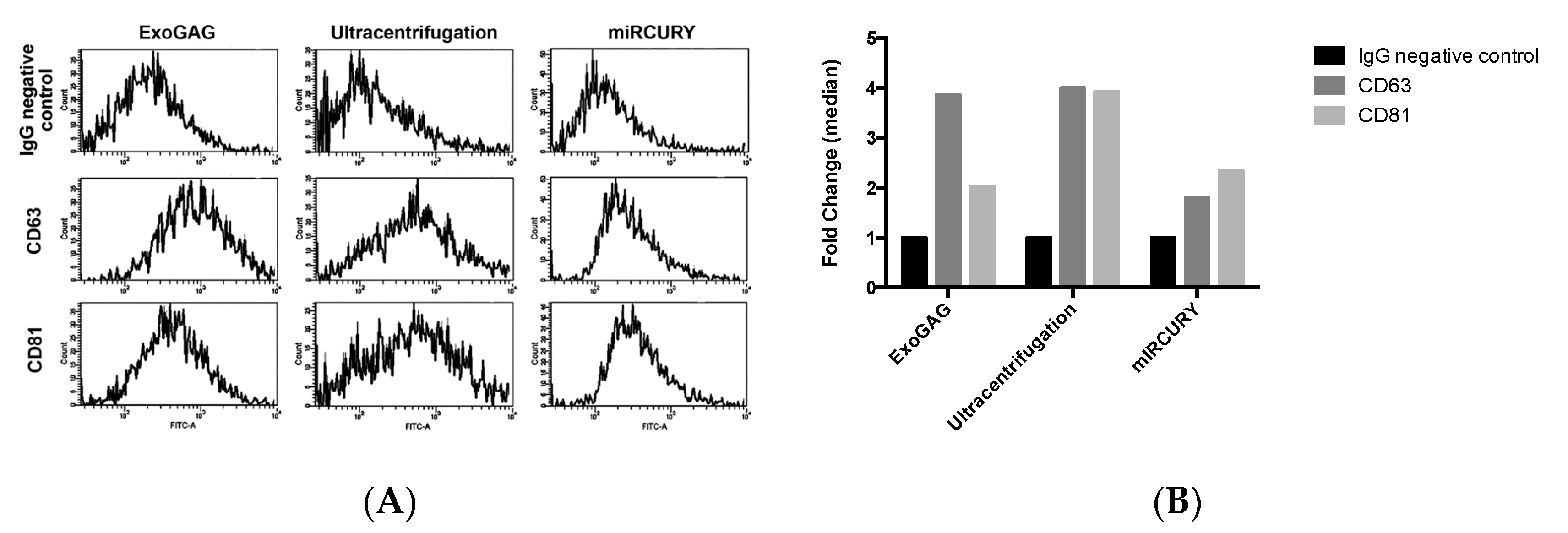
2.1. Characterization of Nanovesicles by Size and Concentration
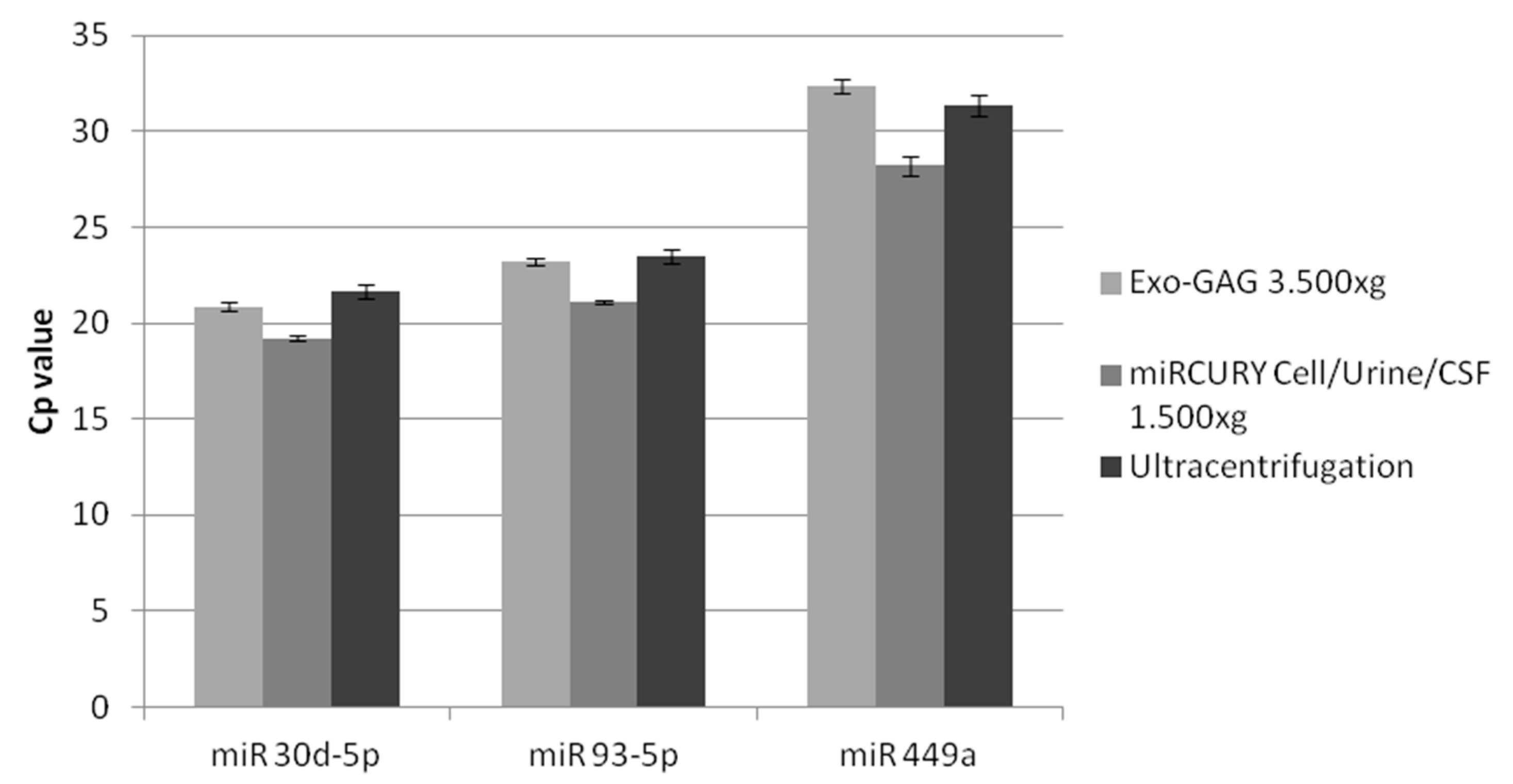
2.2. Nanovesicle RNA Quality and Quantity
2.3. Pre-Study miRNA Expression Pattern
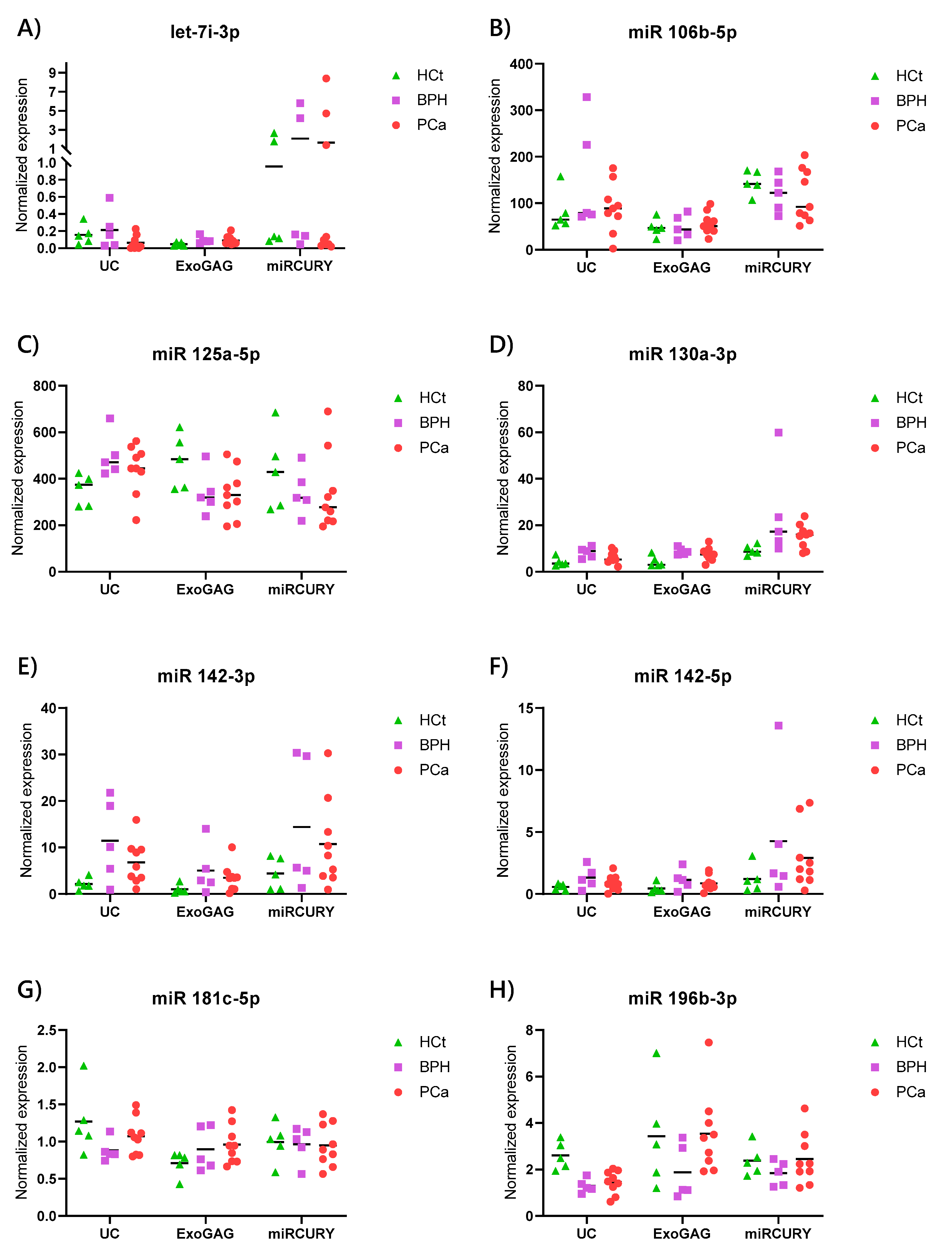
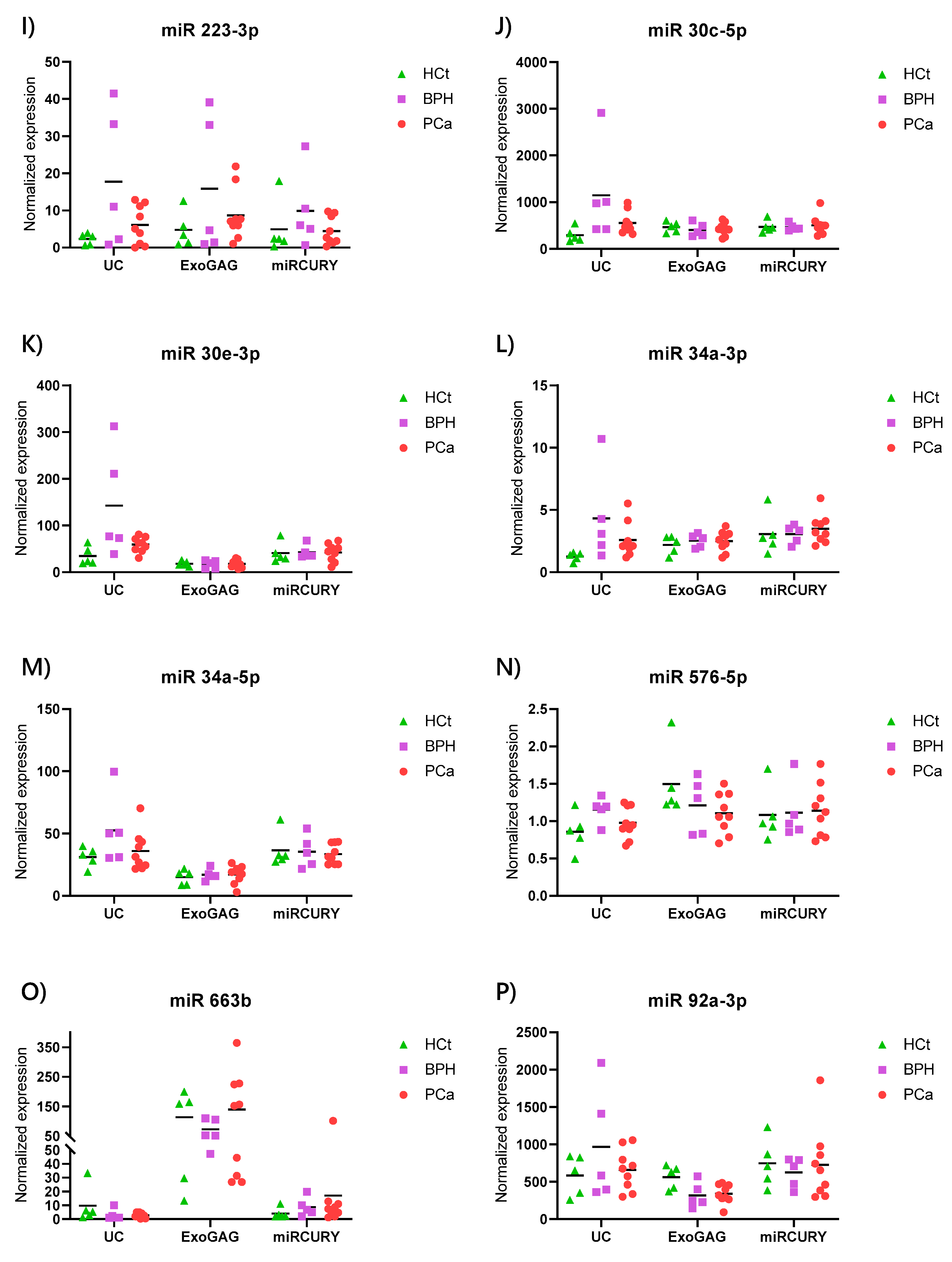
2.4. Exosome/sEV-miRNA Expression Pattern in Prostate Cancer
2.5. Diagnostic Performance of Combined sEV-miRNA-Based Diagnostic Classifiers for PCa
3. Discussion
4. Materials and Methods
4.1. Subjects of Study
4.2. Sample Collection and Processing
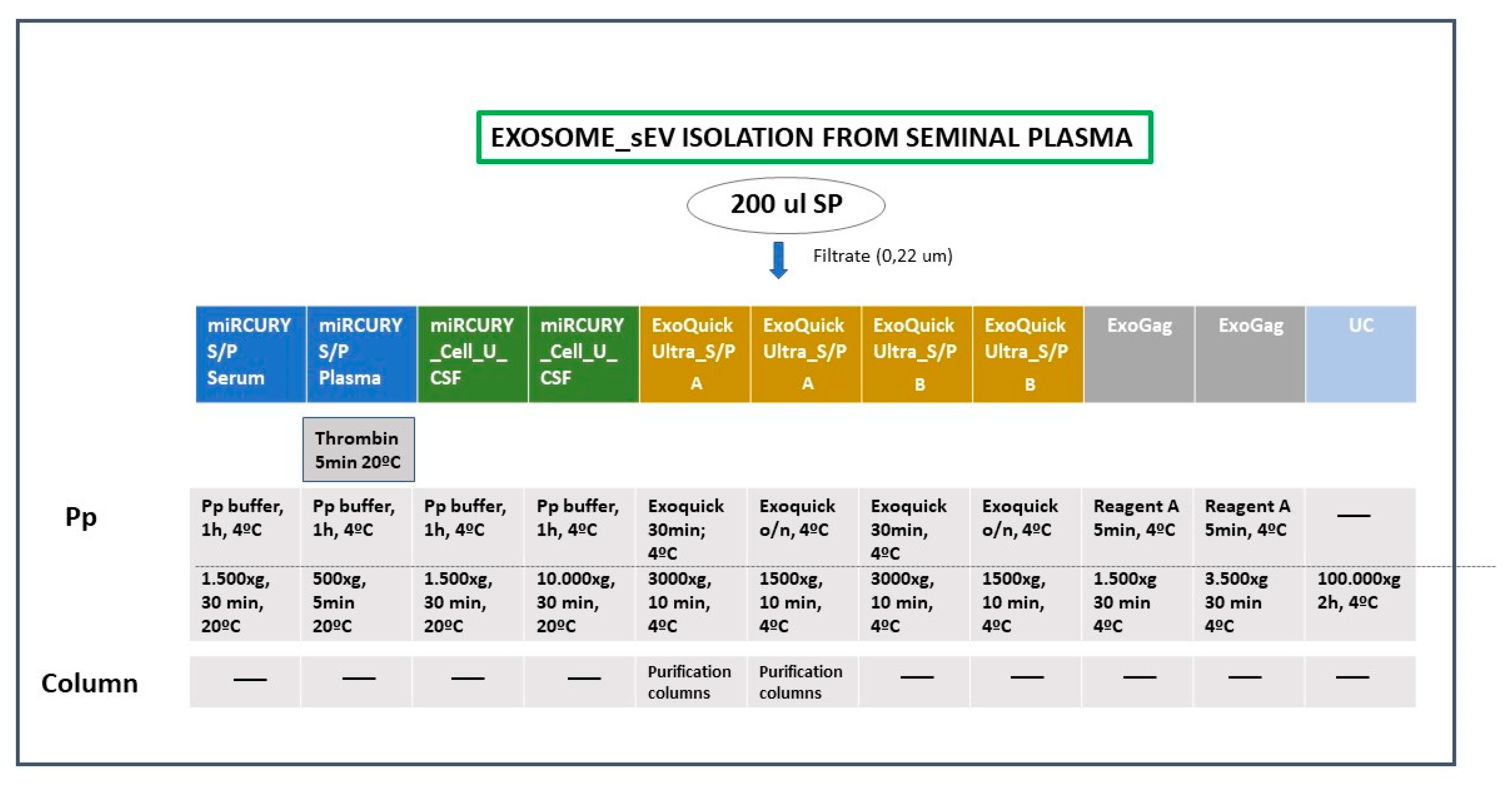
4.3. Exosome/sEV Isolation (Figure 1)
- -
- miRCURY ® Exosome Cell/Urine/CSF kit (miRCURY Cell/Urine/CSF)
- -
- miRCURY ® Exosome Serum/Plasma kit (miRCURY S/P)
- -
- ExoQuick® ULTRA EV Isolation kit for Serum and Plasma (ExoQuick Ultra_S/P)
- -
- ExoGAG
4.4. Characterization of EVs
4.5. Small RNA-Containing Total RNA Isolation from Semen Exosomes/sEVs
4.6. Exosomal miRNA Quantitative Real-Time PCR (qPCR)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BPH | Benign prostate hyperplasia group |
| EV | Extracellular vesicle |
| HCt | Healthy control group |
| mRNA | Messenger RNA |
| miRNA | MicroRNA |
| NTA | Nanoparticle tracking analysis |
| PCa | Prostate cancer |
| PSA | Prostate specific antigen |
| RT-qPCR | Reverse transcription quantitative polymerase chain reaction |
| sEV | Small extracellular vesicle |
| SP | Seminal plasma |
| UC | Ultracentrifugation |
References
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.-J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Wong, W.T.; Wang, N.H. Obesity reshapes stem cell extracellular vesicles. Cytom. Part A 2017, 93, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Barile, L.; Vassalli, G. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 2017, 174, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Renneberg, H.; Konrad, L.; Dammshäuser, I.; Seitz, J.; Aumüller, G. Immunohistochemistry of prostasomes from human semen. Prostate 1997, 30, 98–106. [Google Scholar] [CrossRef]
- Vojtech, L.; Woo, S.; Hughes, S.M.; Levy, C.; Ballweber, L.; Sauteraud, R.P.; Strobl, J.; Westerberg, K.; Gottardo, R.; Tewari, M.; et al. Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res. 2014, 42, 7290–7304. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J. Exosome-Mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Barcelo, M.; Mata, A.; Bassas, L.; Larriba, S. Exosomal microRNAs in seminal plasma are markers of the origin of azoospermia and can predict the presence of sperm in testicular tissue. Hum. Reprod. 2018, 33, 1087–1098. [Google Scholar] [CrossRef]
- Barceló, M.; Castells, M.; Bassas, L.; Vigués, F.; Larriba, S. Semen miRNAs Contained in Exosomes as Non-Invasive Biomarkers for Prostate Cancer Diagnosis. Sci. Rep. 2019, 9, 13772. [Google Scholar] [CrossRef] [Green Version]
- Drabovich, A.P.; Saraon, P.; Jarvi, K.; Diamandis, E.P. Seminal plasma as a diagnostic fluid for male reproductive system disorders. Nat. Rev. Urol. 2014, 11, 278–288. [Google Scholar] [CrossRef]
- Meo, D.A.; Bartlett, J.; Cheng, Y.; Pasic, M.D.; Yousef, G.M. Liquid biopsy: A step forward towards precision medicine in urologic malignancies. Mol. Cancer 2017, 16, 80. [Google Scholar] [CrossRef]
- Filella, X.; Foj, L. Prostate Cancer Detection and Prognosis: From Prostate Specific Antigen (PSA) to Exosomal Biomarkers. Int. J. Mol. Sci. 2016, 17, 1784. [Google Scholar] [CrossRef] [PubMed]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tan, M.; Xiang, Q.; Zhou, Z.; Yan, H.; Lee, T. Thrombin-Activated platelet-derived exosomes regulate endothelial cell expression of ICAM-1 via microRNA-223 during the thrombosis-inflammation response. Thromb. Res. 2017, 154, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Helwa, I.; Cai, J.; Drewry, M.D.; Zimmerman, A.; Dinkins, M.B.; Khaled, M.L.; Seremwe, M.; Dismuke, W.M.; Bieberich, E.; Stamer, W.D.; et al. A Comparative Study of Serum Exosome Isolation Using Differential Ultracentrifugation and Three Commercial Reagents. PLoS ONE 2017, 12, e0170628. [Google Scholar] [CrossRef]
- Tang, Y.-T.; Huang, Y.-Y.; Zheng, L.; Qin, S.-H.; Xu, X.-P.; An, T.-X.; Xu, Y.; Wu, Y.-S.; Hu, X.-M.; Ping, B.-H.; et al. Comparison of isolation methods of exosomes and exosomal RNA from cell culture medium and serum. Int. J. Mol. Med. 2017, 40, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Rekker, K.; Saare, M.; Roost, A.M.; Kubo, A.-L.; Zarovni, N.; Chiesi, A.; Salumets, A.; Peters, M. Comparison of serum exosome isolation methods for microRNA profiling. Clin. Biochem. 2014, 47, 135–138. [Google Scholar] [CrossRef]
- Eldh, M.; Lötvall, J.; Malmhäll, C.; Ekström, K. Importance of RNA isolation methods for analysis of exosomal RNA: Evaluation of different methods. Mol. Immunol. 2012, 50, 278–286. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, Á.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Linares, R.; Tan, S.; Gounou, C.; Arraud, N.; Brisson, A. High-Speed centrifugation induces aggregation of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29509. [Google Scholar] [CrossRef]
- Li, H.; Huang, S.; Guo, C.; Guan, H.; Xiong, C. Cell-Free Seminal mRNA and MicroRNA Exist in Different Forms. PLoS ONE 2012, 7, e34566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, C.; De La Fuente, A.; Casas-Arozamena, C.; Sebastian, V.; Prieto, M.; Arruebo, M.; Abalo, A.; Colas, E.; Moreno-Bueno, G.; Gil-Moreno, A.; et al. Extracellular Vesicles-Based Biomarkers Represent a Promising Liquid Biopsy in Endometrial Cancer. Cancers 2019, 11, 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madison, M.N.; Welch, J.L.; Okeoma, C.M. Isolation of Exosomes from Semen for in vitro Uptake and HIV-1 Infection Assays. Bio-Protocol 2017, 7, e2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Théry, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef] [Green Version]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Skendzel, L.P.; Youden, W.J. Systematic versus random error in laboratory surveys. Am. J. Clin. Pathol. 1970, 54, 448–450. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Study | Study | |||
|---|---|---|---|---|
| Variable | HCt | HCt | BPH | PCa |
| Total, n | 7 | 5 | 5 | 9 |
| Age, mean ± SD | 36.4 ± 6.2 | 42.2 ± 6.3 | 58.6 ± 4.3 | 57 ± 5.8 |
| PSA pre-biopsy (n) | ||||
| ≤10 (ng/mL) | - | - | 5 | 7 |
| >10 (ng/mL) | - | - | 0 | 2 |
| PSA pre-biopsy, mean ± SD (ng/mL) | - | - | 5.5 ± 2.5 | 7.5 ± 3.3 |
| Gleason score -biopsy (n) | ||||
| 6 (3 + 3) | - | - | - | 5 |
| 7 (3 + 4) | - | - | - | 2 |
| 7 (4 + 3) | - | - | - | 1 |
| 8 (4 + 4) | - | - | - | 1 |
| Clinical stage (n) | ||||
| cT1c | - | - | - | 7 |
| cT2a | - | - | - | - |
| cT2c | - | - | - | - |
| cT3a | - | - | - | 2 |
| Vasectomized (n) | - | 5 | 0 | 2 |
| Extraction Method | [RNA] (ng/µL) | 260/280 | 260/230 |
|---|---|---|---|
| Ultracentrifugation (n = 5) | 22.39 ± 7.79 | 1.78 ± 0.17 | 0.60 ± 0.40 |
| ExoGAG 1500× g (n = 2) | 8.17 ± 2.55 | 1.76 ± 0.05 | 0.13 ± 0.05 |
| ExoGAG 3500× g (n = 4) | 12.29 ± 8.6 | 1.79 ± 0.08 | 0.47 ± 0.40 |
| ExoQuick ULTRA A 3000× g (n = 2) | 4.27 ± 0.94 | 1.53 ± 0.04 | 0.39 ± 0.18 |
| ExoQuick ULTRA B 3000× g (n = 2) | 4.25 ± 0.93 | 1.48 ± 0.09 | 0.38 ± 0.08 |
| ExoQuick ULTRA A 1500× g (n = 1) | <2 | 1.52 | 0.51 |
| ExoQuick ULTRA B 1500× g (n = 1) | 7.73 | 1.51 | 0.72 |
| miRCURY cell/urine/CSF 10,000× g (n = 2) | 3.65 ± 3.04 | 1.17 ± 0.01 | 0.10 ± 0.01 |
| miRCURY cell/urine/CSF 1500× g (n = 4) | 31.03 ± 2.94 | 1.63 ± 0.11 | 0.40 ± 0.26 |
| miRCURY Serum (n = 2) | 4.86 ± 3.97 | 1.69 ± 0.09 | 0.54 ± 0.36 |
| A. | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Average Cq ULTRACENTRIFUGATION | p-value | miRNA expression | p-value | ||||||
| Gene Name | HCt | BPH | PCa | CV | (HCt-BPH-PCa) | HCt | BPH | PCa | HCt-PCa |
| let7i-3p | 33.39 | 33.01 | 36.14 | 0.076 | 0.049 | 1 | 1.360 | 0.395 | 0.083 |
| miR 106b-5p | 24.15 | 23.00 | 24.64 | 0.075 | >0.10 | 1 | 1.905 | 1.083 | >0.10 |
| miR 125a-5p | 21.94 | 21.03 | 21.85 | 0.032 | >0.10 | 1 | 1.417 | 1.270 | 0.060 |
| miR 130a-3p | 28.41 | 26.96 | 28.09 | 0.042 | >0.10 | 1 | 1.986 | 1.466 | >0.10 |
| miR 142-3p | 29.51 | 27.10 | 28.19 | 0.056 | 0.059 | 1 | 5.309 | 3.066 | 0.042 |
| miR 142-5p | 31.30 | 29.93 | 31.43 | 0.060 | >0.10 | 1 | 2.291 | 1.443 | >0.10 |
| miR 181c-5p | 30.10 | 30.17 | 30.52 | 0.027 | >0.10 | 1 | 0.694 | 0.845 | >0.10 |
| miR 196b-3p | 29.03 | 29.63 | 30.14 | 0.024 | 0.014 | 1 | 0.498 | 0.562 | 0.004 |
| miR 223-3p | 29.54 | 27.02 | 29.69 | 0.105 | >0.10 | 1 | 7.742 | 2.627 | >0.10 |
| miR 30c-5p | 22.31 | 20.20 | 21.57 | 0.059 | 0.038 | 1 | 3.904 | 1.892 | 0.042 |
| miR 30e-3p | 25.45 | 23.22 | 24.75 | 0.047 | 0.007 | 1 | 4.139 | 1.746 | 0.060 |
| miR 34a-3p | 30.11 | 28.24 | 29.38 | 0.041 | 0.050 | 1 | 3.452 | 2.080 | 0.019 |
| miR 34a-5p | 25.46 | 24.40 | 25.52 | 0.037 | 0.027 | 1 | 1.682 | 1.172 | >0.10 |
| miR 576-5p | 30.66 | 29.78 | 30.65 | 0.026 | 0.055 | 1 | 1.348 | 1.134 | >0.10 |
| miR 663b | 28.03 | 29.03 | 29.68 | 0.045 | 0.078 | 1 | 0.318 | 0.267 | >0.10 |
| miR 92a-3p | 21.34 | 20.41 | 21.34 | 0.037 | 0.034 | 1 | 1.657 | 1.169 | >0.10 |
| miR (576+181) | 0.024 | ||||||||
| B. | |||||||||
| Average Cq miRCURY cell/urine/CSF | p-value | miRNA expression | p-value | ||||||
| Gene Name | HCt | BPH | PCa | CV | (HCt-BPH-PCa) | HCt | BPH | PCa | HCt-PCa |
| let7i-3p | 31.43 | 30.75 | 32.27 | 0.110 | >0.10 | 1 | 2.17 | 1.60 | >0.10 |
| miR 106b-5p | 22.81 | 22.86 | 23.32 | 0.050 | >0.10 | 1 | 0.82 | 0.79 | >0.10 |
| miR 125a-5p | 21.30 | 21.32 | 21.74 | 0.031 | >0.10 | 1 | 0.80 | 0.79 | >0.10 |
| miR 130a-3p | 26.78 | 25.38 | 26.18 | 0.041 | 0.076 | 1 | 2.66 | 1.62 | 0.060 |
| miR 142-3p | 28.34 | 26.69 | 27.19 | 0.069 | >0.10 | 1 | 3.26 | 2.44 | >0.10 |
| miR 142-5p | 30.16 | 28.45 | 29.02 | 0.058 | >0.10 | 1 | 3.52 | 2.41 | >0.10 |
| miR 181c-5p | 30.02 | 29.80 | 30.16 | 0.035 | >0.10 | 1 | 0.97 | 0.96 | >0.10 |
| miR 196b-3p | 28.76 | 28.87 | 28.86 | 0.027 | >0.10 | 1 | 0.77 | 1.04 | >0.10 |
| miR 223-3p | 28.81 | 27.21 | 28.52 | 0.072 | >0.10 | 1 | 2.01 | 0.90 | >0.10 |
| miR 30c-5p | 21.12 | 20.85 | 21.14 | 0.042 | >0.10 | 1 | 0.98 | 1.05 | >0.10 |
| miR 30e-3p | 24.75 | 24.34 | 24.82 | 0.037 | >0.10 | 1 | 1.04 | 1.02 | >0.10 |
| miR 34a-3p | 28.50 | 28.13 | 28.30 | 0.031 | >0.10 | 1 | 1.00 | 1.12 | >0.10 |
| miR 34a-5p | 24.84 | 24.63 | 25.00 | 0.036 | >0.10 | 1 | 0.97 | 0.91 | >0.10 |
| miR 576-5p | 29.91 | 29.60 | 29.90 | 0.029 | >0.10 | 1 | 1.03 | 1.05 | >0.10 |
| miR 663b | 28.33 | 26.97 | 27.19 | 0.043 | 0.077 | 1 | 2.15 | 4.53 | >0.10 |
| miR 92a-3p | 20.53 | 20.48 | 20.77 | 0.031 | >0.10 | 1 | 0.84 | 0.98 | >0.10 |
| miR (576+181) | 0.029 | ||||||||
| C. | |||||||||
| Average Cq EXOGAG | p-value | miRNA expression | p-value | ||||||
| Gene Name | HCt | BPH | PCa | CV | (HCt-BPH-PCa) | HCt | BPH | PCa | HCt-PCa |
| let7i-3p | 35.24 | 34.04 | 34.69 | 0.026 | 0.074 | 1 | 2.01 | 1.90 | >0.10 |
| miR 106b-5p | 25.25 | 25.09 | 25.34 | 0.043 | >0.10 | 1 | 1.04 | 1.17 | >0.10 |
| miR 125a-5p | 21.86 | 22.18 | 22.73 | 0.029 | 0.054 | 1 | 0.71 | 0.72 | 0.060 |
| miR 130a-3p | 28.74 | 27.42 | 28.25 | 0.035 | >0.10 | 1 | 2.03 | 1.69 | 0.083 |
| miR 142-3p | 31.23 | 28.99 | 29.88 | 0.057 | 0.038 | 1 | 5.05 | 3.34 | 0.042 |
| miR 142-5p | 32.27 | 30.78 | 31.72 | 0.043 | >0.10 | 1 | 2.59 | 1.93 | >0.10 |
| miR 181c-5p | 31.25 | 30.77 | 31.17 | 0.022 | >0.10 | 1 | 1.27 | 1.37 | >0.10 |
| miR 196b-3p | 29.19 | 29.87 | 29.37 | 0.031 | >0.10 | 1 | 0.55 | 1.05 | >0.10 |
| miR 223-3p | 29.08 | 27.94 | 28.41 | 0.053 | >0.10 | 1 | 3.31 | 1.83 | >0.10 |
| miR 30c-5p | 21.89 | 21.95 | 22.41 | 0.038 | >0.10 | 1 | 0.87 | 0.90 | >0.10 |
| miR 30e-3p | 26.58 | 26.66 | 27.04 | 0.039 | >0.10 | 1 | 0.93 | 0.96 | >0.10 |
| miR 34a-3p | 29.66 | 29.23 | 29.83 | 0.029 | >0.10 | 1 | 1.15 | 1.12 | >0.10 |
| miR 34a-5p | 26.91 | 26.50 | 27.17 | 0.044 | >0.10 | 1 | 1.13 | 1.11 | >0.10 |
| miR 576-5p | 30.18 | 30.33 | 30.96 | 0.028 | >0.10 | 1 | 0.81 | 0.73 | >0.10 |
| miR 663b | 24.53 | 24.46 | 24.55 | 0.043 | >0.10 | 1 | 0.65 | 1.25 | >0.10 |
| miR 92a-3p | 21.63 | 22.40 | 22.79 | 0.041 | 0.045 | 1 | 0.57 | 0.61 | 0.042 |
| miR (576+181) | 0.022 | ||||||||
| PCa vs. non-PCa (HCt+BPH) | [PSA] model | [miR-142-3p+miR-142-5p] model | [miR-142-3p+miR-142-5p+PSA] model | ||||
| UC | miRCURY Cell_U_CSF | ExoGAG | UC | miRCURY Cell_U_CSF | ExoGAG | ||
| AUC | 0.844 | 0.556 | 0.644 | 0.667 | 0.856 | 0.867 | 0.922 |
| 95% CI | 0.668–1.021 | 0.283–0.828 | 0.385–0.904 | 0.409–0.924 | 0.661–1.050 | 0.681–1.053 | 0.804–1.040 |
| p-value | 0.011 | 0.683 | 0.288 | 0.221 | 0.009 | 0.007 | 0.002 |
| Sensitivity (%) | 66.7 | 22.2 | 22.2 | 11.1 | 77.8 | 77.8 | 66.7 |
| Specificity (%) | 80.0 | 90.0 | 90.0 | 90.0 | 90.0 | 90.0 | 90.0 |
| Positive predictive value | 75.0 | 66.7 | 66.7 | 50.0 | 87.5 | 87.5 | 85.7 |
| Negative predictive value | 72.7 | 56.3 | 56.3 | 52.9 | 81.8 | 81.8 | 75.0 |
| PCa vs. BPH | [PSA] model | [miR-142-3p+miR-142-5p+miR-223-3p] model | [miR-142-3p+miR-142-5p+miR-223-3p+PSA] model | ||||
| UC | miRCURY Cell_U_CSF | ExoGAG | UC | miRCURY Cell_U_CSF | ExoGAG | ||
| AUC | 0.689 | 0.644 | 0.911 | 0.467 | 0.933 | 1 | 1 |
| 95% CI | 0.398–0.979 | 0.309–0.980 | 0.754–1.069 | 0.066–0.867 | 0.790–1.076 | 1.000–1.000 | 1.000–1.000 |
| p-value | 0.257 | 0.386 | 0.014 | 0.841 | 0.009 | 0.003 | 0.003 |
| Sensitivity | 100.0 | 100.0 | 88.9 | 100.0 | 100.0 | 100.0 | 100.0 |
| Specificity | 20.0 | 40.0 | 80.0 | 40.0 | 80.0 | 100.0 | 100.0 |
| Positive predictive value | 100.0 | 75.0 | 88.9 | 75.0 | 90.0 | 100.0 | 100.0 |
| Negative predictive value | 69.2 | 100.0 | 80.0 | 100.0 | 100.0 | 100.0 | 100.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercadal, M.; Herrero, C.; López-Rodrigo, O.; Castells, M.; de la Fuente, A.; Vigués, F.; Bassas, L.; Larriba, S. Impact of Extracellular Vesicle Isolation Methods on Downstream miRNA Analysis in Semen: A Comparative Study. Int. J. Mol. Sci. 2020, 21, 5949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175949
Mercadal M, Herrero C, López-Rodrigo O, Castells M, de la Fuente A, Vigués F, Bassas L, Larriba S. Impact of Extracellular Vesicle Isolation Methods on Downstream miRNA Analysis in Semen: A Comparative Study. International Journal of Molecular Sciences. 2020; 21(17):5949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175949
Chicago/Turabian StyleMercadal, Marina, Carolina Herrero, Olga López-Rodrigo, Manel Castells, Alexandre de la Fuente, Francesc Vigués, Lluís Bassas, and Sara Larriba. 2020. "Impact of Extracellular Vesicle Isolation Methods on Downstream miRNA Analysis in Semen: A Comparative Study" International Journal of Molecular Sciences 21, no. 17: 5949. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175949