Identification of the Lineage Markers and Inhibition of DAB2 in In Vitro Fertilized Porcine Embryos

 , ,
, ,
Abstract
:1. Introduction
2. Results
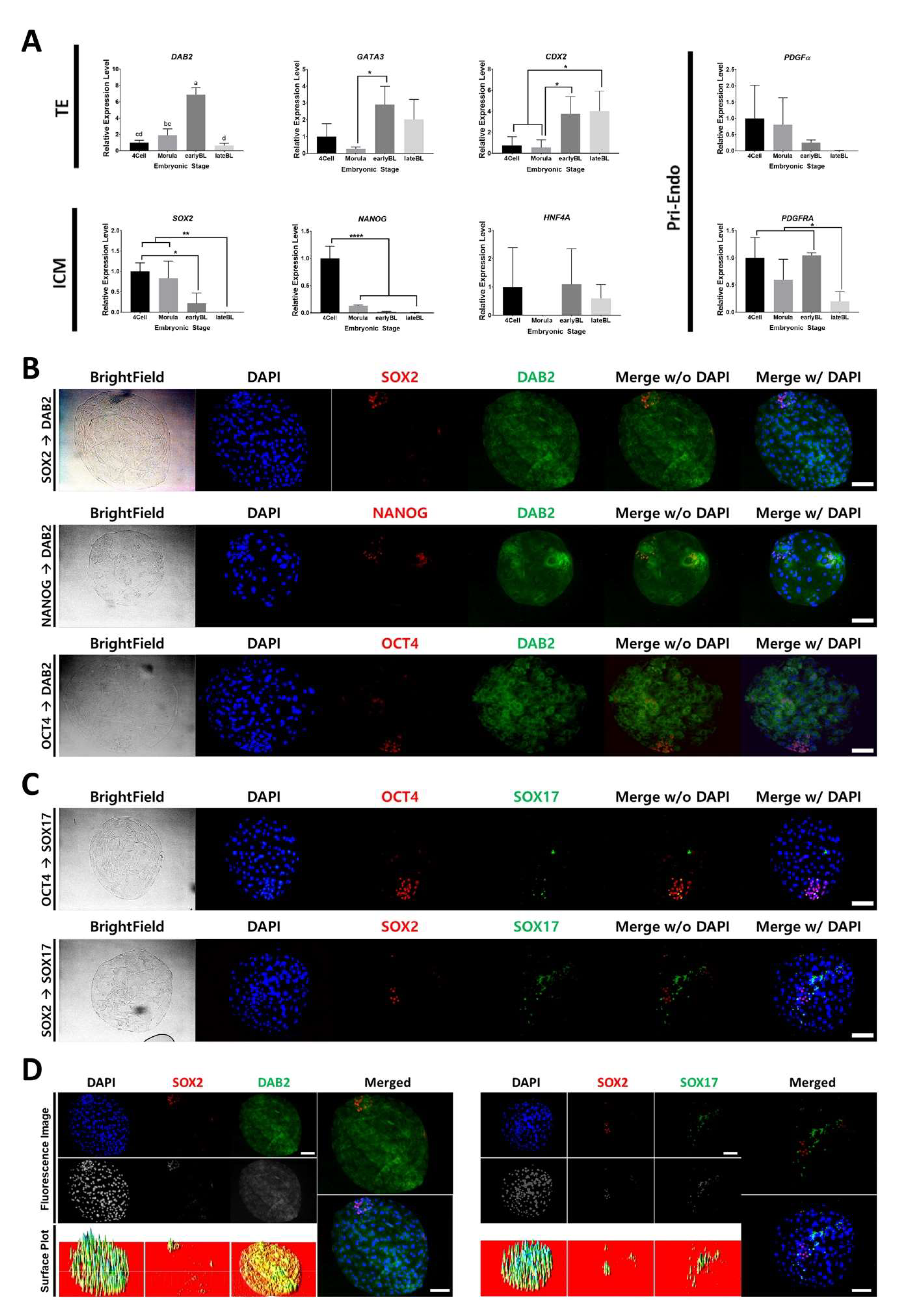
2.1. Marker Gene Expression of IVF Embryos during Development and Distribution of Marker Proteins in Day 7 IVF Embryos
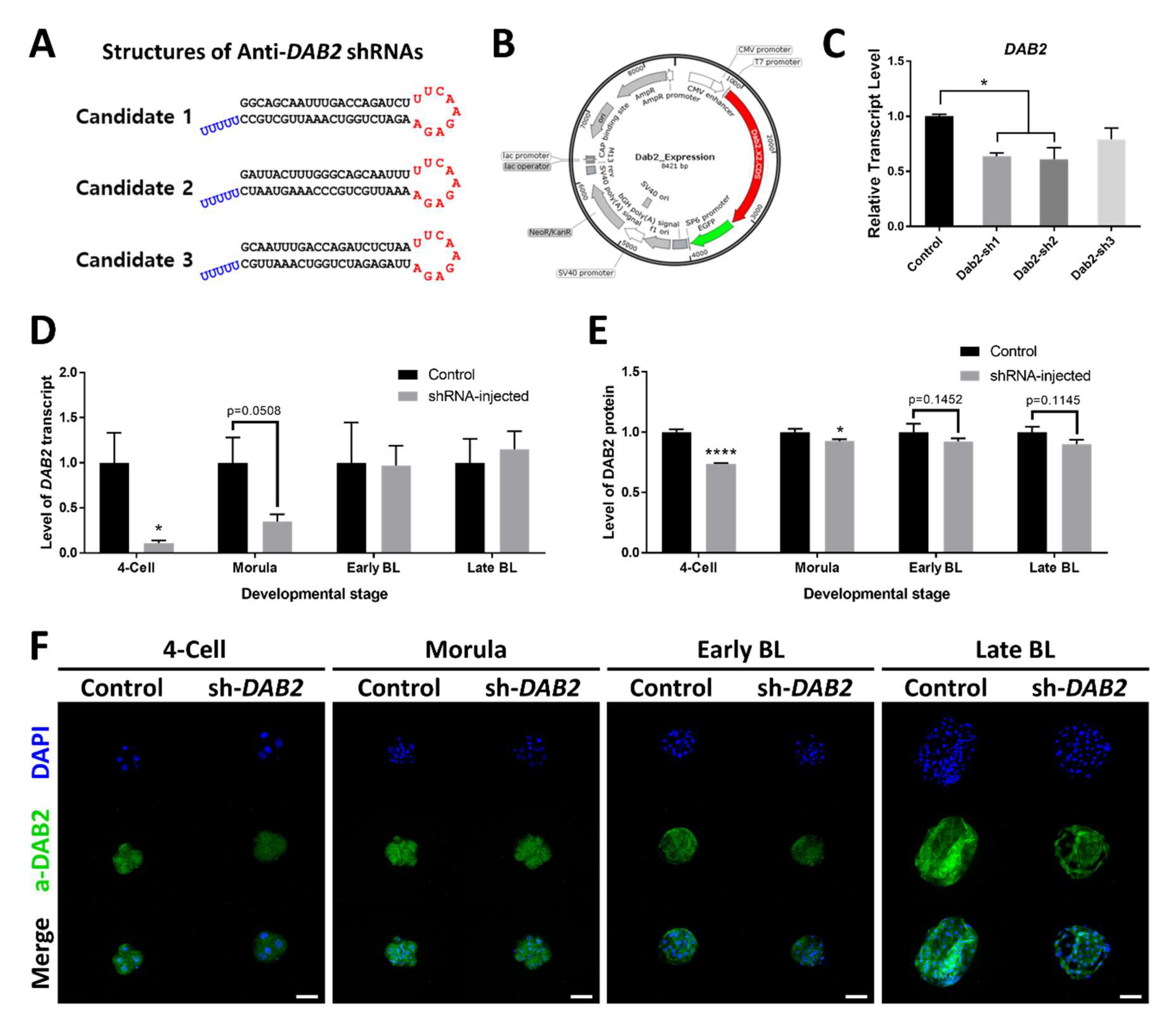
2.2. Cloning of Vectors and Verification of the shRNA Expression Vectors
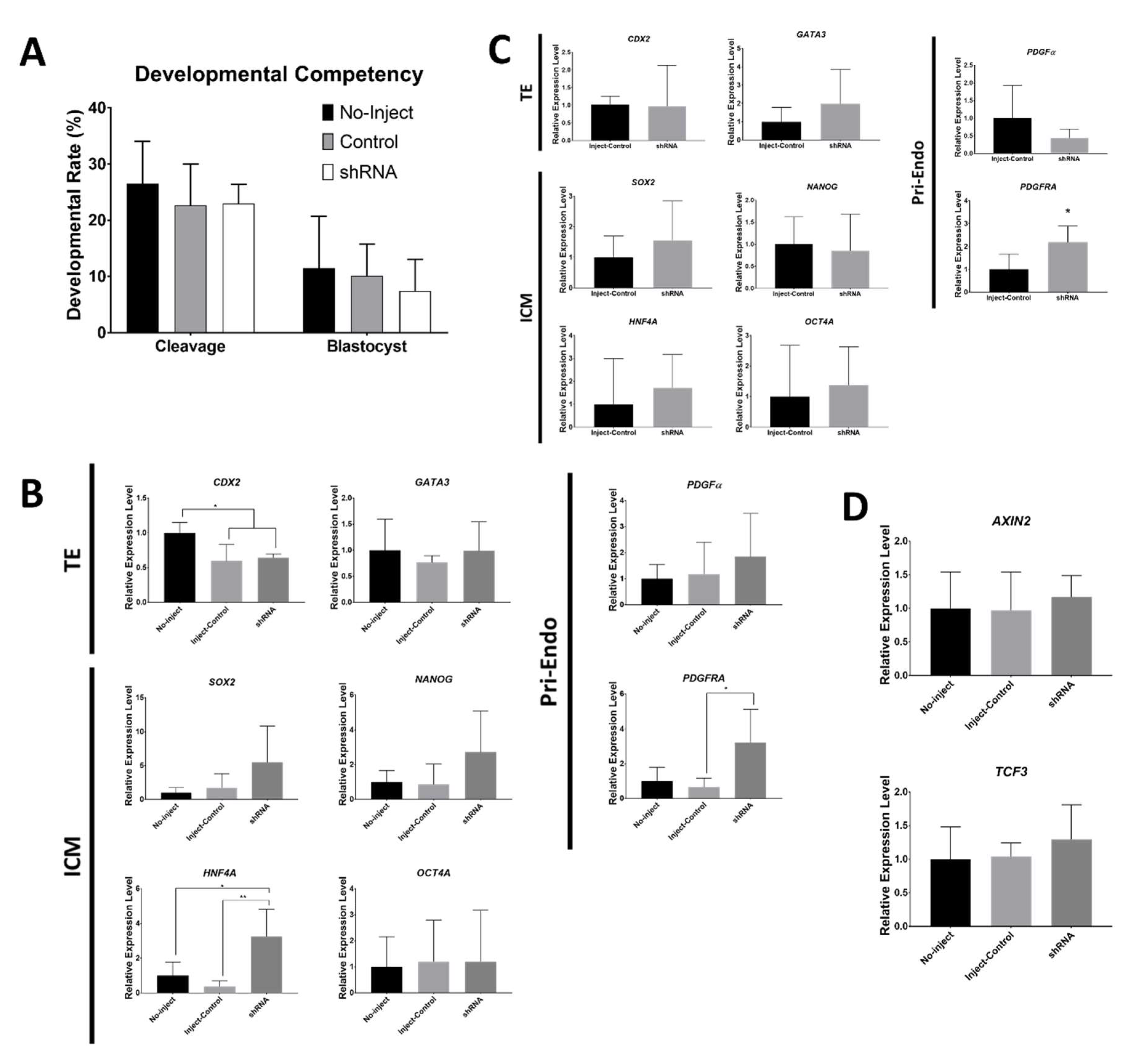
2.3. Marker mRNA Expression and Protein Distribution in shRNA-injected Embryos
3. Discussion
4. Materials and Methods
4.1. In Vitro Production of Fertilized Embryos
4.2. RNA Extraction and Quantitative PCR
4.3. Immunocytochemistry of Embryos
4.4. Production of shRNA-Expressing Vectors and DAB2 Gene Expression Vector
4.5. Culture of Porcine Fetal Fibroblasts and Plasmid Transfection
4.6. Microinjection of Plasmid Vectors into IVF Embryos
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ICM | Inner cell mass |
| TE | Trophectoderm |
| EPI | Epiblast |
| Pri-Endo | Primitive endoderm |
| BL | Blastocyst |
| qPCR | Quantitative PCR |
| IVF | In vitro fertilized |
| pFF | Porcine fetal fibroblasts |
| COCs | Cumulus-oocyte complexes |
| BSA | Bovine serum albumin |
| DPBS | Dulbecco’s Phosphate Buffered Saline |
| mTBM | Modified Tris-buffered medium |
| RT | Room temperature |
Appendix

{kind=link}
{kind=link}
{kind=link}
| Name of oligonucleotide | Sequence | |||
| Anti-Dab2 shRNA-1 | Forward | ccggtGGCAGCAATTTGACCAGATCTTTCAAGAGAAGATCTGGTCAAATTGCTGCCTTTTTg | ||
| Reverse | aattcAAAAAGGCAGCAATTTGACCAGATCTTCTCTTGAAAGATCTGGTCAAATTGCTGCCa | |||
| Anti-Dab2 shRNA-2 | Forward | ccggtGATTACTTTGGGCAGCAATTTTTCAAGAGAAAATTGCTGCCCAAAGTAATCTTTTTg | ||
| Reverse | aattcAAAAAGATTACTTTGGGCAGCAATTTTCTCTTGAAAAATTGCTGCCCAAAGTAATCa | |||
| Anti-Dab2 shRNA-3 | Forward | ccggtGCAATTTGACCAGATCTCTAATTCAAGAGATTAGAGATCTGGTCAAATTGCTTTTTg | ||
| Reverse | aattcAAAAAGCAATTTGACCAGATCTCTAATCTCTTGAATTAGAGATCTGGTCAAATTGCa | |||
| Primers for qPCR | Sequence | Annealing Tm (°C) | Product size (bp) | |
| DAB2 | Forward | TGGGAGTGAGGCCCTAATGA | 58 | 111 bp |
| Reverse | GGACTACTTAGGTCGGGAGGT | 58 | ||
| GATA3 | Forward | GCGGGCTCTACCACAAAA | 61 | 141 bp |
| Reverse | CGTTGGCATTTCTTCTCCA | 61 | ||
| CDX2 | Forward | CAGCGGCGGAACCTGTG | 63 | 92 bp |
| Reverse | ACTCGGTATTTGTCTTTCGTCCTG | 63 | ||
| OCT4A | Forward | GCTGGAGCCGAACCCCGAGG | 68 | 150 bp |
| Reverse | CACCTTCCCAAAGAGAACCCCCAAA | 68 | ||
| SOX2 | Forward | CGGCGGTGGCAACTCTAC | 64 | 100 bp |
| Reverse | TCGGGACCACACCATGAAAG | 64 | ||
| NANOG | Forward | CATCTGCTGAGACCCTCGAC | 60 | 195 bp |
| Reverse | GGGTCTGCGAGAACACAGTT | 60 | ||
| HNF4a | Forward | GCTTCTTTCGGAGGAGTGTG | 58 | 183 bp |
| Reverse | TTGACCTGCGAGTGCTGAT | 58 | ||
| PDGFA | Forward | CAGCGTGGTCTTTGTTCTCC | 60 | 189 bp |
| Reverse | TGTTTCGTGGTTCTGTTTGC | 60 | ||
| PDGFRA | Forward | GGTCACCTGTGCCGTCTTTA | 59 | 115 bp |
| Reverse | TTTGATGGACGGGACCTTGG | 59 | ||
| GAPDH | Forward | TGCTCCTCCCCGTTCGAC | 60 | 100 bp |
| Reverse | ATGCGGCCAAATCCGTTC | 60 | ||
| ACTB | Forward | GTGGACATCAGGAAGGACCTCTA | 63 | 131 bp |
| Reverse | ATGATCTTGATCTTCATGGTGCT | 63 | ||
| Primary Antibodies | Target | Host | Company | Catalog Number | Concentration of Antibody |
| SOX2 | Rabbit | Millipore | AB5603 | 5 μg/mL | |
| DAB2 | Goat | R&D systems | AF8064 | 2 μg/mL | |
| NANOG | Rabbit | Peprotech | 500-P236 | 1 μg/mL | |
| OCT4 | Rabbit | SantaCruz | sc-9081 | 1 μg/mL | |
| SOX17 | Goat | R&D systems | AF1924 | 1 μg/mL | |
| Secondary Antibodies | Fluorescent dye | Target/Host | Company | Catalog Number | Concentration of Antibody |
| Alexa594 | Rabbit/Donkey | Invitrogen | A-21207 | 1:5000 | |
| Alexa488 | Goat/Donkey | Invitrogen | A-11055 | 1:5000 |
References
- Saini, D.; Yamanaka, Y. Cell Polarity-Dependent Regulation of Cell Allocation and the First Lineage Specification in the Preimplantation Mouse Embryo. Curr. Top. Dev. Biol. 2018, 128, 11–35. [Google Scholar] [PubMed]
- Rossant, J. Genetic Control of Early Cell Lineages in the Mammalian Embryo. Ann. Rev. Genet. 2018, 52, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Alberio, R. Transcriptional and epigenetic control of cell fate decisions in early embryos. Reprod. Fertil. Dev. 2017, 30, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.L.; Pera, M.F. Capturing Totipotent Stem Cells. Cell Stem Cell 2018, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Houston, D.W. Vertebrate Axial Patterning: From Egg to Asymmetry. Adv. Exp. Med. Biol. 2017, 953, 209–306. [Google Scholar]
- Leung, C.Y.; Zernicka-Goetz, M. Mapping the journey from totipotency to lineage specification in the mouse embryo. Curr. Opin. Genet. Dev. 2015, 34, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Shi, J.; Tao, Y.; Zernicka-Goetz, M. Tracing the origin of heterogeneity and symmetry breaking in the early mammalian embryo. Nat. Commun. 2018, 9, 1819. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.H. From mouse egg to mouse embryo: Polarities, axes, and tissues. Annu. Rev. Cell Dev. Biol. 2009, 25, 483–512. [Google Scholar] [CrossRef]
- Guo, G.; Huss, M.; Tong, G.Q.; Wang, C.; Li Sun, L.; Clarke, N.D.; Robson, P. Resolution of cell fate decisions revealed by single-cell gene expression analysis from zygote to blastocyst. Dev. Cell 2010, 18, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Lou, X.; Kang, M.; Xenopoulos, P.; Munoz-Descalzo, S.; Hadjantonakis, A.K. A rapid and efficient 2D/3D nuclear segmentation method for analysis of early mouse embryo and stem cell image data. Stem Cell Rep. 2014, 2, 382–397. [Google Scholar] [CrossRef] [Green Version]
- Biase, F.H.; Cao, X.; Zhong, S. Cell fate inclination within 2-cell and 4-cell mouse embryos revealed by single-cell RNA sequencing. Genome Res. 2014, 24, 1787–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Chen, Q.; Li, X.; Zheng, X.; Zhang, Y.; Qiao, J.; Tang, F.; Tao, Y.; Zhou, Q.; Duan, E. Dynamic transcriptional symmetry-breaking in pre-implantation mammalian embryo development revealed by single-cell RNA-seq. Development 2015, 142, 3468–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petropoulos, S.; Edsgard, D.; Reinius, B.; Deng, Q.; Panula, S.P.; Codeluppi, S.; Plaza Reyes, A.; Linnarsson, S.; Sandberg, R.; Lanner, F. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell 2016, 165, 1012–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakeley, P.; Fogarty, N.M.; Del Valle, I.; Wamaitha, S.E.; Hu, T.X.; Elder, K.; Snell, P.; Christie, L.; Robson, P.; Niakan, K.K. Defining the three cell lineages of the human blastocyst by single-cell RNA-seq. Development 2015, 142, 3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiang, Y.; Yin, Q.; Du, Z.; Peng, X.; Wang, Q.; Fidalgo, M.; Xia, W.; Li, Y.; Zhao, Z.A.; et al. Dynamic epigenomic landscapes during early lineage specification in mouse embryos. Nat. Genet. 2018, 50, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Guo, H.; Ren, Y.; Hou, Y.; Dong, J.; Li, R.; Lian, Y.; Fan, X.; Hu, B.; Gao, Y.; et al. Single-cell DNA methylome sequencing of human preimplantation embryos. Nat. Genet. 2018, 50, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-H.; Lee, C.-K. Pig Pluripotent Stem Cells as a Candidate for Biomedical Application. J. Anim. Reprod. Biotechnol. 2019, 34, 8. [Google Scholar] [CrossRef] [Green Version]
- Han, N.R.; Baek, S.; Lee, Y.; Lee, J.; Yun, J.I.; Lee, E.; Lee, S.T. Effects of in vitro Culture Period of Reconstructed Embryos and Genetic Background of Feeder Cells on Establishment of Embryonic Stem Cells Derived from Somatic Cell Nuclear Transfer Blastocysts in Pigs. J. Anim. Reprod. Biotechnol. 2020, 35, 8. [Google Scholar] [CrossRef] [Green Version]
- Lavagi, I.; Krebs, S.; Simmet, K.; Beck, A.; Zakhartchenko, V.; Wolf, E.; Blum, H. Single-cell RNA sequencing reveals developmental heterogeneity of blastomeres during major genome activation in bovine embryos. Sci. Rep. 2018, 8, 4071. [Google Scholar] [CrossRef]
- Wei, Q.; Li, R.; Zhong, L.; Mu, H.; Zhang, S.; Yue, L.; Xiang, J.; Li, E.; Zhi, M.; Cao, S.; et al. Lineage specification revealed by single-cell gene expression analysis in porcine preimplantation embryos. Biol. Reprod. 2018, 99, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Ibeas, P.; Sang, F.; Zhu, Q.; Tang WW, C.; Withey, S.; Klisch, D.; Wood, L.; Loose, M.; Surani, M.A.; Alberio, R. Pluripotency and X chromosome dynamics revealed in pig pre-gastrulating embryos by single cell analysis. Nat. Commun. 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Q.; Yang, X.; Zhang, H.; Liu, S.; Zhao, J.; Zhang, J.; Weng, X.; Jin, J.; Liu, Z. Lineage specification and pluripotency revealed by transcriptome analysis from oocyte to blastocyst in pig. Faseb J. 2020, 34, 691–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, S.J.; Robertson, E.J. Making a commitment: Cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 2009, 10, 91–103. [Google Scholar] [CrossRef]
- Lanner, F. Lineage specification in the early mouse embryo. Exp. Cell Res. 2014, 321, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ralston, A.; Rossant, J. CDX2 acts downstream of cell polarization to cell-autonomously promote trophectoderm fate in the early mouse embryo. Dev. Biol. 2008, 313, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strumpf, D.; Mao, C.A.; Yamanaka, Y.; Ralston, A.; Chawengsaksophak, K.; Beck, F.; Rossant, J. CDX2 is required for correct cell fate specification and differentiation of trophectoderm in the mouse blastocyst. Development 2005, 132, 2093–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou, G.; Liu, S.; Sun, M.; Zhu, J.; Xue, B.; Guo, J.; Zhao, Y.; Qu, B.; Weng, X.; Wei, Y.; et al. CDX2 is essential for cell proliferation and polarity in porcine blastocysts. Development 2017, 144, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Berg, D.K.; Smith, C.S.; Pearton, D.J.; Wells, D.N.; Broadhurst, R.; Donnison, M.; Pfeffer, P.L. Trophectoderm lineage determination in cattle. Dev. Cell 2011, 20, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Chazaud, C.; Yamanaka, Y. Lineage specification in the mouse preimplantation embryo. Development 2016, 143, 1063–1074. [Google Scholar] [CrossRef] [Green Version]
- Kuijk, E.W.; Du Puy, L.; Van Tol, H.T.; Oei, C.H.; Haagsman, H.P.; Colenbrander, B.; Roelen, B.A. Differences in early lineage segregation between mammals. Dev. Dyn. 2008, 237, 918–927. [Google Scholar] [CrossRef]
- Han, N.R.; Baek, S.; Lee, Y.; Lee, J.; Yun, J.I.; Lee, E.; Lee, S.T. Establishment of In-Vitro Culture System for Enhancing Production of Somatic Cell Nuclear Transfer (SCNT) Blastocysts with High Performance in the Colony Formation and Formation of Colonies Derived from SCNT Blastocysts in Pigs. J. Anim. Reprod. Biotechnol. 2019, 34, 9. [Google Scholar] [CrossRef] [Green Version]
- Papaioannou, V.E.; Ebert, K.M. The preimplantation pig embryo: Cell number and allocation to trophectoderm and inner cell mass of the blastocyst in vivo and in vitro. Development 1988, 102, 793–803. [Google Scholar] [PubMed]
- Pomar, F.J.; Teerds, K.J.; Kidson, A.; Colenbrander, B.; Tharasanit, T.; Aguilar, B.; Roelen, B.A. Differences in the incidence of apoptosis between in vivo and in vitro produced blastocysts of farm animal species: A comparative study. Theriogenology 2005, 63, 2254–2268. [Google Scholar] [CrossRef]
- Kashyap, V.; Rezende, N.C.; Scotland, K.B.; Shaffer, S.M.; Persson, J.L.; Gudas, L.J.; Mongan, N.P. Regulation of stem cell pluripotency and differentiation involves a mutual regulatory circuit of the NANOG, OCT4, and SOX2 pluripotency transcription factors with polycomb repressive complexes and stem cell microRNAs. Stem Cells Dev. 2009, 18, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Artus, J.; Kang, M.; Cohen-Tannoudji, M.; Hadjantonakis, A.K. PDGF signaling is required for primitive endoderm cell survival in the inner cell mass of the mouse blastocyst. Stem Cells 2013, 31, 1932–1941. [Google Scholar] [CrossRef] [Green Version]
- Artus, J.; Panthier, J.J.; Hadjantonakis, A.K. A role for PDGF signaling in expansion of the extra-embryonic endoderm lineage of the mouse blastocyst. Development 2010, 137, 3361–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.A.; Plath, K.; Zeitlinger, J.; Brambrink, T.; Medeiros, L.A.; Lee, T.I.; Levine, S.S.; Wernig, M.; Tajonar, A.; Ray, M.K.; et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 2006, 441, 349–353. [Google Scholar] [CrossRef]
- Liu, Z.; Scannell, D.R.; Eisen, M.B.; Tjian, R. Control of embryonic stem cell lineage commitment by core promoter factor, TAF3. Cell 2011, 146, 720–731. [Google Scholar] [CrossRef] [Green Version]
- Hofsteen, P.; Robitaille, A.M.; Chapman, D.P.; Moon, R.T.; Murry, C.E. Quantitative proteomics identify DAB2 as a cardiac developmental regulator that inhibits WNT/beta-catenin signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.; Cai, K.Q.; Tao, W.; Smith, E.R.; Xu, X.X. Differential requirement for Dab2 in the development of embryonic and extra-embryonic tissues. BMC Dev. Biol. 2013, 13, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, L.; Feng, G.; Wang, Y.; Li, Y.; Li, X.; Liu, C.; Jiao, G.; Huang, C.; Shi, J.; et al. Asymmetric Expression of LincGET Biases Cell Fate in Two-Cell Mouse Embryos. Cell 2018, 175, 1887–1901.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.H.; Smith, E.R.; Roland, I.H.; Sheng, Z.; He, J.; Martin, W.D.; Hamilton, T.C.; Lambeth, J.D.; Xu, X.X. Disabled-2 is essential for endodermal cell positioning and structure formation during mouse embryogenesis. Dev. Biol. 2002, 251, 27–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, G.M.; Dalvit, G.C.; Achi, M.V.; Miguez, M.S.; Cetica, P.D. Immature oocyte quality and maturational competence of porcine cumulus-oocyte complexes subpopulations. Biocell 2009, 33, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, K.; Hoshino, Y.; Tanemura, K.; Sato, E. Selection of in vitro-matured porcine oocytes based on localization patterns of lipid droplets to evaluate developmental competence. J. Reprod. Dev. 2013, 59, 405–408. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, K.; Suzuki, C.; Tanaka, A.; Anas, I.M.; Iwamura, S. Birth of piglets derived from porcine zygotes cultured in a chemically defined medium. Biol. Reprod. 2002, 66, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.N.; Choi, K.H.; Lee, C.K. Multi-resistance strategy for viral diseases and in vitro short hairpin RNA verification method in pigs. Asian-Australas J. Anim. Sci. 2018, 31, 489–498. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.-N.; Lee, M.; Choe, G.C.; Lee, D.-K.; Choi, K.-H.; Kim, S.-H.; Jeong, J.; Lee, C.-K. Identification of the Lineage Markers and Inhibition of DAB2 in In Vitro Fertilized Porcine Embryos. Int. J. Mol. Sci. 2020, 21, 7275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197275
Oh J-N, Lee M, Choe GC, Lee D-K, Choi K-H, Kim S-H, Jeong J, Lee C-K. Identification of the Lineage Markers and Inhibition of DAB2 in In Vitro Fertilized Porcine Embryos. International Journal of Molecular Sciences. 2020; 21(19):7275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197275
Chicago/Turabian StyleOh, Jong-Nam, Mingyun Lee, Gyung Cheol Choe, Dong-Kyung Lee, Kwang-Hwan Choi, Seung-Hun Kim, Jinsol Jeong, and Chang-Kyu Lee. 2020. "Identification of the Lineage Markers and Inhibition of DAB2 in In Vitro Fertilized Porcine Embryos" International Journal of Molecular Sciences 21, no. 19: 7275. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197275