The Hirudo Medicinalis Microbiome Is a Source of New Antimicrobial Peptides

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Metagenome-Wide Search for Antimicrobial Peptides
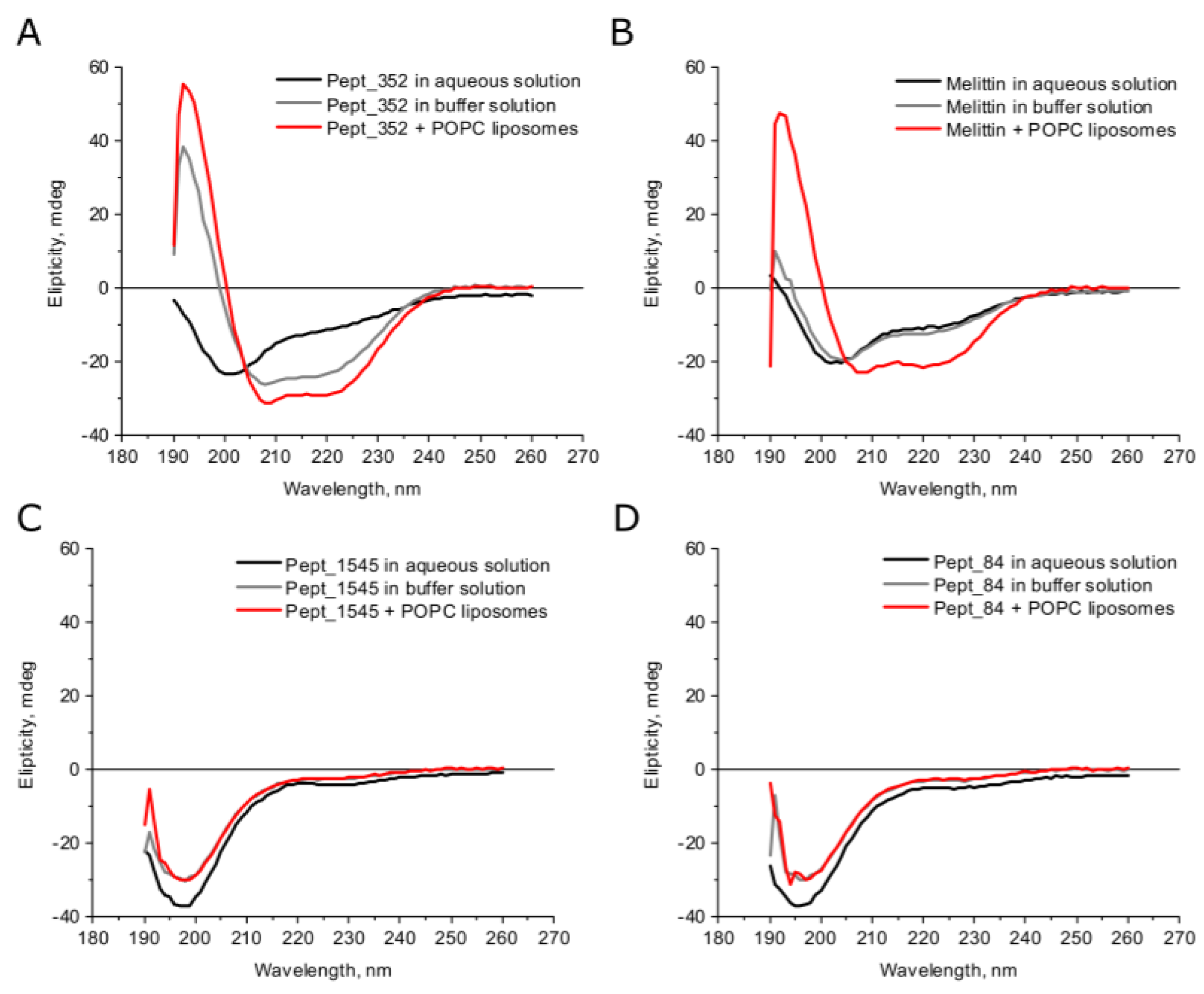
2.2. Secondary Structures of AMPs
2.3. Antimicrobial Activity of AMPs
2.4. Cytotoxic Assays of AMPs
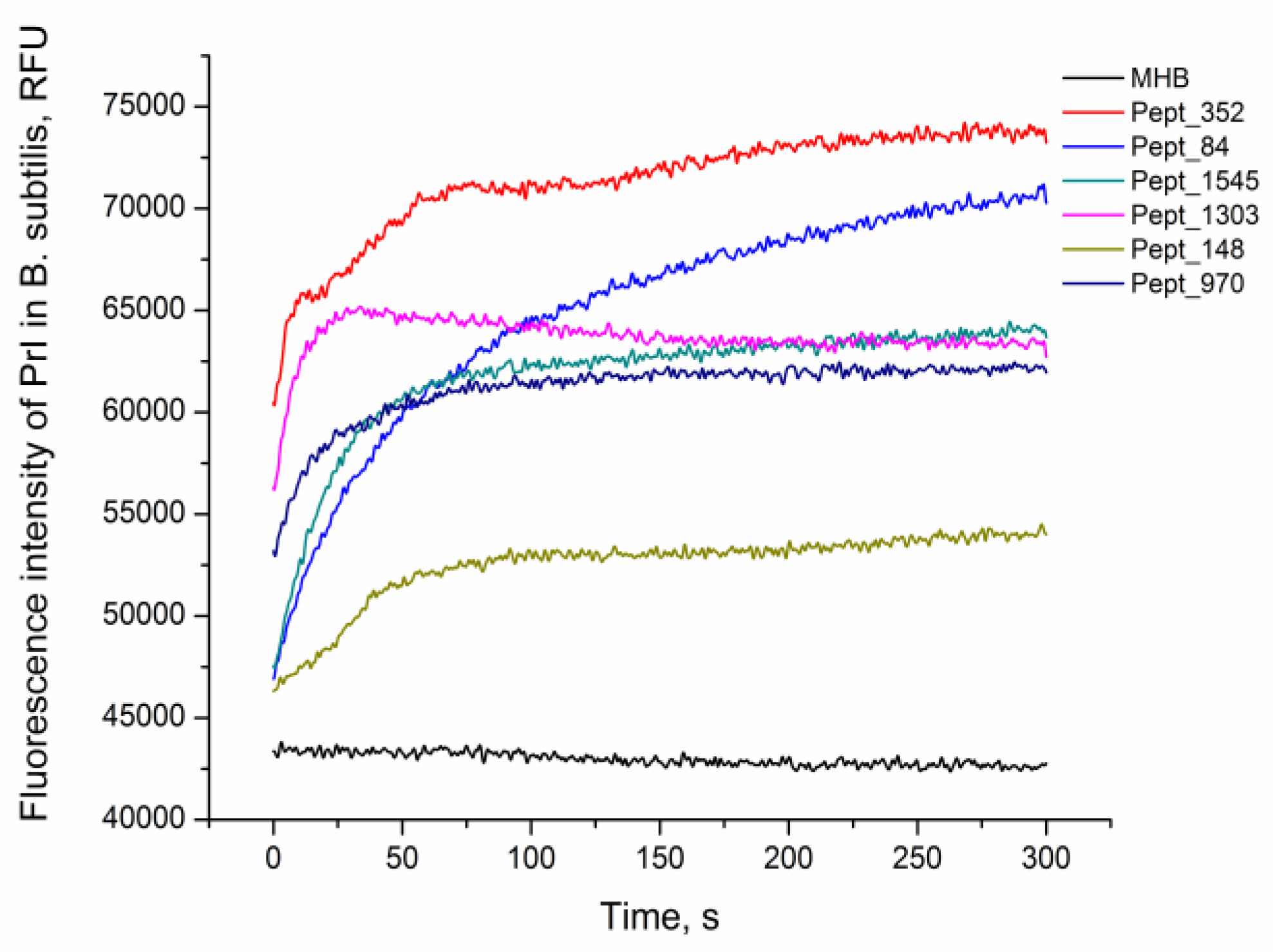
2.5. AMP Activity by Propidium Iodide Penetration Assay
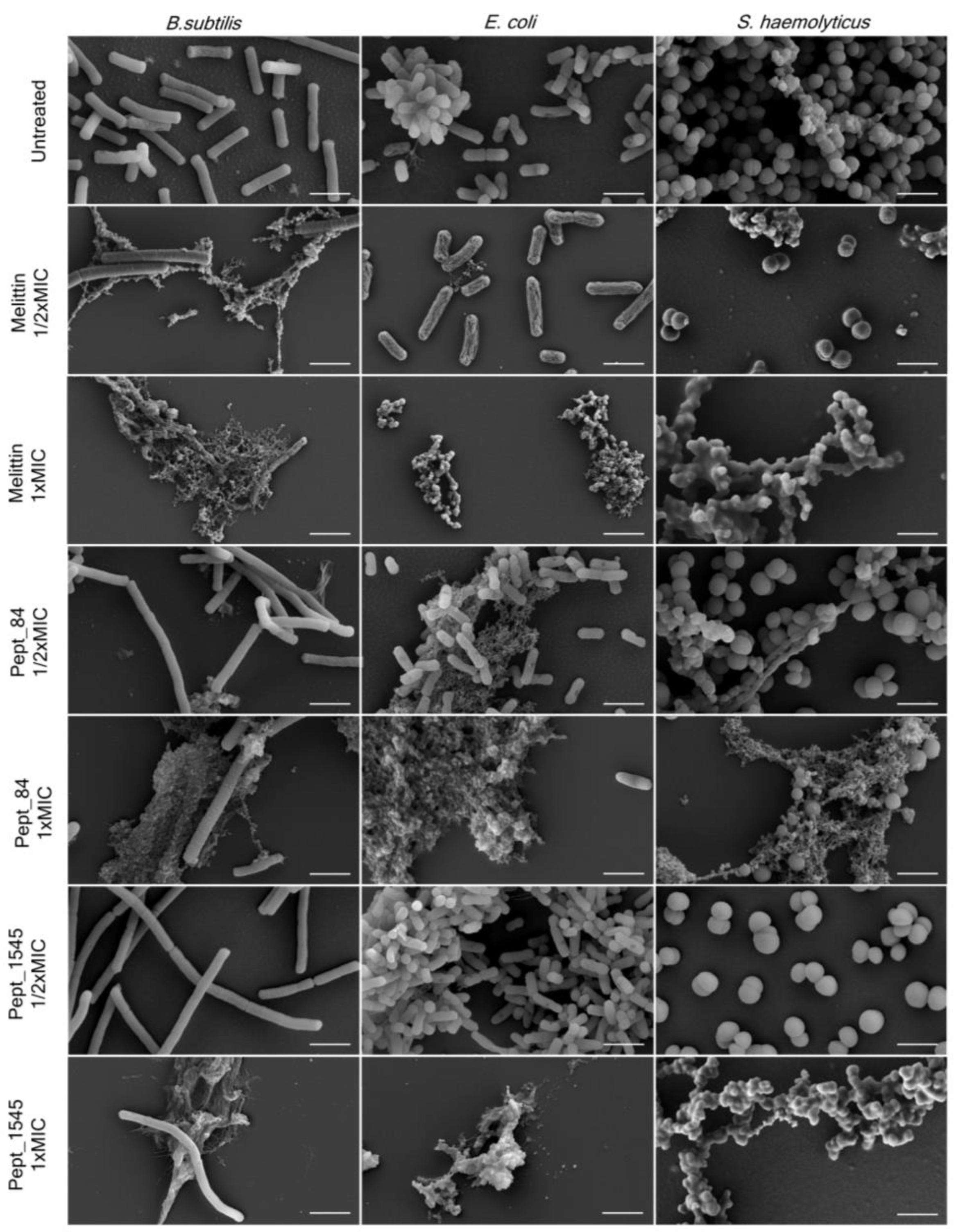
2.6. Bacterial Cell Morphological Changes after AMPs Exposure
3. Discussion
4. Materials and Methods
4.1. Data Set Analysis
4.2. Peptide Synthesis
4.3. Antimicrobial Activity Assay
4.4. Circular Dichroism Spectroscopy
4.5. Viability of Bacteria by PI Penetration
4.6. Cytotoxic and Cell Viability Assays
4.7. Scanning Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMP | Antimicrobial peptides |
| CD | Circular dichroism |
| MIC | Minimum inhibitory concentration |
| POPC | Phosphatidylcholine |
| HMDS | Hexamethyldisilazane |
| LDH | Lactate dehydrogenase |
References
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Lohner, K.; Hilpert, K. Antimicrobial peptides: Cell membrane and microbial surface interactions. Biochim. Biophys. Acta-Biomembr. 2016, 1858, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Campagna, S.; Saint, N.; Molle, G.; Aumelas, A. Structure and mechanism of action of the antimicrobial peptide piscidin. Biochemistry 2007, 46, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula–Thomas, S. CAMP R3 : A database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 2015, 44, D1094–D1097. [Google Scholar] [CrossRef] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defense peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020. [Google Scholar] [CrossRef]
- Cheng, K.T.; Wu, C.L.; Yip, B.S.; Chih, Y.H.; Peng, K.L.; Hsu, S.Y.; Yu, H.Y.; Cheng, J.W. The interactions between the antimicrobial peptide p–113 and living Candida albicans cells shed light on mechanisms of antifungal activity and resistance. Int. J. Mol. Sci. 2020, 21, 2654. [Google Scholar] [CrossRef] [Green Version]
- Bouza, E.; Valerio, M.; Soriano, A.; Morata, L.; Carus, E.G.; Rodríguez–González, C.; Hidalgo–Tenorio, M.C.; Plata, A.; Muñoz, P.; Vena, A.; et al. Dalbavancin in the treatment of different Gram–positive infections: A real–life experience. Int. J. Antimicrob. Agents 2018, 51, 571–577. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Sothiselvam, S.; Lu, T.K.; de la Fuente–Nunez, C. Peptide design principles for antimicrobial applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef]
- Ohtsuka, Y.; Inagaki, H. In silico identification and functional validation of linear cationic α–helical antimicrobial peptides in the ascidian Ciona intestinalis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Grafskaia, E.N.; Nadezhdin, K.D.; Talyzina, I.A.; Polina, N.F.; Podgorny, O.V.; Pavlova, E.R.; Bashkirov, P.V.; Kharlampieva, D.D.; Bobrovsky, P.A.; Latsis, I.A.; et al. Medicinal leech antimicrobial peptides lacking toxicity represent a promising alternative strategy to combat antibiotic–resistant pathogens. Eur. J. Med. Chem. 2019, 180, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Marden, J.N.; McClure, E.A.; Beka, L.; Graf, J. Host matters: Medicinal leech digestive–tract symbionts and their pathogenic potential. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, J.P.; Lemke, S. Small bite, large impact–saliva and salivary molecules in the medicinal leech, Hirudo medicinalis. Naturwissenschaften 2011, 98, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Indergand, S.; Graf, J. Ingested blood contributes to the specificity of the symbiosis of Aeromonas veronii biovar sobria and Hirudo medicinalis, the medicinal leech. Appl. Environ. Microbiol. 2000, 66, 4735–4741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beka, L.; Fullmer, M.S.; Colston, S.M.; Nelson, M.C.; Talagrand–Reboul, E.; Walker, P.; Ford, B.; Whitaker, I.S.; Lamy, B.; Gogarten, J.P.; et al. Low–level antimicrobials in the medicinal leech select for resistant pathogens that spread to patients. MBio 2018, 9, e01328-18. [Google Scholar] [CrossRef] [Green Version]
- Babenko, V.V.; Podgorny, O.V.; Manuvera, V.A.; Kasianov, A.S.; Manolov, A.I.; Grafskaia, E.N.; Shirokov, D.A.; Kurdyumov, A.S.; Vinogradov, D.V.; Nikitina, A.S.; et al. Draft genome sequences of Hirudo medicinalis and salivary transcriptome of three closely related medicinal leeches. BMC Genomics 2020, 21, 20–21. [Google Scholar] [CrossRef]
- Pirtskhalava, M.; Gabrielian, A.; Cruz, P.; Griggs, H.L.; Squires, R.B.; Hurt, D.E.; Grigolava, M.; Chubinidze, M.; Gogoladze, G.; Vishnepolsky, B.; et al. DBAASP v.2: An enhanced database of structure and antimicrobial/cytotoxic activity of natural and synthetic peptides. Nucleic Acids Res. 2016, 44, D1104–D1112. [Google Scholar] [CrossRef]
- Torrent, M.; Di Tommaso, P.; Pulido, D.; Nogués, M.V.; Notredame, C.; Boix, E.; Andreu, D. AMPA: An automated web server for prediction of protein antimicrobial regions. Bioinformatics 2012, 28, 130–131. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-T.; Lee, C.-C.; Yang, J.-R.; Lai, J.Z.C.; Chang, K.Y. A large–scale structural classification of antimicrobial peptides. Biomed. Res. Int. 2015, 2015, 475062. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, P.; Lin, W.Z.; Jia, J.H.; Chou, K.C. IAMP–2L: A two–level multi–label classifier for identifying antimicrobial peptides and their functional types. Anal. Biochem. 2013, 436, 168–177. [Google Scholar] [CrossRef]
- Bhadra, P.; Yan, J.; Li, J.; Fong, S.; Siu, S.W.I. AmPEP: Sequence–based prediction of antimicrobial peptides using distribution patterns of amino acid properties and random forest. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Virtanen, J.; Xue, Z.; Zhang, Y. I–TASSER–MR: Automated molecular replacement for distant–homology proteins using iterative fragment assembly and progressive sequence truncation. Nucleic Acids Res. 2017, 45, W429–W434. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.C.; Strandberg, E.; Grau–Campistany, A.; Wadhwani, P.; Reichert, J.; Bürck, J.; Rabanal, F.; Auger, M.; Paquin, J.F.; Ulrich, A.S. Influence of the length and charge on the activity of α–helical amphipathic antimicrobial peptides. Biochemistry 2017, 56, 1680–1695. [Google Scholar] [CrossRef] [PubMed]
- Hristova, K.; Dempsey, C.E.; White, S.H. Structure, location, and lipid perturbations of melittin at the membrane interface. Biophys. J. 2001, 80, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Wiedman, G.; Khan, A.; Ulmschneider, M.B. Absorption and folding of melittin onto lipid bilayer membranes via unbiased atomic detail microsecond molecular dynamics simulation. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 2243–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meylan, P.; Lang, C.; Mermoud, S.; Johannsen, A.; Norrenberg, S.; Hohl, D.; Vial, Y.; Prod’hom, G.; Greub, G.; Kypriotou, M.; et al. Skin colonization by staphylococcus aureus precedes the clinical diagnosis of atopic dermatitis in infancy. J. Invest. Dermatol. 2017, 137, 2497–2504. [Google Scholar] [CrossRef] [Green Version]
- Moravej, H.; Moravej, Z.; Yazdanparast, M.; Heiat, M.; Mirhosseini, A.; Moosazadeh Moghaddam, M.; Mirnejad, R. Antimicrobial peptides: Features, action, and their resistance mechanisms in bacteria. Microb. Drug Resist. 2018, 24, 747–767. [Google Scholar] [CrossRef]
- Sun, E.; Belanger, C.R.; Haney, E.F.; Hancock, R.E.W. Host defense (antimicrobial) peptides. In Peptide Applications in Biomedicine, Biotechnology and Bioengineering; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; pp. 253–285. [Google Scholar] [CrossRef]
- Torres, M.D.T.; de la Fuente–Nunez, C. Toward computer–made artificial antibiotics. Curr. Opin. Microbiol. 2019, 51, 30–38. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, Á.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In silico optimization of a guava antimicrobial peptide enables combinatorial exploration for peptide design. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Oshiro, K.G.N.; Cândido, E.S.; Chan, L.Y.; Torres, M.D.T.; Monges, B.E.D.; Rodrigues, S.G.; Porto, W.F.; Ribeiro, S.M.; Henriques, S.T.; Lu, T.K.; et al. Computer–aided design of mastoparan–like peptides enables the generation of nontoxic variants with extended antibacterial properties. J. Med. Chem. 2019, 62, 8140–8151. [Google Scholar] [CrossRef]
- Hoeksema, M.; Van Eijk, M.; Haagsman, H.P.; Hartshorn, K.L. Histones as mediators of host defense, inflammation and thrombosis. Future Microbiol. 2016, 11, 441–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Hwang, J.S.; Lee, D.G. Antifungal effect and pore–forming action of lactoferricin b like peptide derived from centipede scolopendra Subspinipes mutilans. Biochim. Biophys. Acta-Biomembr. 2013, 1828, 2745–2750. [Google Scholar] [CrossRef] [Green Version]
- Maltz, M.A.; Bomar, L.; Lapierre, P.; Morrison, H.G.; McClure, E.A.; Sogin, M.L.; Graf, J. Metagenomic analysis of the medicinal leech gut microbiota. Front. Microbiol. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desriac, F.; Jégou, C.; Balnois, E.; Brillet, B.; Le Chevalier, P.; Fleury, Y. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddall, M.E.; Perkins, S.L.; Desser, S.S. Leech mycetome endosymbionts are a new lineage of alphaproteobacteria related to the Rhizobiaceae. Mol. Phylogenet. Evol. 2004, 30, 178–186. [Google Scholar] [CrossRef]
- Ott, B.M.; Rickards, A.; Gehrke, L.; Rio, R.V.M. Characterization of shed medicinal leech mucus reveals a diverse microbiota. Front. Microb. 2015, 5, 757. Available online: https://www.frontiersin.org/article/10.3389/fmicb.2014.00757 (accessed on 1 September 2015). [CrossRef] [Green Version]
- McClure, E.A.; Nelson, M.C.; Lin, A.; Graf, J. Macrobdella decora: Old world leech gut microbial community structure conserved in a new world leech. bioRxiv 2019, 687418. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.C.; Graf, J. Bacterial symbioses of the medicinal leech Hirudo verbana. Gut Microbes 2012, 3, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Modry, D.; Pafčo, B.; Zurek, L. Bacterial Community of the Digestive Tract of the European Medicinal Leech (Hirudo verbana) from the Danube River. Microb. Ecol. 2019, 77, 1082–1090. [Google Scholar] [CrossRef]
- Polina, N.; Babenko, V.; Grafskaia, E.; Podgorny, O.; Bobrovsky, P.; Kharlampieva, D.; Belova, A.; Shirokov, D.; Miroshina, O.; Manuvera, V.; et al. A digestive system microbiota of medicinal leech hirudo medicinalis. FEBS OPEN BIO 2018, 8 (Suppl. 1), 462. [Google Scholar]
- Urushibara, T. Effect of liposome surface charge and peptide side chain charge density on antimicrobial peptide–membrane binding as determined by circular dichroism. J. Membr. Sci. Technol. 2013, 3, 124. [Google Scholar] [CrossRef]
- Durham, D.R.; Kloos, W.E. Comparative study of the total cellular fatty acids of Staphylococcus species of human origin. Int. J. Syst. Bateriol. 1978, 28, 223–228. [Google Scholar]
- Tiwari, K.B.; Gatto, C.; Wilkinson, B.J. Plasticity of Coagulase-Negative Staphylococcal Membrane Fatty Acid Composition and Implications for Responses to Antimicrobial Agents. Antibiotics 2020, 9, 214. [Google Scholar] [CrossRef] [PubMed]
- Jamasbi, E.; Lucky, S.S.; Li, W.; Hossain, M.A.; Gopalakrishnakone, P.; Separovic, F. Effect of dimerized melittin on gastric cancer cells and antibacterial activity. Amino Acids 2018, 50, 1101–1110. [Google Scholar] [CrossRef]
- Zhang, S.-K.; Song, J.; Gong, F.; Li, S.-B.; Chang, H.-Y.; Xie, H.-M.; Gao, H.-W.; Tan, Y.-X.; Ji, S.-P. Design of an α–helical antimicrobial peptide with improved cell-selective and potent anti-biofilm activity. Sci. Rep. 2016, 6, 27394. [Google Scholar] [CrossRef] [Green Version]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane–active mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Hale, J.D.F.; Hancock, R.E.W. Alternative mechanisms of action of cationic antimicrobial peptides on bacteria. Expert Rev. Anti. Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (mic) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Norma, J. Greenfield. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda–Carrera, I.; Frise, E.; Verena, K.; Mark, L.; Tobias, P.; Stephan, P.; Curtis, R.; Stephan, S.; Benjamin, S.; et al. Fiji-an open platform for biological image analysis. Nat. Methods 2009, 9, 10–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolova, A.I.; Pavlova, E.R.; Khramova, Y.V.; Klinov, D.V.; Shaitan, K.V. Imaging human keratinocytes grown on electrospun mats by scanning electron microscopy. Microsc. Res. Tech. 2019, 82, 544–549. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peptide | Organism | Amino Acid Sequence | Length, a. a. | Molecular Weight, Da |
|---|---|---|---|---|
| pept_1303 | Aminobacter | IGRHFKRRNSIWGICWF | 17 | 2176.58 |
| pept_148 | Aminobacter | VLIRGLIHMLRGG | 13 | 1434.81 |
| pept_970 | Sphingobacteriia | FVKILAKLVNYAKN | 14 | 1621 |
| pept_1545 | Chitinophagia | FLIGKAIKRKFCLRSVWNA | 19 | 2250.78 |
| pept_352 | Sphingobacteriia | KKGKSFKQLHIIVHLVKSWLRTILTHI | 27 | 3224.98 |
| pept_84 | Chitinophagia | IVKRFFRISYKLQSLKIIKGKRTFT | 25 | 3071.79 |
| MIC (μM) | ||||
|---|---|---|---|---|
| Peptide | E. coli K12 substr MG1655 | B. subtilis 168HT | S. aureus ST 88 | S. haemolyticus 527 |
| pept_1303 | 14.8 | 14.8 | >100 | 14.8 |
| pept_148 | >100 | 22.4 | >100 | >100 |
| pept_970 | 79 | 19.8 | >100 | >100 |
| pept_1545 | 14.3 | 7.2 | >100 | 14.3 |
| pept_352 | 10 | 10 | 19.9 | 10 |
| pept_84 | 10.5 | 5.3 | >100 | 5.3 |
| Melittin | 5.7 | 1.5 | 1.5 | 5.7 |
| Peptide | Cell Viability, % |
|---|---|
| pept_1303 | 25.1 ± 8.3 * |
| pept_148 | 98.3 ± 0.9 |
| pept_970 | 2.7 ± 2.1 ** |
| pept_1545 | 98.7 ± 1.3 |
| pept_352 | 0.2 ± 0.1 *** |
| pept_84 | 11.2 ± 7.4 * |
| Melittin | 0.3 ± 0.1 *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grafskaia, E.; Pavlova, E.; Babenko, V.V.; Latsis, I.; Malakhova, M.; Lavrenova, V.; Bashkirov, P.; Belousov, D.; Klinov, D.; Lazarev, V. The Hirudo Medicinalis Microbiome Is a Source of New Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197141
Grafskaia E, Pavlova E, Babenko VV, Latsis I, Malakhova M, Lavrenova V, Bashkirov P, Belousov D, Klinov D, Lazarev V. The Hirudo Medicinalis Microbiome Is a Source of New Antimicrobial Peptides. International Journal of Molecular Sciences. 2020; 21(19):7141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197141
Chicago/Turabian StyleGrafskaia, Ekaterina, Elizaveta Pavlova, Vladislav V. Babenko, Ivan Latsis, Maja Malakhova, Victoria Lavrenova, Pavel Bashkirov, Dmitrii Belousov, Dmitry Klinov, and Vassili Lazarev. 2020. "The Hirudo Medicinalis Microbiome Is a Source of New Antimicrobial Peptides" International Journal of Molecular Sciences 21, no. 19: 7141. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197141