Novel Insights into the Biochemical Mechanism of CK1ε and its Functional Interplay with DDX3X


Abstract
:1. Introduction
2. Results
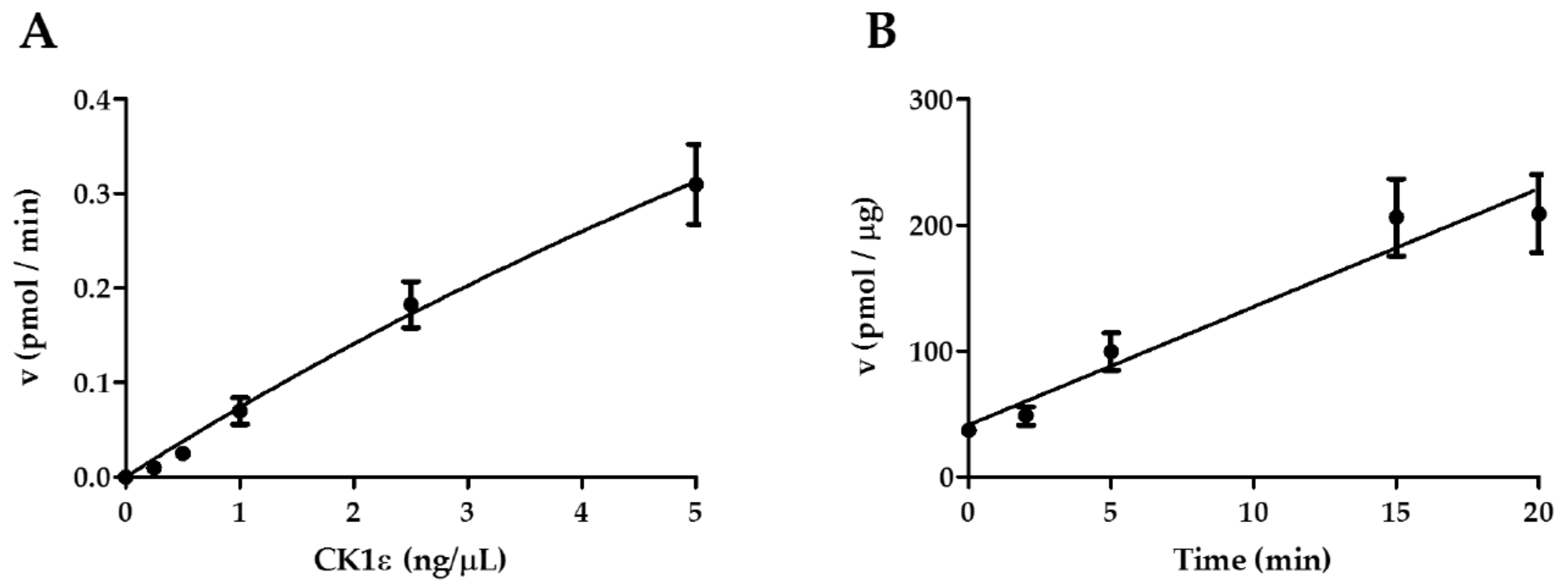
2.1. Determination of the Optimal Initial Conditions of the Reaction Catalyzed by CK1ε
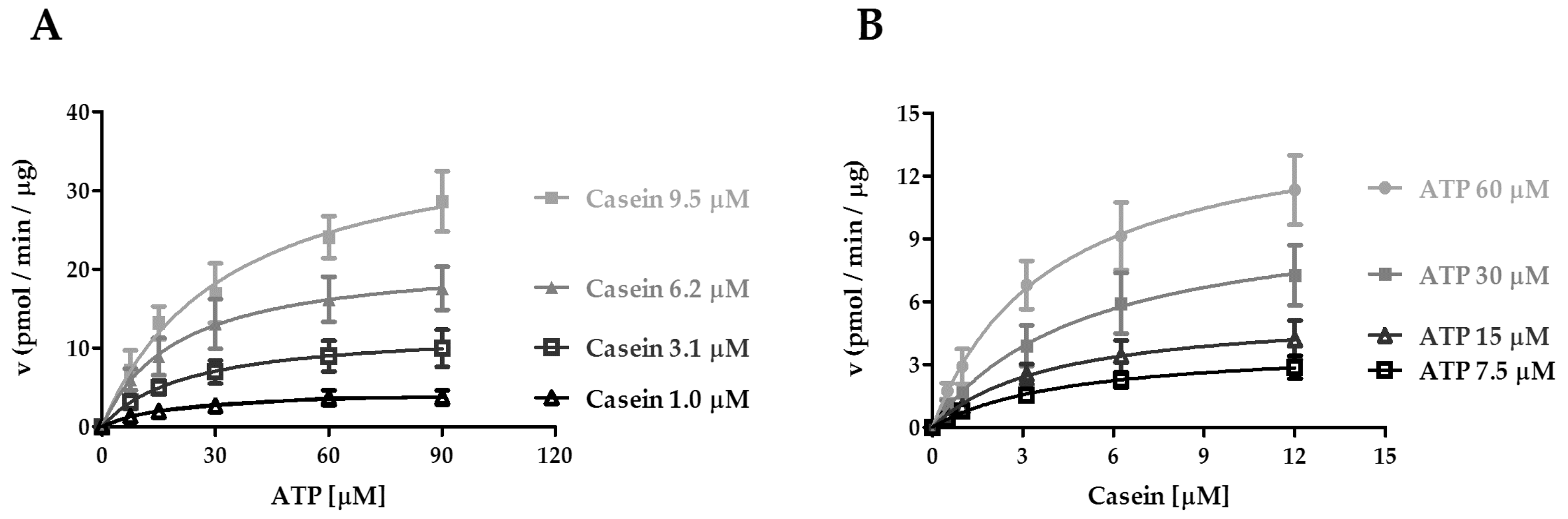
2.2. Kinetic Analysis of CK1ε Acting on ATP and Casein
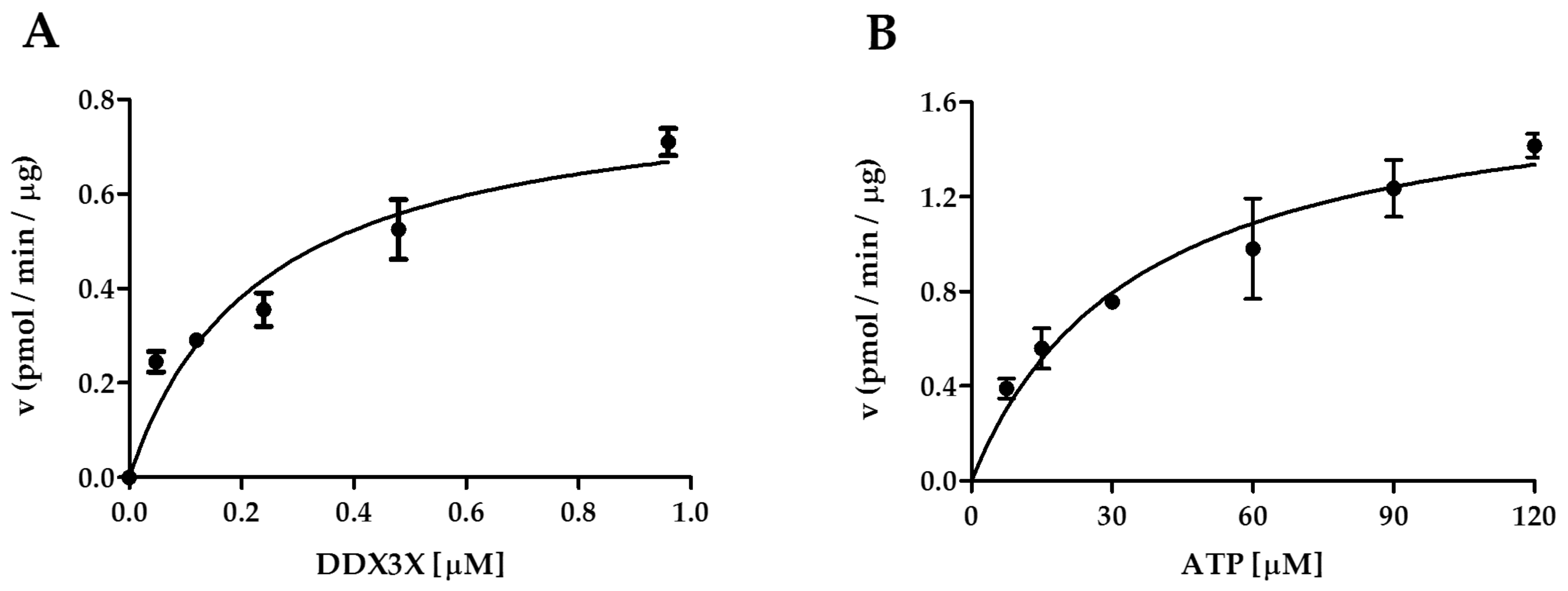
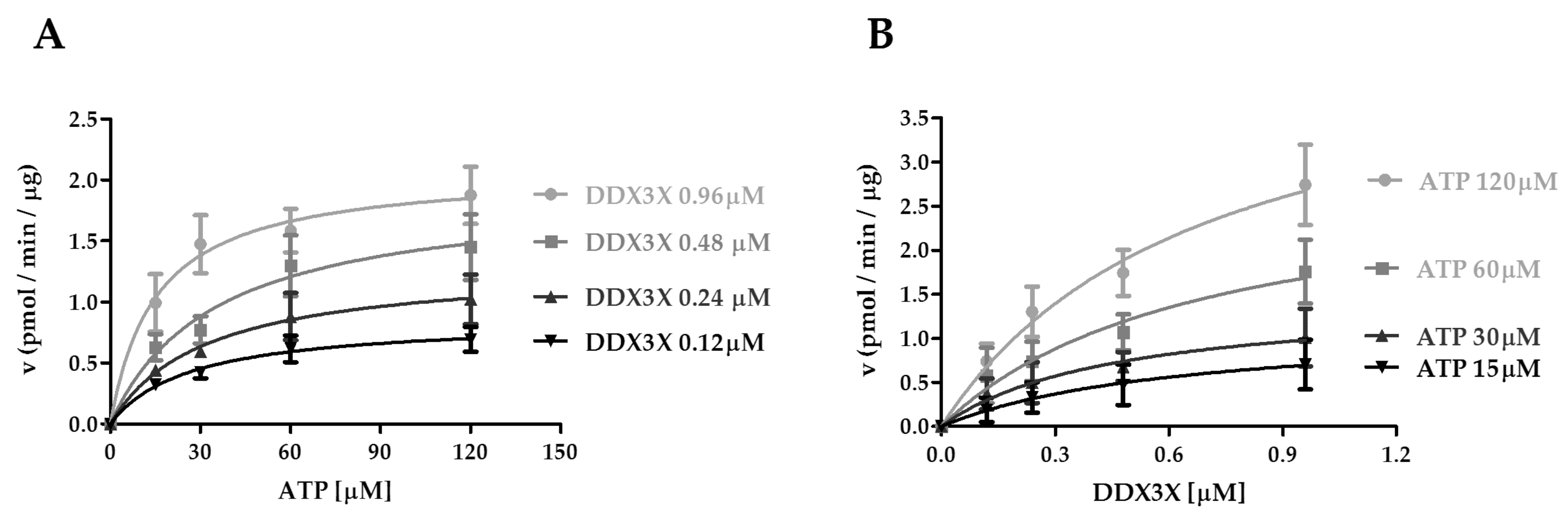
2.3. Interplay of CK1ε and the Human DDX3X Recombinant Protein in the Phosphorylation Reaction
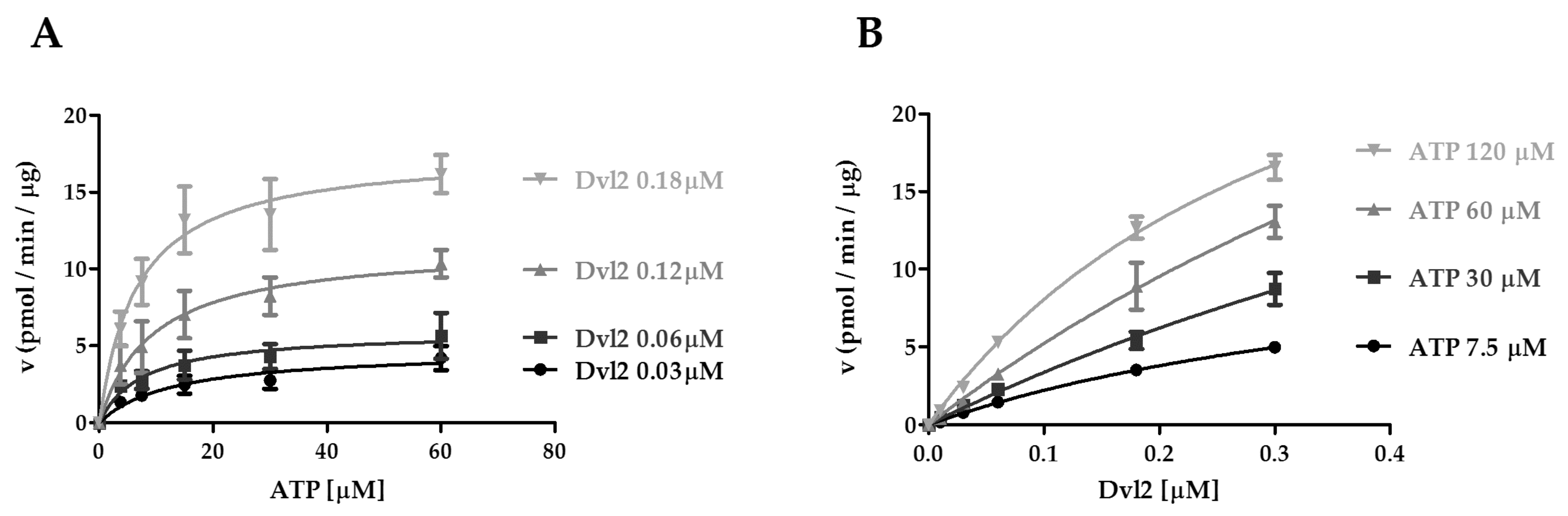
2.4. Kinetic Analysis of CK1ε Acting on ATP and the Dvl2 Protein
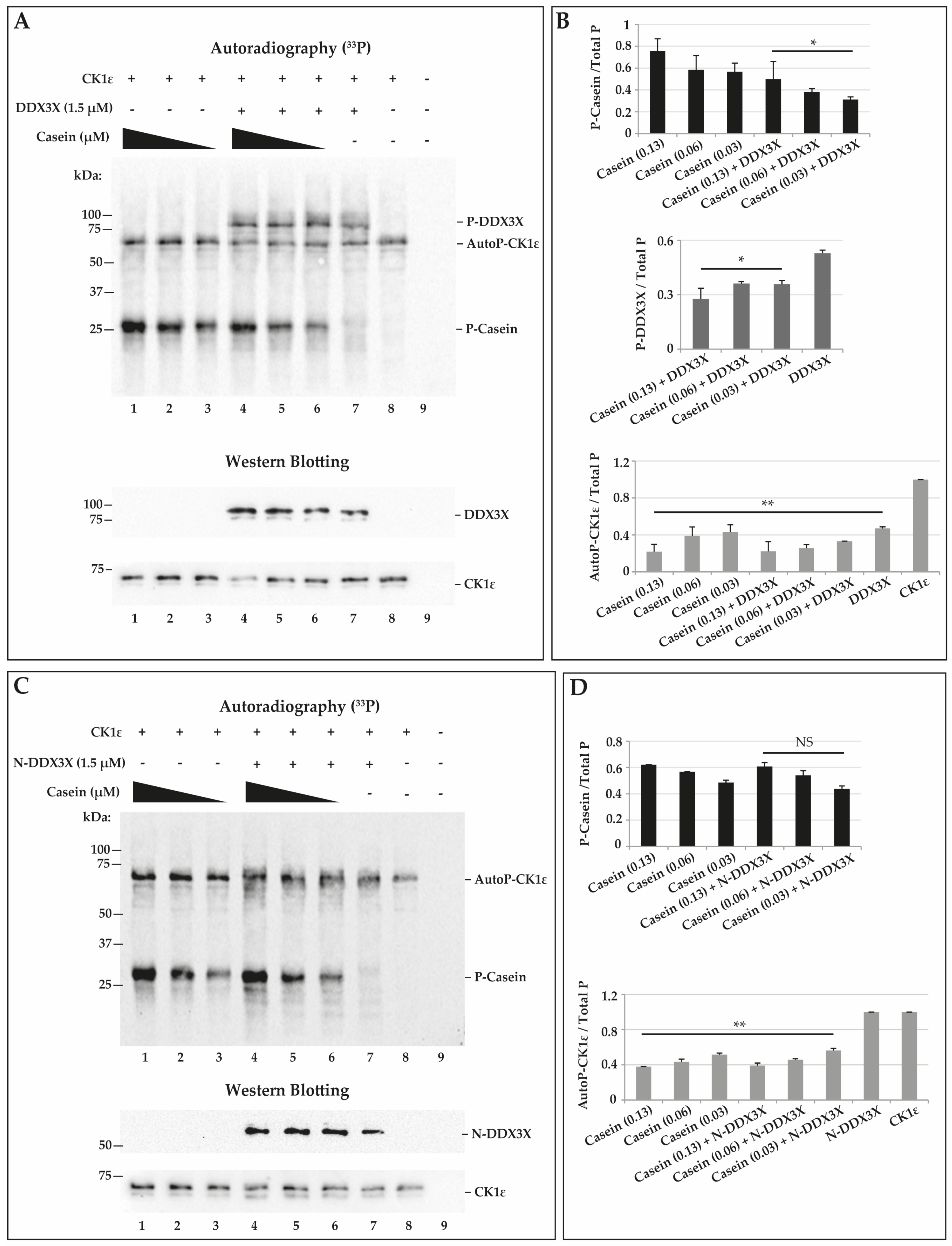
2.5. CK1ε Phosphorylates Casein and DDX3X in a Competitive Way
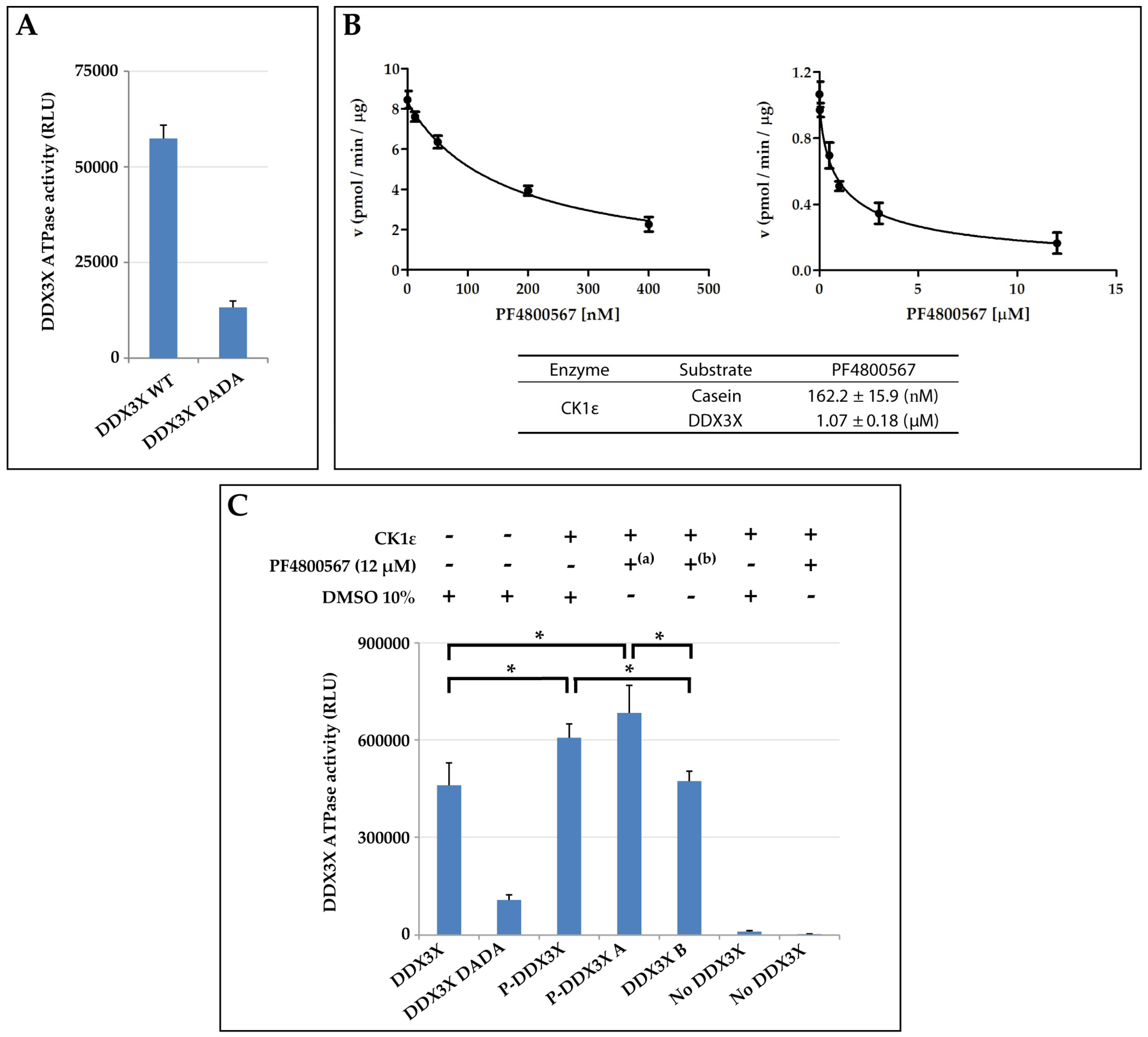
2.6. CK1ε Positively Modulates the DDX3X ATPase Activity
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Enzymes and Proteins
4.3. Kinase Activity Assay
4.4. Kinetic Analysis
4.5. Autoradiography Assay
4.6. Western Blot Analysis
4.7. ATPase Activity Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Wnt | Wingless type family protein |
| Km | Michaelis constant |
| Vmax | maximal velocity of the enzymatic reaction |
| PMSF | phenylmethylsulfonyl fluoride |
| MOPS | 3-(N-morpholino)propanesulfonic acid) |
| EGTA | ethylene glycol tetraacetic acid |
| EDTA | Ethylenediaminetetraacetic acid |
| SDS | Sodium dodecyl sulfate |
References
- Gross, S.D.; Anderson, R.A. Casein kinase I: Spatial organization and positioning of a multifunctional protein kinase family. Cell Signal. 1998, 10, 699–711. [Google Scholar] [CrossRef]
- Knippschild, U.; Wolff, S.; Giamas, G.; Brockschmidt, C.; Wittau, M.; Würl, P.U.; Eismann, T.; Stöter, M. The role of the casein kinase 1 (CK1) family in different signaling pathways linked to cancer development. Onkologie 2005, 28, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.; Virshup, D.M. Post-translational modifications regulate the ticking of the circadian clock. Nat. Rev. Mol. Cell Biol. 2007, 8, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Cheong, J.K.; Virshup, D.M. Casein kinase 1: Complexity in the family. Int. J. Biochem. Cell Biol. 2011, 43, 465–469. [Google Scholar] [CrossRef]
- Schittek, B.; Sinnberg, T. Biological functions of casein kinase 1 isoforms and putative roles in tumorigenesis. Mol. Cancer 2014, 13, 231. [Google Scholar] [CrossRef] [Green Version]
- Burnett, G.; Kennedy, E.P. The enzymatic phosphorylation of proteins. J. Biol. Chem. 1954, 211, 969–980. [Google Scholar]
- Budini, M.; Jacob, G.; Jedlicki, A.; Pérez, C.; Allende, C.C.; Allende, J.E. Autophosphorylation of carboxy-terminal residues inhibits the activity of protein kinase CK1alpha. J. Cell. Biochem. 2009, 106, 399–408. [Google Scholar] [CrossRef]
- Knippschild, U.; Krüger, M.; Richter, J.; Xu, P.; García-Reyes, B.; Peifer, C.; Halekotte, J.; Bakulev, V.; Bischof, J. The CK1 Family: Contribution to Cellular Stress Response and Its Role in Carcinogenesis. Front. Oncol. 2014, 4, 96. [Google Scholar] [CrossRef] [Green Version]
- Vancura, A.; Sessler, A.; Leichus, B.; Kuret, J. A prenylation motif is required for plasma membrane localization and biochemical function of casein kinase I in budding yeast. J. Biol. Chem. 1994, 269, 19271–19278. [Google Scholar]
- Gietzen, K.F.; Virshup, D.M. Identification of inhibitory autophosphorylation sites in casein kinase I epsilon. J. Biol. Chem. 1999, 274, 32063–32070. [Google Scholar] [CrossRef] [Green Version]
- Venerando, A.; Ruzzene, M.; Pinna, L.A. Casein kinase: The triple meaning of a misnomer. The Biochem. J. 2014, 460, 141–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, D.I.; Gil, C.; Martinez, A. Protein kinases CK1 and CK2 as new targets for neurodegenerative diseases. Med. Res. Rev. 2011, 31, 924–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milne, D.M.; Looby, P.; Meek, D.W. Catalytic activity of protein kinase CK1 delta (casein kinase 1delta) is essential for its normal subcellular localization. Exp. Cell Res. 2001, 263, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, T.; Zhang, Y.; Qin, X. Molecular basis for the regulation of the circadian clock kinases CK1δ and CK1ε. Cell Signal. 2017, 31, 58–65. [Google Scholar] [CrossRef]
- Foldynová-Trantírková, S.; Sekyrová, P.; Tmejová, K.; Brumovská, E.; Bernatík, O.; Blankenfeldt, W.; Krejcí, P.; Kozubík, A.; Dolezal, T.; Trantírek, L.; et al. Breast cancer-specific mutations in CK1epsilon inhibit Wnt/beta-catenin and activate the Wnt/Rac1/JNK and NFAT pathways to decrease cell adhesion and promote cell migration. Breast Cancer Res. 2010, 12, R30. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, N.; Yang, J.; Hasselblatt, K.; Liu, S.; Zhou, Y.; Rauh-Hain, J.A.; Ng, S.K.; Choi, P.W.; Fong, W.P.; Agar, N.Y.; et al. Casein kinase I epsilon interacts with mitochondrial proteins for the growth and survival of human ovarian cancer cells. EMBO Mol. Med. 2012, 4, 952–963. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, Q.; Wang, Q.; Liu, H.; Zhou, F.; Zhang, Y.; Yuan, M.; Zhao, C.; Guan, Y.; Wang, X. DDX3 binding with CK1ε was closely related to motor neuron degeneration of ALS by affecting neurite outgrowth. Am. J. Transl. Res. 2017, 9, 4627–4639. [Google Scholar]
- Guan, H.; Zhu, T.; Wu, S.; Liu, S.; Liu, B.; Wu, J.; Cai, J.; Zhu, X.; Zhang, X.; Zeng, M.; et al. Long noncoding RNA LINC00673-v4 promotes aggressiveness of lung adenocarcinoma via activating WNT/β-catenin signaling. Proc. Natl. Acad. Sci. USA 2019, 116, 14019–14028. [Google Scholar] [CrossRef] [Green Version]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Bryja, V.; Schulte, G.; Rawal, N.; Grahn, A.; Arenas, E. Wnt-5a induces Dishevelled phosphorylation and dopaminergic differentiation via a CK1-dependent mechanism. J. Cell Sci. 2007, 120 Pt 4, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Dunn, I.F.; Firestein, R.; Gupta, P.; Wardwell, L.; Repich, K.; Schinzel, A.C.; Wittner, B.; Silver, S.J.; Root, D.E.; et al. CK1epsilon is required for breast cancers dependent on beta-catenin activity. PLoS ONE 2010, 5, e8979. [Google Scholar]
- He, T.Y.; Wu, D.W.; Lin, P.L.; Wang, L.; Huang, C.C.; Chou, M.C.; Lee, H. DDX3 promotes tumor invasion in colorectal cancer via the CK1ε/Dvl2 axis. Sci. Rep. 2016, 6, 21483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, A.; Rinkevich, B. The DDX3 subfamily of the DEAD box helicases: Divergent roles as unveiled by studying different organisms and in vitro assays. Curr. Med. Chem. 2007, 14, 2517–2525. [Google Scholar] [CrossRef]
- Schröder, M. Human DEAD-box protein 3 has multiple functions in gene regulation and cell cycle control and is a prime target for viral manipulation. Biochem. Pharmacol. 2010, 79, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Riva, V.; Maga, G. From the magic bullet to the magic target: Exploiting the diverse roles of DDX3X in viral infections and tumorigenesis. Future Med. Chem. 2019, 11, 1357–1381. [Google Scholar] [CrossRef] [PubMed]
- Cruciat, C.M.; Dolde, C.; de Groot, R.E.; Ohkawara, B.; Reinhard, C.; Korswagen, H.C.; Niehrs, C. RNA helicase DDX3 is a regulatory subunit of casein kinase 1 in Wnt-β-catenin signaling. Science 2013, 339, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Lin, Y.M.; Yeh, C.M.; Chen, C.J.; Chen, M.W.; Hung, H.F.; Yeh, K.T.; Yang, S.F. Casein kinase 1 epsilon expression predicts poorer prognosis in low T-stage oral cancer patients. Int. J. Mol. Sci. 2014, 15, 2876–2891. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.H.; Yeh, C.M.; Hsieh, M.J.; Lin, Y.M.; Chen, M.W.; Chen, C.J.; Lin, C.Y.; Hung, H.F.; Yeh, K.T.; Yang, S.F. Low cytoplasmic casein kinase 1 epsilon expression predicts poor prognosis in patients with hepatocellular carcinoma. Tumour Biol. 2016, 37, 3997–4005. [Google Scholar] [CrossRef]
- Brai, A.; Martelli, F.; Riva, V.; Garbelli, A.; Fazi, R.; Zamperini, C.; Pollutri, A.; Falsitta, L.; Ronzini, S.; Maccari, L.; et al. DDX3X Helicase Inhibitors as a New Strategy To Fight the West Nile Virus Infection. J. Med. Chem. 2019, 62, 2333–2347. [Google Scholar] [CrossRef]
- Sharma, M.; Castro-Piedras, I.; Simmons, G.E., Jr.; Pruitt, K. Dishevelled: A masterful conductor of complex Wnt signals. Cell Signal. 2018, 47, 52–64. [Google Scholar] [CrossRef]
- Dolde, C.; Bischof, J.; Grüter, S.; Montada, A.; Halekotte, J.; Peifer, C.; Kalbacher, H.; Baumann, U.; Knippschild, U.; Suter, B. A CK1 FRET biosensor reveals that DDX3X is an essential activator of CK1ε. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbelli, A.; Beermann, S.; Di Cicco, G.; Dietrich, U.; Maga, G. A motif unique to the human DEAD-box protein DDX3 is important for nucleic acid binding, ATP hydrolysis, RNA/DNA unwinding and HIV-1 replication. PLoS ONE 2011, 6, e19810. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.M.; Fisher, K.; Rubitski, D.; Marconi, M.; Meng, Q.J.; Sládek, M.; Adams, J.; Bass, M.; Chandrasekaran, R.; Butler, T.; et al. Selective inhibition of casein kinase 1 epsilon minimally alters circadian clock period. J. Pharmacol. Exp. Ther. 2009, 330, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.J.; Maywood, E.S.; Bechtold, D.A.; Lu, W.Q.; Li, J.; Gibbs, J.E.; Dupré, S.M.; Chesham, J.E.; Rajamohan, F.; Knafels, J.; et al. Entrainment of disrupted circadian behavior through inhibition of casein kinase 1 (CK1) enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 15240–15245. [Google Scholar] [CrossRef] [Green Version]
- Long, A.M.; Zhao, H.; Huang, X. Structural basis for the potent and selective inhibition of casein kinase 1 epsilon. J. Med. Chem. 2012, 55, 10307–10311. [Google Scholar] [CrossRef]
- Toyoshima, M.; Howie, H.L.; Imakura, M.; Walsh, R.M.; Annis, J.E.; Chang, A.N.; Frazier, J.; Chau, B.N.; Loboda, A.; Linsley, P.S.; et al. Functional genomics identifies therapeutic targets for MYC-driven cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 9545–9550. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.; Ullah, K.; Xu, P.; Alscher, V.; Blatz, A.; Peifer, C.; Halekotte, J.; Leban, J.; Vitt, D.; Holzmann, K.; et al. Effects of altered expression and activity levels of CK1δ and ɛ on tumor growth and survival of colorectal cancer patients. Int. J. Cancer 2015, 136, 2799–2810. [Google Scholar] [CrossRef] [Green Version]
- Varghese, R.T.; Young, S.; Pham, L.; Liang, Y.; Pridham, K.J.; Guo, S.; Murphy, S.; Kelly, D.F.; Sheng, Z. Casein Kinase 1 Epsilon Regulates Glioblastoma Cell Survival. Sci. Rep. 2018, 8, 13621. [Google Scholar] [CrossRef]
- Whitehouse, S.; Feramisco, J.R.; Casnellie, J.E.; Krebs, E.G.; Walsh, D.A. Studies on the kinetic mechanism of the catalytic subunit of the cAMP-dependent protein kinase. J. Biol. Chem. 1983, 258, 3693–3701. [Google Scholar]
- Zhang, X.; Zhang, S.; Yamane, H.; Wahl, R.; Ali, A.; Lofgren, J.A.; Kendall, R.L. Kinetic mechanism of AKT/PKB enzyme family. J. Biol. Chem. 2006, 281, 13949–13956. [Google Scholar] [CrossRef] [Green Version]
- Soulat, D.; Bürckstümmer, T.; Westermayer, S.; Goncalves, A.; Bauch, A.; Stefanovic, A.; Hantschel, O.; Bennett, K.L.; Decker, T.; Superti-Furga, G. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008, 27, 2135–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Fullam, A.; Brennan, R.; Schröder, M. Human DEAD box helicase 3 couples IκB kinase ε to interferon regulatory factor 3 activation. Mol. Cell. Biol. 2013, 33, 2004–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocak, S.; Linder, P. DEAD-box proteins: The driving forces behind RNA metabolism. Nat. Rev. Mol. Cell Biol. 2004, 5, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Ji, X. The mechanism of RNA duplex recognition and unwinding by DEAD-box helicase DDX3X. Nat. Commun. 2019, 10, 3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franca, R.; Belfiore, A.; Spadari, S.; Maga, G. Human DEAD-box ATPase DDX3 shows a relaxed nucleoside substrate specificity. Proteins 2007, 67, 1128–1137. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Casein [μM] | 1 | 3.1 | 6.2 | 9.5 |
|---|---|---|---|---|
| VmaxATP(pmol min−1μg−1) | 4.7 ± 0.7 a | 12.4 ± 2.3 a | 21.6 ± 3.6 a | 38.2 ± 5.6 a |
| KmATP[μM] | 21.1 ± 6.6 | 20.5 ± 1.2 | 21.1 ± 3.7 | 35.9 ± 7.9 |
| kcat(min−1) | 13.5 ± 2.1 | 35.7 ± 6.8 | 62.2 ± 10.3 | 110.1 ± 16.2 |
| kcat / KmATP(min−1 μM−1) | 0.6 | 1.7 | 2.9 | 3.1 |
| ATP [μM] | 7.5 | 15 | 30 | 60 |
|---|---|---|---|---|
| VmaxCasein(pmol min−1μg−1) | 3.9 ± 0.4 a | 5.6 ± 0.7 a | 10.5 ± 1.4 a | 15.0 ± 1.4 a |
| KmCasein[μM] | 4.6 ± 0.6 | 3.9 ± 0.9 | 4.8 ± 0.8 | 4.0 ± 1.1 |
| kcat(min−1) | 11.3 ± 1.3 | 16.0 ± 1.9 | 30.1 ± 4.2 | 43.2 ± 3.9 |
| kcat / KmCasein(min−1 μM−1) | 2.5 | 4.1 | 6.3 | 10.8 |
| Enzyme | Substrate | Vmax (pmol min−1 μg−1) | Km [μM] |
|---|---|---|---|
| CK1ε | DDX3X | 0.8 ± 0.1 a | 0.2 ± 0.1 a |
| CK1ε | ATP | 1.7 ± 0.1 | 35.4 ± 8.5 |
| DDX3X [μM] | 0.12 | 0.24 | 0.48 | 0.96 |
|---|---|---|---|---|
| VmaxATP(pmol min−1μg−1) | 0.9 ± 0.1 a | 1.3 ± 0.2 a | 1.9 ± 0.4 a | 2.1 ± 0.2 a |
| KmATP[μM] | 26.7 ± 3.3 | 31.6 ± 12.5 | 34.9 ± 7.8 | 16.2 ± 6.0 |
| kcat(min−1) | 2.5 ± 0.3 | 3.7 ± 0.7 | 5.5 ± 1.0 | 6.0 ± 0.7 |
| kcat / KmATP(min−1 μM−1) | 0.09 | 0.11 | 0.16 | 0.37 |
| ATP [μM] | 15 | 30 | 60 | 120 |
|---|---|---|---|---|
| VmaxDDX3X(pmol min−1μg−1) | 1.2 ± 0.4 a | 1.5 ± 0.4 a | 2.9 ± 0.7 a | 4.6 ± 0.8 a |
| KmDDX3X[μM] | 0.6 ± 0.3 | 0.5 ± 0.2 | 0.7 ± 0.3 | 0.7 ± 0.2 |
| kcat(min−1) | 3.3 ± 1.2 | 4.2 ± 1.0 | 8.3 ± 2.0 | 13.2 ± 2.3 |
| kcat / KmDDX3X(min−1 μM−1) | 5.5 | 8.4 | 11.9 | 18.9 |
| Dvl2 [μM] | 0.03 | 0.06 | 0.12 | 0.18 |
|---|---|---|---|---|
| VmaxATP(pmol min−1 μg−1) | 4.7 ± 0.8 a | 5.9 ± 1.0 a | 11.5 ± 1.5a | 17.7 ± 1.8a |
| KmATP[μM] | 13.8 ± 3.8 | 7.8 ± 3.5 | 9.5 ± 3.2 | 6.7 ± 1.9 |
| kcat(min−1) | 13.6 ± 2.3 | 17.0 ± 2.9 | 33.1 ± 4.4 | 51.0 ± 5.1 |
| kcat / KmATP(min−1 μM−1) | 1.0 | 2.2 | 3.5 | 7.6 |
| ATP [μM] | 7.5 | 30 | 60 | 120 |
|---|---|---|---|---|
| VmaxDvl2(pmol min−1 μg−1) | 13.3 ± 2.8a | 36.7 ± 23.1a | 49.8 ± 27.3a | 36.3 ± 4.4a |
| KmDvl2[μM] | 0.5 ± 0.1 | 1.1 ± 0.6 | 0.9 ± 0.5 | 0.4 ± 0.1 |
| kcat(min−1) | 38.2 ± 8.0 | 105.7 ± 64.3 | 143.4 ± 68.0 | 104.7 ± 12.6 |
| kcat / KmDvl2(min−1 μM−1) | 76.4 | 98.8 | 159.3 | 261.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bono, B.; Franco, G.; Riva, V.; Garbelli, A.; Maga, G. Novel Insights into the Biochemical Mechanism of CK1ε and its Functional Interplay with DDX3X. Int. J. Mol. Sci. 2020, 21, 6449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176449
Bono B, Franco G, Riva V, Garbelli A, Maga G. Novel Insights into the Biochemical Mechanism of CK1ε and its Functional Interplay with DDX3X. International Journal of Molecular Sciences. 2020; 21(17):6449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176449
Chicago/Turabian StyleBono, Bartolo, Giulia Franco, Valentina Riva, Anna Garbelli, and Giovanni Maga. 2020. "Novel Insights into the Biochemical Mechanism of CK1ε and its Functional Interplay with DDX3X" International Journal of Molecular Sciences 21, no. 17: 6449. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176449