1. Introduction
Interleukin-1 (IL-1) family cytokines play important roles in regulating and initiating inflammatory and immunological responses [
1]. The IL-1 family includes eleven cytokines comprising seven agonist ligands, three receptor antagonists, and an anti-inflammatory cytokine [
2]. Interleukin 33 (hereafter called as “IL-33”) cytokine is identified as one of the IL-1 family agonist ligands [
3]. It was first regarded as an alarmin that is released to signal immune system when a cell or tissue is damaged or stressed [
4]. Recently, IL-33 has been considered as an important factor of the immune system involved in allergic inflammation and chronic diseases such as asthma, atopic dermatitis, and allergic rhinitis [
5,
6,
7].
IL-33, expressed in endothelial cells, fibroblasts, epithelial cells, and other cells, binds to its receptor suppressor of tumorigenicity 2 (ST2)/interleukin 1 receptor-like 1 (IL1RL1), which formed heterodimer with co-receptor, IL-1 receptor accessory protein (IL1RAcP) [
4,
8]. There are two types of ST2 isoforms: the transmembrane form, ST2, and soluble form, sST2, covering residues 19 through 321 of the ectodomain (hereafter, we call all isoforms as simply “ST2”). The ST2 is expressed on various immune cells including innate lymphoid group 2 cells (ILC2s), mast cells, dendritic cells, macrophages, basophils, and type 2 helper T cells (T
h2), and it is linked to T
h2 effector functions [
9,
10]. IL-33 exerts its biological functions followed by binding to ST2 expressed in immune cells, and it is mainly associated with T
h2 responses through the production of inflammatory cytokines IL-5 and IL-13 [
3,
11]. The heterodimer complex formation activates downstream signaling complex formation. Myeloid differentiation primary response 88 (MyD88) first binds to heterodimeric receptor and leads to the recruitment of interleukin-1 receptor-associated kinase 1 (IRAK1), IRAK4 and tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6), and these subsequently activate mitogen-activated protein kinases (MAPKs) and nuclear factor κB (NF-κB) signaling pathways to promote inflammatory cytokine production [
9,
12,
13]. It seems that IL-33 has the potential to activate T
h2 cytokine-mediated allergic inflammation and related diseases, suggesting that blockade of the IL-33/ST2 signaling axis can be a new therapeutic strategy for allergic inflammation and chronic inflammatory diseases [
14,
15,
16].
Several strategies have been developed to suppress the IL-33 mediated downstream signaling pathway to prevent chronic diseases and allergic inflammation: antagonists against IL-33, and antagonists against ST2 or sST2 binding to IL-33 [
17]. Here, we describe the discovery and characterization of single-chain variable fragment (scFv) monoclonal antibodies (mAbs) directly targeting IL-33 to inhibit IL-33 binding to ST2. Although there are diverse antibodies against IL-33 for various purposes, they are mainly derived from immunizing living organisms with immunoglobulin G forms [
16,
18] or monoclonal antibodies for IL-33 detection [
15,
19]. Since immunoglobulin G (IgG) forms of antibodies are hard to handle and not suitable to further engineering, we used a human synthetic single-chain variable fragment (scFv) antibody library to screen IL-33 specific mAbs in vitro.
The discovery of mAbs using phage display library was performed with five rounds of biopanning, and enzyme-linked immunosorbent assay (ELISA) was used to determine the antibodies affinity. Using immunoblotting, we observed their cross-reactivity, and two types of mutant-based epitope mapping were implemented to identify the binding epitope domains. The inhibition effect of antibody was verified by glutathione S-transferase (GST) pull-down assay and human cell-expressing ST2 and IL1RAcP-based assay. The antibody seems to have therapeutic function by interfering with IL-33 binding to the ST2 receptor, heterodimeric receptor complex formation, and blocking the IL-33/ST2 signaling axis.
4. Materials and Methods
4.1. Plasmid Constructs Cloning
Genes encoding the mature form of IL-33 (residues 112–270), hereafter called simply “IL-33”, and the ectodomain of ST2 (residues 19–321) were synthesized (Cosmo Genetech, Seoul, Korea) and cloned into BamHI/StuI sites of parallel GST-2 vector [
26] and
NotI/
NcoI sites of the pSF vector, respectively. CH2 and CH3 domains of human the IgG Fc region were cloned into XhoI/BsgI sites of pSF-ST2 plasmid for ST2-Fc fusion protein expression. Plasmids encoding IL-33 mutants (deletion mutants, alanine-scanning mutants, and an oxidation-resistant mutant C208S/C232S) were prepared by following the protocol for QuikChange kit (Agilent, Santa Clara, CA, USA). Identities of all the constructs were verified by DNA sequencing.
4.2. Expression and Purification of Recombinant Proteins
The plasmid encoding GST-IL-33 was transformed into E. coli BL21 (DE3) cells. A single colony was inoculated into 10 mL Luria broth (LB) media containing 100 μg·mL−1 ampicillin and grown at 37 °C overnight. After 16–18 h, the pre-cultured cells were transferred to 500 mL LB media containing 100 μg·mL−1 of ampicillin, grown at 37 °C until OD600 0.6–1.0, induced with 0.6 mM isopropyl-β-d-1-thiogalactopyranoside (IPTG), and further grown at 25 °C overnight with gentle shaking. Cells were harvested by centrifugation and re-suspended in lysis buffer (50 mM Tris-HCl pH = 7.5 and 150 mM NaCl). Cells were disrupted by ultrasonication, cleared by centrifugation, and the supernatant containing GST-IL-33 was transferred to Glutathione Sepharose 4B resin (GE Healthcare, Chicago, IL, USA) pre-equilibrated with the lysis buffer. After washing the resin with the lysis buffer, GST-IL-33 was eluted in GST elution buffer (50 mM Tris-HCl pH 8.0, 150 mM NaCl, and 10 mM reduced glutathione). IL-33 was relieved from the GST fusion protein using a recombinant His6-tagged tobacco etch virus (TEV) protease during dialysis at 4 °C overnight. The resulting IL-33 was further purified on a Superdex 75 10/300 GL size-exclusion chromatography column (GE HealthCare, Chicago, IL, USA) pre-equilibrated with the lysis buffer. A ST2-Fc fusion protein, containing the ectodomain (residues 19–321) of ST2 and Fc from human IgG, was expressed in Expi293F cells (Thermo Fisher Scientific, Waltham, MA, USA) maintained in Expi293 expression medium (Thermo Fisher Scientific, Waltham, MA, USA). The day before transfection, cells were seeded to a final density of 2 × 106 viable cells ml−1 in a 125 mL Erlenmeyer flask and grown at 37 °C for 24 h. After 24 h, cells were transfected with 30 μg of pSF-ST2-Fc plasmid DNA diluted in Opti-MEM™ I Medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 80 μL of ExpiFectamine™ 293 Reagent (Thermo Fisher Scientific, Waltham, MA, USA). Four days post-transfection, the cells were harvested by centrifugation, and its supernatant containing ST2-Fc was transferred to protein A agarose (Thermo Fisher Scientific, Waltham, MA, USA) pre-equilibrated by phosphate-buffered saline (PBS). The resin was washed using PBS, and ST2-Fc was eluted in Fc elution buffer (100 mM glycine pH = 3 with 1/10 volume of 1 M Tris-HCl pH = 8.0). The eluted ST2-Fc was dialyzed in the lysis buffer at 4 °C overnight.
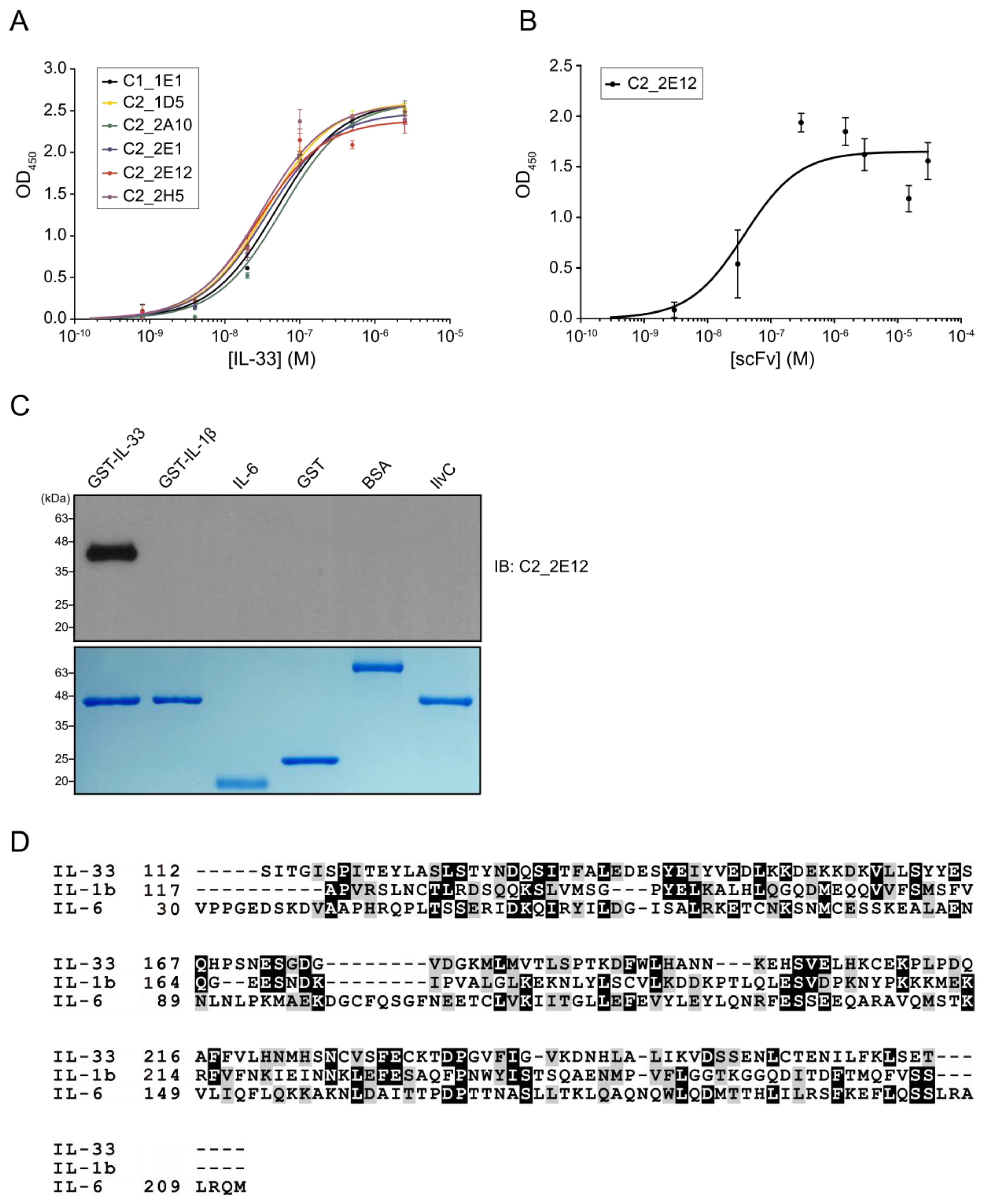
4.3. Biopanning Using Phage Display
A synthetic human scFv library encoding His
6- and HA-tagged scFv clones was used for biopanning [
27]. Biopanning was performed as described previously with some modifications [
28]. Library biopanning was performed in immuno tubes coated with the recombinant GST–IL-33 as an antigen in two conditions: inclusion of a negative selection with GST in every round (condition 1), and in the first round only (condition 2). In condition 1, five rounds of biopanning were performed in immuno tubes coated with GST protein for a negative selection and the recombinant GST-IL-33 at a gentle decrease in antigen concentrations (50, 10, 7.5, 5, and 2.5 μg·mL
−1). In condition 2, the negative selection using GST protein is performed only in the first round of biopanning and coated with GST-IL-33 in the other four rounds of biopanning using the same concentration with condition 1. To select scFv clones that specifically bind to IL-33, single colonies from the final round of a biopanning output plate were grown in a 96-well cell culture plate until OD
600 reached 0.6–1.0 and induced with 1 mM IPTG grown overnight at 30 °C with shaking. The harvested cells in each 96-well were re-suspended in cold 1× TES buffer (50 mM Tris-HCl pH = 8.0, 1 mM ethylenediaminetetraacetic acid (EDTA) and 20% (
w/
v) sucrose) for 30 min on ice, and cold 0.2×TES buffer was added to the re-suspended cells for 1 h on ice. The recombinant GST-IL-33 protein at 10 μg·mL
−1 in PBS was coated on a 96-well ELISA plate. ELISA assay for scFv screening with horseradish peroxidase (HRP) conjugated anti-HA secondary antibody (1:3000 dilution, Santa Cruz Biotechnology, Dallas, TX, USA) was performed with a final reading of signals recorded at OD
450. The OD
450 values with GST-IL-33 were divided by the OD
450 value with GST, and the ratio of OD
450 values was compared.
4.4. Expression and Purification of scFvs in E. coli
Cells were pre-cultured from the single colonies of scFv-expressing E. coli BL21 (DE3) at 37 °C overnight, transferred to 500 mL super broth (SB) media containing 100 μg·mL−1 ampicillin. Cells were grown at 37 °C until OD600 0.5–0.8 and induced with 1 mM IPTG at 30 °C with vigorous shaking. After 16–18 h, cells were harvested by centrifugation and re-suspended in cold 1× TES buffer for 30 min on ice, and cold 0.2× TES buffer was added to the re-suspended cells for 1 h on ice. The re-suspended cells supplemented with 5 mM MgCl2 to block EDTA were centrifuged, and their supernatants containing each scFvs were transferred to Ni-NTA agarose resin (Qiagen, Hilden, Germany). Each resin was washed by wash buffer A (PBS supplanted with 20 mM imidazole and 0.5 mM DTT) and scFvs were eluted by His-tag elution buffer A (PBS supplemented with 300 mM imidazole and 0.5 mM DTT). Size exclusion chromatography was performed on a Superdex 75 increase 10/300 GL column (GE HealthCare, Chicago, IL, USA) pre-equilibrated with PBS.
4.5. Reformatting, Expression, and Purification of C2_2E12 in Mammalian Cells
The gene encoding C2_2E12 in the pComb3X vector [
27] was cloned into the pSF vector to express three different formats of C2_2E12 in mammalian cells: scFv, antigen-binding fragment (Fab), and immunoglobulin G (IgG). The C2_2E12 scFv was cloned into NotI/XhoI sites of the pSF vector for expression in Expi293F cells (Thermo Fisher Scientific, Waltham, MA, USA) maintained in Expi293 expression medium (Thermo Fisher Scientific, Waltham, MA, USA). A variable heavy chain and variable light chain of C2_2E12 were cloned into NotI/NcoI sites and HindIII/XhoI sites of the pSF vector, respectively, for the expression of Fab and IgG formats in Expi293F cells (Thermo Fisher Scientific, Waltham, MA, USA). Transfection and preparation steps for Fab and IgG were the same with those for scFv except for the amount and ratio of DNA plasmid used for transfection. The expression of scFv, Fab, and IgG formats of C2_2E12 in mammalian cells was performed in the same way as for ST2-Fc. Supernatants containing C2_2E12 scFv and Fab were transferred to Ni-NTA agarose resin (Qiagen, Hilden, Germany) pre-equilibrated with PBS. Each resin was washed by wash buffer B (PBS supplemented with 20 mM imidazole) and eluted by His-tag elution buffer B (PBS supplemented with 300 mM imidazole). Supernatant containing C2_2E12 IgG was transferred to a protein A agarose (Thermo Fisher Scientific, Waltham, MA, USA) pre-equilibrated by PBS. The resin was washed using PBS, and the C2_2E12 IgG was eluted in the Fc elution buffer. The antibodies were dialyzed at 4 °C overnight, concentrated, and loaded to a Superdex 75 10/300 GL size-exclusion chromatography column (GE HealthCare, Chicago, IL, USA) pre-equilibrated with PBS.
4.6. Immunoblot
Purified proteins were subjected to 12% SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck Millipore, Burlington, MA, USA). Membranes were blocked with 5% (w/v) skim milk in Tris-buffered saline (pH = 7.5) containing 0.1% (v/v) tween-20 for 1 h at room temperature. Different primary antibodies were incubated at 4 °C overnight, and secondary antibodies were incubated at room temperature for 1 h. The bound antibody was detected by enhanced chemiluminescence (ECL) reaction with EZ-Western Lumi Pico kit (DoGen, Seoul, Korea).
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
GST-IL-33 was coated on half the total area of a Costar® 96-well plate in the clear flat bottom polystyrene high bind microplate (Corning, Corning, NY, USA) and incubated at 4 °C for overnight. In the next day, the resulting culture was washed using PBST (PBS supplemented with 0.1% (v/v) tween-20) and blocked using blocking buffer (5% (w/v) skim milk in PBST). Serial dilutions of purified scFvs as the primary antibody with approximately 8 ng·mL−1 to 0.8 mg·mL−1 were added to the wells. The plate was incubated at ambient temperature for 1 h and washed using PBST. Subsequently, HRP-conjugated anti-HA antibody as the secondary antibody (1:3000 dilution, Santa Cruz Biotechnology) was added to the wells and incubated at ambient temperature for 1 h. The incubated plate was washed using PBST, and tetramethylbenzidine (TMB) substrate solution (GenDEPOT, Katy, TX, USA) was added for color development. After incubation for 10 min, 1 M H2SO4 was added to the plate to stop the color development reaction. The final signal readings were recorded at 450 nm and plotted using Prism 5 (GraphPad, San Diego, CA, USA).
4.8. Biolayer Interferometry (BLI)
Binding kinetics was measured by BLI experiments using a BLItz system (ForteBio, Fremont, CA, USA).
E. coli cell lysate containing His
6-tagged antibodies was prepared in 0.5× TES buffer. GST-IL-33 was prepared in BLI buffer (PBS supplemented with 20 mM imidazole, 0.05% (
v/
v) Triton X-100 and 0.1 mg·mL
−1 BSA) to reduce the nonspecific binding signal. The BLI buffer was also used as the kinetics buffer. The cell lysate was immobilized to Ni-NTA biosensors (ForteBio, Fremont, CA, USA) and washed using the kinetics buffer. The sensors were subsequently reacted with various concentrations (2, 1, 0.5, and 0.25 μM) of GST-IL33 (association step) and washed using the kinetics buffer (dissociation step). These assays were performed twice each. All real-time recorded sensograms were analyzed by the ‘global fitting’ method in BLItz Pro 1.2 (ForteBio, Fremont, CA, USA) to calculate
kon (association rate constant) and
koff (dissociation rate constant) values. The
Kd (dissociation constant) value of each antibody was calculated using the following equation:
r2 analysis, an indication of goodness of graph curve fitting, was performed using BLItz Pro 1.2, and the
r2 values of all experiments were above 0.98. The graphs of raw sensograms were prepared by Prism 5 (GraphPad, San Diego, CA, USA).
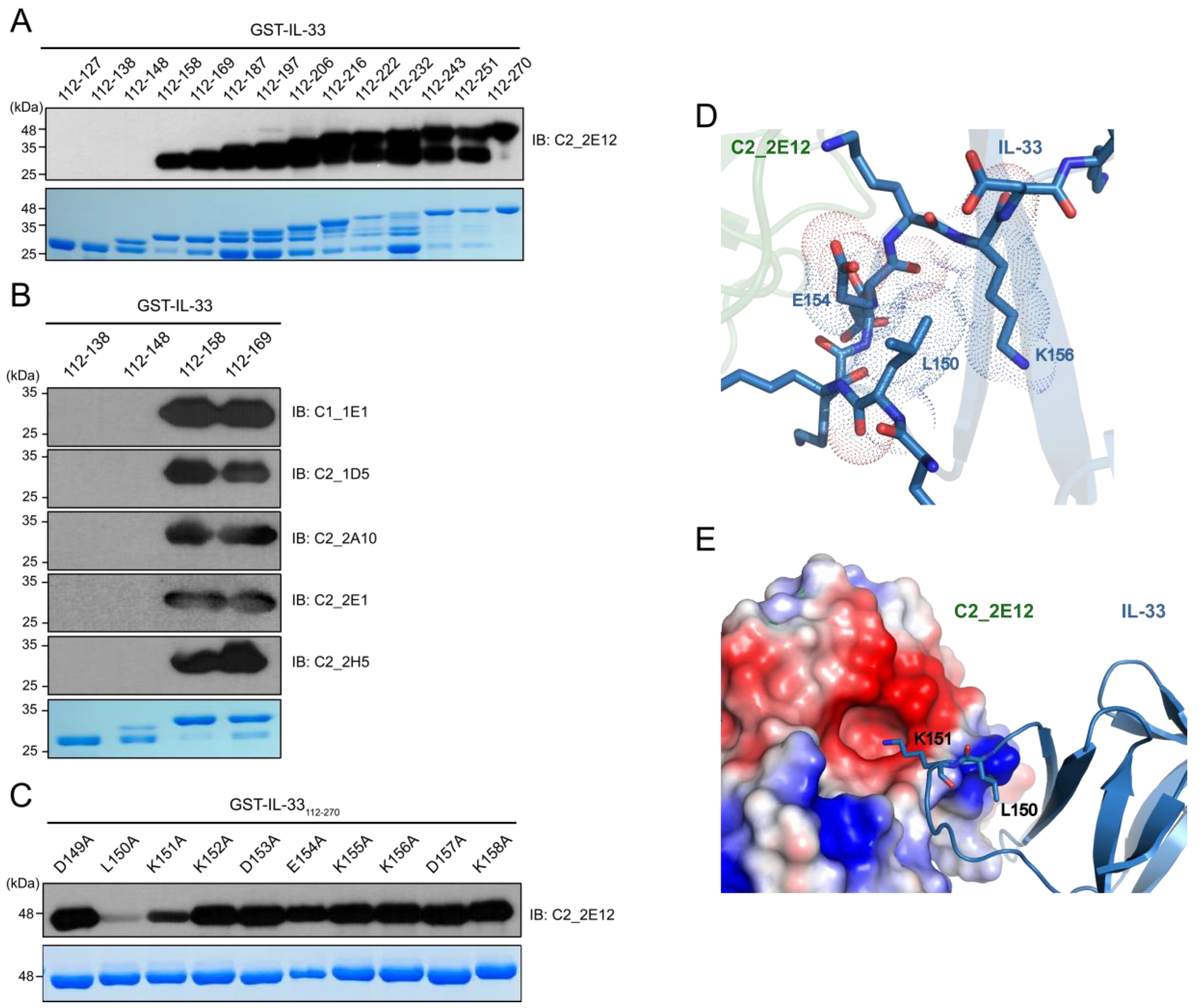
4.9. Structural Modeling
A homology structural model for C2_2E12 that was generated using the SWISS-MODEL server [
24] and the crystal structure of IL-33:ST2 complex (PDB ID: 4KC3) were used as templates for the docking of C2_2E12 to IL-33. Protein–protein docking modeling was performed using the HADDOCK server [
29]. To perform the HADDOCK modeling, a restraint was applied such that the two key epitope residues of IL-33, L150 and K151, must interact with the complementary determination region of C2_2E12. The Z-score of clustering and other modeling parameters are listed in
Table S2. Structural analysis of the interface between C2_2E12 and IL-33 was performed using PyMOL 1.8 (Schrödinger, New York, NY, USA).
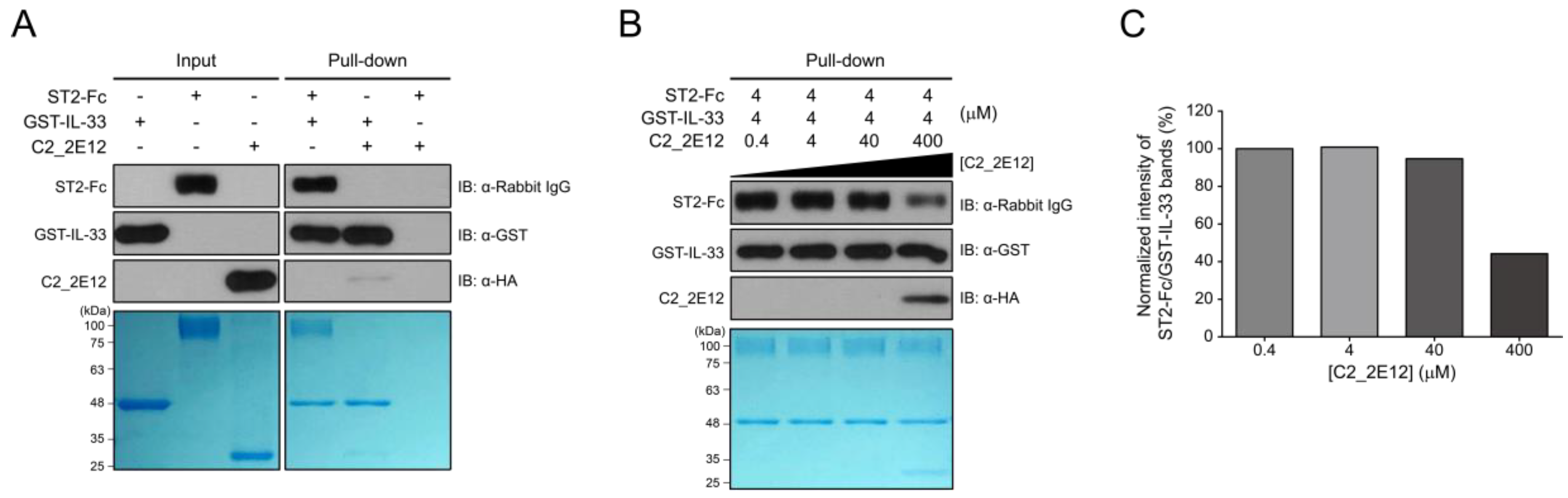
4.10. Pull-Down Assay
Pull-down assay was performed using Glutathione Sepharose 4B resin (GE Healthcare, Chicago, IL, USA). First, 4 µM of GST-IL-33 was added to the resin and incubated for 30 min at 4 °C with gentle shaking. The resin was washed 4 times with wash buffer C (50 mM Tris-HCl pH = 7.5 and 150 mM NaCl) and incubated for 30 min at 4 °C with gentle shaking upon the addition of 4 µM of ST2-Fc. Subsequently, C2_2E12 at a series of concentrations (0.4, 4, 40, and 400 µM) was added to the resulting resin with further incubation for 30 min at 4 °C. After 4 times of washing, proteins were separated by SDS-PAGE on a 12% gel, stained with Coomassie Brilliant Blue, or transferred onto a PVDF membrane (45 mA for 60 min). Protein bands on the PVDF membrane were visualized by immunoblotting by ECL reaction using anti-ST2 (1:5000 dilution, Abcam, Cambridge, United Kingdom) and mouse anti-rabbit IgG–HRP (1:10000 dilution, Santa Cruz Biotechnology, Dallas, TX, USA), anti-GST-HRP (1:10000 dilution, Santa Cruz Biotechnology, Dallas, TX, USA), and anti-His6–HRP (1:10000 dilution, Santa Cruz Biotechnology, Dallas, TX, USA). Gels were quantified using ImageJ (National Institutes of Health, Bethesda, MD, USA).
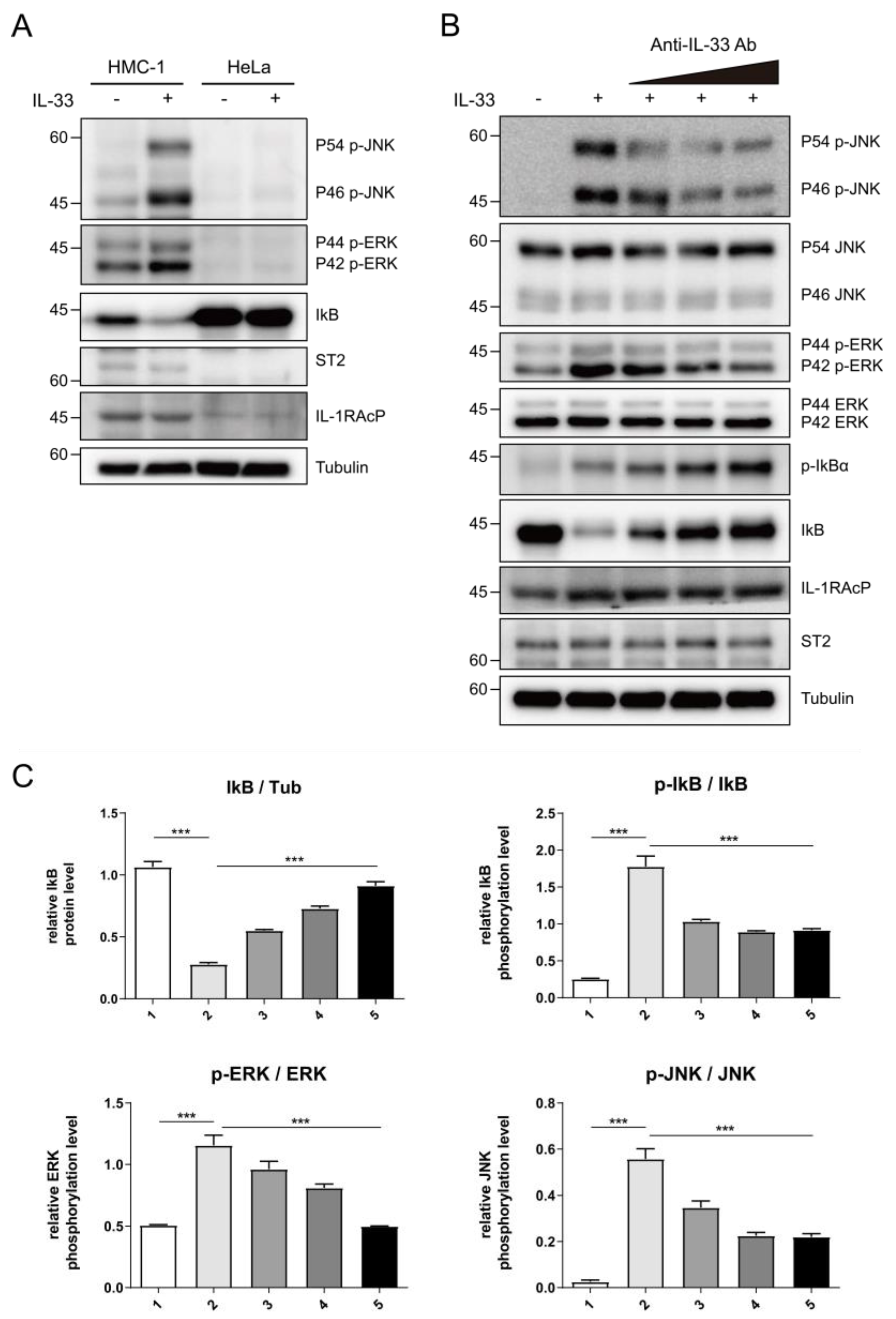
4.11. Cell Signaling Analysis
HMC-1 cells were a kind gift from Prof. Soohyun Kim, Konkuk University, South Korea. HMC-1 cells were cultured in Iscove’s modified Dulbecco’s medium (IMDM) containing 10% FBS. HeLa cells were cultured in Dulbecco′s modified Eagle′s media (DMEM) containing 10% FBS. Cells were centrifuged at 800× g for 5 min at 4°C, and they were lysed with NETN lysis buffer (100 mM NaCl, 20 mM Tris-HCl pH = 8.0, 0.5 mM EDTA, 0.1% NP-40, 5 mM NEM, 10 mM NaF, 1 mM Na3VO4, a protein inhibitor cocktail) for 15 min on ice. Cell lysates were centrifuged at 15,000× g at 4°C for 10 min. Protein extracts were separated by SDS-PAGE and transferred onto PVDF membranes. Membranes were immunoblotted with the indicated antibodies, and the signals were visualized with ImageQuant™ LAS 4000 mini (GE Healthcare, Chicago, IL, USA). Anti-p-IĸB (#2859), anti-IĸB (#4814), anti-p-S6K (#9251), and anti-S6K (#9252) antibodies were purchased from Cell signaling. Anti-p-ERK (sc-7383), anti-ERK (sc-27129), and anti-IL-1RAcP (sc-376872) were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). Anti-ST2 (#D065-3) were purchased from MBL (Woburn, MA, USA).


{kind=link}
{kind=link}
{kind=link}
{kind=link}



