In oat and maize crosses, some maize chromosomes are not extruded during embryogenesis, but are ultimately stabilized and act as oat chromosomes in mitosis [
20]. Studies of Riera-Lizarazu et al. [
21] explained that maize chromosome elimination in crosses between oat and maize is more gradual than, for example, in wheat and maize. A single maize chromosome in an oat genome background represents only about 2% of the total DNA [
22]. Moreover, the OMA plants look like oat with some morphological modifications, and the amount of maize gene expression or gene silencing is difficult to intuitively assess. However, Dong et al. [
10] actually found that more than 70% of the genes from the alien maize chromosomes maintained the original expression or transcription pattern under the oat genomic environment. They speculated that the maize genes maintaining the original transcription in OMA lines (introgression expression) may be predominantly regulated by a local
cis element in the alien maize chromosome [
10]. Gene expression of alien chromosomes has also been detected in wheat × barley addition lines [
23].
3.2. Functioning of the OMA Line Photosynthetic Apparatus in Drought Conditions
Drought is the most severe stress and the main cause of significant losses in growth and productivity of crop plants [
25], inducing meaningful alterations in plant physiology and biochemistry. Various physiological responses of plants to drought are linked with their tolerance mechanisms such as pigment content and stability, as well as high relative water content [
26]. The sensitivity of plants to drought depends on the intensity of stress, species, genotype, the presence of other stress factors, and the plant developmental phase [
19].
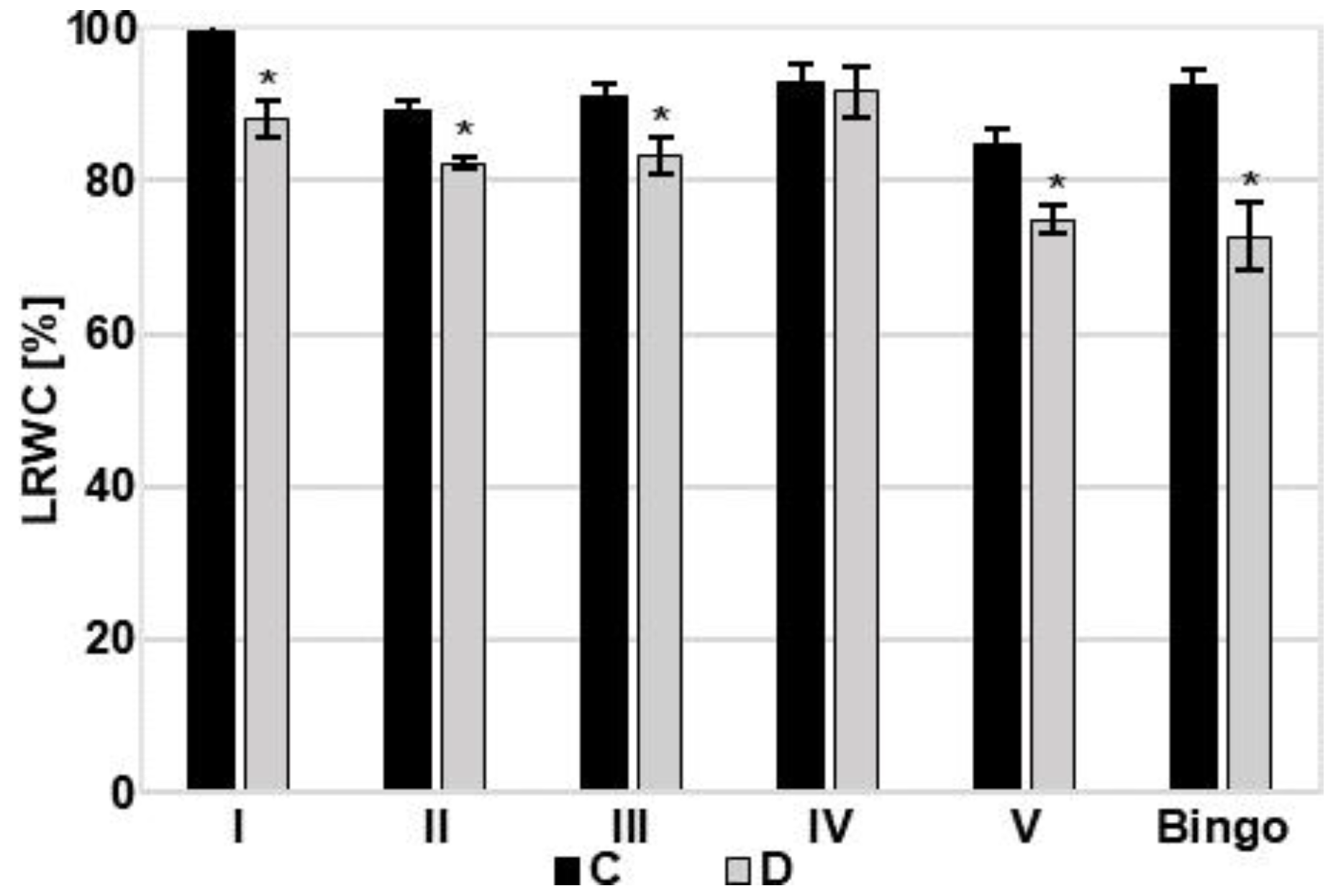
Under drought stress, LRWC is proposed as a more important indicator of water status than other water potential parameters [
27]. A significant decrease in LRWC value in response to drought stress has been reported by many authors [
28,
29,
30,
31]. Depending on the reduction of LRWC, water stress can be divided into mild (LRWC decreases by 8–10%), moderate (10–20%), and severe (over 20%) [
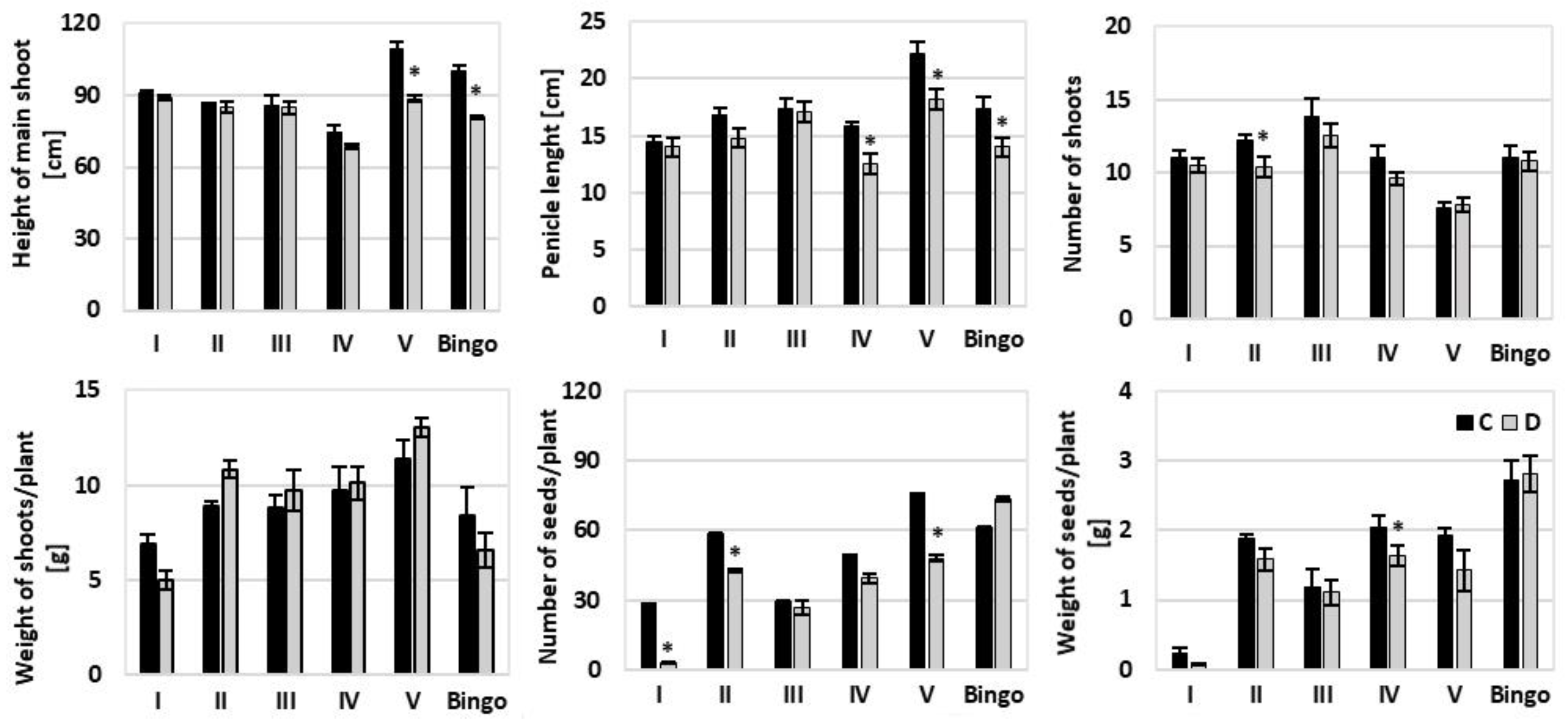
32]. According to this classification, the soil drought applied in our experiment caused severe/moderate stress in cv. Bingo (LRWC decreased by 21%), and mild/moderate in tested OMA lines (LRWC declined by 7–11%). This deviation in LRWC may be attributed to differences in the ability of water absorption from the soil and/or controlling water loss through the stomata. It may also be caused by differentiated accumulation and osmotic adjustment to maintain tissue turgor and hence physiological activities [
27]. Soluble sugars and proline are well known as the two most important organic solutes and their increase is commonly observed in response to environmental stress. Many studies indicate that evaluation of proline accumulation can be useful in the assessment of drought tolerance in cereals [
33].
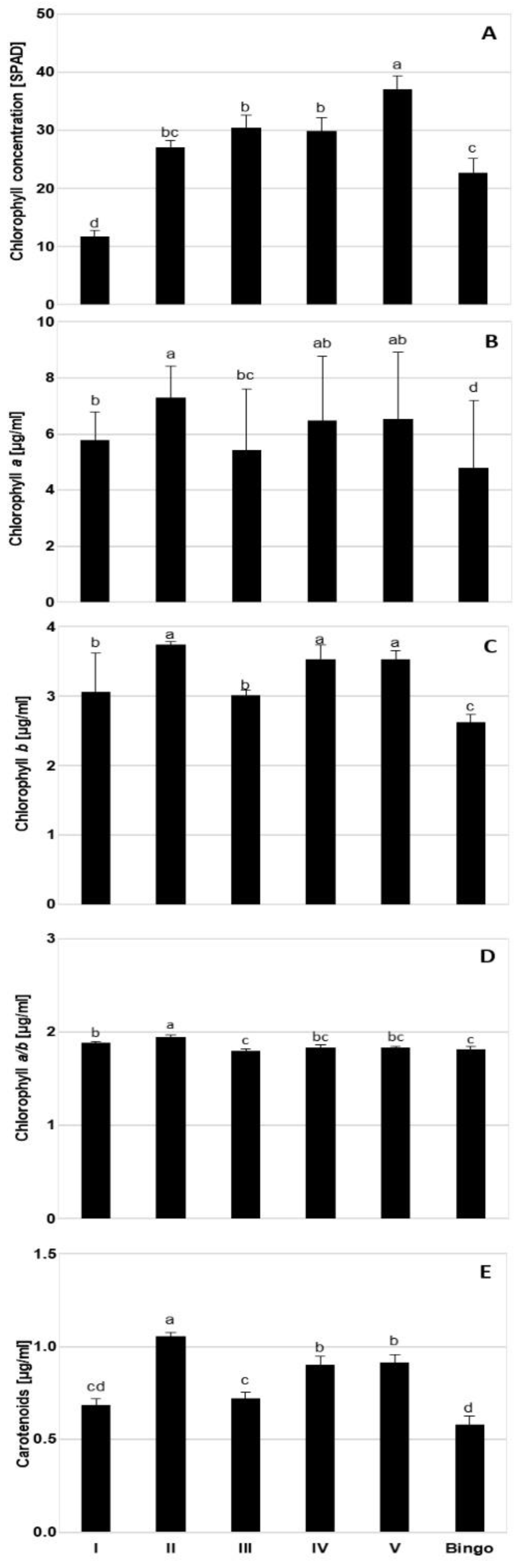
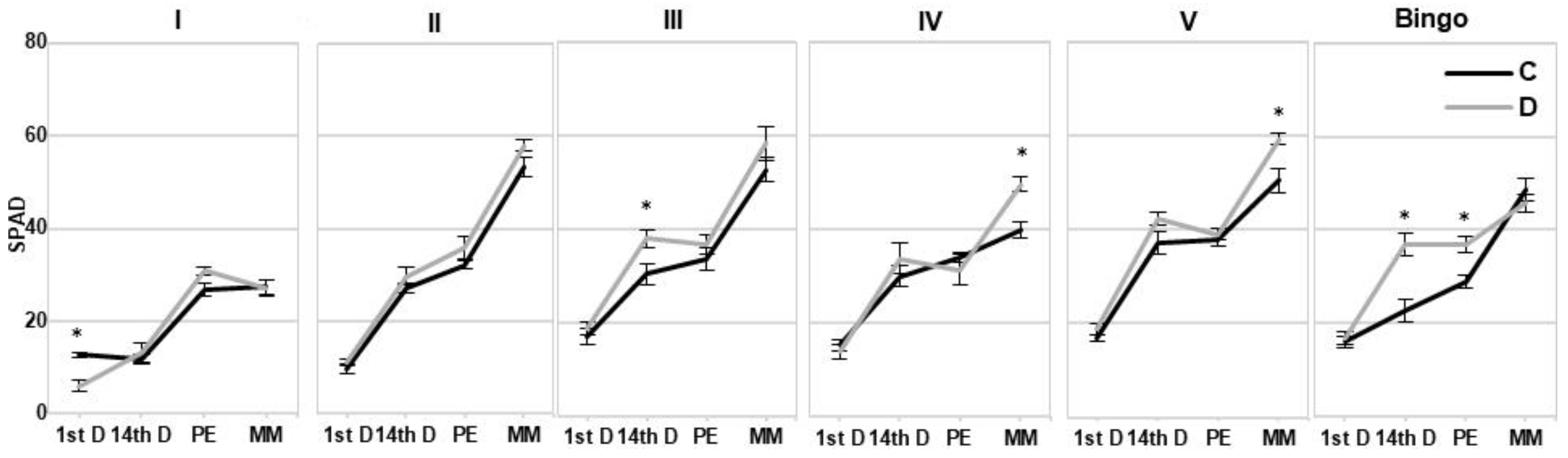
Despite the observed reduction in LRWC values we did not note such a clear response in the chlorophyll content under drought conditions. Chl degradation is listed as one of the consequences of drought stress that may result from sustained photoinhibition and photo-bleaching [
34], even though other plant processes, such as cell division and cell expansion, are the earliest to respond to water deficit stress [
35]. According to O’Neill et al. [
36] a decline in SPAD index is a sensitive and readily measurable trait that could be used to screen for stress tolerance. Cassol et al. [
37] showed that the SPAD index was linear and positively correlated with the Chl content, despite the fact that the reading of the instrument cannot be regarded as the absolute content of the Chl in the leaves. However, our results did not confirm these conclusions. Moreover, presented studies showed a significant discrepancy between SPAD index and chlorophyll content, particularly in lines IV and V, at kernel milk maturity phase—an increase of SPAD values was not reflected in the content of Chl measured spectrophotometrically. Therefore, it seems that the SPAD should not be treated as an independent indicator of tolerance, especially under mild drought stress, which was also observed in our earlier studies [
31]. Based on our experience, this parameter can be used to illustrate some general physiological conditions in plants.
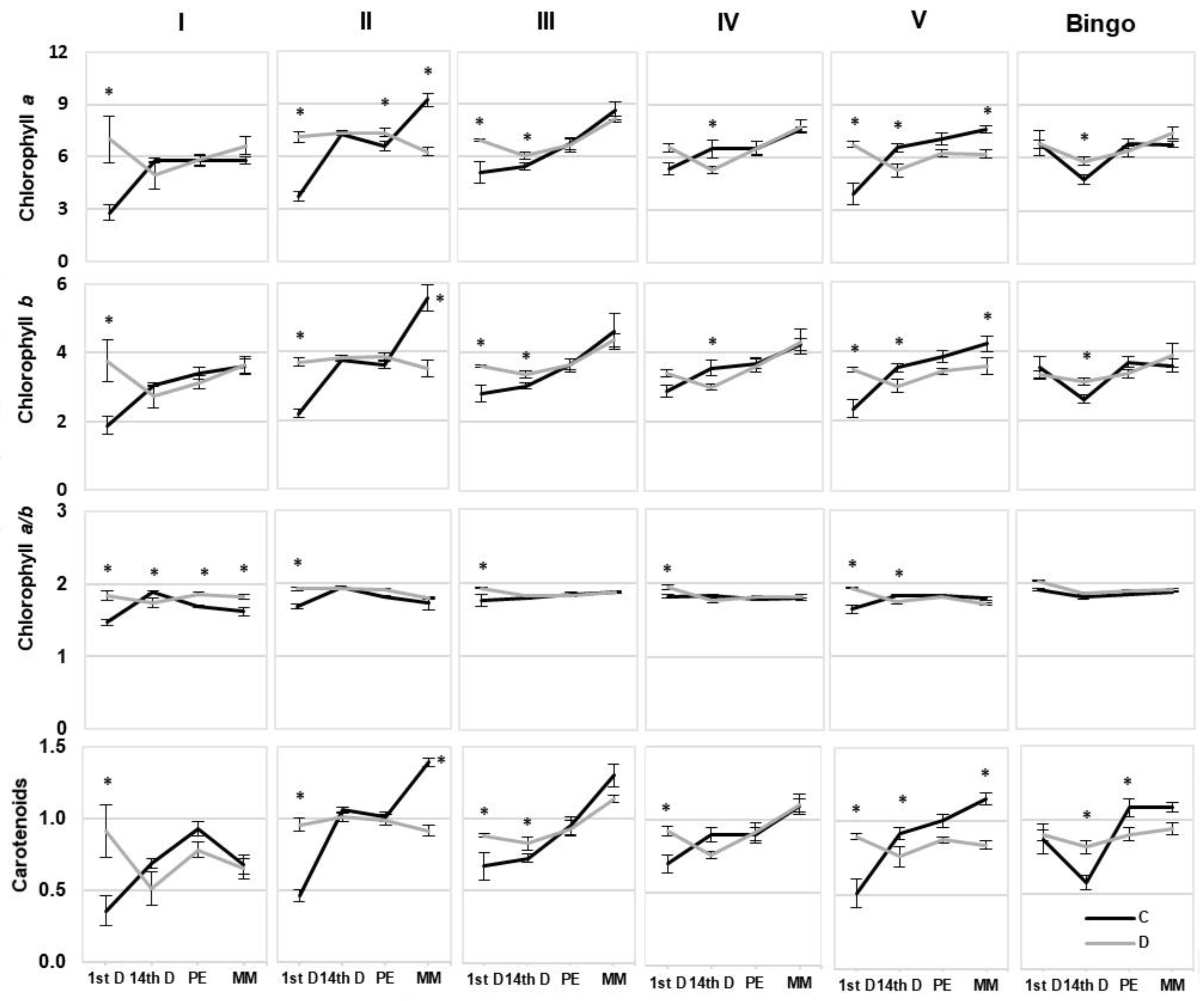
It is known that drought stress causes not only substantial damage to photosynthetic pigments, but also leads to deterioration of thylakoid membranes [
38,
39]. Thus, a reduction in photosynthetic capacity in plants exposed to drought stress is expected [
40], and the decrease in Chl content is a commonly observed phenomenon [
41,
42]. However, there are also some reports which show an enhanced accumulation of Chl under drought stress [
43,
44]. Ashraf and Karim [
45] suggested that it may be due to the variation in Chl synthesis among the cultivars mediated by the variation in the activities of specific enzymes involved in the biosynthesis of Chl. Chl levels vary with foliage maturity, nutrient levels, environmental conditions, light availability, and also seasonally. Thus, plants continuously regulate their photosynthetic processes in response to the changing environmental conditions on diurnal and seasonal timescales [
46]. Our results also showed varied changes in photosynthetic pigment content, through an increase on the 1st day of drought in tested OMA lines, with no alteration in cv. Bingo. We observed a significant decrease in Chl content in lines II and V in the kernel milk maturity phase, which probably is related to the greater sensitivity of these lines to drought associated with accelerated plant aging. On the other hand, line III and cv. Bingo were the only plants characterized by an increase in photosynthetic pigments, both chlorophylls and carotenoids, on the 14th day of drought. Moreover, it is generally known that under drought stress the reduction of Chl
b is greater than that of Chl
a which directly translates into Chl
a/b value [
47]. In our study both Chl forms presented quite a similar course of changes within the tested line/cv. Surprisingly, in line I an increase in Chl
a/b occurred despite no significant changes in the content of individual Chl. Similar results were obtained in wheat, where a slight increase in Chl
a/b ratio in drought tolerant cultivars under water deficit conditions was observed [
48].
Chl absorbed light energy which is then transformed into Chl fluorescence (CF) [
49]. In vivo Chl
a fluorescence may be used as a direct indicator of photosynthetic activity giving a valuable insight into the exploitation of the excitation energy by PSII, and indirectly by other protein complexes of the thylakoid membranes [
50]. CF delivers the information about the energy absorption, the utilization, and the electron transport in PSII [
51]. Therefore, the functional state of the photosynthetic apparatus, based on fluorescence methods, is a useful physiological indicator to study the sensibility of plants to environmental abiotic stress, including drought [
52,
53,
54,
55]. Moreover, CF is considered to be a non-invasive tool for the detection of plant photosynthetic performance both under optimal and adverse conditions [
33]. CF-modulated parameters are commonly used in the JIP-test. In our study, we assayed CF parameters to characterize the functioning of the photosynthetic apparatus of oat × maize hybrids.
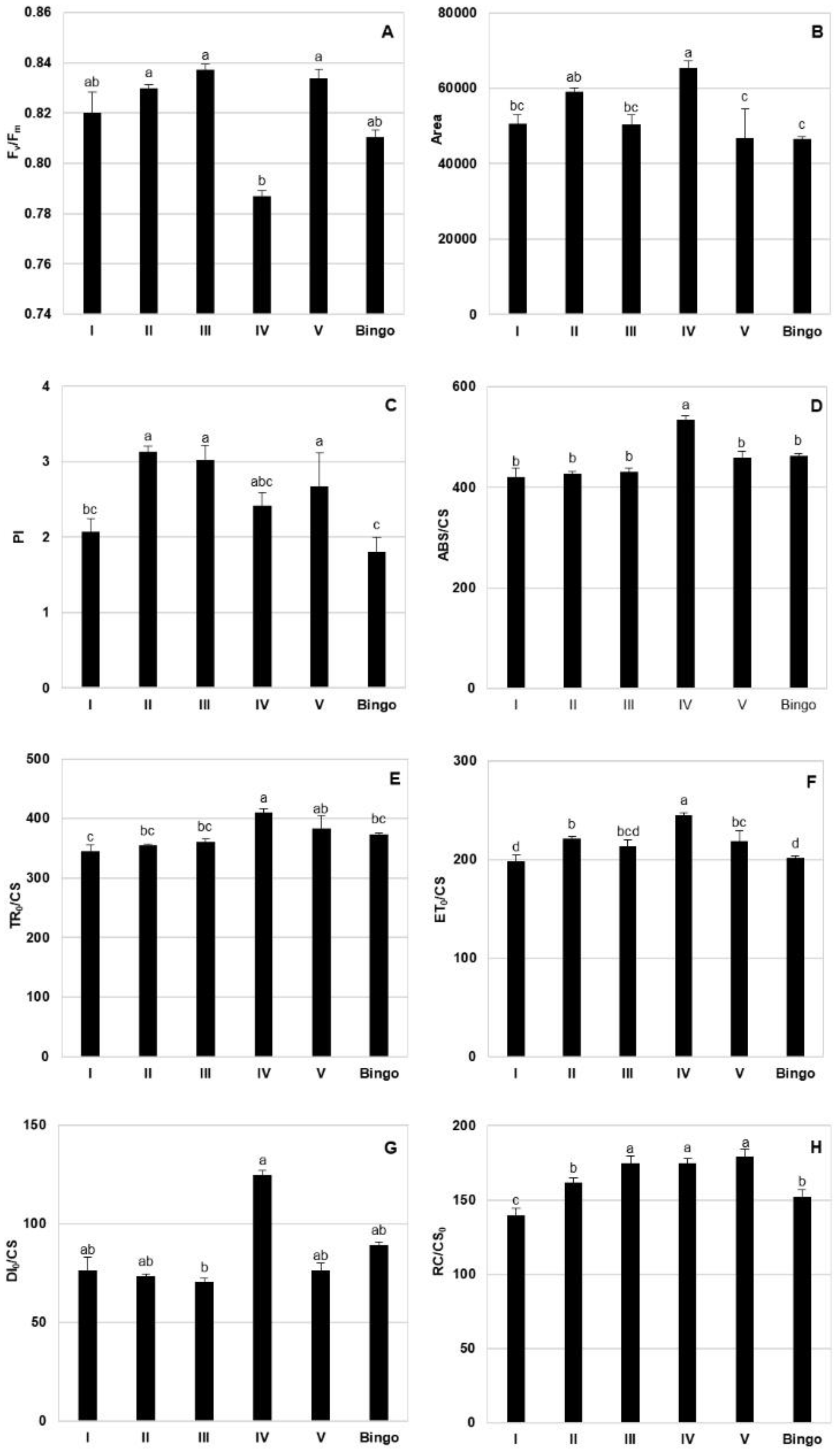
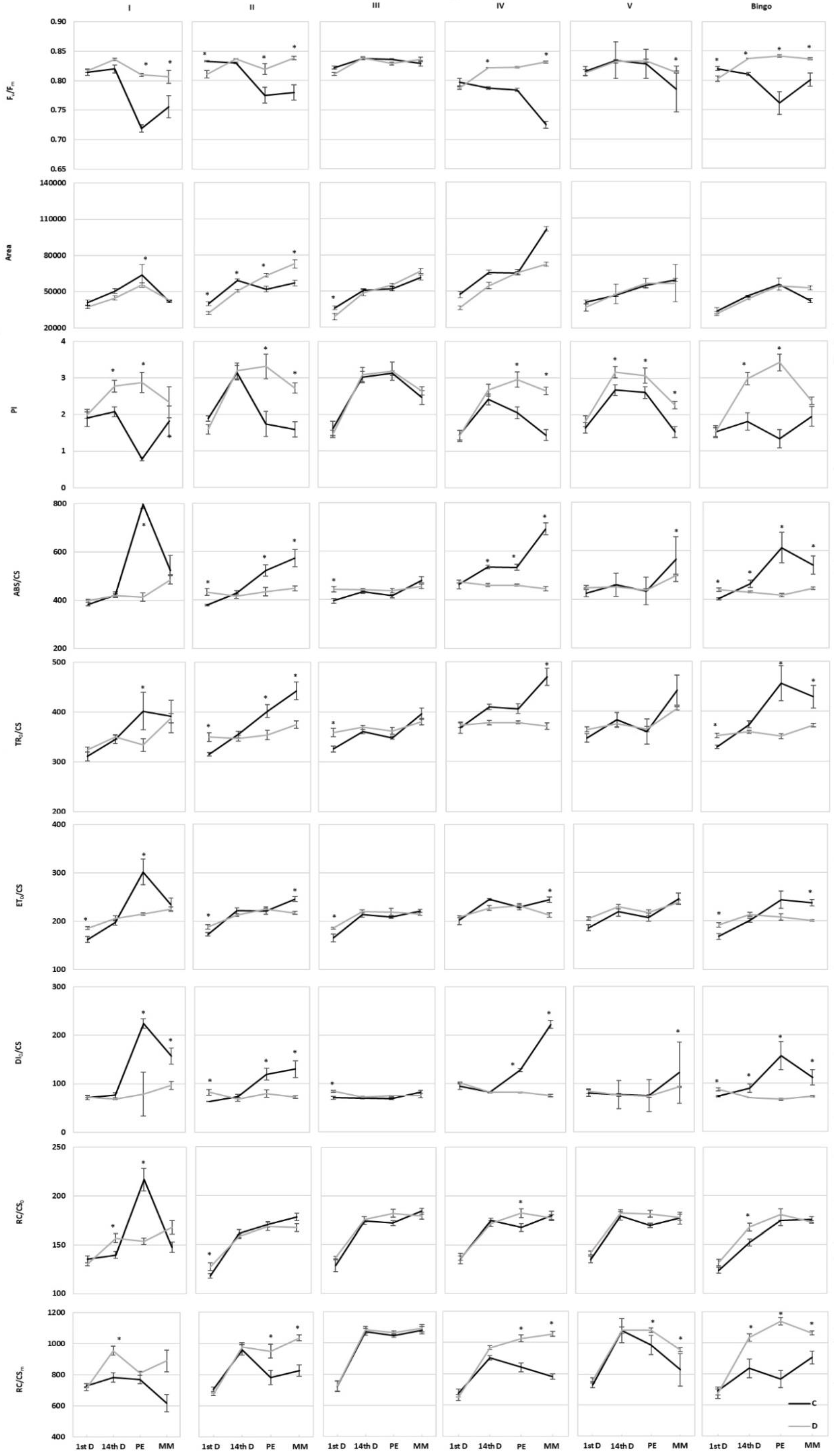
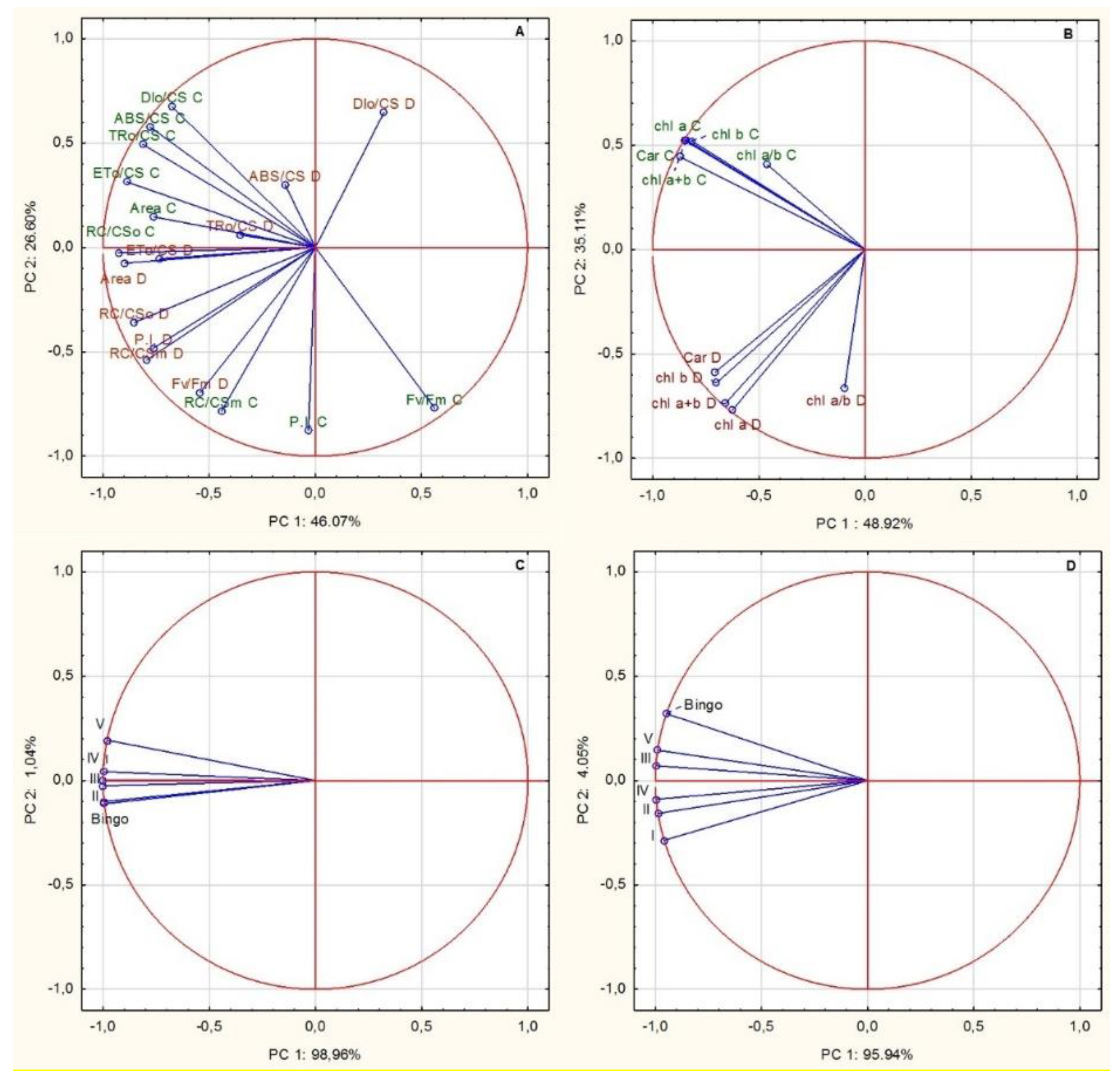
The presented results seem to indicate a different response to drought stress of examined lines, allowing us to divide them, based on the similarity of the value and direction of changes in CF parameters, into two groups: (a) line III and V, in which hardly any significant changes in most parameters were shown, and (b) other hybrid lines and cv. Bingo, characterized by a similar response to applied drought stress (similar course of parameter changes and change of their values i.e., increase/decrease, in drought conditions). Nevertheless, plants of the V line differed from line III with a higher PI and RC/CS
m, which probably may indicate a more efficient functioning of the photosynthetic apparatus in drought conditions despite the observed decrease in chlorophyll content in drought conditions. Our results revealed some changes in the PI parameter which is sensitive to changes in either antenna properties, trapping efficiency or electron transport beyond QA. The F
v/F
m ratio is the most commonly used CF parameter characterizing the maximum quantum yield of PSII, which in most higher plants under physiological conditions ranges from 0.78 to 0.84. In the presented study, its values were within the range indicated by the literature, regardless of the conditions of plant growth and the date of taken measurements. Furthermore, in oat DH lines after 14 days of drought, F
v/F
m did not differ significantly [
55]. In most of the tested OMA lines in drought condition, increased F
v/F
m was not linked to an increase in yield as Liang et al. [
56] reported in wheat. There are also some studies where stress did not alter this parameter [
57,
58]. Its increase observed in several cases in drought conditions seems to be more associated with adaptation to stress than the reflection of damage. Our results are consistent with Antonkiewicz and Rapacz [
59] which consider F
v/F
m alteration as a symptom of adaptive or developmental changes. The direction of changes seems to depend on the level of stress because Souza et al. [
60] proved that a significant decrease of F
v/F
m occurred only in case of severe drought stress.
Area, ET
0/CS and RC/CS
0 showed only slight changes during plant development. An exception was line I, in which these values decreased in drought at panicle ejection phase. Therefore, reduced ET
0/CS indicates that active reaction centers (RCs) are converted into inactive RCs, reducing the efficiency of trapping and a decline in PSII activity [
61]. According to Czyczyło-Mysza et al. [
62] higher values of ET
0/CS, RC/CS
m, as well as ABS/CS, TR
0/CS, and PI, indicate better functioning of photosynthetic apparatus. The observed decrease of the ABS/CS ratio under drought stress in our experiment was possibly due to inactivation of some PSII RCs or a decrease in antenna size. In some of the tested OMA lines we observed an increase of RC/CS
m which provides information about the number of active reaction centers. In our study most of the tested lines showed higher performance index (PI) values in drought condition which occurred with a simultaneous decrease of energy dissipated from PSII (DI
0/CS). A similar relationship was observed in the oat DH lines [
55]. The relatively constant number of active reaction centers remaining in the tested lines despite drought conditions may probably explain the lack of a significant decrease of energy used for electron transport (ET
0/CS).
According to the literature data, OMA lines can be screened for the effects of individual maize chromosomes on photosynthesis in oat [
7,
9,
63]. Since oat is a C3 species and maize is C4, the OMAs may provide a way for the introgression of C4 photosynthesis into oat [
7,
9,
10]. When genes encoding phosphoenolpyruvate carboxylase (PEPC), orthophosphate dikinase (PPdK), pyruvate and the 29-oxoglutarate/malate transporter in maize were expressed in oat, larger bundle sheath cells with increased cell wall lipid deposition were observed in oat leaves [
63]. In particular, OMA lines that contained maize chromosomes 6 and 9 were shown to accumulate maize PEPCase and PPdK [
7]. Furthermore, both enzymes were active, suggesting that oat PEPC and PPdK regulatory protein can phosphorylate the maize proteins. Moreover, high expression of C4 enzymes in C3 plants may play an important role in improvement of the protection of the photosynthetic apparatus against environmental stress, and hence increase grain yield [
64]. However, even in lines with both chromosomes present, photosynthesis was more C3-like than C4 [
65]. Despite OMA lines providing insight into certain aspects of C4 regulation they failed to reveal global regulators of the pathway [
65], thus the actual function of C4 enzymes in C3 plants still remains unknown [
64].

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}