Exercise Training and Vitamin C Supplementation Affects Ferritin mRNA in Leukocytes without Affecting Prooxidative/Antioxidative Balance in Elderly Women

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Methods
2.1. Subjects
2.2. Ethics Approval and Consent to Participate
2.3. Diet Analysis
2.4. Training
2.5. Supplementation
2.6. Report of Determination of VO2 Max
2.7. Blood Collection, Analysis of Vitamin C Concentration, Total Oxidative Status (TOS)/Total Oxidative Capacity (TOC), Total Antioxidative Status (TAS)/Total Antioxidative Capacity (TAC) Analysis and Genetic Research
2.8. Genetic Evaluation
- For TUBB (tubulin beta class I, NM_001293213): forward primer: TCCACGGCCTTGCTCTTGTTT and reverse primer: GACATCAAGGCGCATGTGAAC;
- For FTH1 (NM_002032): forward primer: TCCTACGTTTACCTGTCCATG and reverse primer: CTGCAGCTTCATCAGTTTCTC;
- For FTL (NM_000146): forward primer: GTCAATTTGTACCTGCAGGCC and reverse primer: CTCGGCCAATTCGCGGAA;
- For PCBP1 (NM_006196): forward primer: AGAGTCATGACCATTCCGTAC and reverse primer: TCCTTGAATCGAGTAGGCATC;
- For PCBP2 (NM_001128913): forward primer: TCCAGCTCTCCGGTCATCTTT and reverse primer: ACTGAATCCGGTGTTGCCATG;
- For CAT (NM_001752): forward primer: GATGGACATCGCCACATGAAT and reverse primer: AAGATCCCGGATGCCATAGTC;
- For FOXO3A: forward primer: TTCAAGGATAAGGGCGACAGC and reverse primer: CCCATCAGGGTTGATGATCCA.
2.9. Vitamin C Plasma Concentration Measurement
2.10. TOS/TOC and TAS/TAC Analysis
2.11. Statistical Analysis
3. Results
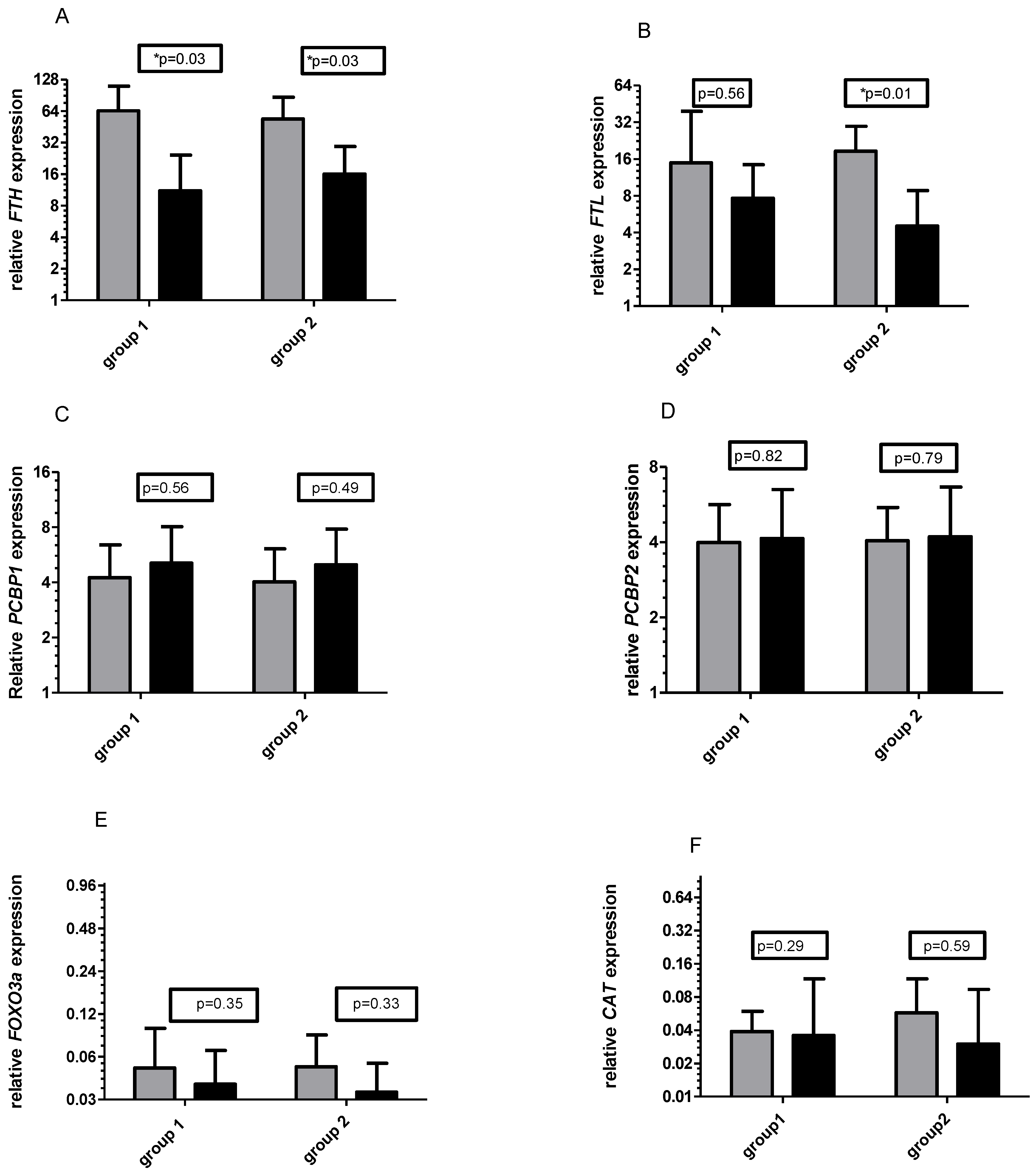
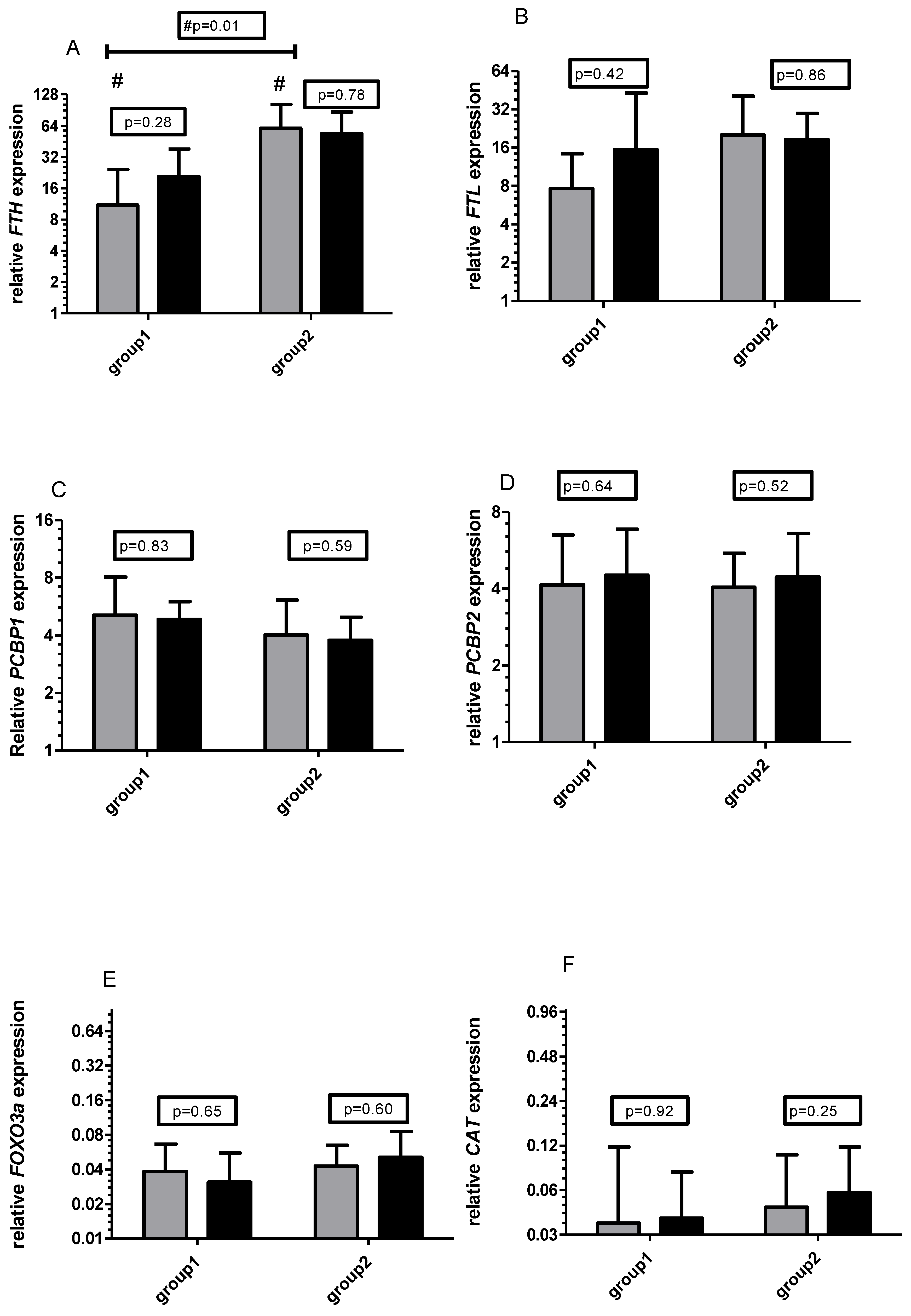
3.1. Gene Expression
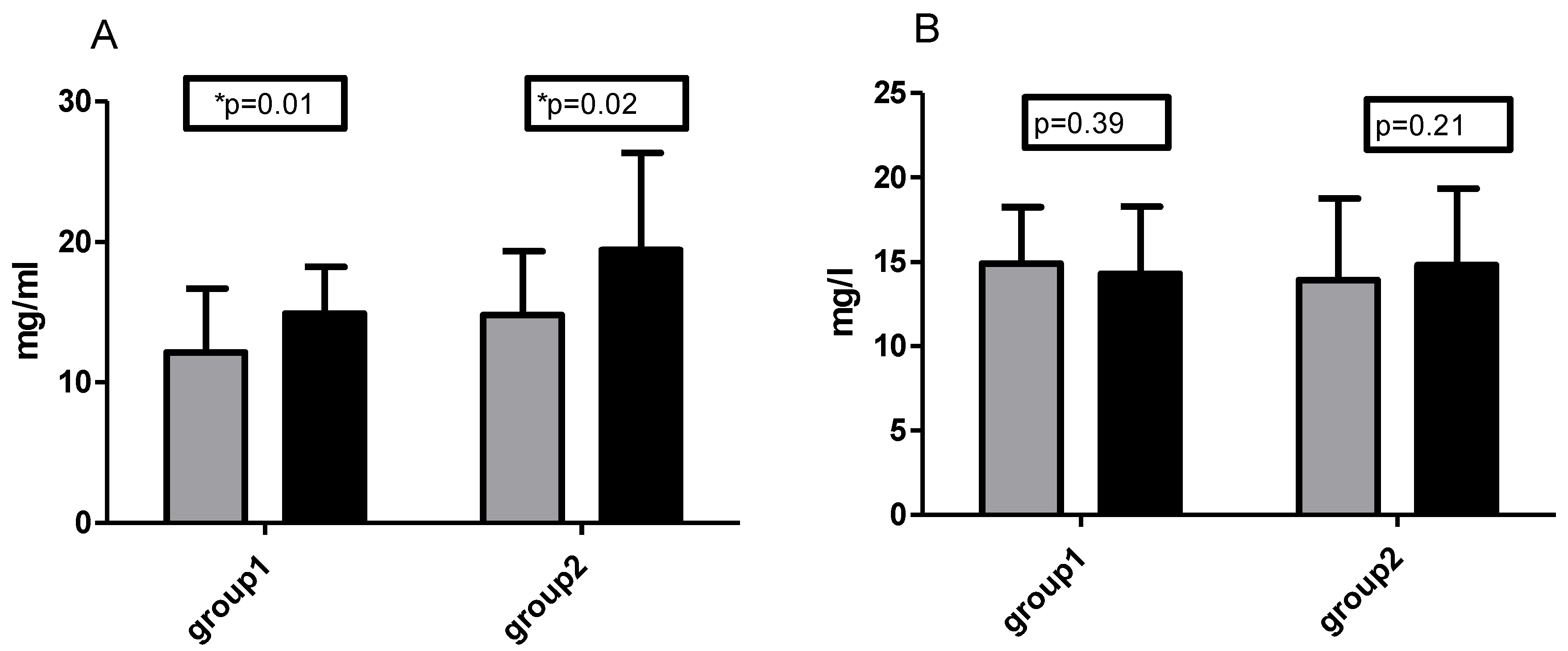
3.2. Control of Diet and Vitamin C Concentration
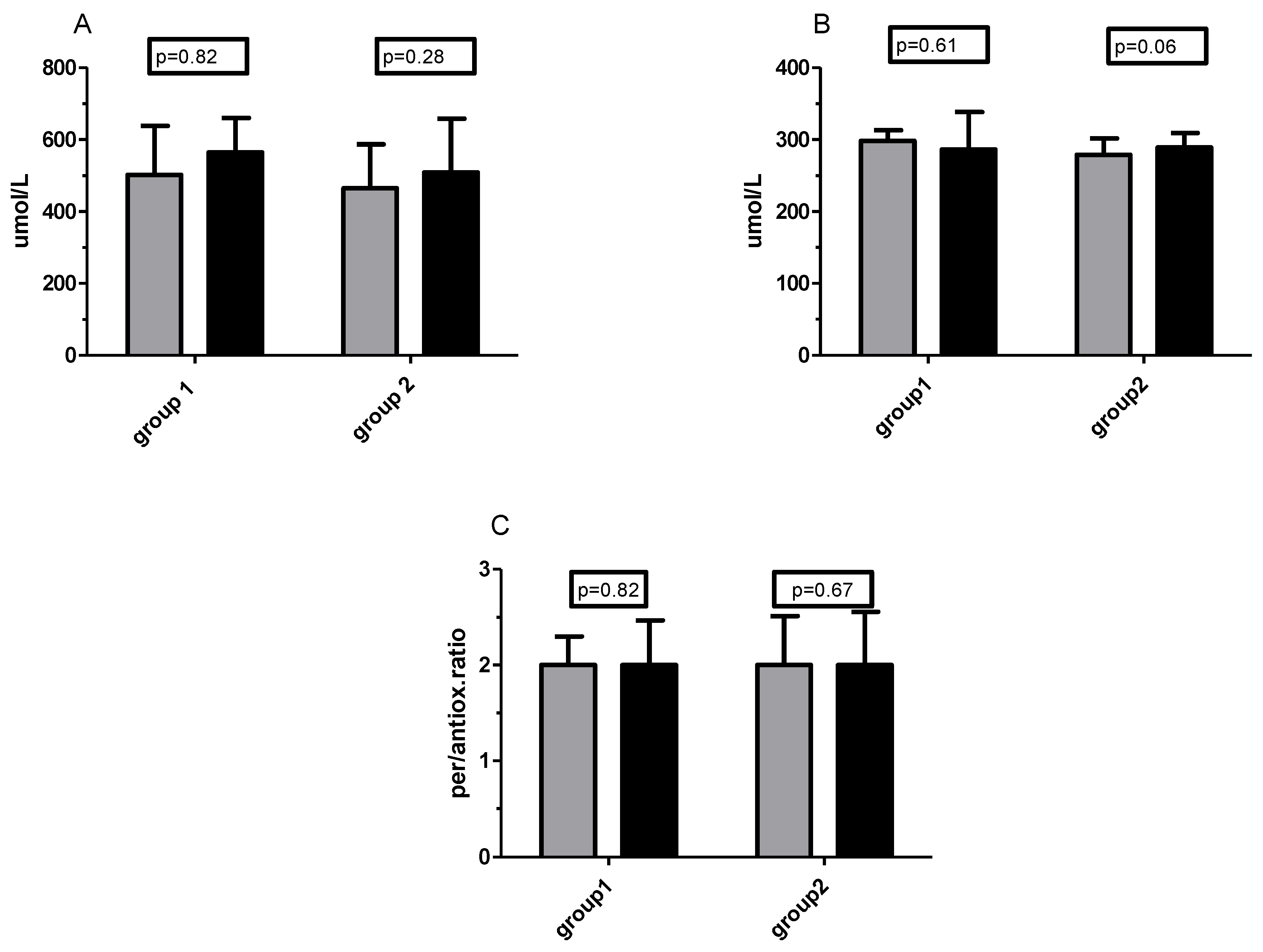
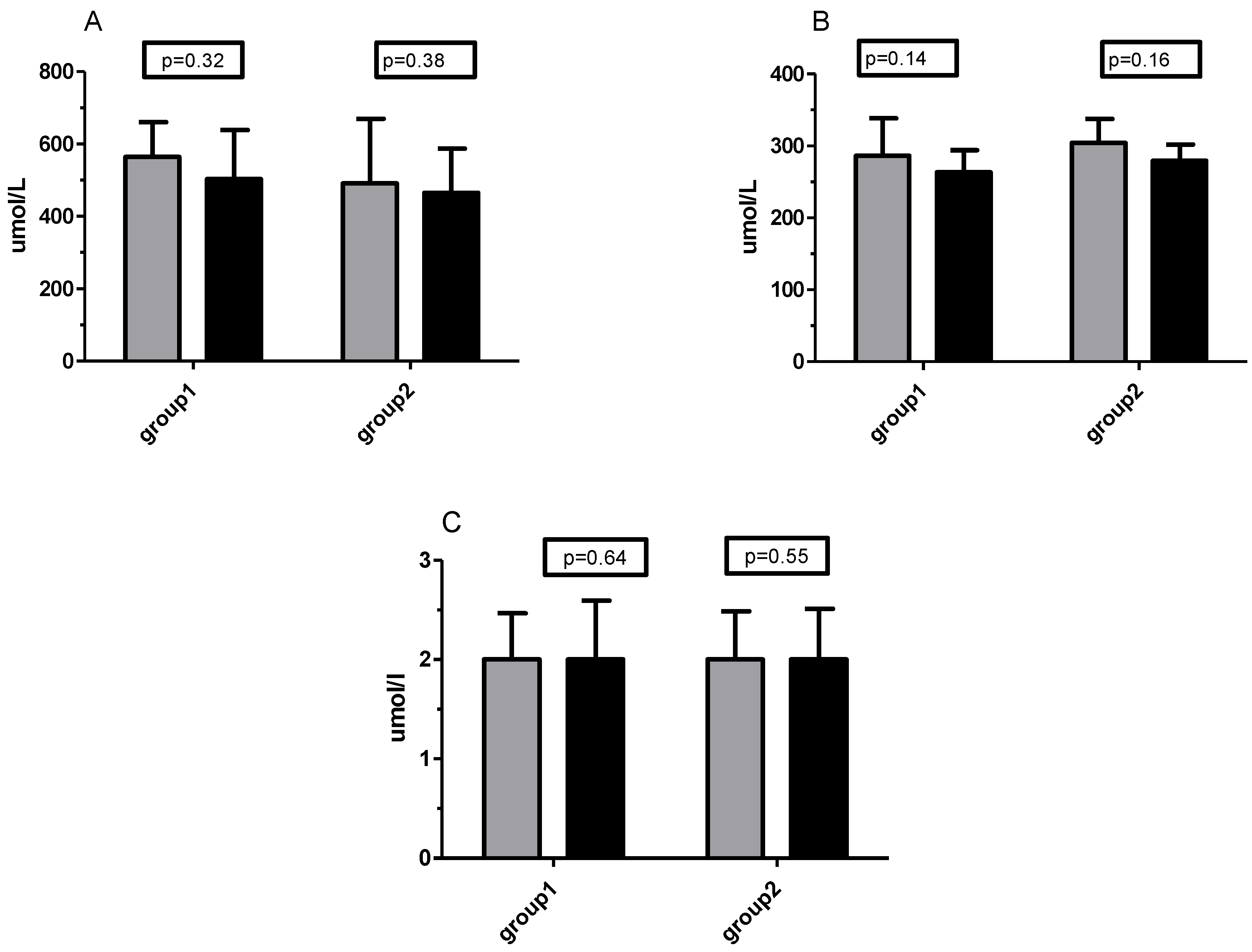
3.3. TOS/TOC and TAS/TAC Analysis
4. Discussion
Vitamin C Concentration and Plasma Pro-/Antioxidant Capacity
5. Conclusions
Study Limitations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kell, D.B.; Pretorius, E. Serum Ferritin Is an Important Inflammatory Disease Marker, as It Is Mainly a leakage Product from Damaged Cells. Metallomics 2014, 6, 748–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, Y.; Ayaki, H.; Whitman, S.P.; Morrow, C.S.; Torti, S.V.; Torti, F.M. Coordinate transcriptional and translational regulation of ferritin in response to oxidative stress. Mol. Cell Biol. 2000, 20, 5818–5827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairo, G.; Recalcati, S. Iron-regulatory proteins: Molecular biology and pathophysiological implications. Expert Rev. Mol. Med. 2007, 9, 1–13. [Google Scholar] [CrossRef]
- Cairo, G.; Castrusini, E.; Minotti, G.; Bernelli-Zazzera, A. Superoxide and hydrogen peroxide-dependent inhibition of iron regulatory protein activity: A protective stratagem against oxidative injury. FASEB J. 1996, 10, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Mieszkowski, J.; Kochanowicz, M.; Żychowska, M.; Kochanowicz, A.; Grzybkowska, A.; Anczykowska, K.; Sawicki, P.; Borkowska, A.; Niespodzińsk, I.B.; Antosiewicz, J. Ferritin Genes Overexpression in PBMC and a Rise in Exercise Performance as an Adaptive Response to Ischaemic Preconditioning in Young Men. BioMed Res. Int. 2019, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Grzybkowska, A.; Anczykowska, K.; Ratkowski, W.; Aschenbrenner, P.; Antosiewicz, J.; Bonisławska, I.; Żychowska, M. Changes in Serum Iron and Leukocyte mRNA Levels of Genes Involved in Iron Metabolism in Amateur Marathon Runners—Effect of the Running Pace. Genes 2019, 10, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Bencze, K.Z.; Stemmler, T.L.; Philpott, C.C. A Cytosolic Iron Chaperone That Delivers Iron to Ferritin. Science 2008, 320, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Maqueda, M.; Roca, E.; Brotons, D.; Soria, J.M.; Perera, A. Affected Pathways and Transcriptional Regulators in Gene Expression Response to an Ultra-Marathon Trail: Global and Independent Activity Approaches. PLoS ONE 2017, 12, e0180322. [Google Scholar] [CrossRef] [Green Version]
- Neufer, P.D.; Bamman, M.M.; Muoio, D.M.; Bouchard, C.; Cooper, D.M.; Goodpaster, B.H.; Booth, F.W.; Kohrt, W.M.; Gerzsten, R.E.; Mattson, M.P.; et al. Understanding the Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Balsano, C.; Alisi, A. Antioxidant Effects of Natural Bioactive Compounds. Curr. Pharm. Des. 2009, 15, 3063–3073. [Google Scholar] [CrossRef]
- Lee, C.H.; Chan, R.S.M.; Wan, H.Y.L.; Woo, Y.C.; Cheung, C.Y.Y.; Fong, C.H.Y.; Cheung, B.M.Y.; Lam, T.-H.; Janus, E.; Woo, J.; et al. Dietary Intake of Anti-Oxidant Vitamins A, C, and E is Inversely Associated with Adverse Cardiovascular Outcomes in Chinese-A 22-Years Population-Based Prospective Study. Nutrients 2018, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.C.; Sung, Y.C.; Davison, G.; Chen, C.Y.; Liao, Y.H. Short-Term High-Dose Vitamin C and E Supplementation Attenuates Muscle Damage and Inflammatory Responses to Repeated Taekwondo Competitions: A Randomized Placebo-Controlled Trial. Int. J. Med. Sci. 2018, 30, 1217–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, G.; Cumming, K.T.; Holden, G.; Hallén, J.; Rønnestad, B.R.; Sveen, O.; Skaug, A.; Paur, I.; Bastani, N.E.; Østgaard, H.N.; et al. Vitamin C and E supplementation hampers cellular adaptation to endurance training in humans: A double-blind, randomised, controlled trial. J. Physiol. 2014, 15, 1887–1901. [Google Scholar] [CrossRef] [PubMed]
- Yfanti, C.; Soren, A.T.; Anders, N.R.; Remi, M.; Ole, M.H.; Jens, L.; Rose, A.J.; Fischer, C.P.; Pedersen, B.K. Antioxidant Supplementation Does Not Alter Endurance Training Adaptation. Med. Sci. Sports Exerc. 2010, 42, 1388–1395. [Google Scholar] [CrossRef]
- Morrison, D.; Hughes, J.; Della Gatta, P.A.; Mason, S.; Lamon, S.; Russell, A.P.; Wadley, G.D. Vitamin C and E supplementation prevents some of the cellular adaptations to endurance-training in humans. Free Radic. J. Biol. Med. 2015, 89, 852–862. [Google Scholar] [CrossRef]
- Mankowski, R.T.; Anton, S.D.; Buford, T.W.; Leeuwenburgh, C. Dietary Antioxidants as Modifiers of Physiologic Adaptations to Exercise. Med. Sci. Sports Exerc. 2015, 47, 1857–1868. [Google Scholar] [CrossRef] [Green Version]
- Szponar, L.; Wolnicka, K.; Rychlik, E. A Photo Album of Products and Dishes; Institute of Food and Nutrition: Warszawa, Poland, 2008. (In Polish) [Google Scholar]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Robilaile, R.; Hoffer, J. A simple method for plasma total vitamin C analysis suitable for routine clinical laboratory use. Nutr. J. 2016, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Natur Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Jarosz, M. Normy Żywienia dla Populacji Polski; Institute of Food and Nutrition: Warszawa, Poland, 2017. (In Polish) [Google Scholar]
- Imam, M.U.; Zhang, S.; Ma, J.; Wang, H.; Wang, F. Antioxidants Mediate Both Iron Homeostasis and Oxidative Stress. Nutrients 2017, 9, 671. [Google Scholar] [CrossRef]
- Bresgen, N.; Eckl, P.M. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 11, 808–847. [Google Scholar] [CrossRef] [PubMed]
- Emerit, J.; Beaumont, C.; Trivin, F. Iron metabolism, free radicals, and oxidative injury. Biomed. Pharmacother. 2001, 55, 333–339. [Google Scholar] [CrossRef]
- Kohgo, Y.; Ikuta, K.; Ohtake, T.; Torimoto, T.; Kato, J. Body iron metabolism and pathophysiology of iron overload. Int. J. Hematol. 2008, 88, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortas, J.; Kuchta, A.; Prusik, K.; Prusik, K.; Ziemann, E.; Labudda, S.; Ćwiklińska, A.; Wieczorek, E.; Jankowski, M.; Antosiewicz, J. Nordic walking training attenuation of oxidative stress in association with a drop in body iron stores in elderly women. Biogerontology 2017, 18, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortas, J.; Prusik, K.; Flis, D.; Prusik, K.; Ziemann, E.; Leaver, N.; Antosiewicz, J. Effect of Nordic Walking training on iron metabolism in elderly women. Clin. Interv. Aging 2015, 27, 1889–1896. [Google Scholar]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Grennberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Donniacuo, M.; Urbanek, K.; Nebbioso, A.; Sodano, L.; Gallo, L.; Altucci, L.; Rinaldi, B. Cardioprotective effect of a moderate and prolonged exercise training involves sirtuin pathway. Life Sci. 2019, 222, 140–147. [Google Scholar] [CrossRef]
- Halon-Golabek, M.; Borkowska, A.; Herman-Antosiewicz, A.; Antosiewicz, J. Iron Metabolism of the Skeletal Muscle and Neurodegeneration. Front. Neurosci. 2019, 15, 165. [Google Scholar] [CrossRef] [Green Version]
- Halon-Golabek, M.; Borkowska, A.; Kaczor, J.J.; Ziółowski, W.; Flis, D.J.; Knap, N.; Kasperuk, K.; Antosiewicz, J. hmSOD1 Gene Mutation-Induced Disturbance in Iron Metabolism Is Mediated by Impairment of Akt Signalling Pathway. J. Cachexia Sarcopenia Muscle 2018, 9, 557–569. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.F.; Pullar, J.M.; Wilson, R.; Spittlehouse, J.K.; Vissers, M.C.M.; Skidmore, P.M.L.; Willis, J.; Cameron, V.A.; Carr, A.C. Vitamin C Status Correlates with Markers of Metabolic and Cognitive Health in 50-Year-Olds: CHALICE Cohort Study. Nutrients 2017, 9, 831. [Google Scholar] [CrossRef]
- Bunpo, P.; Anthony, T.G. Ascorbic acid supplementation does not alter oxidative stress markers in healthy volunteers engaged in a supervised exercise program. Appl. Physiol. Nutr. Metab. 2016, 41, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadtman, E.R. Ascorbic acid and oxidative inactivation of proteins. Am. J. Clin. Nutr. 1991, 54, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Espey, M.G.; Krishna, M.C.; Mitchell, J.B.; Corpe, C.P.; Buettner, G.R.; Shactor, E.; Levine, M. Pharmacologic ascorbic acid concentrations selectively kill cancer cells: Action as a pro-drug to deliver hydrogen peroxide to tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 13604–13609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Chen, Q.; Espey, M.G.; Sun, A.Y.; Lee, J.H.; Krishna, M.C.; Shactor, E.; Choyke, P.L.; Pooput, C.; Kirk, K.L.; et al. Ascorbate in pharmacologic concentrations selectively generates ascorbate radical and hydrogen peroxide in extracellular fluid in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 8749–8754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuma-Kaneko, M.; Sawanobori, M.; Kawakami, S.; Uno, T.; Nakamura, Y.; Onizuka, M.; Ando, K.; Kawada, H. Iron removal enhances vitamin C-induced apoptosis and growth inhibition of K-562 leukemic cells. Sci. Rep. 2018, 26, 17377. [Google Scholar] [CrossRef]
- Gaman, A.M.; Buga, A.M.; Gaman, M.A.; Popa-Wagner, A. The Role of Oxidative Stress and the Effects of Antioxidants on the Incidence of Infectious Complications of Chronic Lymphocytic Leukemia. Oxidative Med. Cell. Longev. 2014. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cabrera, M.C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Viña, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 2008, 87, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Meier, P.; Renga, M.; Hoppeler, H.; Baum, O. The impact of antioxidant supplements and endurance exercise on genes of the carbohydrate and lipid metabolism in skeletal muscle of mice. Cell Biochem. Funct. 2013, 31, 51–59. [Google Scholar] [CrossRef]
- Tofolean, I.T.; Ganea, C.; Ionescu, D.; Filippi, A.; Garaiman, A.; Goicea, A.; Gaman, M.A.; Dimancea, A.; Baran, I. Cellular determinants involving mitochondrial dysfunction, oxidative stress and apoptosis correlate with the synergic cytotoxicity of epigallocatechin-3-gallate and menadione in human leukemia Jurkat T cells. Pharmacol. Res. 2016, 103, 300–317. [Google Scholar] [CrossRef]
- Gheorghe, G.; Stoian, A.P.; Gaman, M.A.; Socea, B.; Neagu, T.P.; Stanescu, A.M.A.; Bratu, O.B.; Mischianu, D.L.D.; Suceveanu, A.J.; Diaconu, C.C. The Benefits and Risks of Antioxidant Treatment in Liver Diseases. Rev. Chim. 2019, 70, 651–655. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, X.; Gaman, M.A.; Kord-Varkaneh, H.; Rahmani, J.; Salehi-Sahlabadi, A.; Day, A.S.; Xu, Y. Serum Vitamin D Levels and Risk of Liver Cancer: A Systematic Review and Dose-Response Meta-Analysis of Cohort Studies. J. Nutr. Cancer 2020. [Google Scholar] [CrossRef]
- Redman, L.M.; Smith, S.R.; Burton, J.H.; Martin, C.K.; Il’yasova, D.; Ravussin, E. Metabolic Slowing and Reduced Oxidative Damage with Sustained Caloric Restriction Support the Rate of Living and Oxidative Damage Theories of Aging. Cell Metab. 2018, 27, 805–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Wegman, M.P.; Guo, M.H.; Bennion, D.M.; Shankar, M.N.; Chrzanowski, S.M.; Goldberg, L.A.; Xu, J.; Williams, T.A.; Lu, X.; Hsu, S.I.; et al. Practicality of intermittent fasting in humans and its effect on oxidative stress and genes related to aging and metabolism. Rejuvenation Res. 2015, 18, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Oxidative stress and vascular inflammation in aging. Free Radic. Biol. Med. 2013, 65, 380–401. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | I (Baseline) | ||
|---|---|---|---|
| Group 1 | Group 2 | p | |
| VO2 max. (mL/kg/min) | 20.69 ± 4.05 | 19.73 ± 2.45 | 0.61 |
| Body mass (kg) | 74.26 ± 16.87 | 71.46 ± 5.39 | 0.68 |
| Fat (kg) | 29.86 ± 13.86 | 29.74 ± 3.12 | 0.98 |
| Muscle mass (kg) | 24.07 ± 3.76 | 22.46 ± 2.17 | 0.34 |
| Parameter | II | p (I vs. II) | III | p (I vs. III) | p (II vs. III) | |
|---|---|---|---|---|---|---|
| VO2 max. (mL/kg/min) | Group 1 | 20.96 ± 4.47 | 0.67 | 21.63 ± 4.66 | 0.13 | 0.49 |
| Group 2 | 19.62 ± 2.04 | 0.79 | 20.62 ± 1.48 | 0.30 | 0.27 | |
| p (Group 1 vs. Group 2) = 0.51 | p (Group 1 vs. Group 2) = 0.62 | |||||
| Body mass (kg) | Group 1 | 73.33 ± 16.15 | 0.55 | 71.79 ± 16.24 | 0.13 | 0.07 |
| Group 2 | 70.07 ± 7.12 | 0.38 | 65.50 ± 6.95 *,# | 0.03 | 0.04 | |
| p (Group 1 vs. Group 2) = 0.63 | p (Group 1 vs. Group 2) = 0.36 | |||||
| Fat (kg) | Group 1 | 26.87± 10.67 | 0.14 | 24.98 ± 14.37 * | 0.02 | 0.45 |
| Group 2 | 26.60 ± 6.18 | 0.12 | 23.07 ± 7.68 * | 0.02 | 0.13 | |
| p (Group 1 vs. Group 2) = 0.95 | p (Group 1 vs. Group2) = 0.75 | |||||
| Muscle mass (kg) | Group 1 | 25.58 ± 4.79 | 0.55 | 25.89 ± 4.75 * | 0.04 | 0.85 |
| Group 2 | 23.57 ± 2.24 | 0.07 | 23.07 ± 1.91 | 0.45 | 0.45 | |
| p (Group 1 vs. Group 2) = 0.60 | p (Group 1 vs. Group 2) = 0.15 | |||||
| Diet Analysis | Group 1 | Group 2 |
|---|---|---|
| Energy (kcal) | 2069 ± 702 | 1634 ± 245 |
| Proteins (g/day) | 90 ± 32 | 71 ± 20 |
| Fat (g/day) | 75 ± 29 | 62 ± 12 |
| Carbohydrates (g/day) | 260 ± 130 | 196 ± 22 |
| Vit C (mg) | 154 ± 105 | 141 ± 56 |
| Vi C (mg/kg b. m.) | 2.13 ± 1.5 | 2.00 ± 0.79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żychowska, M.; Grzybkowska, A.; Wiech, M.; Urbański, R.; Pilch, W.; Piotrowska, A.; Czerwińska-Ledwig, O.; Antosiewicz, J. Exercise Training and Vitamin C Supplementation Affects Ferritin mRNA in Leukocytes without Affecting Prooxidative/Antioxidative Balance in Elderly Women. Int. J. Mol. Sci. 2020, 21, 6469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186469
Żychowska M, Grzybkowska A, Wiech M, Urbański R, Pilch W, Piotrowska A, Czerwińska-Ledwig O, Antosiewicz J. Exercise Training and Vitamin C Supplementation Affects Ferritin mRNA in Leukocytes without Affecting Prooxidative/Antioxidative Balance in Elderly Women. International Journal of Molecular Sciences. 2020; 21(18):6469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186469
Chicago/Turabian StyleŻychowska, Małgorzata, Agata Grzybkowska, Monika Wiech, Robert Urbański, Wanda Pilch, Anna Piotrowska, Olga Czerwińska-Ledwig, and Jędrzej Antosiewicz. 2020. "Exercise Training and Vitamin C Supplementation Affects Ferritin mRNA in Leukocytes without Affecting Prooxidative/Antioxidative Balance in Elderly Women" International Journal of Molecular Sciences 21, no. 18: 6469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186469